
Alleviative Effect of Probiotic Ferment on Lawsonia intracellularis Infection in Piglets
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Treatments
2.2. Growth Performance, Gross Pathology, and Sample Collection
2.3. Digestive Enzyme Activities and Permeability
2.4. Microscopic Pathology and Intestinal Morphology
2.5. Periodic Acid Schiff Staining and Immunohistochemistry
2.6. RNA Chromogenic In Situ Hybridization
2.7. Measurement of Gene Expression by Real-Time Quantitative PCR (qRT-PCR)
2.8. Western Blot
2.9. Ileal and Colonic Luminal Microbiome Analysis
2.10. Statistical Analysis
3. Results
3.1. Piglets Infected with LI Showed Lower Growth Performance and Typical Pathological Symptoms
3.2. Intestinal Morphological Damage Induced by LI Could be Repaired by FAM
3.3. FAM Promoted Nutrient Utilization by Enhancing Digestive Enzyme Activity and Transporter Expression
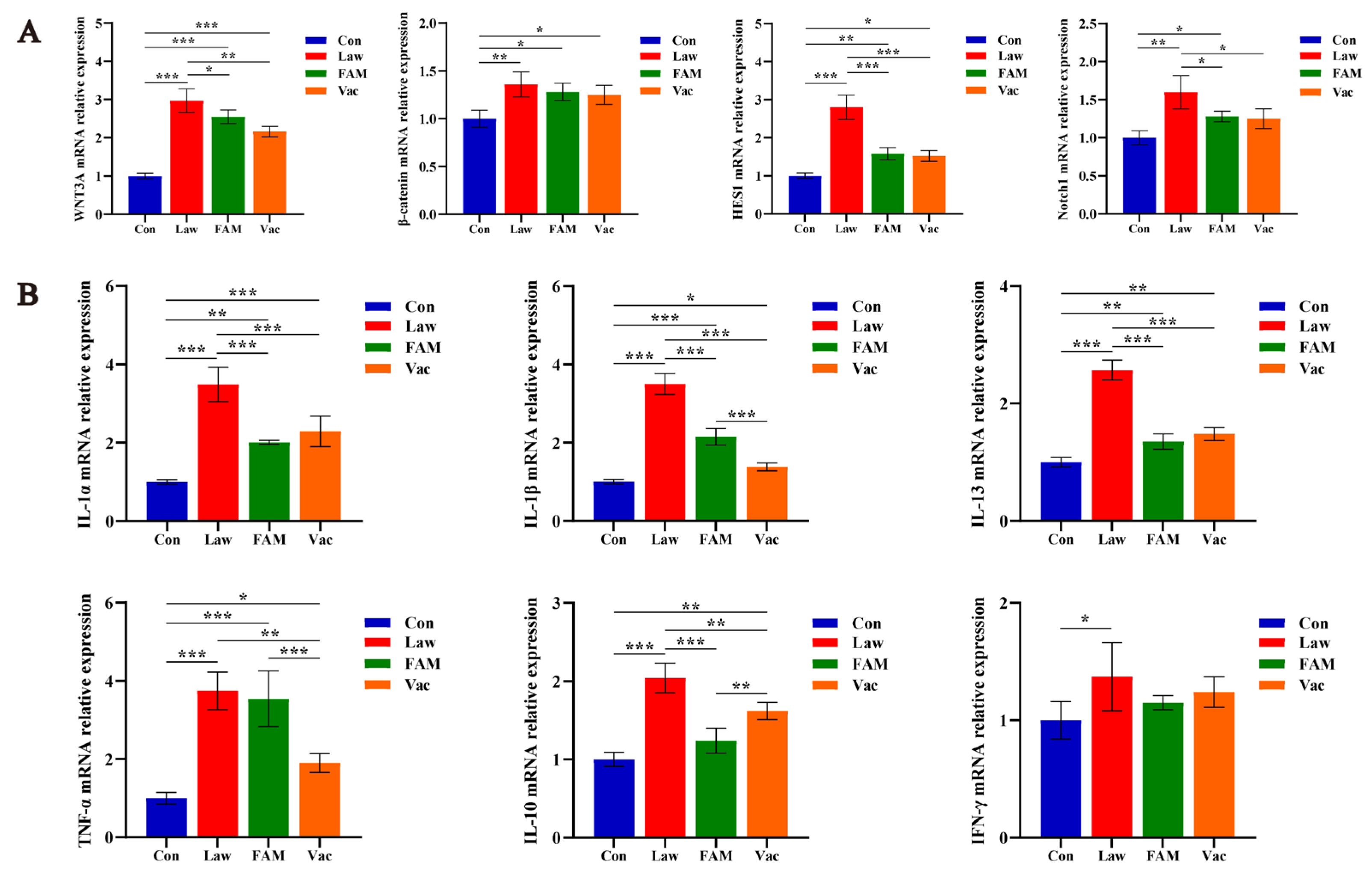
3.4. Reduction of LI Colonization Ameliorated Abnormal Differentiation and Function of Intestinal Epithelial Cells and Alleviated Severe Inflammatory Response in Piglets
3.5. Structural and Functional Changes in the Ileal Microbiota of Infected Piglets Following FAM Supplementation
3.6. FAM Altered Diversity and Structure of Gut Microbiota in the Colon of Infected Piglets
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Content, % | Nutrient Composition 2 | Content |
|---|---|---|---|
| Corn | 63 | Digestible energy, kcal·kg−1 | 3500 |
| Soybean meal | 18 | Crude protein, % | 18.8 |
| Puffed soybeans | 8 | Calcium, % | 0.75 |
| Fish meal | 4 | Available phosphorus, % | 0.4 |
| Whey powder | 3 | Lysine, % | 1.4 |
| Dicalcium phosphate | 1 | Threonine, % | 0.87 |
| Limestone | 0.77 | Methionine, % | 0.42 |
| Salt | 0.4 | Tryptophan, % | 0.21 |
| L-Lysine | 0.5 | ||
| Threonine | 0.2 | ||
| DL-methionine | 0.1 | ||
| Tryptophan | 0.03 | ||
| Premix 1 | 1 | ||
| Total | 100 |
Appendix B
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | GenBank Number |
|---|---|---|---|
| GAPDH | CCAAGGAGAACTCAAGATTTGGC | TGCCGTGGGTGGAATCATAC | XM_021091114.1 |
| IL-1β | ATGCCAACGTGCAGTCTATG | CATGCAGAACACCACTTCTCT | NM_214055.1 |
| TNF-α | GGCCCAAGGACTCAGATCAT | GGCATACCCACTCTGCCATT | NM_214022.1 |
| IL-1α | AACCTGGATGAGGCAGTGAAA | AGCACTCACAAACAGTCGGG | NM_214029.1 |
| IL-10 | GAAGACGTAATGCCGAAGGC | AGGGCAGAAATTGATGACAGC | NM_214041.1 |
| IL-13 | TTGCTCTCACCTGCTTTGGT | GCATAGGGGTGTCTTCTGGT | XM_005661643.3 |
| IFN-γ | GTTTTTCTGGCTCTTACTGC | CTTCCGCTTTCTTAGGTTAG | NM_213948.1 |
| HES1 | AACACGACACCGGATAAACCA | GGAATGCCTCGAGCTATCTTTT | NM_001195231.1 |
| WNT3A | GCTCTCGGGGCGGACT | CAACAGCCAGGGACCACC | XM_003123621.4 |
| Notch1 | CCCGAGTGGACAGGTCAGT | GTCGTCGATGTTTTCGCTGC | XM_021081037.1 |
| β-catenin | GGCTTGGAATGAGACTGCTG | GGGTCCATACCCAAGGCATC | XM_013981492.2 |
| SGLT1 | CGTCATCTACTTCGTGGTGGT | AGAAGCTCCAACCGGCCA | NM_001164021.1 |
| TCN2 | TGCCTGGACCTGCGAAATAG | TTGAGGCTGTGCAGGTAGTG | XM_021071885.1 |
| ASBT | TGCCGCCCTCCCCACAAC | GGACTGACCCTGGACTCTGACC | NM_000452.3 |
| CAT1 | TCACGCTCATGATGCCCTAC | TGGAGCCTAAAAGGCTGGTG | NM_001012613.1 |
| EAAC1 | AACCCTTTCCGATTCGCCAT | GCTGTGGCGGTGATACTGAT | NM_001164649.1 |
| FABPS | AGTTGACCATCACTACCGGG | AACTGAACCACTGTCTTGACC | NM_001004046.2 |
Appendix C
| ITEM | Con | Law | FAM | Vac | p Value |
|---|---|---|---|---|---|
| d −28~d 0 | |||||
| Initial BW (kg) | 9.15 ± 0.44 | 9.12 ± 0.39 | 9.14 ± 0.47 | 9.06 ± 0.34 | 0.872 |
| Final BW(kg) | 20.61 ± 2.17 | 20.41 ± 2.63 | 21.20 ± 1.85 | 20.33 ± 2.58 | 0.534 |
| ADG (g/d) | 409.3 ± 13.5 b | 403.2 ± 8.7 b | 430.7 ± 15.2 a | 402.5 ± 14.4 b | 0.040 |
| ADFI (g/d) | 646.4 | 619.2 | 648.1 | 623.9 | |
| F/G | 1.58 | 1.54 | 1.5 | 1.55 | |
| d 0~d 10 | |||||
| Initial BW (kg) | 20.61 ± 2.17 | 20.41 ± 2.63 | 21.2 ± 1.85 | 20.33 ± 2.58 | 0.477 |
| Final BW (kg) | 26.48 ± 2.33 a | 21.68 ± 3.39 b | 23.26 ± 2.94 b | 22.57 ± 2.85 b | 0.000 |
| ADG (g/d) | 586.6 ± 14.9 a | 127.5 ± 7.9 d | 205.7± 12.6 c | 224.4 ± 14.7 b | 0.000 |
| ADFI (g/d) | 1020.7 | 698.3 | 791.6 | 823.5 | |
| F/G | 1.74 | 5.48 | 3.85 | 3.67 | |
| Diarrhea incidence, % | 3.3 | 58.3 | 43.3 | 31.7 |
References
- Kroll, J.J.; Roof, M.B.; Hoffman, L.J.; Dickson, J.S.; Harris, D.L.H. Proliferative enteropathy: A global enteric disease of pigs caused by Lawsonia intracellularis. Anim. Health Res. Rev. 2005, 6, 173–197. [Google Scholar] [CrossRef] [Green Version]
- Guedes, R.M.C.; Gebhart, C.J. Onset and duration of fecal shedding, cell-mediated and humoral immune responses in pigs after challenge with a pathogenic isolate or attenuated vaccine strain of Lawsonia intracellularis. Vet. Microbiol. 2003, 91, 135–145. [Google Scholar] [CrossRef]
- Komine, M.; Cunha, T.O.; Mullaney, T.P.; Smedley, R.C.; Langohr, I.M. Pathology in Practice. JAVMA-J. Am. Vet. Med. Assoc. 2016, 248, 897–899. [Google Scholar] [CrossRef]
- Lawson, G.H.K.; Gebhart, C.J. Proliferative enteropathy. J. Comp. Pathol. 2000, 22, 77–100. [Google Scholar] [CrossRef] [PubMed]
- Gabardo, M.d.P.; Sato, J.P.H.; Daniel, A.G.d.S.; Andrade, M.R.; Pereira, C.E.R.; Rezende, T.P.; Otoni, L.V.A.; Rezende, L.A.; Guedes, R.M.C. Evaluation of the involvement of mice (Mus musculus) in the epidemiology of porcine proliferative enteropathy. Vet. Microbiol. 2017, 205, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Crienen, A.; Swam, H.; von Berg, S.; Jolie, R.; Nathues, H. Prevalence of Lawsonia intracellularis in pig herds in different European countries. Porcine Health Manag. 2019, 5, 31. [Google Scholar] [CrossRef]
- Vannucci, F.A.; Gebhart, C.J. Recent Advances in Understanding the Pathogenesis of Lawsonia intracellularis Infections. Vet. Pathol. 2014, 51, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.G.E.; Lawson, G.H.K. Lawsonia intracellularis: Getting inside the pathogenesis of proliferative enteropathy. Vet. Microbiol. 2001, 82, 331–345. [Google Scholar] [CrossRef]
- Vannucci, F.A.; Foster, D.N.; Gebhart, C.J. Comparative Transcriptional Analysis of Homologous Pathogenic and Non-Pathogenic Lawsonia intracellularis Isolates in Infected Porcine Cells. PLoS ONE. 2012, 7, e46708. [Google Scholar] [CrossRef] [Green Version]
- Leite, F.L.; Abrahante, J.E.; Vasquez, E.; Vannucci, F.; Gebhart, C.J.; Winkelman, N.; Mueller, A.; Torrison, J.; Rambo, Z.; Isaacson, R.E. A cell proliferation and inflammatory signature is induced by Lawsonia intracellularis infection in swine. Mbio 2019, 10, e01605-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huan, Y.W.; Bengtsson, R.J.; MacIntyre, N.; Guthrie, J.; Finlayson, H.; Smith, S.H.; Archibald, A.L.; Ait-Ali, T. Lawsonia intracellularis exploits beta-catenin/Wnt and Notch signalling pathways during infection of intestinal crypt to alter cell homeostasis and promote cell proliferation. PLoS ONE. 2017, 12, e173782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karuppannan, A.K.; Opriessnig, T. Lawsonia intracellularis: Revisiting the Disease Ecology and Control of This Fastidious Pathogen in Pigs. Front. Vet. Sci. 2018, 5, 181. [Google Scholar] [CrossRef] [PubMed]
- Walter, D.; Gebhart, C.; Kroll, J.; Holck, J.T.; Chittick, W. Serologic profiling and vaccination timing for Lawsonia intracellularis. J Swine Health Prod. 2004, 12, 310–313. [Google Scholar]
- Nogueira, M.G.; Collins, A.M.; Dunlop, R.H.; Emery, D. Effect of the route of administration on the mucosal and systemic immune responses to Lawsonia intracellularis vaccine in pigs. Aust. Vet. J. 2015, 93, 124–126. [Google Scholar] [CrossRef] [PubMed]
- Roerink, F.; Morgan, C.L.; Knetter, S.M.; Passat, M.H.; Archibald, A.L.; Ait-Ali, T.; Strait, E.L. A novel inactivated vaccine against Lawsonia intracellularis induces rapid induction of humoral immunity, reduction of bacterial shedding and provides robust gut barrier function. Vaccine 2018, 36, 1500–1508. [Google Scholar] [CrossRef]
- Pereira, C.E.R.; Resende, T.P.; Vasquez, E.; Marshall-Lund, L.; Guedes, R.M.C.; Gebhart, C.J. In vitro antimicrobial activity against equine Lawsonia intracellularis strains. Equine. Vet. J. 2019, 51, 665–668. [Google Scholar] [CrossRef]
- Collins, A.M.; Love, R.J. Re-challenge of pigs following recovery from proliferative enteropathy. Vet. Microbiol. 2007, 120, 381–386. [Google Scholar] [CrossRef]
- Whitney, M.H.; Shurson, G.C.; Guedes, R.C. Effect of dietary inclusion of distillers dried grains with solubles, soybean hulls, or a polyclonal antibody product on the ability of growing pigs to resist a Lawsonia intracellularis challenge. J. Anim. Sci. 2006, 84, 1880–1889. [Google Scholar] [CrossRef] [Green Version]
- Le Bourgot, C.; Le Normand, L.; Formal, M.; Respondek, F.; Blat, S.; Apper, E.; Ferret-Bernard, S.; Le Huerou-Luron, I. Maternal short-chain fructo-oligosaccharide supplementation increases intestinal cytokine secretion, goblet cell number, butyrate concentration and Lawsonia intracellularis humoral vaccine response in weaned pigs. Br. J. Nutr. 2017, 117, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xie, Q.; Sun, S.; Huang, B.; Zhang, Y.; Xu, Y.; Zhang, S.; Xiang, H. Probiotics-fermented Massa Medicata Fermentata ameliorates weaning stress in piglets related to improving intestinal homeostasis. Appl. Microbiol. Biotechnol. 2018, 102, 10713–10727. [Google Scholar] [CrossRef]
- Park, J.H.; Song, T.H.; Kim, I. Egg production, egg quality, and cecal microbial populations of layers fed diets supplemented with fermented phytogenic feed additive. Turk. Vet. Anim. Sci. 2016, 40, 660–666. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Swine, 11th ed.; National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Xu, T.; Ma, X.; Zhou, X.; Qian, M.; Yang, Z.; Cao, P.; Han, X. Coated tannin supplementation improves growth performance, nutrients digestibility, and intestinal function in weaned piglets. J. Anim. Sci. 2022, 100, skac088. [Google Scholar] [CrossRef]
- Hu, L.S.; Geng, S.J.; Li, Y.; Cheng, S.S.; Fu, X.F.; Yue, X.J.; Han, X.Y. Exogenous Fecal Microbiota Transplantation from Local Adult Pigs to Crossbred Newborn Piglets. Front. Microbiol. 2018, 8, 2663. [Google Scholar] [CrossRef]
- D’Annunzio, G.; Bardini, R.; Ostanello, F.; Sarli, G. Porcine Proliferative Enteropathy Caused by Lawsonia intracellularis. Large Anim. Rev. 2021, 27, 149–163. [Google Scholar]
- Xie, Z.; Li, M.; Qian, M.; Yang, Z.; Han, X. Co-Cultures of Lactobacillus acidophilus and Bacillus subtilis Enhance Mucosal Barrier by Modulating Gut Microbiota-Derived Short-Chain Fatty Acids. Nutrients. 2022, 14, 4475. [Google Scholar] [CrossRef]
- Chance, J.A.; DeRouchey, J.M.; Amachawadi, R.G.; Ishengoma, V.; Nagaraja, T.G.; Goodband, R.D.; Woodworth, J.C.; Tokach, M.D.; Kang, Q.; Loughmiller, J.A.; et al. Influence of yeast-based pre- and probiotics in lactation and nursery diets on nursery pig performance and antimicrobial resistance of fecal Escherichia coli. J. Anim. Sci. 2022, 100, skac166. [Google Scholar] [CrossRef]
- Helm, E.T.; Curry, S.M.; Schwartz, K.J.; Lonergan, S.M.; Gabler, N.K. Mycoplasma hyopneumoniae-Lawsonia intracellularis dual challenge modulates intestinal integrity and function. J. Anim. Sci. 2019, 97, 2376–2384. [Google Scholar] [CrossRef] [PubMed]
- Draskovic, V.; Stanimirovic, Z.; Glisic, M.; Bosnjak-Neumuller, J.; Teodorovic, R.; Teodorovic, V.; Kukolj, V. The effects of a phytogenic additive on the histomorphometric characteristics of the intestines in weaned pigs with a subclinical natural infection with Lawsonia intracellularis. Acta Vet. Beogr. 2020, 70, 81–91. [Google Scholar] [CrossRef]
- Chwen, L.T.; Foo, H.L.; Nguyen Tien, T.; Choe, D.W. Growth Performance, Plasma Fatty Acids, Villous Height and Crypt Depth of Preweaning Piglets Fed with Medium Chain Triacylglycerol. Asian Australas. J. Anim. Sci. 2013, 26, 700–704. [Google Scholar]
- Lhoste, E.F.; Fiszlewicz, M.; Gueugneau, A.M.; Wickerplanquart, C.; Puigserver, A.; Corring, T. Effects of dietary proteins on some pancreatic messenger-rnas encoding digestive enzymes in the pig. J. Nutr. Biochem. 1993, 4, 143–152. [Google Scholar] [CrossRef]
- Helm, E.T.; Burrough, E.R.; Leite, F.L.; Gabler, N.K. Lawsonia intracellularis infected enterocytes lack sucrase-isomaltase which contributes to reduced pig digestive capacity. Vet Res. 2021, 52, 90–106. [Google Scholar] [CrossRef] [PubMed]
- Jiao, N.; Baker, S.S.; Chapa-Rodriguez, A.; Liu, W.; Nugent, C.A.; Tsompana, M.; Mastrandrea, L.; Buck, M.J.; Baker, R.D.; Genco, R.J.; et al. Suppressed hepatic bile acid signalling despite elevated production of primary and secondary bile acids in NAFLD. Gut. 2018, 67, 1881–1891. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.; Robertson, J.; Gallagher, N. Humoral regulation of vitamin-B12 absorption by pregnant mouse small-intestine. Gastroenterology. 1977, 72, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Vannucci, F.A.; Foster, D.N.; Gebhart, C.J. Laser microdissection coupled with RNA-seq analysis of porcine enterocytes infected with an obligate intracellular pathogen (Lawsonia intracellularis). BMC Genom. 2013, 14, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollegioni, L.; Piubelli, L.; Sacchi, S.; Pilone, M.S.; Molla, G. Physiological functions of D-amino acid oxidases: From yeast to humans. Cell Mol. Life. Sci. 2007, 64, 1373–1394. [Google Scholar] [CrossRef]
- Encarnacao, P.; de Lange, C.; Bureau, D.P. Diet energy source affects lysine utilization for protein deposition in rainbow trout (Oncorhynchus mykiss). Aquaculture 2006, 261, 1371–1381. [Google Scholar] [CrossRef]
- Smith, S.H.; Wilson, A.D.; Van Ettinger, I.; MacIntyre, N.; Archibald, A.L.; Ait-Ali, T. Down-regulation of mechanisms involved in cell transport and maintenance of mucosal integrity in pigs infected with Lawsonia intracellularis. Vet Res. 2014, 45, 55. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.Y.; Jung, J.H.; Kim, I.H. Effect of mannan oligosaccharides and fructan on growth performance, nutrient digestibility, blood profile, and diarrhea score in weanling pigs. J. Anim. Sci. 2012, 90, 833–839. [Google Scholar] [CrossRef]
- Zhao, P.; Li, H.; Lei, Y.; Li, T.; Kim, S.; Kim, I. Effect of fermented medicinal plants on growth performance, nutrient digestibility, fecal noxious gas emissions, and diarrhea score in weanling pigs. J. Sci. Food Agric. 2016, 96, 1269–1274. [Google Scholar] [CrossRef]
- Draskovic, V.; Bosnjak-Neumuller, J.; Vasiljevic, M.; Petrujkic, B.; Aleksic, N.; Kukolj, V.; Stanimirovic, Z. Influence of phytogenic feed additive on Lawsonia intracellularis infection in pigs. Prev. Vet. Med. 2018, 151, 46–51. [Google Scholar] [CrossRef]
- Opriessnig, T.; Karuppannan, A.K.; Beckler, D.; Ait-Ali, T.; Cubas-Atienzar, A.; Halbur, P.G. Bacillus pumilus probiotic feed supplementation mitigates Lawsonia intracellularis shedding and lesions. Vet. Res. 2019, 50, 8. [Google Scholar] [CrossRef] [Green Version]
- Bansil, R.; Turner, B.S. Mucin structure, aggregation, physiological functions and biomedical applications. Curr. Opin. Colloid Interface Sci. 2006, 11, 164–170. [Google Scholar] [CrossRef]
- Kim, J.J.; Khan, W.I. Goblet cells and mucins: Role in innate defense in enteric infections. Pathogens 2013, 2, 55–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengtsson, R.J.; MacIntyre, N.; Guthrie, J.; Wilson, A.D.; Finlayson, H.; Matika, O.; Pong-Wong, R.; Smith, S.H.; Archibald, A.L.; Ait-Ali, T. Lawsonia intracellularis infection of intestinal crypt cells is associated with specific depletion of secreted MUC2 in goblet cells. Vet. Immunol. Immunopathol. 2015, 168, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Takiishi, T.; Morales Fenero, C.I.; Saraiva Camara, N.O. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef] [PubMed]
- Yi, Z.; Fan, H.; Liu, X.; Tang, Q.; Zuo, D.; Yang, J. Adrenomedullin improves intestinal epithelial barrier function by downregulating myosin light chain phosphorylation in ulcerative colitis rats. Mol. Med. Rep. 2015, 12, 3615–3620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Rando, T.A. Manifestations and mechanisms of stem cell aging. J. Cell Biol. 2011, 193, 257–266. [Google Scholar] [CrossRef] [PubMed]
- de Lau, W.; Barker, N.; Low, T.Y.; Koo, B.-K.; Li, V.S.W.; Teunissen, H.; Kujala, P.; Haegebarth, A.; Peters, P.J.; van de Wetering, M.; et al. Lgr5 homologues associate with Wnt receptors and mediate R-spondin signalling. Nature. 2011, 476, 293-U257. [Google Scholar] [CrossRef]
- van Es, J.H.; van Gijn, M.E.; Riccio, O.; van den Born, M.; Vooijs, M.; Begthel, H.; Cozijnsen, M.; Robine, S.; Winton, D.J.; Radtke, F.; et al. Notch/gamma-secretase inhibition turns proliferative cells in intestinal crypts and adenomas into goblet cells. Nature 2005, 435, 959–963. [Google Scholar] [CrossRef]
- Fre, S.; Huyghe, M.; Mourikis, P.; Robine, S.; Louvard, D.; Artavanis-Tsakonas, S. Notch signals control the fate of immature progenitor cells in the intestine. Nature 2005, 435, 964–968. [Google Scholar] [CrossRef]
- Stanger, B.Z.; Datar, R.; Murtaugh, L.C.; Melton, D.A. Direct regulation of intestinal fate by Notch. Proc Natl. Acad. Sci. USA 2005, 102, 12443–12448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reya, T.; Clevers, H. Wnt signalling in stem cells and cancer. Nature 2005, 434, 843–850. [Google Scholar] [CrossRef]
- Pinto, D.; Gregorieff, A.; Begthel, H.; Clevers, H. Canonical Wnt signals are essential for homeostasis of the intestinal epithelium. Genes. Dev. 2003, 17, 1709–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obradovic, M.R.; Wilson, H.L. Immune response and protection against Lawsonia intracellularis infections in pigs. Vet. Immunol. Immunopathol. 2020, 219, 109959. [Google Scholar] [CrossRef]
- Yeh, J.-Y.; Ga, A.R. Systemic cytokine response in pigs infected orally with a Lawsonia intracellularis isolate of South Korean origin. J. Vet. Med. Sci. 2018, 80, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogueira, M.G.; Collins, A.M.; Donahoo, M.; Emery, D. Immunological responses to vaccination following experimental Lawsonia intracellularis virulent challenge in pigs. Vet. Microbiol. 2013, 164, 131–138. [Google Scholar] [CrossRef]
- Jianmin, W.; Ying, Z.; Xian, W.; Lingchang, K.; Johnston, L.J.; Lin, L.; Xi, M. Dietary nutrients shape gut microbes and intestinal mucosa via epigenetic modifications. Crit. Rev. Food Sci. Nutr. 2022, 62, 783–797. [Google Scholar]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.A.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; et al. The gut microbiota and host health: A new clinical frontier. Gut 2016, 65, 330–339. [Google Scholar] [CrossRef] [Green Version]
- D’Afflitto, M.; Upadhyaya, A.; Green, A.; Peiris, M. Association Between Sex Hormone Levels and Gut Microbiota Composition and Diversity-A Systematic Review. J. Clin. Gastroenterol. 2022, 56, 384–392. [Google Scholar] [CrossRef]
- Chen, Y.-r.; Jing, Q.-l.; Chen, F.-l.; Zheng, H.; Chen, L.-d.; Yang, Z.-c. Desulfovibrio is not always associated with adverse health effects in the Guangdong Gut Microbiome Project. Peerj. 2021, 9, e12033. [Google Scholar] [CrossRef]
- He, Y.; Mao, C.; Wen, H.; Chen, Z.; Lai, T.; Li, L.; Lu, W.; Wu, H. Influence of ad Libitum Feeding of Piglets with Bacillus Subtilis Fermented Liquid Feed on Gut Flora, Luminal Contents and Health. Sci Rep. 2017, 7, 44553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houtman, T.A.; Eckermann, H.A.; Smidt, H.; de Weerth, C. Gut microbiota and BMI throughout childhood: The role of firmicutes, bacteroidetes, and short-chain fatty acid producers. Sci. Rep. 2022, 12, 3140. [Google Scholar] [CrossRef]
- Touch, S.; Godefroy, E.; Rolhion, N.; Danne, C.; Oeuvray, C.; Straube, M.; Galbert, C.; Brot, L.; Alonso Salgueiro, I.; Chadi, S.; et al. Human CD4+/CD8alpha+ regulatory T cells induced by Faecalibacterium prausnitzii protect against intestinal inflammation. JCI Insight 2022, 7, e154722. [Google Scholar] [CrossRef]
- Zawistowska-Rojek, A.; Kosmider, A.; Stepien, K.; Tyski, S. Adhesion and aggregation properties of Lactobacillaceae strains as protection ways against enteropathogenic bacteria. Arch. Microbiol. 2022, 204, 285. [Google Scholar] [CrossRef]
- Rodriguez-Palacios, A.; Harding, A.; Menghini, P.; Himmelman, C.; Retuerto, M.; Nickerson, K.P.; Lam, M.; Croniger, C.M.; McLean, M.H.; Durum, S.K.; et al. The Artificial Sweetener Splenda Promotes Gut Proteobacteria, Dysbiosis, and Myeloperoxidase Reactivity in Crohn’s Disease-Like Ileitis. Inflamm. Bowel Dis. 2018, 24, 1005–1020. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, M.; Zhou, M.; Wu, L.; Yang, H.; Huang, L.; Chen, C. Isolation and genomic characterization of five novel strains of Erysipelotrichaceae from commercial pigs. BMC Microbiol. 2021, 21, 125. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Lin, Y.; Wang, Z.; Pan, Q.; Cai, G.; He, Q.; Xu, X.; Cai, X. Campylobacter jejuni Cytolethal Distending Toxin Induces GSDME-Dependent Pyroptosis in Colonic Epithelial Cells. Front. Cell. Infect. Microbiol. 2022, 12, 853204. [Google Scholar] [CrossRef]
- Henke, M.T.; Brown, E.M.; Cassilly, C.D.; Vlamakis, H.; Xavier, R.J.; Clardy, J. Capsular polysaccharide correlates with immune response to the human gut microbe Ruminococcus gnavus. Proc. Natl. Acad. Sci. USA 2021, 118, e2007595118. [Google Scholar] [CrossRef]
- Pires, R.H.; Venceslau, S.S.; Morais, F.; Teixeira, M.; Xavier, A.V.; Pereira, I.A.C. Characterization of the Desulfovibrio desulfuricans ATCC 27774 DsrMKJOP complex—A membrane-bound redox complex involved in the sulfate respiratory pathway. Biochemistry 2006, 45, 249–262. [Google Scholar] [CrossRef]
- Kushkevych, I.; Vitezova, M.; Fedrova, P.; Vochyanova, Z.; Parakova, L.; Hosek, J. Kinetic properties of growth of intestinal sulphate-reducing bacteria isolated from healthy mice and mice with ulcerative colitis. Acta. Vet. Brno. 2017, 86, 405–411. [Google Scholar] [CrossRef] [Green Version]
- Kushkevych, I.; Dordevic, D.; Kollar, P. Analysis of physiological parameters of Desulfovibrio strains from individuals with colitis. Open Life Sci. 2018, 13, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Stephens, M.; von der Weid, P.-Y. Lipopolysaccharides modulate intestinal epithelial permeability and inflammation in a species-specific manner. Gut Microbes 2020, 11, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Im, E.; Riegler, F.M.; Pothoulakis, C.; Rhee, S.H. Elevated lipopolysaccharide in the colon evokes intestinal inflammation, aggravated in immune modulator-impaired mice. Am. J. Physiol.-Gastroint. Liver Physiol. 2012, 303, G490–G497. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.; Ham, H.-I.; Lee, S.-H.; Cho, Y.-J.; Kim, Y.-R.; Yoon, C.-K.; Seok, Y.-J. Functional dissection of the phosphotransferase system provides insight into the prevalence of Faecalibacterium prausnitzii in the host intestinal environment. Environ. Microbiol. 2021, 23, 4726–4740. [Google Scholar] [CrossRef] [PubMed]










| Item | Con | Law | FAM | Vac | p Value |
|---|---|---|---|---|---|
| Jejunum | |||||
| Villus height (μm) | 523.4 ± 21.6 | 505.5 ± 22.5 | 518.4 ± 18.1 | 513.3 ± 20.4 | 0.349 |
| Crypt depth (μm) | 254.5 ± 11.0 | 278.3 ± 11.4 | 270.5 ± 12.0 | 262.4 ± 10.4 | 0.350 |
| Villus height/Crypt depth | 2.08 ± 0.11 a | 1.82 ± 0.09 b | 1.92 ± 0.09 a,b | 1.96 ± 0.07 a,b | 0.099 |
| Ileum | |||||
| Villus height (μm) | 473.3 ± 18.2 a | 407.3 ± 15.6 c | 427.2 ± 14.7 b,c | 454.1 ± 20.7 a,b | 0.007 |
| Crypt depth (μm) | 212.1 ± 9.7 c | 297.0 ± 14.0 a | 269.1 ± 13.3 b | 265.4 ± 12.4 b | 0.000 |
| Villus height/Crypt depth | 2.23 ± 0.13 a | 1.37 ± 0.03 d | 1.59 ± 0.02 c | 1.71 ± 0.08 b | 0.000 |
| Colon | |||||
| Crypt depth (μm) | 480.3 ± 16.1 c | 597.3 ± 20.9 a | 553.4 ± 18.3 b | 559.4 ± 19.5 b | 0.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, T.; Guo, Y.; Zhang, Y.; Cao, K.; Zhou, X.; Qian, M.; Han, X. Alleviative Effect of Probiotic Ferment on Lawsonia intracellularis Infection in Piglets. Biology 2023, 12, 879. https://doi.org/10.3390/biology12060879
Xu T, Guo Y, Zhang Y, Cao K, Zhou X, Qian M, Han X. Alleviative Effect of Probiotic Ferment on Lawsonia intracellularis Infection in Piglets. Biology. 2023; 12(6):879. https://doi.org/10.3390/biology12060879
Chicago/Turabian StyleXu, Tingting, Yong Guo, Yuanyuan Zhang, Kai Cao, Xinchen Zhou, Mengqi Qian, and Xinyan Han. 2023. "Alleviative Effect of Probiotic Ferment on Lawsonia intracellularis Infection in Piglets" Biology 12, no. 6: 879. https://doi.org/10.3390/biology12060879