Effect of Acute High-Intensity Interval Training on Immune Function and Oxidative Stress in Canoe/Kayak Athletes


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
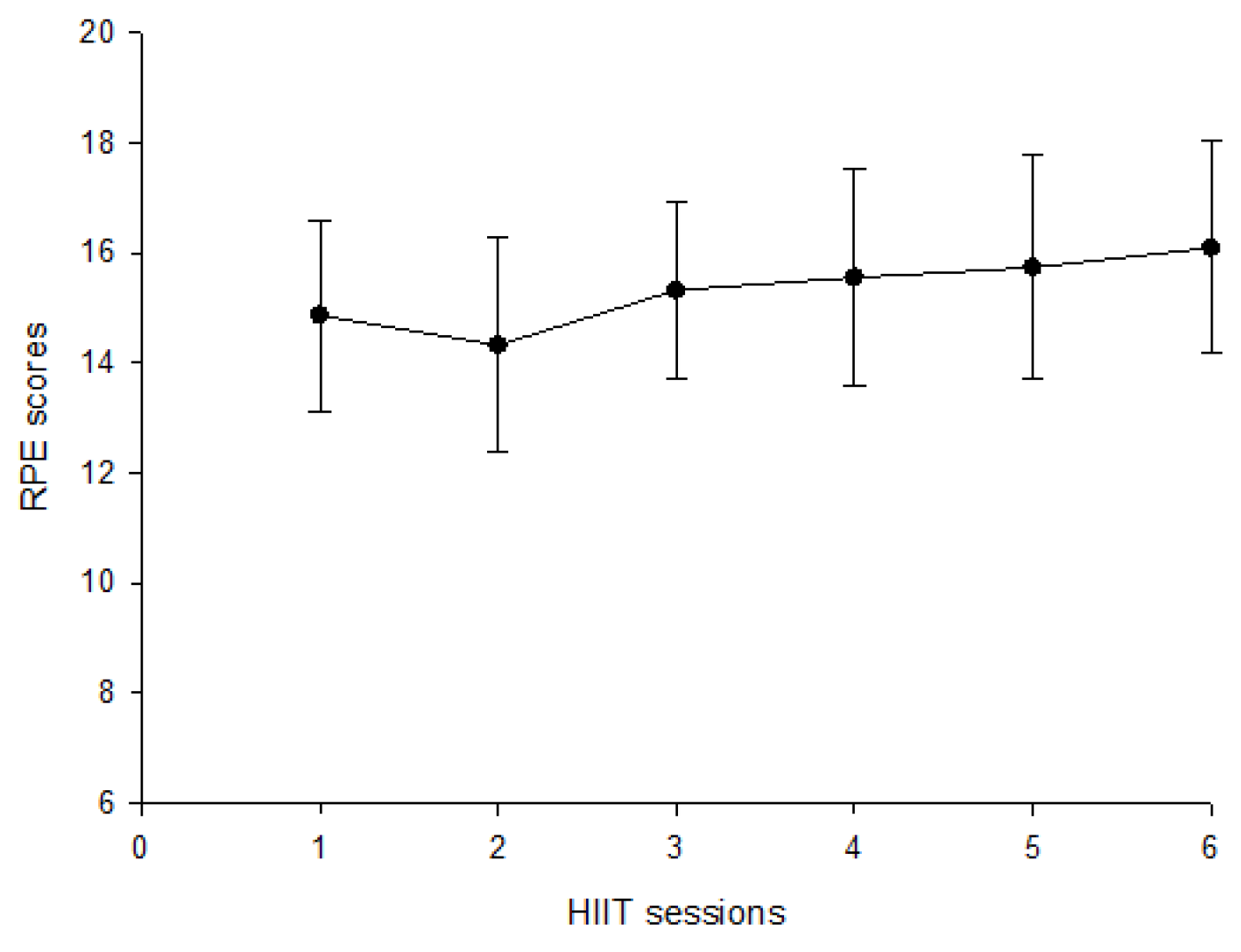
2.2. Experimental Protocol
2.3. Blood Collection and Treatment
2.4. Determination of Neutrophil Degranulation
2.5. Determination of Lymphocyte Subsets
2.6. Index of Lipid Peroxidation
2.7. Determination of Antioxidant Enzymes
2.8. Saliva Collection and Treatment
2.9. Determination of Salivary Immunoglobin A
2.10. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- International Canoe Federation. ICF Canoe Sprint Rules; International Canoe Federation: Lausanne, Switzerland, 2022; Available online: https://www.canoeicf.com/sites/default/files/rules_canoe_sprint_2022.pdf (accessed on 1 January 2022).
- Byrnes, W.C.; Kearney, J.T. Aerobic and anaerobic contributions during simulated canoe/kayak sprint events. Med. Sci. Sports Exerc. 1997, 29, S220. [Google Scholar] [CrossRef]
- Van Someren, K.A.; Howatson, G. Prediction of flatwater kayaking performance. Int. J. Sports Physiol. Perform. 2008, 3, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Van Someren, K.A.; Palmer, G.S. Prediction of 200-m sprint kayaking performance. Can. J. Appl. Physiol. 2003, 28, 505–517. [Google Scholar] [CrossRef]
- Tesch, P.A. Physiological characteristics of elite kayak paddlers. Can. J. Appl. Sport Sci. 1983, 8, 87–91. [Google Scholar] [PubMed]
- Fry, R.W.; Morton, A.R. Physiological and Kinanthroprnetric attributes of elite flatwater kayakists. Med. Sci. Sports Exerc. 1991, 23, 1297–1301. [Google Scholar] [CrossRef] [PubMed]
- Gray, G.L.; Matherson, G.O.; MeKenzie, D.C. The metabolic cost of two kayaking techniques. Int. J. Sports Med. 1995, 16, 250–254. [Google Scholar] [CrossRef]
- Bishop, D. Physiological predictors of flat-water kayak performance in women. Eur. J. Appl. Physiol. 2000, 82, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Zouhal, H.; Lahaye, L.D.; Abderrahaman, B.; Minter, G.; Herbez, R.; Castagna, C. Energy system contribution to Olympic distances in flat water kayaking (500 and 1,000 m) in highly trained subjects. J. Strength Cond. Res. 2000, 26, 825–831. [Google Scholar] [CrossRef]
- Fernandez, B.; Perez-Landaluce, J.; Rodriguez, M.; Terrados, N. Metabolic contribution in Olympic kayaking events. Med. Sci. Sports Exerc. 1995, 27, 24. [Google Scholar] [CrossRef]
- Viana, R.B.; de Lira, C.A.B.; Naves, J.P.A.; Coswig, V.S.; Del Vecchio, F.B.; Ramirez-Campillo, R.; Vieira, C.A.; Gentil, P. Can we draw general conclusions from interval training studies? Sports Med. 2018, 48, 2001–2009. [Google Scholar] [CrossRef]
- Billat, V.L.; Slawinski, J.; Bocquet, V.; Demarle, A.; Lafitte, L.; Chassaing, P.; Koralsztein, J.P. Intermittent runs at the velocity associated with maxmal oxygen uptake enables subjects to reman at maximal oxygen uptake for a longer time than intense but submaximal runs. Eur. J. Appl. Physiol. 2000, 81, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Billat, L.V. Interval training for performance: A scientific and empirical practice. Sports Med. 2001, 31, 13–31. [Google Scholar] [CrossRef] [PubMed]
- Nieman, W.L.M. The compelling link between physical activity and the body’s defense system. J. Sport Health Sci. 2018, 8, 201–217. [Google Scholar] [CrossRef]
- Gonçalves, C.A.M.; Dantas, P.M.S.; Dos Santos, I.K.; Dantas, M.; da Silva, D.C.P.; Cabral, B.G.D.; Guerra, R.O.; Cavalcanti, G.B. Effect of acute and chronic aerobic exercise on immunological markers: A systematic review. Front. Physol. 2020, 10, 1602. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Rohde, T.; Ostrowski. Recovery of the immune system after exercise. Acta Physiol. Scand. 1998, 162, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C. The effect of exercise on immune function. Bull. Rheum. Dis. 1994, 43, 5–8. [Google Scholar] [PubMed]
- Walsh, G.; Shephard, J. Part one: Immune function and exercise. Exerc. Immunol. Rev. 2011, 17, 6–63. [Google Scholar]
- Davison, G. Innate immune responses to a single session of sprint interval training. Appl. Physiol. Nutr. Metab. 2011, 36, 395–404. [Google Scholar] [CrossRef]
- Ottone, V.D.O.; Costa, K.B.; Tossige-Gomes, R.; De Matos, M.A.; Brito-Melo, G.; Magalhaes, F.D.C.; Esteves, E.A.; Amorim, F.; Rocha-Vieira, E. Late neutrophil priming following a single session of high-intensity interval exercise. Int. J. Sports Med. 2019, 40, 171–179. [Google Scholar] [CrossRef]
- Tossige-Gomes, R.; Costa, K.B.; Ottone, V.D.O.; Magalhães, F.D.C.; Amorim, F.T.; Rocha-Vieira, E. Lymphocyte redox imbalance and reduced proliferation after a single session of high intensity interval exercise. PLoS ONE 2016, 11, e0153647. [Google Scholar] [CrossRef] [PubMed]
- Born, D.P.; Zinner, C.; Sperlich, B. The mucosal immune function is not compromised during a period of high-intensity interval training. Is it time to reconsider an old assumption? Front. Physiol. 2017, 8, 485. [Google Scholar] [CrossRef] [PubMed]
- Khammassi, M.; Ouerghi, N.; Said, M.; Feki, M.; Khammassi, Y.; Pereira, B.; Thivel, D.; Bouassida, A. Continuous moderate-intensity but not high-intensity interval training improves immune function biomarkers in healthy young men. J. Strength Cond. Res. 2020, 34, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Shek, P.N.; Sabiston, B.H.; Buguet, A.; Radomski, M.W. Strenuous exercise and immunological changes: A multiple-time-point analysis of leukocyte subsets, CD4/CD8 ratio, immunoglobulin production and NK cell response. Int. J. Sports Med. 1995, 16, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M.; Bishop, N.; Walish, N. Exercise Immunology; Routledge: London, UK, 2013. [Google Scholar]
- Novas, A.M.; Rowbottom, D.G.; Jenkins, D.G. Tennis, incidence of URTI and salivary IgA. Int. J. Sports Med. 2003, 24, 223–239. [Google Scholar] [CrossRef] [PubMed]
- Davison, G.; Diment, B.C. Bovine colostrum supplementation attenuates the decrease of salivary lysozyme and enhances the recovery of neutrophil function after prolonged exercise. Br. J. Nutr. 2010, 103, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.L. Exercise-induced modulation of antioxidant defense. Ann. N. Y. Acad. Sci. 2002, 959, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C. Exercise immunology: Nutritional countermeasures. Can. J. Appl. Physiol. 2001, 26, S45–S55. [Google Scholar] [CrossRef]
- Ji, L.L. Oxidative stress during exercise: Implication of antioxidant nutrients. Free Radic. Biol. Med. 1995, 18, 1079–1086. [Google Scholar] [CrossRef]
- Radak, Z.; Chung, H.Y.; Koltai, E.; Taylor, A.W.; Goto, S. Exercise, oxidative stress and hormesis. Ageing Res. Rev. 2008, 7, 34–42. [Google Scholar] [CrossRef]
- Johnson, B.D.; Padilla, J.; Wallace, J.P. The exercise dose affects oxidative stress and brachial artery flow-mediated dilation in trained men. Eur. J. Appl. Physiol. 2012, 112, 33–42. [Google Scholar] [CrossRef]
- Li, T.L. Exercise and salivary IgA response. J. Exerc. Physiol. Fitness 2007, 6, 35–50. [Google Scholar] [CrossRef]
- Dill, D.B.; Costill, D.L. Calculation of percentage changes in volumes of blood, plasma, and red cells in dehydration. J. Appl. Physiol. 1974, 37, 247–248. [Google Scholar] [CrossRef] [PubMed]
- Chicharro, J.L.; Lucia, A.; Perez, M.; Vaquero, A.F.; Urena, R. Saliva composition and exercise. Sports Med. 1998, 26, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Li, T.L.; Gleeson, M. The effect of single and repeated bouts of prolonged cycling on leukocyte redistribution, neutrophil degranulation, IL-6, and plasma stress hormone responses. Int. J. Sport Nutr. Exerc. Metabol. 2004, 14, 501–516. [Google Scholar] [CrossRef] [PubMed]
- Adams, G.R.; Zaldivar, F.P.; Nance, D.M.; Kodesh, E.; Radom-Aizik, S.; Cooper, D.M. Exercise and leukocyte interchange among central circulation, lung, spleen, and muscle. Brain Behav. Immun. 2011, 25, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Kappel, M.; Tvede, N.; Galbo, H.; Haahr, P.M.; Kjaer, M.; Linstow, M.; Klarlund, K.; Pedersen, B.K. Evidence that the effect of physical exercise on NK cell activity is mediated by epinephrine. J. Appl. Physiol. 1991, 70, 2530–2534. [Google Scholar] [CrossRef]
- Tvede, N.; Kappel, M.; Klarlund, K.; Duhn, S.; Halkjaer-Kristensen, J.; Kjaer, M.; Galbo, H.; Pedersen, B.K. Evidence that the effect of bicycle exercise on blood mononuclear cell proliferative responses and subsets is mediated by epinephrine. Int. J. Sports Med. 1994, 15, 100–104. [Google Scholar] [CrossRef]
- Fisher, G.; Schwartz, D.D.; Quindry, J.; Barberio, M.D.; Foster, E.B.; Jones, K.W.; Pascoe, D.D. Lymphocyte enzymatic antioxidant responses to oxidative stress following high-intensity interval exercise. J. Appl. Physiol. 2011, 110, 730–737. [Google Scholar] [CrossRef]
- Walsh, N.P.; Blannin, A.K.; Clark, A.M.; Cook, L.; Robson, P.; Gleeson, M. The effects of high-intensity intermittent exercise on the plasma concentrations of glutamine and organic acids. Eur. J. Appl. Physiol. Occup. Physiol. 1998, 77, 434–438. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Akerstrom, T.C.; Nielsen, A.R.; Fischer, C.P. Role of myokines in exercise and metabolism. J. Appl. Physiol. 2007, 103, 1093–1098. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Hoffman-Goetz, L. Exercise and the immune system: Regulation, integration, and adaptation. Physiol. Rev. 2000, 80, 1055–1081. [Google Scholar] [CrossRef] [PubMed]
- Kargotich, S.; Keast, D.; Goodman, C.; Crawford, G.P.; Morton, A.R. The influence of blood volume changes on leucocyte and lymphocyte subpopulations in elite swimmers following interval training of varying intensities. Int. J. Sports Med. 1997, 18, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Wahl, P.; Mathes, S.; Bloch, W.; Zimmer, P. Acute impact of recovery on the restoration of cellular immunological homeostasis. Laryngo-Rhino-Otol. 2019, 41, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Belviranli, M.; Okudan, N.; Kabak, B. The effects of acute high-intensity interval training on hematological parameters in sedentary subjects. Med. Sci. 2017, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Gray, A.B.; Telford, R.G.; Collins, M.; Baker, M.S.; Weidemann, M.J. Granulocyte activation induced by intense interval running. J. Leukocyte Biol. 1993, 53, 591–597. [Google Scholar] [CrossRef]
- Souza, D.; Vale, A.F.; Silva, A.; Araújo, M.A.S.; de Paula Júnior, C.A.; de Lira, C.A.B.; Ramirez-Campillo, R.; Martins, W.; Gentil, P. Acute and chronic effects of interval training on the immune system: A systematic review with meta-analysis. Biology 2001, 10, 868. [Google Scholar] [CrossRef]
- Rowbottom, D.G.; Green, K.J. Acute exercise effects on the immune system. Med. Sci. Sports Exerc. 2000, 32, 396–405. [Google Scholar] [CrossRef]
- Robson, P.J.; Blannin, A.K.; Walsh, N.P.; Castell, L.M.; Gleeson, M. Effects of exercise intensity, duration and recovery on in vitro neutrophil function in male athletes. Int. J. Sports Med. 1999, 20, 128–135. [Google Scholar] [CrossRef]
- Gabriel, H.; Schwarz, L.; Borm, P.; Kindermann, W. Differential mobilization of leucocyte and lymphocyte subpopulations into the circulation during endurance exercise. Eur. J. Appl. Physiol. Occup. Physiol. 1992, 65, 529–534. [Google Scholar] [CrossRef]
- Gabriel, H.; Kindermann, W. The acute immune response to exercise: What does it mean? Int. J. Sports Med. 1997, 18, 28–45. [Google Scholar] [CrossRef]
- Hinton, J.R.; Rowbottom, D.G.; Keast, D.; Morton, A.R. Acute intensive interval training and in vitro t-lymphocyte function. Int. J. Sports Med. 1997, 18, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Kakanis, M.W.; Peake, J.; Brenu, E.W.; Simmonds, M.; Gray, B.; Hooper, S.L.; Marshall-Gradisnik, S.M. The open window of susceptibility to infection after acute exercise in healthy young male elite athletes. Exerc. Immunol. Rev. 2010, 16, 119–137. [Google Scholar] [CrossRef] [PubMed]
- Frisina, J.P.; Gaudieri, S.; Cable, T.; Keast, D.; Palmer, T.N. Effects of acute exercise on lymphocyte subsets and metabolic activity. Int. J. Sports Med. 1994, 15, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Fry, R.W.; Morton, A.R.; Crawford, G.P.; Keast, D. Cell numbers and in vitro responses of leucocytes and lymphocyte subpopulations following maximal exercise and interval training sessions of different intensities. Eur. J. Appl. Physiol. Occup. Physiol. 1992, 64, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Mackinnon, L.T.; Chick, T.W.; van As, A.; Tomasi, T.B. The effect of exercise on secretory and natural immunity. Adv. Experi. Med. Biol. 1987, 216A, 869–876. [Google Scholar] [CrossRef]
- Mackinnon, L.T.; Ginn, E.; Seymour, G.J. Decreased salivary immunoglobulin A secretion rate after intense interval exercise in elite kayakers. Eur. J. Appl. Physiol. Occup. Physiol. 1993, 67, 180–184. [Google Scholar] [CrossRef]
- Nieman, D.C.; Miller, A.R.; Henson, D.A.; Warren, B.J.; Gusewitch, G.; Johnson, R.L.; Davis, J.M.; Butterworth, D.E.; Nehlsen-Cannarella, S.L. Effects of high- vs. moderate-intensity exercise on natural killer cell activity. Med. Sci. Sports Exerc. 1993, 25, 1126–1134. [Google Scholar] [CrossRef]
- Krüger, K.; Alack, K.; Ringseis, R.; Mink, L.; Pfeifer, E.; Schinle, M.; Gindler, K.; Kimmelmann, L.; Walscheid, R.; Muders, K.; et al. Apoptosis of T-cell subsets after acute high-intensity interval exercise. Med. Sci. Sports Exerc. 2016, 48, 2021–2029. [Google Scholar] [CrossRef]
- Gleeson, M.; Bishop, N.; Oliveira, M.; McCauley, T.; Tauler, P. Sex differences in immune variables and respiratory infection incidence in an athletic population. Exerc. Immunol. Rev. 2011, 17, 122–135. [Google Scholar]
- Mendonca, G.V.; Heffernan, K.S.; Rossow, L.; Guerra, M.; Pereira, F.M.D.C.D.; Fernhall, B. Sex differences in linear and nonlinear heart rate variability during early recovery from supramaximal exercise. Appl. Physiol. Nutr. Metab. 2010, 35, 439–446. [Google Scholar] [CrossRef]
- Fahlman, M.M.; Engels, H.J.; Morgan, A.L.; Kolokouri, I. Mucosal IgA response to repeated Wingate tests in females. Int. J. Sports Med. 2001, 22, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Mackinnon, L.T.; Jenkins, D.G. Decreased salivary immunoglobulins after intense interval exercise before and after training. Med. Sci. Sports Exerc. 1993, 25, 678–683. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.J.; Edwards, R.H.; Symons, M.C. Electron spin resonance studies of intact mammalian skeletal muscle. Biochim. Biophys. Acta 1985, 847, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Gohil, K.; Rothfuss, L.; Lang, J.; Packer, L. Effect of exercise training on tissue vitamin E and ubiquinone content. J. Appl. Physiol. 1987, 63, 1638–1641. [Google Scholar] [CrossRef] [PubMed]
- Lew, H.; Pyke, S.; Quintanilha, A. Changes in the glutathione status of plasma, liver and muscle following exhaustive exercise in rats. FEBS Lett. 1985, 185, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Anzueto, A.; Andrade, F.H.; Maxwell, L.C.; Levine, S.M.; Lawrence, R.A.; Jenkinson, S.G. Diaphragmatic function after resistive breathing in vitamin E-deficient rats. J. Appl. Physiol. 1993, 74, 267–271. [Google Scholar] [CrossRef]
- Chang, C.K.; Tseng, H.F.; Hsuuw, Y.D.; Chan, W.H.; Shieh, L.C. Higher LDL oxidation at rest and after a rugby game in weekend warriors. Ann. Nutr. Metabol. 2002, 46, 103–107. [Google Scholar] [CrossRef]
- Knez, W.L.; Jenkins, D.G.; Coombes, J.S. Oxidative stress in half and full Ironman triathletes. Med. Sci. Sports Exerc. 2007, 39, 283–288. [Google Scholar] [CrossRef]
- Vider, J.; Lehtmaa, J.; Kullisaar, T.; Vihalemm, T.; Zilmer, K.; Kairane, C.; Landõr, A.; Karu, T.; Zilmer, M. Acute immune response in respect to exercise-induced oxidative stress. Pathophysiology 2001, 7, 263–270. [Google Scholar] [CrossRef]
- Elosua, R.; Molina, L.; Fito, M.; Arquer, A.; Sanchez-Quesada, J.L.; Covas, M.I.; Ordoñez-Llanos, J.; Marrugat, J. Response of oxidative stress biomarkers to a 16-week aerobic physical activity program, and to acute physical activity, in healthy young men and women. Atherosclerosis 2003, 167, 327–334. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variables | Pre-T | Post-T | Post-3 h |
|---|---|---|---|
| Total leucocytes (109·L−1) | 5.09 ± 1.15 | 9.04 ± 2.12 * | 8.47 ± 1.94 * |
| Lymphocyte percentage (%) | 35.46 ± 8.11 | 45.73 ± 8.66 * | 21.05 ± 7.18 # |
| Lymphocyte counts (109·L−1) | 1.77 ± 0.44 | 4.14 ± 1.27 * | 1.71 ± 0.43 # |
| Neutrophil percentage (%) | 53.53 ± 8.87 | 44.11 ± 9.21 * | 72.41 ± 8.16 *# |
| Neutrophil counts (109·L−1) | 2.79 ± 0.96 | 4.02 ± 1.28 * | 6.24 ± 2.04 *# |
| Total LPS-stimulated elastase release (ug·L−1) | 232.72 ± 91.80 | 297.90 ± 108.86 * | 486.30 ± 205.09 *# |
| LPS-stimulated elastase release per neutrophil (fg·cell−1) | 84.94 ± 26.36 | 74.72 ± 14.18 | 79.27 ± 26.38 |
| Variables | Pre-T | Post-T | Post-3 h |
|---|---|---|---|
| CD4+ percentage (%) | 33.95 ± 10.37 | 23.23 ± 8.95 * | 35.24 ± 11.58 # |
| CD4+ counts (109·L−1) | 0.60 ± 0.20 | 0.95 ± 0.45 * | 0.59 ± 0.22 # |
| CD8+ percentage (%) | 25.03 ± 10.92 | 22.85 ± 9.81 | 25.07 ± 10.21 |
| CD8+ counts (109·L−1) | 0.41 ± 0.23 | 0.89 ± 0.41 * | 0.38 ± 0.23 # |
| CD19+ percentage (%) | 14.16 ± 6.23 | 9.32 ± 4.26 * | 15.56 ± 6.43 # |
| CD19+ counts (109·L−1) | 0.26 ± 0.14 | 0.40 ± 0.18 * | 0.27 ± 0.12 # |
| CD56+ percentage (%) | 11.24 ± 5.91 | 25.97 ± 14.22 * | 9.40 ± 6.78 # |
| CD56+ counts (109·L−1) | 0.19 ± 0.12 | 1.17 ± 0.91 * | 0.16 ± 0.12 # |
| CD4/CD8 ratio | 1.63 ± 0.75 | 2.93 ± 1.85 * | 1.70 ± 1.00 # |
| Variables | Pre-T | Post-T | Post-3 h |
|---|---|---|---|
| Saliva flow rate (mg·min−1) | 263.89 ± 103.23 | 282.46 ± 114.79 | 241.12 ± 119.32 |
| SIgA concentration (mg·L−1) | 0.39 ± 0.40 | 0.26 ± 0.17 | 0.37 ± 0.25 |
| SIgA secretion rate (mg·min−1) | 0.07 ± 0.04 | 0.08 ± 0.07 | 0.08 ± 0.07 |
| Variables | Pre-T | Post-T | Post-3 h |
|---|---|---|---|
| SOD (U·mL−1) | 3.61 ± 2.48 | 4.03 ± 3.22 | 4.84 ± 3.42 *# |
| CAT (U·mL−1) | 506.16 ± 191.11 | 693.09 ± 202.62 * | 337.54 ± 121.82 *# |
| TBARS (U·mL−1) | 2.51 ± 0.72 | 3.08 ± 1.17 * | 2.40 ± 0.55 # |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, T.-T.; Li, T.-L.; Ko, B.-J.; Chien, L.-H. Effect of Acute High-Intensity Interval Training on Immune Function and Oxidative Stress in Canoe/Kayak Athletes. Biology 2023, 12, 1144. https://doi.org/10.3390/biology12081144
Lee T-T, Li T-L, Ko B-J, Chien L-H. Effect of Acute High-Intensity Interval Training on Immune Function and Oxidative Stress in Canoe/Kayak Athletes. Biology. 2023; 12(8):1144. https://doi.org/10.3390/biology12081144
Chicago/Turabian StyleLee, Ting-Ting, Tzai-Li Li, Bo-Jen Ko, and Li-Hui Chien. 2023. "Effect of Acute High-Intensity Interval Training on Immune Function and Oxidative Stress in Canoe/Kayak Athletes" Biology 12, no. 8: 1144. https://doi.org/10.3390/biology12081144