
Xenopus laevis (Daudin, 1802) as a Model Organism for Bioscience: A Historic Review and Perspective

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. The Advantages of the Xenopus Model
- It is easy to obtain from dedicated companies and to raise and feed in large colonies in the laboratory;
- It is possible to induce, in the female, repeated spawning at any season, and therefore to obtain embryos throughout the year;
- Thanks to their relatively large dimensions (~1.0–1.3 mm in diameter), it is possible to easily handle the oocytes before the fertilization process; moreover, oocytes are released in large numbers, making in vitro fertilization possible;
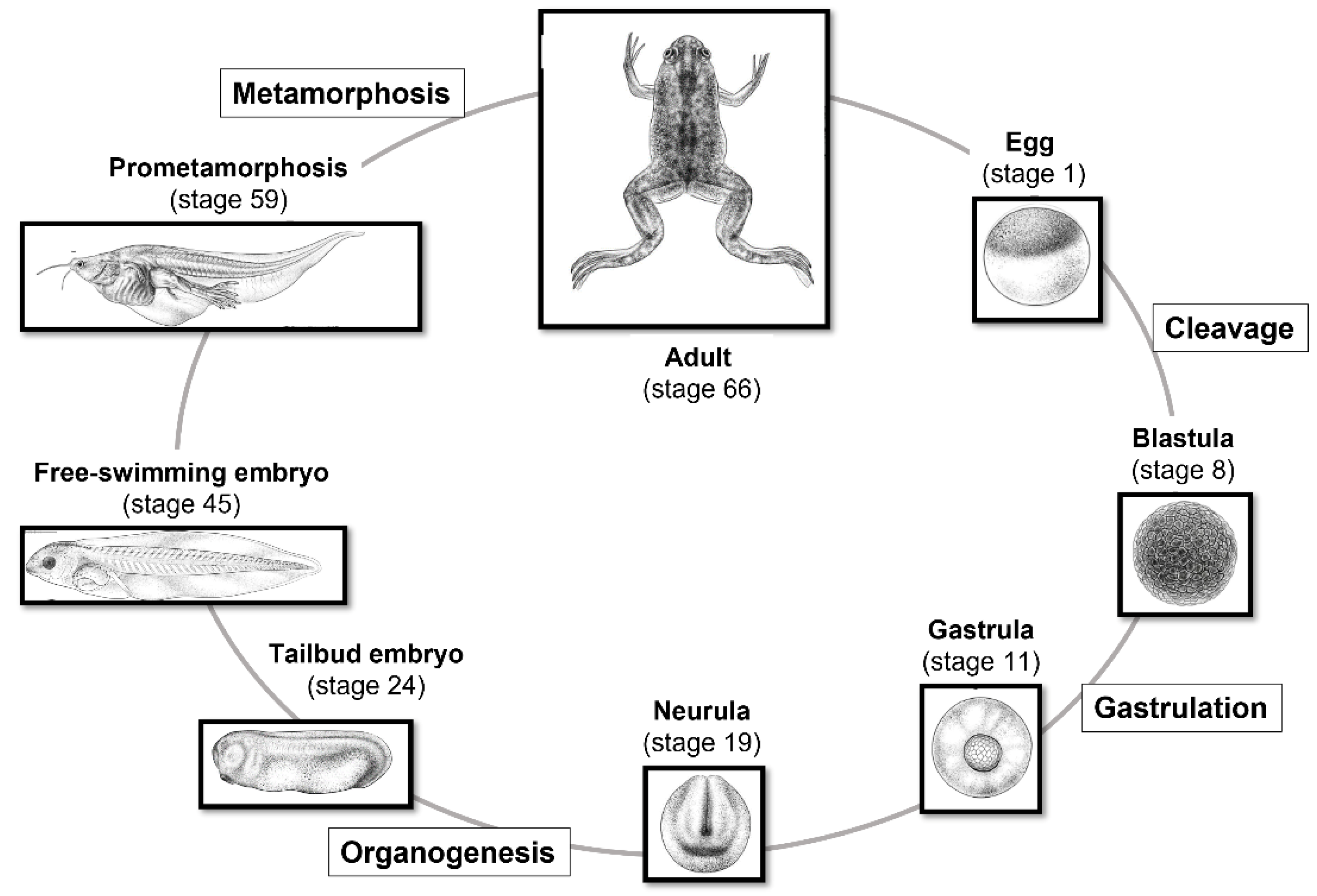
- Since both fertilization and embryonic development take place outside the mother’s body, it is possible to observe and manipulate the process at every stage (Figure 2);
- Its early development is very fast, making it possible to study the developmental progress, until the tadpole stage, in approximately four days at 22/24 °C;
- It is an excellent model for the study of body axis formation;
- It has a high degree of conservation of most essential cellular and molecular mechanisms.
3. History of Xenopus Use in Laboratory
4. Xenopus in Developmental Studies
4.1. Xenopus in the Study of Oogenesis and Role of Cytoskeletal Proteins
4.2. Xenopus as a Model for the Study of Organogenesis
4.3. Interactions of Specific Proteins in Organ Formation: The Eye and Kidney
4.4. Xenopus and Studies of Apoptosis in Development
5. Xenopus in Environmental Studies
5.1. Xenopus in the Study of Microplastic Environmental Pollution
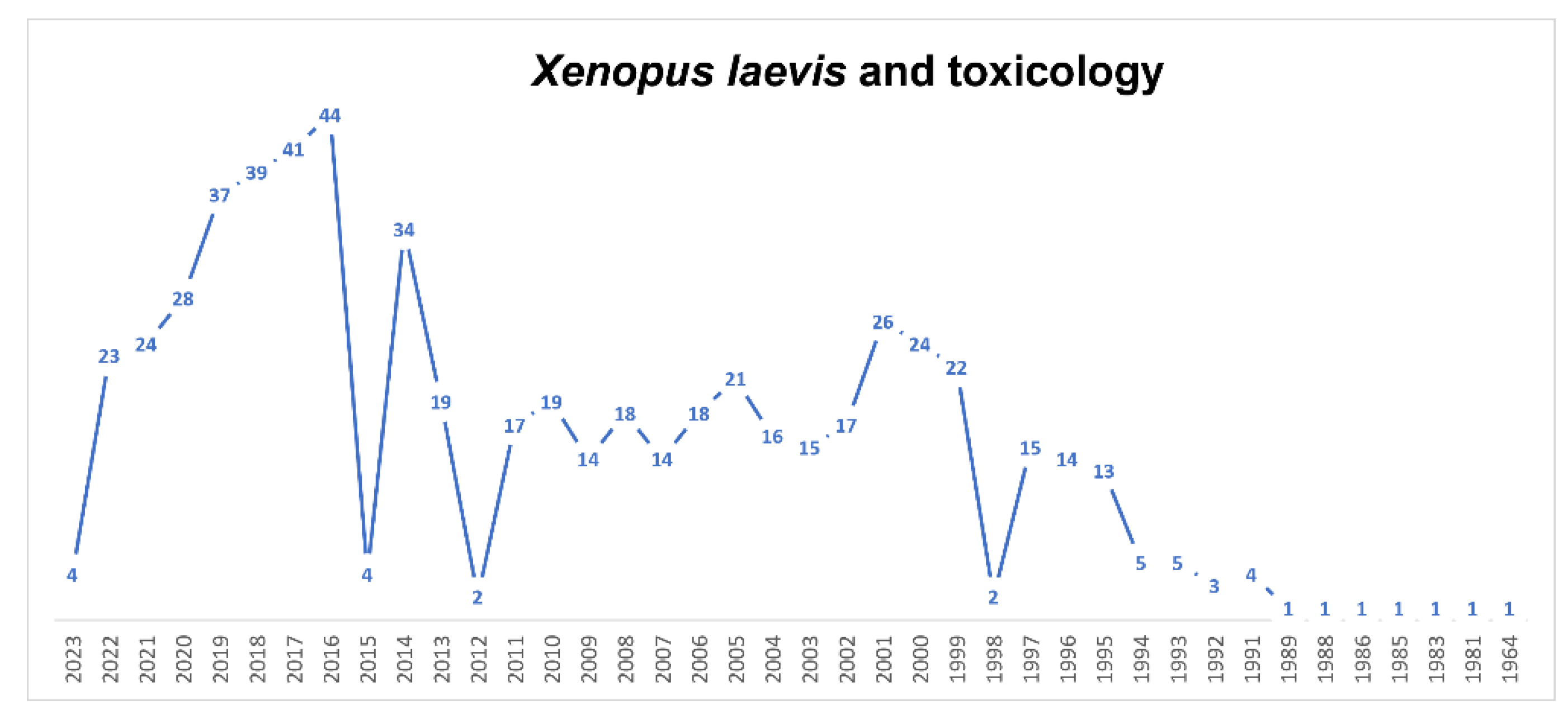
5.2. Xenopus in the Study of Embryo Toxicology
6. Xenopus in Emerging Areas of Bioscience
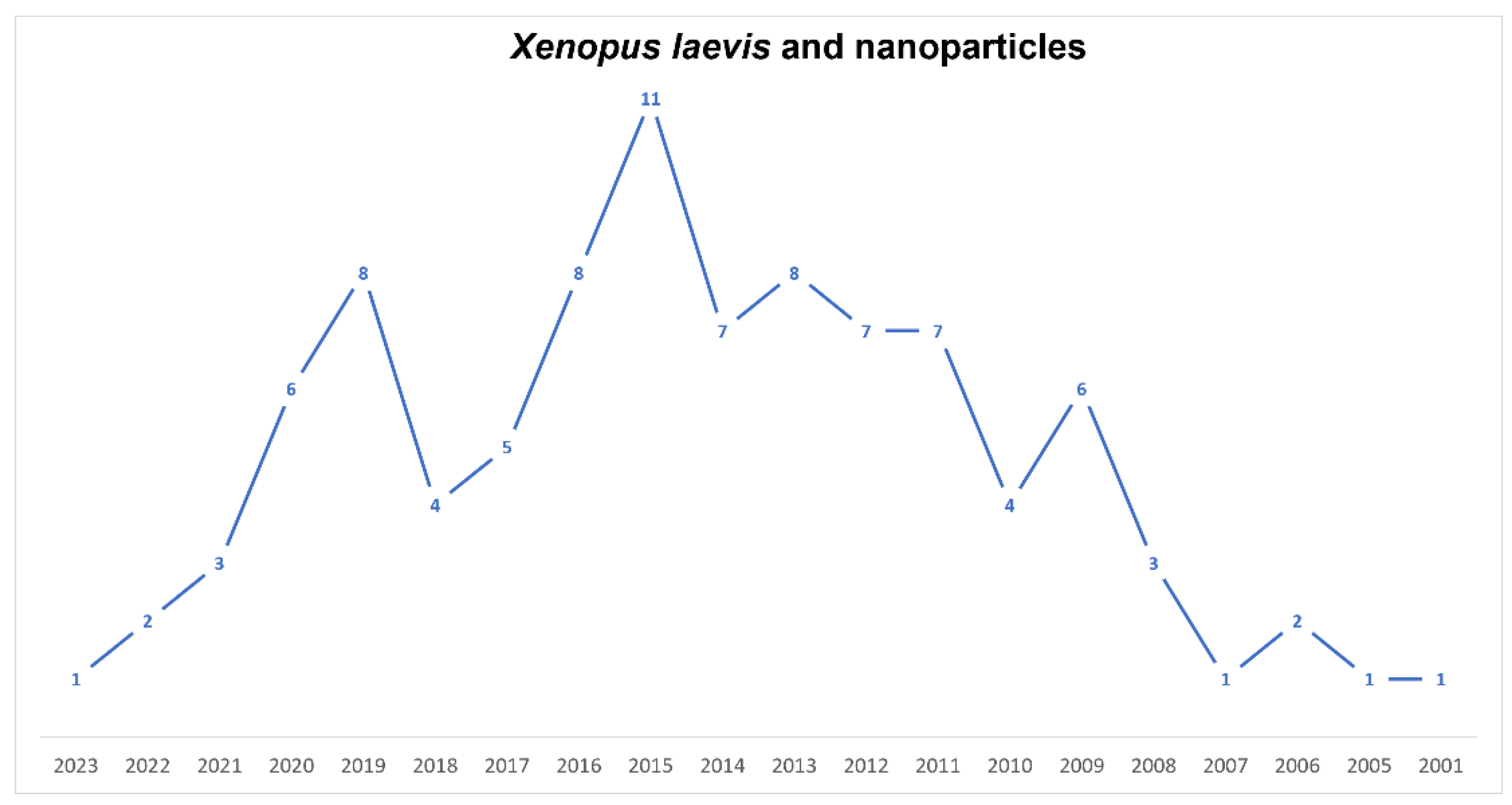
Nanoscience and Xenopus Developmental Studies
7. Xenopus in Immunology, Genetic and Disease Studies
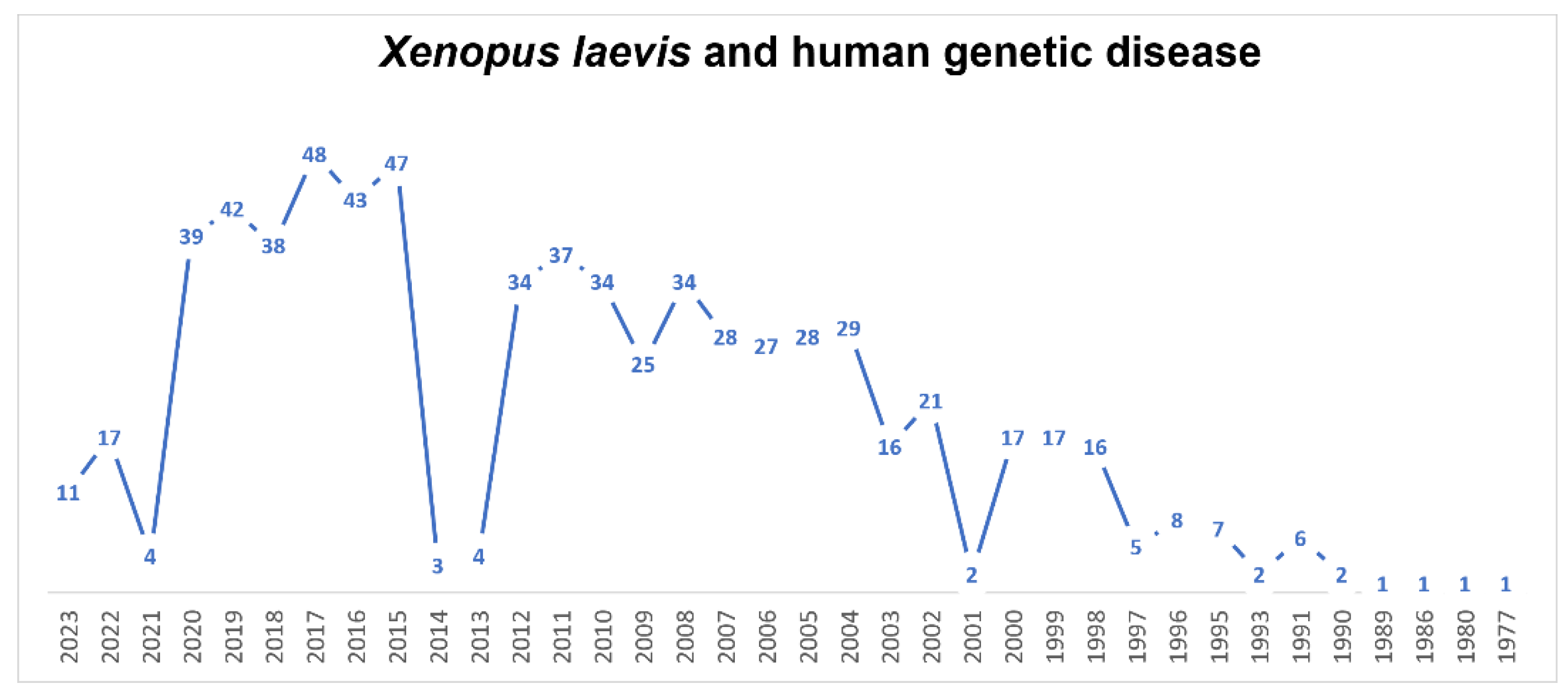
7.1. Xenopus laevis as a Model for Human Genetic Disease
7.2. Xenopus laevis in Immunologic Studies
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gurdon, J.B.; Hopwood, N. The introduction of Xenopus laevis into developmental biology: Of empire, pregnancy testing and ribosomal genes. Int. J. Dev. Biol. 2000, 44, 43–50. [Google Scholar] [PubMed]
- Beck, C.W.; Slack, J.M. An amphibian with ambition: A new role for Xenopus in the 21st century. Genome Biol. 2001, 2, reviews1029.1. [Google Scholar] [CrossRef] [Green Version]
- Tadjuidje, E.; Heasman, J. Xenopus as an Experimental Organism; John Wiley & Sons: Hoboken, NJ, USA, 2010. [Google Scholar] [CrossRef]
- Showell, C.; Conlon, F.L. Egg collection and in vitro fertilization of the western clawed frog Xenopus tropicalis. Cold Spring Harb. Protoc. 2009, 2009, pdb.prot5293. [Google Scholar] [CrossRef] [Green Version]
- American Society for Testing and Materials. Standard guide for Conducting frog Embryo Teratogenesis Assay-Xenopus, ASTM E1439-98: Annual Book of ASTM Standards; ASTM: Philadelphia, PA, USA, 2014; Volume 11.05, pp. 826–836. [Google Scholar]
- Nieuwkoop, P.D.; Faber, J. Normal Table of Xenopus laevis (Daudin): A Systematical and Chronologica Survey of the Development from the Fertilized Egg till the End of Metamorphosis; Elsevier: Amsterdam, The Netherlands, 1956. [Google Scholar]
- De Marco, N.; Tussellino, M.; Carotenuto, R.; Ronca, R.; Rizzolio, S.; Biffo, S.; Campanella, C. Eukaryotic initiation factor eIF6 modulates the expression of Kermit 2/XGIPC in IGF- regulated eye development. Dev. Biol. 2017, 427, 148–154. [Google Scholar] [CrossRef]
- Tussellino, M.; De Marco, N.; Campanella, C.; Carotenuto, R. Involvement of the eukaryotic initiation factor 6 and kermit2/gipc2 in Xenopus laevis pronephros formation. Int. J. Dev. Biol. 2012, 56, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Carotenuto, R.; Capriello, T.; Cofone, R.; Galdiero, G.; Fogliano, C.; Ferrandino, I. Impact of copper in Xenopus laevis liver: Histological damages and atp7b downregulation. Ecotoxicol. Environ. Saf. 2020, 188, 109940. [Google Scholar] [CrossRef] [PubMed]
- Blum, M.; Ott, T. Xenopus: An Undervalued Model Organism to Study and Model Human Genetic Disease. Cells Tissues Organs 2019, 205, 303–313. [Google Scholar] [CrossRef]
- Hoppler, S.; Conlon, F.L. Xenopus: Experimental Access to Cardiovascular Development, Regeneration Discovery, and Cardiovascular Heart-Defect Modeling. Cold Spring Harb. Perspect. Biol. 2020, 12, a037200. [Google Scholar] [CrossRef]
- Nenni, M.J.; Fisher, M.E.; James-Zorn, C.; Pells, T.J.; Ponferrada, V.; Chu, S.; Fortriede, J.D.; Burns, K.A.; Wang, Y.; Lotay, V.S.; et al. Xenbase: Facilitating the Use of Xenopus to Model Human Disease. Front. Physiol. 2019, 10, 154. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Xu, X. Application of Xenopus laevis in ecotoxicology (I)—Introduction and quality control of laboratory animal. Chin. Sci. Bull. 2006, 51, 1273–1280. [Google Scholar] [CrossRef]
- Burlibașa, L.; Gavrilă, L. Amphibians as model organisms for study environmental genotoxicity. Appl. Ecol. Environ. Res. 2011, 9, 1–15. [Google Scholar] [CrossRef]
- Noble, G.K. The Biology of the Amphibia; McGraw-Hill: New York, NY, USA, 1931. [Google Scholar]
- Hamburger, V. The Heritage of Experimental Embryology: Hans Spemann and the Organizer; Oxford University Press: New York, NY, USA; Oxford, UK, 1988. [Google Scholar]
- Gard, D.L. Gamma-tubulin is asymmetrically distributed in the cortex of Xenopus oocytes. Dev. Biol. 1994, 161, 131–140. [Google Scholar] [CrossRef]
- Takagi, C.; Sakamaki, K.; Morita, H.; Hara, Y.; Suzuki, M.; Kinoshita, N.; Ueno, N. Transgenic Xenopus laevis for live imaging in cell and developmental biology. Develop. Dev. Growth Differ. 2013, 55, 422–433. [Google Scholar] [CrossRef]
- Moosmann, J.; Ershov, A.; Altapova, V.; Baumbach, T.; Prasad, M.S.; LaBonne, C.; Xiao, X.; Kashef, J.; Hofmann, R. X-ray phase-contrast in vivo microtomography probes new aspects of Xenopus gastrulation. Nature 2013, 497, 374–377. [Google Scholar] [CrossRef] [Green Version]
- Moosmann, J.; Ershov, A.; Weinhardt, V.; Baumbach, T.; Prasad, M.S.; LaBonne, C.; Xiao, X.; Kashef, J.; Hofmann, R. Time-lapse X-ray phase-contrast microtomography for in vivo imaging and analysis of morphogenesis. Nat. Protoc. 2014, 9, 294–304. [Google Scholar] [CrossRef]
- Session, A.M.; Uno, Y.; Kwon, T.; Chapman, J.A.; Toyoda, A.; Takahashi, S.; Fukui, A.; Hikosaka, A.; Suzuki, A.; Kondo, M.; et al. Genome evolution in the allotetraploid frog Xenopus laevis. Nature 2016, 538, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Baldessari, D.; Shin, Y.; Krebs, O.; König, R.; Koide, T.; Vinayagam, A.; Fenger, U.; Mochii, M.; Terasaka, C.; Kitayama, A.; et al. Global gene expression profiling and cluster analysis in Xenopus laevis. Mech. Dev. 2005, 122, 441–475. [Google Scholar] [CrossRef]
- Sczyrba, A.; Beckstette, M.; Brivanlou, A.H.; Giegerich, R.; Altmann, C.R. XenDB: Full length cDNA prediction and cross species mapping in Xenopus laevis. BMC Genom. 2005, 6, 123. [Google Scholar] [CrossRef] [Green Version]
- Amin, N.M.; Tandon, P.; Osborne Nishimura, E.; Conlon, F.L. RNA-seq in the tetraploid Xenopus laevis enables genome-wide insight in a classic developmental biology model organism. Methods 2014, 66, 398–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Bertke, M.M.; Champion, M.M.; Zhu, G.; Huber, P.W.; Dovichi, N.J. Quantitative proteomics of Xenopus laevis embryos: Expression kinetics of nearly 4000 proteins during early development. Sci. Rep. 2014, 4, 4365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearl, E.J.; Grainger, R.M.; Guille, M.; Horb, M.E. Development of Xenopus resource centers: The National Xenopus Resource and the European Xenopus Resource Center. Genesis 2012, 50, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Naert, T.; Tulkens, D.; Edwards, N.A.; Carron, M.; Shaidani, N.I.; Wlizla, M.; Boel, A.; Demuynck, S.; Horb, M.E.; Coucke, P.; et al. Maximizing CRISPR/Cas9 phenotype penetrance applying predictive modeling of editing outcomes in Xenopus and zebrafish embryos. Sci. Rep. 2020, 10, 14662. [Google Scholar] [CrossRef] [PubMed]
- Guille, M.; Grainger, R. Genetics and Gene Editing Methods in Xenopus laevis and Xenopus tropicalis. Cold Spring Harb. Protoc. 2022; advance online publication. [Google Scholar] [CrossRef]
- Hardwick, L.J.; Philpott, A. An oncologist’s friend: How Xenopus contributes to cancer research. Dev. Biol. 2015, 408, 180–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; Kuriyama, S.; Itoh, G.; Kohyama, A.; Iwabuchi, Y.; Shibata, H.; Yashiro, M.; Aiba, N. Identification of anti-cancer chemical compounds using Xenopus embryos. Cancer Sci. 2016, 107, 803–811. [Google Scholar] [CrossRef] [Green Version]
- Wallingford, J.B. Tumors in tadpoles: The Xenopus embryo as a model system for the study of tumorigenesis. Trends Genet. 1999, 15, 385–388. [Google Scholar] [CrossRef]
- Griffin, J.N.; Liu, K.J.; Sempou, E. Editorial: Xenopus Models of Organogenesis and Disease. Front. Physiol. 2020, 11, 534. [Google Scholar] [CrossRef]
- Gundersen, C.; Miledi, R.; Parker, I. Messenger RNA from human brain induces drug- and voltage-operated channels in Xenopus oocytes. Nature 1984, 308, 421–424. [Google Scholar] [CrossRef]
- Mais, C.; McStay, B.; Scheer, U. On the formation of amplified nucleoli during early Xenopus oogenesis. J. Struct. Biol. 2002, 140, 214–226. [Google Scholar] [CrossRef]
- Kloc, M.; Larabell, C.; Chan, A.P.; Etkin, L.D. Contribution of METRO pathway localized molecules to the organization of the germ cell lineage. Mech. Dev. 1998, 75, 81–93. [Google Scholar] [CrossRef]
- Kloc, M.; Etkin, L.D. Two distinct pathways for the localization of RNAs at the vegetal cortex in Xenopus oocytes. Development 1995, 121, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Carotenuto, R.; Tussellino, M. Xenopus laevis oocyte as a model for the study of the cytoskeleton. Comptes Rendus Biol. 2018, 341, 219–227. [Google Scholar] [CrossRef]
- Carotenuto, R.; Maturi, G.; Infante, V.; Capriglione, T.; Petrucci, T.C.; Campanella, C. A novel protein cross-reacting with antibodies against spectrin is localised in the nucleoli of amphibian oocytes. J. Cell Sci. 1997, 110, 2683–2690. [Google Scholar] [CrossRef]
- Vaccaro, M.C.; Gigliotti, S.; Graziani, F.; Carotenuto, R.; De Angelis, C.; Tussellino, M.; Campanella, C. A transient asymmetric distribution of XNOA 36 mRNA and the associated spectrin network bisects Xenopus laevis stage I oocytes along the future A/V axis. Eur. J. Cell Biol. 2010, 89, 525–536. [Google Scholar] [CrossRef]
- Gard, D.L.; Cha, B.J.; King, E. The organization and animal-vegetal asymmetry of cytokeratin filaments in stage VI Xenopus oocytes is dependent upon F-actin and microtubules. Dev. Biol. 1997, 184, 95–114. [Google Scholar] [CrossRef] [Green Version]
- Klymkowsky, M.W. Intermediate filament organization, reorganization, and function in the clawed frog Xenopus. Curr. Top. Dev. Biol. 1995, 31, 455–486. [Google Scholar] [CrossRef] [PubMed]
- Carotenuto, R.; Petrucci, T.C.; Correas, I.; Vaccaro, M.C.; De Marco, N.; Dale, B.; Wilding, M. Protein 4.1 and its interaction with other cytoskeletal proteins in Xenopus laevis oogenesis. Eur. J. Cell Biol. 2009, 88, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Hovland, R.; Hesketh, J.E.; Pryme, I.F. The compartmentalization of protein synthesis: Importance of cytoskeleton and role in mRNA targeting. Int. J. Biochem. Cell Biol. 1996, 28, 1089–1105. [Google Scholar] [CrossRef]
- Jansen, R.P. RNA-cytoskeletal associations. FASEB J. 1999, 13, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Chierchia, L.; Tussellino, M.; Guarino, D.; Carotenuto, R.; DeMarco, N.; Campanella, C.; Biffo, S.; Vaccaro, M.C. Cytoskeletal proteins associate with components of the ribosomal maturation and translation apparatus in Xenopus stage I oocytes. Zygote 2015, 23, 669–682. [Google Scholar] [CrossRef]
- Lo, K.Y.; Li, Z.; Bussiere, C.; Bresson, S.; Marcotte, E.M.; Johnson, A.W. Defining the pathway of cytoplasmic maturation of the 60S ribosomal subunit. Mol. Cell 2010, 39, 196–208. [Google Scholar] [CrossRef] [Green Version]
- Brina, D.; Grosso, S.; Miluzio, A.; Biffo, S. Translational control by 80S formation and 60S availability: The central role of eIF6, a rate limiting factor in cell cycle progression and tumorigenesis. Cell Cycle 2011, 10, 3441–3446. [Google Scholar] [CrossRef] [PubMed]
- Heasman, J.; Quarmby, J.; Wylie, C.C. The mitochondrial cloud of Xenopus oocytes: The source of germinal granule material. Dev. Biol. 1984, 105, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Wilding, M.; Carotenuto, R.; Infante, V.; Dale, B.; Marino, M.; Di Matteo, L.; Campanella, C. Confocal microscopy analysis of the activity of mitochondria contained within the ‘mitochondrial cloud’ during oogenesis in Xenopus laevis. Zygote 2001, 9, 347–352. [Google Scholar] [CrossRef]
- Gard, D.L. Confocal microscopy and 3-D reconstruction of the cytoskeleton of Xenopus oocytes. Microsc. Res. Tech. 1999, 44, 388–414. [Google Scholar] [CrossRef]
- Kloc, M.; Etkin, L.D. Apparent continuity between the messenger transport organizer and late RNA localization pathways during oogenesis in Xenopus. Mech. Dev. 1998, 73, 95–106. [Google Scholar] [CrossRef]
- Carotenuto, R.; Vaccaro, M.C.; Capriglione, T.; Petrucci, T.C.; Campanella, C. alpha-Spectrin has a stage-specific asymmetrical localization during Xenopus oogenesis. Mol. Reprod. Dev. 2000, 55, 229–239. [Google Scholar] [CrossRef]
- Bolívar, J.; Díaz, I.; Iglesias, C.; Valdivia, M.M. Molecular cloning of a zinc finger autoantigen transiently associated with interphase nucleolus and mitotic centromeres and midbodies. Orthologous proteins with nine CXXC motifs highly conserved from nematodes to humans. J. Biol. Chem. 1999, 274, 36456–36464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, G.H.; Melton, D.A. Processed Vg1 protein is an axial mesoderm inducer in Xenopus. Cell 1993, 74, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Tatone, C.; Carotenuto, R.; Colonna, R.; Chaponnier, C.; Gabbiani, G.; Giorgi, M. Spectrin and ankyrin-like proteins in the egg of Disco-glossus pictus (Anura): Their identification and localization in the site of sperm entrance versus the rest of the egg. Dev. Growth Differ. 1993, 35, 161–171. [Google Scholar] [CrossRef]
- Edwards-Faret, G.; González-Pinto, K.; Cebrián-Silla, A.; Peñailillo, J.; García-Verdugo, J.M.; Larraín, J. Cellular response to spinal cord injury in regenerative and non-regenerative stages in Xenopus laevis. Neural Dev. 2021, 16, 2. [Google Scholar] [CrossRef] [PubMed]
- Horb, M.E.; Slack, J.M. Endoderm specification and differentiation in Xenopus embryos. Dev. Biol. 2001, 236, 330–343. [Google Scholar] [CrossRef] [Green Version]
- Sokol, S.Y. Mechanotransduction During Vertebrate Neurulation. Curr. Top. Dev. Biol. 2016, 117, 359–376. [Google Scholar] [CrossRef] [PubMed]
- Borodinsky, L.N. Xenopus laevis as a Model Organism for the Study of Spinal Cord Formation, Development, Function and Regeneration. Front. Neural Circuits 2017, 11, 90. [Google Scholar] [CrossRef] [Green Version]
- Slater, P.G.; Palacios, M.; Larraín, J. Xenopus, a Model to Study Wound Healing and Regeneration: Experimental Approaches. Cold Spring Harb. Protoc. 2021, 2021, top100966. [Google Scholar] [CrossRef]
- Aztekin, C.; Hiscock, T.W.; Marioni, J.C.; Gurdon, J.B.; Simons, B.D.; Jullien, J. Identification of a regeneration-organizing cell in the Xenopus tail. Science 2019, 364, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Lee-Liu, D.; Méndez-Olivos, E.E.; Muñoz, R.; Larraín, J. The African clawed frog Xenopus laevis: A model organism to study regeneration of the central nervous system. Neurosci. Lett. 2017, 652, 82–93. [Google Scholar] [CrossRef]
- Xiang, Y.; Li, Y.; Zhang, Z.; Cui, K.; Wang, S.; Yuan, X.B.; Wu, C.P.; Poo, M.M.; Duan, S. Nerve growth cone guidance mediated by G protein-coupled receptors. Nat. Neurosci. 2002, 5, 843–848. [Google Scholar] [CrossRef]
- Song, H.J.; Ming, G.L.; Poo, M.M. cAMP-induced switching in turning direction of nerve growth cones. Nature 1997, 388, 275–279. [Google Scholar] [CrossRef]
- Dickson, B.J.; Senti, K.A. Axon guidance: Growth cones make an unexpected turn. Curr. Biol. 2002, 12, R218–R220. [Google Scholar] [CrossRef] [Green Version]
- Lowery, L.; Vactor, D. The trip of the tip: Understanding the growth cone machinery. Nat. Rev. Mol. Cell Biol. 2009, 10, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Erclik, T.; Hartenstein, V.; McInnes, R.R.; Lipshitz, H.D. Eye evolution at high resolution: The neuron as a unit of homology. Dev. Biol. 2009, 332, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Gehring, W.J. New perspectives on eye development and the evolution of eyes and photoreceptors. J. Hered. 2005, 96, 171–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuber, M.E.; Gestri, G.; Viczian, A.S.; Barsacchi, G.; Harris, W.A. Specification of the vertebrate eye by a network of eye field transcription factors. Development 2003, 130, 5155–5167. [Google Scholar] [CrossRef] [Green Version]
- Pera, E.M.; Ikeda, A.; Eivers, E.; De Robertis, E.M. Integration of IGF, FGF, and anti-BMP signals via Smad1 phosphorylation in neural induction. Genes Dev. 2003, 17, 3023–3028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard-Parpaillon, L.; Héligon, C.; Chesnel, F.; Boujard, D.; Philpott, A. The IGF pathway regulates head formation by inhibiting Wnt signaling in Xenopus. Dev. Biol. 2002, 244, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; O’Donnell, M.; Gitler, A.D.; Klein, P.S. Kermit 2/XGIPC, an IGF1 receptor interacting protein, is required for IGF signaling in Xenopus eye development. Development 2006, 133, 3651–3660. [Google Scholar] [CrossRef] [Green Version]
- Vize, P.D.; Seufert, D.W.; Carroll, T.J.; Wallingford, J.B. Model systems for the study of kidney development: Use of the pronephros in the analysis of organ induction and patterning. Dev. Biol. 1997, 188, 189–204. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.A. Xenopus: A prince among models for pronephric kidney development. J. Am. Soc. Nephrol. 2005, 16, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Lienkamp, S.S. Using Xenopus to study genetic kidney diseases. Semin. Cell Dev. Biol. 2016, 51, 117–124. [Google Scholar] [CrossRef]
- Taira, M.; Otani, H.; Jamrich, M.; Dawid, I.B. Expression of the LIM class homeobox gene Xlim-1 in pronephros and CNS cell lineages of Xenopus embryos is affected by retinoic acid and exogastrulation. Development 1994, 120, 1525–1536. [Google Scholar] [CrossRef] [PubMed]
- Carroll, T.J.; Vize, P.D. Synergism between Pax-8 and lim-1 in embryonic kidney development. Dev. Biol. 1999, 214, 46–59. [Google Scholar] [CrossRef] [Green Version]
- Cirio, M.C.; Hui, Z.; Haldin, C.E.; Cosentino, C.C.; Stuckenholz, C.; Chen, X.; Hong, S.K.; Dawid, I.B.; Hukriede, N.A. Lhx1 is required for specification of the renal progenitor cell field. PLoS ONE 2011, 6, e18858. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.S.; Wang, T.T.; Zhang, S.L.; Chen, X.; Carrière, S. Catecholamines and angiotensinogen gene expression in kidney proximal tubular cells. Mol. Cell. Biochem. 2000, 212, 73–79. [Google Scholar] [CrossRef]
- Kobayashi, A.; Kwan, K.M.; Carroll, T.J.; McMahon, A.P.; Mendelsohn, C.L.; Behringer, R.R. Distinct and sequential tissue-specific activities of the LIM-class homeobox gene Lim1 for tubular morphogenesis during kidney development. Development 2005, 132, 2809–2823. [Google Scholar] [CrossRef] [Green Version]
- Droz, S.T.; McLaughlin, K.A. Use of Xenopus Frogs to Study Renal Development/Repair. Results Probl. Cell Differ. 2017, 60, 77–107. [Google Scholar] [CrossRef]
- Coles, H.S.; Burne, J.F.; Raff, M.C. Large-scale normal cell death in the developing rat kidney and its reduction by epidermal growth factor. Development 1993, 118, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Burek, M.J.; Oppenheim, R.W. Programmed cell death in the developing nervous system. Brain Pathol. 1996, 6, 427–446. [Google Scholar] [CrossRef]
- Hensey, C.; Gautier, J. A developmental timer that regulates apoptosis at the onset of gastrulation. Mech. Dev. 1997, 69, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M.D.; Weil, M.; Raff, M.C. Programmed cell death in animal development. Cell 1997, 88, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Hensey, C.; Gautier, J. Programmed cell death during Xenopus development: A spatio-temporal analysis. Dev. Biol. 1998, 203, 36–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkielstein, C.V.; Lewellyn, A.L.; Maller, J.L. The midblastula transition in Xenopus embryos activates multiple pathways to prevent apoptosis in response to DNA damage. Proc. Natl. Acad. Sci. USA 2001, 98, 1006–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeo, W.; Gautier, J. A role for programmed cell death during early neurogenesis in xenopus. Dev. Biol. 2003, 260, 31–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Marco, N.; Iannone, L.; Carotenuto, R.; Biffo, S.; Vitale, A.; Campanella, C. p27(BBP)/eIF6 acts as an anti-apoptotic factor upstream of Bcl-2 during Xenopus laevis development. Cell Death Differ. 2010, 17, 360–372. [Google Scholar] [CrossRef] [PubMed]
- Pekmezekmek, A.B.; Emre, M.; Erdogan, S.; Yilmaz, B.; Tunc, E.; Sertdemir, Y.; Emre, Y. Effects of high-molecular-weight polyvinyl chloride on Xenopus laevis adults and embryos: The mRNA expression profiles of Myf5, Esr1, Bmp4, Pax6, and Hsp70 genes during early embryonic development. Environ. Sci. Pollut. Res. Int. 2022, 29, 14767–14779. [Google Scholar] [CrossRef] [PubMed]
- Ruthsatz, K.; Domscheit, M.; Engelkes, K.; Vences, M. Microplastics ingestion induces plasticity in digestive morphology in larvae of Xenopus laevis. Comparative biochemistry and physiology. Part A Mol. Integr. Physiol. 2022, 269, 111210. [Google Scholar] [CrossRef]
- Ruthsatz, K.; Schwarz, A.; Gomez-Mestre, I.; Meyer, R.; Domscheit, M.; Bartels, F.; Schaeffer, S.M.; Engelkes, K. Life in plastic, it’s not fantastic: Sublethal effects of polyethylene microplastics ingestion throughout amphibian metamorphosis. Sci. Total Environ. 2023, 885, 163779. [Google Scholar] [CrossRef]
- De Felice, B.; Bacchetta, R.; Santo, N.; Tremolada, P.; Parolini, M. Polystyrene microplastics did not affect body growth and swimming activity in Xenopus laevis tadpoles. Environ. Sci. Pollut. Res. Int. 2018, 25, 34644–34651. [Google Scholar] [CrossRef]
- Bonfanti, P.; Colombo, A.; Saibene, M.; Motta, G.; Saliu, F.; Catelani, T.; Mehn, D.; La Spina, R.; Ponti, J.; Cella, C.; et al. Microplastics from miscellaneous plastic wastes: Physico-chemical characterization and impact on fish and amphibian development. Ecotoxicol. Environ. Saf. 2021, 225, 112775. [Google Scholar] [CrossRef]
- Bacchetta, R.; Winkler, A.; Santo, N.; Tremolada, P. The Toxicity of Polyester Fibers in Xenopuslaevis. Water 2021, 13, 3446. [Google Scholar] [CrossRef]
- Venâncio, C.; Gabriel, A.; Oliveira, M.; Lopes, I. Feeding exposure and feeding behaviour as relevant approaches in the assessment of the effects of micro(nano)plastics to early life stages of amphibians. Environ. Res. 2022, 212, 113476. [Google Scholar] [CrossRef]
- Wheeler, G.N.; Brändli, A.W. Simple vertebrate models for chemical genetics and drug discovery screens: Lessons from zebrafish and Xenopus. Dev. Dyn. 2009, 238, 1287–1308. [Google Scholar] [CrossRef]
- Tussellino, M.; Ronca, R.; Formiggini, F.; De Marco, N.; Fusco, S.; Netti, P.A.; Carotenuto, R. Polystyrene nanoparticles affect Xenopus laevis development. J. Nanopart. Res. 2015, 17, 70. [Google Scholar] [CrossRef]
- Carotenuto, R.; Tussellino, M.; Ronca, R.; Benvenuto, G.; Fogliano, C.; Fusco, S.; Netti, P.A. Toxic effects of SiO2NPs in early embryogenesis of Xenopuslaevis. Chemosphere 2022, 289, 133233. [Google Scholar] [CrossRef]
- Fort, D.J.; Mathis, M. Frog Embryo Teratogenesis Assay-Xenopus (FETAX): Use in Alternative Preclinical Safety Assessment. Cold Spring Harb. Protoc. 2018, 2018, pdb-prot098319. [Google Scholar] [CrossRef] [PubMed]
- Hoke, R.A.; Ankley, G.T. Application of frog embryo teratogenesis assay-Xenopus to ecological risk assessment. Environ. Toxicol. Chem. 2005, 24, 2677–2690. [Google Scholar] [CrossRef] [PubMed]
- Babalola, O.O.; Truter, J.C.; Van Wyk, J.H. Lethal and Teratogenic Impacts of Imazapyr, Diquat Dibromide, and Glufosinate Ammonium Herbicide Formulations Using Frog Embryo Teratogenesis Assay-Xenopus (FETAX). Arch. Environ. Contam. Toxicol. 2021, 80, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Huyck, R.W.; Nagarkar, M.; Olsen, N.; Clamons, S.E.; Saha, M.S. Methylmercury exposure during early Xenopus laevis development affects cell proliferation and death but not neural progenitor specification. Neurotoxicology Teratol. 2015, 47, 102–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouchet, F.; Teaniniuraitemoana, V.; Baudrimont, M.; Daffe, G.; Gauthier, L.; Gonzalez, P. Recovery capabilities of Xenopus laevis after exposure to Cadmium and Zinc. Chemosphere 2015, 139, 117–125. [Google Scholar] [CrossRef]
- Gao, J.; Ruan, H.; Qi, X.; Guo, X.; Zheng, J.; Liu, C.; Fang, Y.; Huang, M.; Xu, M.; Shen, W. Increased apoptosis and abnormal visual behavior by histone modifications with exposure to para-xylene in developing Xenopus. Neuroscience 2016, 331, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Luo, Y.; Ding, N.; Gao, J.; Wang, X.; Shen, W. d-Glucuronolactone attenuates para-xylene-induced defects in neuronal development and plasticity in Xenopus tectum in vivo. Toxicology 2020, 430, 152341. [Google Scholar] [CrossRef] [PubMed]
- Cruz, S.L.; Balster, R.L.; Woodward, J.J. Effects of volatile solvents on recombinant N-methyl-D-aspartate receptors expressed in Xenopus oocytes. Br. J. Pharmacol. 2000, 131, 1303–1308. [Google Scholar] [CrossRef] [Green Version]
- van Kleef, R.G.; Vijverberg, H.P.; Westerink, R.H. Selective inhibition of human heteromeric alpha9alpha10 nicotinic acetylcholine receptors at a low agonist concentration by low concentrations of ototoxic organic solvents. Toxicol. Vitr. 2008, 22, 1568–1572. [Google Scholar] [CrossRef]
- Christin, M.S.; Ménard, L.; Gendron, A.D.; Ruby, S.; Cyr, D.; Marcogliese, D.J.; Rollins-Smith, L.; Fournier, M. Effects of agricultural pesticides on the immune system of Xenopus laevis and Rana pipiens. Aquat. Toxicol. 2004, 67, 33–43. [Google Scholar] [CrossRef]
- Asouzu Johnson, J.; Ihunwo, A.; Chimuka, L.; Mbajiorgu, E.F. Cardiotoxicity in African clawed frog (Xenopus laevis) sub-chronically exposed to environmentally relevant atrazine concentrations: Implications for species survival. Aquat. Toxicol. 2019, 213, 105218. [Google Scholar] [CrossRef] [PubMed]
- Bonfanti, P.; Colombo, A.; Orsi, F.; Nizzetto, I.; Andrioletti, M.; Bacchetta, R.; Mantecca, P.; Fascio, U.; Vailati, G.; Vismara, C. Comparative teratogenicity of chlorpyrifos and malathion on Xenopus laevis development. Aquat. Toxicol. 2004, 70, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Hayes, T.B.; Khoury, V.; Narayan, A.; Nazir, M.; Park, A.; Brown, T.; Adame, L.; Chan, E.; Buchholz, D.; Stueve, T.; et al. Atrazine induces complete feminization and chemical castration in male African clawed frogs (Xenopus laevis). Proc. Natl. Acad. Sci. USA 2010, 107, 4612–4617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimayi, C.; Odusanya, D.; Weiss, J.M.; de Boer, J.; Chimuka, L.; Mbajiorgu, F. Effects of environmentally relevant sub-chronic atrazine concentrations on African clawed frog (Xenopus laevis) survival, growth and male gonad development. Aquat. Toxicol. 2018, 199, 1–11. [Google Scholar] [CrossRef]
- Sai, L.; Li, Y.; Zhang, Y.; Zhang, J.; Qu, B.; Guo, Q.; Han, M.; Jia, Q.; Yu, G.; Li, K.; et al. Distinct m6A methylome profiles in poly(A) RNA from Xenopus laevis testis and that treated with atrazine. Chemosphere 2020, 245, 125631. [Google Scholar] [CrossRef]
- Uçkun, M.; Özmen, M. Evaluating Multiple Biochemical Markers in Xenopus laevis Tadpoles Exposed to the Pesticides Thiacloprid and Trifloxystrobin in Single and Mixed Forms. Environ. Toxicol. Chem. 2021, 40, 2846–2860. [Google Scholar] [CrossRef]
- Li, M.; Zhu, J.; Fang, H.; Wang, M.; Wang, Q.; Zhou, B. Coexposure to environmental concentrations of cis-bifenthrin and graphene oxide: Adverse effects on the nervous system during metamorphic development of Xenopus laevis. J. Hazard. Mater. 2020, 381, 120995. [Google Scholar] [CrossRef] [PubMed]
- Tussellino, M.; Ronca, R.; Carotenuto, R.; Pallotta, M.M.; Furia, M.; Capriglione, T. Chlorpyrifos exposure affects fgf8, sox9, and bmp4 expression required for cranial neural crest morphogenesis and chondrogenesis in Xenopus laevis embryos. Environ. Mol. Mutagen. 2016, 57, 630–640. [Google Scholar] [CrossRef]
- Iuga, A.; Lerner, E.; Shedd, T.R.; van der Schalie, W.H. Rapid responses of a melanophore cell line to chemical contaminants in water. J. Appl. Toxicol. 2009, 29, 346–349. [Google Scholar] [CrossRef]
- Wilkinson, J.L.; Boxall, A.B.A.; Kolpin, D.W.; Leung, K.M.Y.; Lai, R.W.S.; Galbán-Malagón, C.; Adell, A.D.; Mondon, J.; Metian, M.; Marchant, R.A.; et al. Pharmaceutical pollution of the world’s rivers. Proc. Natl. Acad. Sci. USA 2022, 119, e2113947119. [Google Scholar] [CrossRef]
- Derksen, J.G.; Rijs, G.B.; Jongbloed, R.H. Diffuse pollution of surface water by pharmaceutical products. Water Sci. Technol. 2004, 49, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Fatta-Kassinos, D.; Meric, S.; Nikolaou, A. Pharmaceutical residues in environmental waters and wastewater: Current state of knowledge and future research. Anal. Bioanal. Chem. 2011, 399, 251–275. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, S. Removal of pharmaceuticals and personal care products (PPCPs) from wastewater: A review. J. Environ. Manag. 2016, 182, 620–640. [Google Scholar] [CrossRef]
- Islas-Flores, H.; Pérez-Alvaréz, I.; Gómez-Oliván, L.M. Evaluation of Teratogenicity of Pharmaceuticals Using FETAX. Methods Mol. Biol. 2018, 1797, 299–307. [Google Scholar] [CrossRef]
- Richards, S.M.; Cole, S.E. A toxicity and hazard assessment of fourteen pharmaceuticals to Xenopus laevis larvae. Ecotoxicology 2006, 15, 647–656. [Google Scholar] [CrossRef]
- Conners, D.E.; Rogers, E.D.; Armbrust, K.L.; Kwon, J.W.; Black, M.C. Growth and development of tadpoles (Xenopus laevis) exposed to selective serotonin reuptake inhibitors, fluoxetine and sertraline, throughout metamorphosis. Environ. Toxicol. Chem. 2009, 28, 2671–2676. [Google Scholar] [CrossRef]
- Isidori, M.; Piscitelli, C.; Russo, C.; Smutná, M.; Bláha, L. Teratogenic effects of five anticancer drugs on Xenopus laevis embryos. Ecotoxicol. Environ. Saf. 2016, 133, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Cardoso-Vera, J.D.; Islas-Flores, H.; SanJuan-Reyes, N.; Montero-Castro, E.I.; Galar-Martínez, M.; García-Medina, S.; Elizalde-Velázquez, A.; Dublán-García, O.; Gómez-Oliván, L.M. Comparative study of diclofenac-induced embryotoxicity and teratogenesis in Xenopus laevis and Lithobates catesbeianus, using the frog embryo teratogenesis assay: Xenopus (FETAX). Sci. Total Environ. 2017, 574, 467–475. [Google Scholar] [CrossRef]
- Pérez-Alvarez, I.; Islas-Flores, H.; Gómez-Oliván, L.M.; Barceló, D.; López De Alda, M.; Pérez Solsona, S.; Sánchez-Aceves, L.; SanJuan-Reyes, N.; Galar-Martínez, M. Determination of metals and pharmaceutical compounds released in hospital wastewater from Toluca, Mexico, and evaluation of their toxic impact. Environ. Pollut. 2018, 240, 330–341. [Google Scholar] [CrossRef]
- Fogliano, C.; Motta, C.M.; Venditti, P.; Fasciolo, G.; Napolitano, G.; Avallone, B.; Carotenuto, R. Environmental concentrations of a delorazepam-based drug impact on embryonic development of non-target Xenopus laevis. Aquat. Toxicol. 2022, 250, 106244. [Google Scholar] [CrossRef]
- Hussain, S.M.; Warheit, D.B.; Ng, S.P.; Comfort, K.K.; Grabinski, C.M.; Braydich-Stolle, L.K. At the Crossroads of Nanotoxicology in vitro: Past Achievements and Current Challenges. Toxicol. Sci. 2015, 147, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Clift, M.J.; Endes, C.; Vanhecke, D.; Wick, P.; Gehr, P.; Schins, R.P.; Petri-Fink, A.; Rothen-Rutishauser, B. A comparative study of different in vitro lung cell culture systems to assess the most beneficial tool for screening the potential adverse effects of carbon nanotubes. Toxicol. Sci. 2014, 137, 55–64. [Google Scholar] [CrossRef]
- Schimpel, C.; Passegger, C.; Egger, S.; Tam-Amersdorfer, C.; Strobl, H. A novel 3D cell culture model to study the human small intestinal immune landscape. Eur. J. Immunol. 2023, 53, e2250131. [Google Scholar] [CrossRef]
- Giannaccini, M.; Cuschieri, A.; Dente, L.; Raffa, V. Non-mammalian vertebrate embryos as models in nanomedicine. Nanomed. Nanotechnol. Biol. Med. 2014, 10, 703–719. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Zhao, Y.; Nel, A.E.; Lin, S. Zebrafish: An in vivo model for nano EHS studies. Small 2013, 9, 1608–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Lovera, C.; Martínez-Val, J.; Cabezas-Sainz, P.; López, R.; Rubiolo, J.A.; Sánchez, L. In vivo toxicity assays in zebrafish embryos: A pre-requisite for xenograft preclinical studies. Toxicol. Mech. Methods 2019, 29, 478–487. [Google Scholar] [CrossRef]
- Asare, N.; Duale, N.; Slagsvold, H.H.; Lindeman, B.; Olsen, A.K.; Gromadzka-Ostrowska, J.; Meczynska-Wielgosz, S.; Kruszewski, M.; Brunborg, G.; Instanes, C. Genotoxicity and gene expression modulation of silver and titanium dioxide nanoparticles in mice. Nanotoxicology 2016, 10, 312–321. [Google Scholar] [CrossRef]
- Carew, A.C.; Hoque, M.E.; Metcalfe, C.D.; Peyrot, C.; Wilkinson, K.J.; Helbing, C.C. Chronic sublethal exposure to silver nanoparticles disrupts thyroid hormone signaling during Xenopus laevis metamorphosis. Aquat. Toxicol. 2015, 159, 99–108. [Google Scholar] [CrossRef]
- Dubertret, B.; Skourides, P.; Norris, D.J.; Noireaux, V.; Brivanlou, A.H.; Libchaber, A. In vivo imaging of quantum dots encapsulated in phospholipid micelles. Science 2002, 298, 1759–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stylianou, P.; Skourides, P.A. Imaging morphogenesis, in Xenopus with Quantum Dot nanocrystals. Mech. Dev. 2009, 126, 828–841. [Google Scholar] [CrossRef] [PubMed]
- Mogi, K.; Adachi, T.; Izumi, S.; Toyoizumi, R. Visualisation of cerebrospinal fluid flow patterns in albino Xenopus larvae in vivo. Fluids Barriers CNS 2012, 9, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, Y.I.; Mitchell, T.; Smolyakov, G.A.; Osiński, M.; Hartley, R.S. Quantum dot assisted tracking of the intracellular protein Cyclin E in Xenopus laevis embryos. J. Nanobiotechnology 2015, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Umanzor-Alvarez, J.; Wade, E.C.; Gifford, A.; Nontapot, K.; Cruz-Reese, A.; Gotoh, T.; Sible, J.C.; Khodaparast, G.A. Near-infrared laser delivery of nanoparticles to developing embryos: A study of efficacy and viability. Biotechnol. J. 2011, 6, 519–524. [Google Scholar] [CrossRef] [Green Version]
- Galdiero, E.; Falanga, A.; Siciliano, A.; Maselli, V.; Guida, M.; Carotenuto, R.; Tussellino, M.; Lombardi, L.; Benvenuto, G.; Galdiero, S. Daphnia magna and Xenopus laevis as in vivo models to probe toxicity and uptake of quantum dots functionalized with gH625. Int. J. Nanomed. 2017, 12, 2717–2731. [Google Scholar] [CrossRef] [Green Version]
- Carotenuto, R.; Fogliano, C.; Rienzi, M.; Siciliano, A.; Salvatore, M.M.; De Tommaso, G.; Benvenuto, G.; Galdiero, E.; Guida, M. Comparative Toxicological Evaluation of Tattoo Inks on Two Model Organisms. Biology 2021, 10, 1308. [Google Scholar] [CrossRef]
- Zaidi, S.; Choi, M.; Wakimoto, H.; Ma, L.; Jiang, J.; Overton, J.D.; Romano-Adesman, A.; Bjornson, R.D.; Breitbart, R.E.; Brown, K.K.; et al. De novo mutations in histone-modifying genes in congenital heart disease. Nature 2013, 498, 220–223. [Google Scholar] [CrossRef] [Green Version]
- Glessner, J.T.; Bick, A.G.; Ito, K.; Homsy, J.; Rodriguez-Murillo, L.; Fromer, M.; Mazaika, E.; Vardarajan, B.; Italia, M.; Leipzig, J.; et al. Increased frequency of de novo copy number variants in congenital heart disease by integrative analysis of single nucleotide polymorphism array and exome sequence data. Circ. Res. 2014, 115, 884–896. [Google Scholar] [CrossRef]
- Homsy, J.; Zaidi, S.; Shen, Y.; Ware, J.S.; Samocha, K.E.; Karczewski, K.J.; DePalma, S.R.; McKean, D.; Wakimoto, H.; Gorham, J.; et al. De novo mutations in congenital heart disease with neurodevelopmental and other congenital anomalies. Science 2015, 350, 1262–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manheimer, K.B.; Richter, F.; Edelmann, L.J.; D’Souza, S.L.; Shi, L.; Shen, Y.; Homsy, J.; Boskovski, M.T.; Tai, A.C.; Gorham, J.; et al. Robust identification of mosaic variants in congenital heart disease. Hum. Genet. 2018, 137, 183–193. [Google Scholar] [CrossRef]
- Hwang, W.Y.; Marquez, J.; Khokha, M.K. Xenopus: Driving the Discovery of Novel Genes in Patient Disease and Their Underlying Pathological Mechanisms Relevant for Organogenesis. Front. Physiol. 2019, 10, 953. [Google Scholar] [CrossRef] [Green Version]
- Lane, M.; Mis, E.K.; Khokha, M.K. Microinjection of Xenopus tropicalis Embryos. Cold Spring Harb. Protoc. 2022, 2022, pdb.prot107644. [Google Scholar] [CrossRef] [PubMed]
- Treimer, E.; Kalayci, T.; Schumann, S.; Suer, I.; Greco, S.; Schanze, D.; Schmeisser, M.J.; Kühl, S.J.; Zenker, M. Functional characterization of a novel TP53RK mutation identified in a family with Galloway-Mowat syndrome. Hum. Mutat. 2022, 43, 1866–1871. [Google Scholar] [CrossRef]
- Mann, N.; Mzoughi, S.; Schneider, R.; Kühl, S.J.; Schanze, D.; Klämbt, V.; Lovric, S.; Mao, Y.; Shi, S.; Tan, W.; et al. Mutations in PRDM15 Are a Novel Cause of Galloway-Mowat Syndrome. J. Am. Soc. Nephrol. 2021, 32, 580–596. [Google Scholar] [CrossRef]
- Choukair, D.; Eberle, B.; Vick, P.; Hermanns, P.; Weiss, B.; Paramasivam, N.; Schlesner, M.; Lornsen, K.; Roeth, R.; Klutmann, C.; et al. Identification of Transient Receptor Potential Channel 4-Associated Protein as a Novel Candidate Gene Causing Congenital Primary Hypothyroidism. Horm. Res. Paediatr. 2020, 93, 16–29. [Google Scholar] [CrossRef]
- Feehan, J.M.; Stanar, P.; Tam, B.M.; Chiu, C.; Moritz, O.L. Generation and Analysis of Xenopus laevis Models of Retinal Degeneration Using CRISPR/Cas9. Methods Mol. Biol. 2019, 1834, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Macken, W.L.; Godwin, A.; Wheway, G.; Stals, K.; Nazlamova, L.; Ellard, S.; Alfares, A.; Aloraini, T.; AlSubaie, L.; Alfadhel, M.; et al. Biallelic variants in COPB1 cause a novel, severe intellectual disability syndrome with cataracts and variable microcephaly. Genome Med. 2021, 13, 34. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Z.; Chen, M.; Fan, N.; Yang, J.; Liu, L.; Wang, Y.; Liu, X. Mutational Analysis of the TYR and OCA2 Genes in Four Chinese Families with Oculocutaneous Albinism. PLoS ONE 2015, 10, e0125651. [Google Scholar] [CrossRef] [Green Version]
- Viet, J.; Reboutier, D.; Hardy, S.; Lachke, S.A.; Paillard, L.; Gautier-Courteille, C. Modeling ocular lens disease in Xenopus. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2020, 249, 610–621. [Google Scholar] [CrossRef]
- Sega, A.G.; Mis, E.K.; Lindstrom, K.; Mercimek-Andrews, S.; Ji, W.; Cho, M.T.; Juusola, J.; Konstantino, M.; Jeffries, L.; Khokha, M.K.; et al. De novo pathogenic variants in neuronal differentiation factor 2 (NEUROD2) cause a form of early infantile epileptic encephalopathy. J. Med. Genet. 2019, 56, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.D.; Raymond, D.A.; Peterson, R.D.; South, M.A.; Good, R.A. The functions of the thymus system and the bursa system in the chicken. J. Exp. Med. 1966, 123, 75–102. [Google Scholar] [CrossRef] [Green Version]
- Criscitiello, M.F.; Ohta, Y.; Saltis, M.; McKinney, E.C.; Flajnik, M.F. Evolutionarily conserved TCR binding sites, identification of T cells in primary lymphoid tissues, and surprising trans-rearrangements in nurse shark. J. Immunol. 2010, 184, 6950–6960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, T.L.; Stewart, R.; Cohen, N.; Rau, L.; Ritchie, P.; Watson, M.D.; Robert, J.; Horton, J.D. Ontogeny of Xenopus NK cells in the absence of MHC class I antigens. Dev. Comp. Immunol. 2003, 27, 715–726. [Google Scholar] [CrossRef]
- Rollins-Smith, L.A.; Needham, D.A.; Davis, A.T.; Blair, P.J. Late thymectomy in Xenopus tadpoles reveals a population of T cells that persists through metamorphosis. Dev. Comp. Immunol. 1996, 20, 165–174. [Google Scholar] [CrossRef]
- Sakuraoka, J.; Tochinai, S. Demonstration of cells involved in rejection of tolerogenic grafts in tolerant Xenopus. Dev. Comp. Immunol. 1993, 17, 439–447. [Google Scholar] [CrossRef]
- Ono, M.; Tochinai, S. Demonstration of cells possessing tolerance-inducing activity in Xenopus laevis rendered tolerant perimetamorphically. Transplantation 1995, 60, 66–70. [Google Scholar] [CrossRef]
- Mashoof, S.; Breaux, B.; Criscitiello, M.F. Larval Thymectomy of Xenopus laevis. Cold Spring Harb. Protoc. 2018, 2018, pdb.prot099192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foulkrod, A.M.; Appasamy, P.M. Expression of TCR genes in adult and larval Xenopus laevis. Dev. Comp. Immunol. 2019, 96, 78–82. [Google Scholar] [CrossRef]
- McGuire, C.C.; Lawrence, B.P.; Robert, J. Thyroid Disrupting Chemicals in Mixture Perturb Thymocyte Differentiation in Xenopus laevis Tadpoles. Toxicol. Sci. Off. J. Soc. Toxicol. 2021, 181, 262–272. [Google Scholar] [CrossRef]
- Dimitrakopoulou, D.; Khwatenge, C.N.; James-Zorn, C.; Paiola, M.; Bellin, E.W.; Tian, Y.; Sundararaj, N.; Polak, E.J.; Grayfer, L.; Barnard, D.; et al. Advances in the Xenopus immunome: Diversification, expansion, and contraction. Dev. Comp. Immunol. 2023, 145, 104734. [Google Scholar] [CrossRef] [PubMed]
- Edholm, E.S.; Albertorio Saez, L.M.; Gill, A.L.; Gill, S.R.; Grayfer, L.; Haynes, N.; Myers, J.R.; Robert, J. Nonclassical MHC class I-dependent invariant T cells are evolutionarily conserved and prominent from early development in amphibians. Proc. Natl. Acad. Sci. USA 2013, 110, 14342–14347. [Google Scholar] [CrossRef] [Green Version]
- Rhoo, K.H.; Edholm, E.S.; Forzán, M.J.; Khan, A.; Waddle, A.W.; Pavelka, M.S., Jr.; Robert, J. Distinct Host-Mycobacterial Pathogen Interactions between Resistant Adult and Tolerant Tadpole Life Stages of Xenopus laevis. J. Immunol. 2019, 203, 2679–2688. [Google Scholar] [CrossRef] [Green Version]
- Rollins-Smith, L.A.; Fites, J.S.; Reinert, L.K.; Shiakolas, A.R.; Umile, T.P.; Minbiole, K.P. Immunomodulatory metabolites released by the frog-killing fungus Batrachochytrium dendrobatidis. Infect. Immun. 2015, 83, 4565–4570. [Google Scholar] [CrossRef] [Green Version]
- Hyoe, R.K.; Robert, J. A Xenopus tadpole alternative model to study innate-like T cell-mediated anti-mycobacterial immunity. Dev. Comp. Immunol. 2019, 92, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Banach, M.; Robert, J. Evolutionary Underpinnings of Innate-Like T Cell Interactions with Cancer. Immunol. Investig. 2019, 48, 737–758. [Google Scholar] [CrossRef] [PubMed]
- Yaparla, A.; Koubourli, D.V.; Popovic, M.; Grayfer, L. Exploring the relationships between amphibian (Xenopus laevis) myeloid cell subsets. Dev. Comp. Immunol. 2020, 113, 103798. [Google Scholar] [CrossRef] [PubMed]
- Yaparla, A.; Wendel, E.S.; Grayfer, L. The unique myelopoiesis strategy of the amphibian Xenopus laevis. Dev. Comp. Immunol. 2016, 63, 136–143. [Google Scholar] [CrossRef]
- Hassnain Waqas, S.F.; Noble, A.; Hoang, A.C.; Ampem, G.; Popp, M.; Strauß, S.; Guille, M.; Röszer, T. Adipose tissue macrophages develop from bone marrow-independent progenitors in Xenopus laevis and mouse. J. Leukoc. Biol. 2017, 102, 845–855. [Google Scholar] [CrossRef] [Green Version]
- Paredes, R.; Ishibashi, S.; Borrill, R.; Robert, J.; Amaya, E. Xenopus: An in vivo model for imaging the inflammatory response following injury and bacterial infection. Dev. Biol. 2015, 408, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Hossainey, M.R.H.; Yaparla, A.; Hauser, K.A.; Moore, T.E.; Grayfer, L. The Roles of Amphibian (Xenopus laevis) Macrophages during Chronic Frog Virus 3 Infections. Viruses 2021, 13, 2299. [Google Scholar] [CrossRef] [PubMed]
- Kalia, N.; Hauser, K.A.; Burton, S.; Hossainey, M.R.H.; Zelle, M.; Horb, M.E.; Grayfer, L. Endogenous Retroviruses Augment Amphibian (Xenopus laevis) Tadpole Antiviral Protection. J. Virol. 2022, 96, e0063422. [Google Scholar] [CrossRef] [PubMed]
- Nagata, S. Xenopus laevis macrophage-like cells produce XCL-1, an intelectin family serum lectin that recognizes bacteria. Immunol. Cell Biol. 2018, 96, 872–878. [Google Scholar] [CrossRef]
- Koubourli, D.V.; Yaparla, A.; Popovic, M.; Grayfer, L. Amphibian (Xenopus laevis) Interleukin-8 (CXCL8): A Perspective on the Evolutionary Divergence of Granulocyte Chemotaxis. Front. Immunol. 2018, 9, 2058. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carotenuto, R.; Pallotta, M.M.; Tussellino, M.; Fogliano, C. Xenopus laevis (Daudin, 1802) as a Model Organism for Bioscience: A Historic Review and Perspective. Biology 2023, 12, 890. https://doi.org/10.3390/biology12060890
Carotenuto R, Pallotta MM, Tussellino M, Fogliano C. Xenopus laevis (Daudin, 1802) as a Model Organism for Bioscience: A Historic Review and Perspective. Biology. 2023; 12(6):890. https://doi.org/10.3390/biology12060890
Chicago/Turabian StyleCarotenuto, Rosa, Maria Michela Pallotta, Margherita Tussellino, and Chiara Fogliano. 2023. "Xenopus laevis (Daudin, 1802) as a Model Organism for Bioscience: A Historic Review and Perspective" Biology 12, no. 6: 890. https://doi.org/10.3390/biology12060890