
Characterization and Homology Modeling of Catalytically Active Recombinant PhaCAp Protein from Arthrospira platensis

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
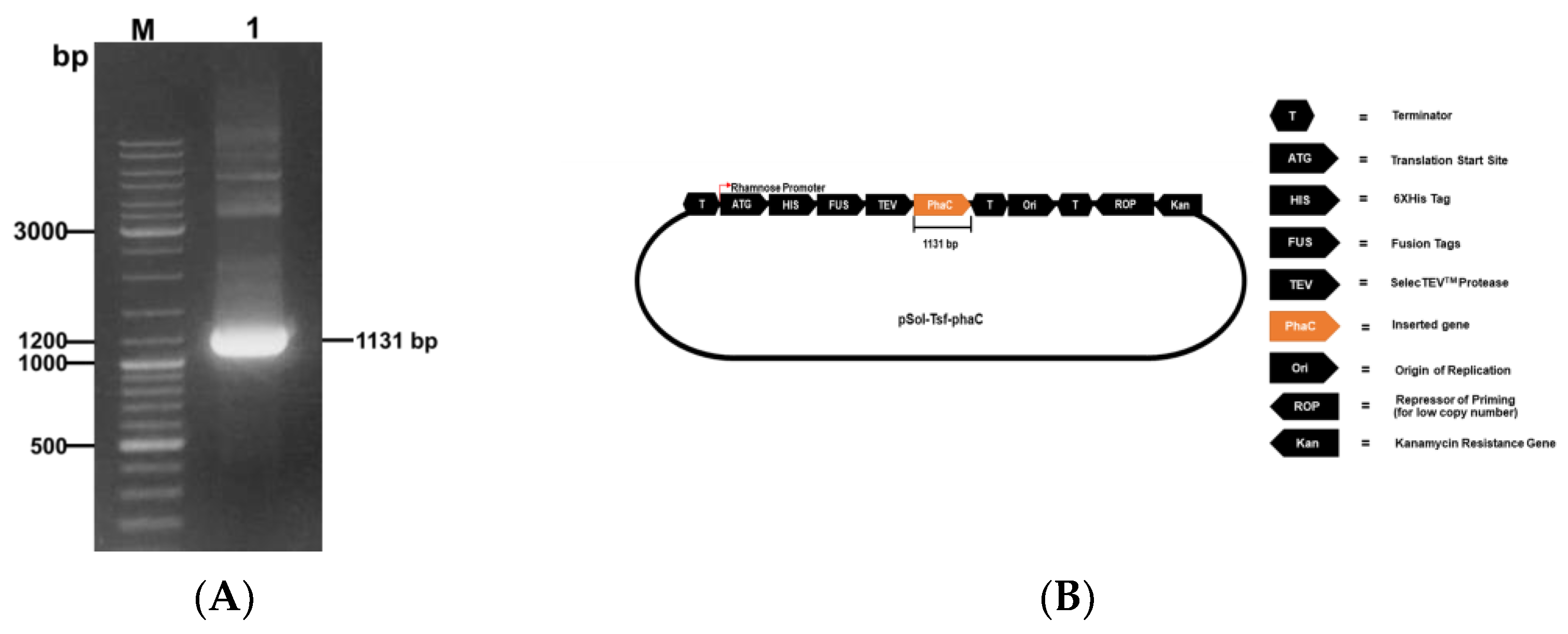
2.2. Cloning of phaCAp in pSol-Tsf Plasmid
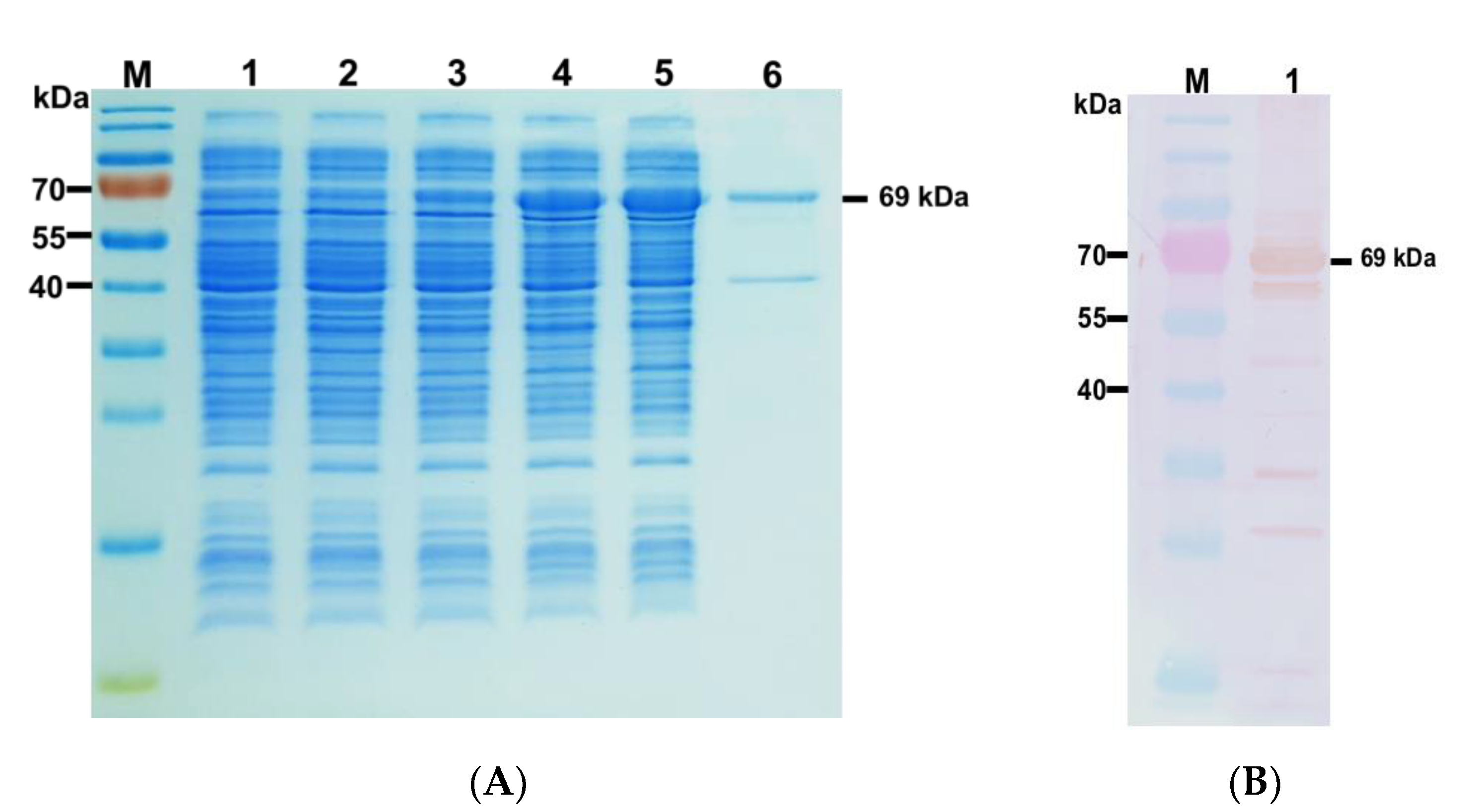
2.3. Expression and Purification of rPhaCAp
2.4. In Vitro Enzymatic Assay of the Purified rPhaCAp
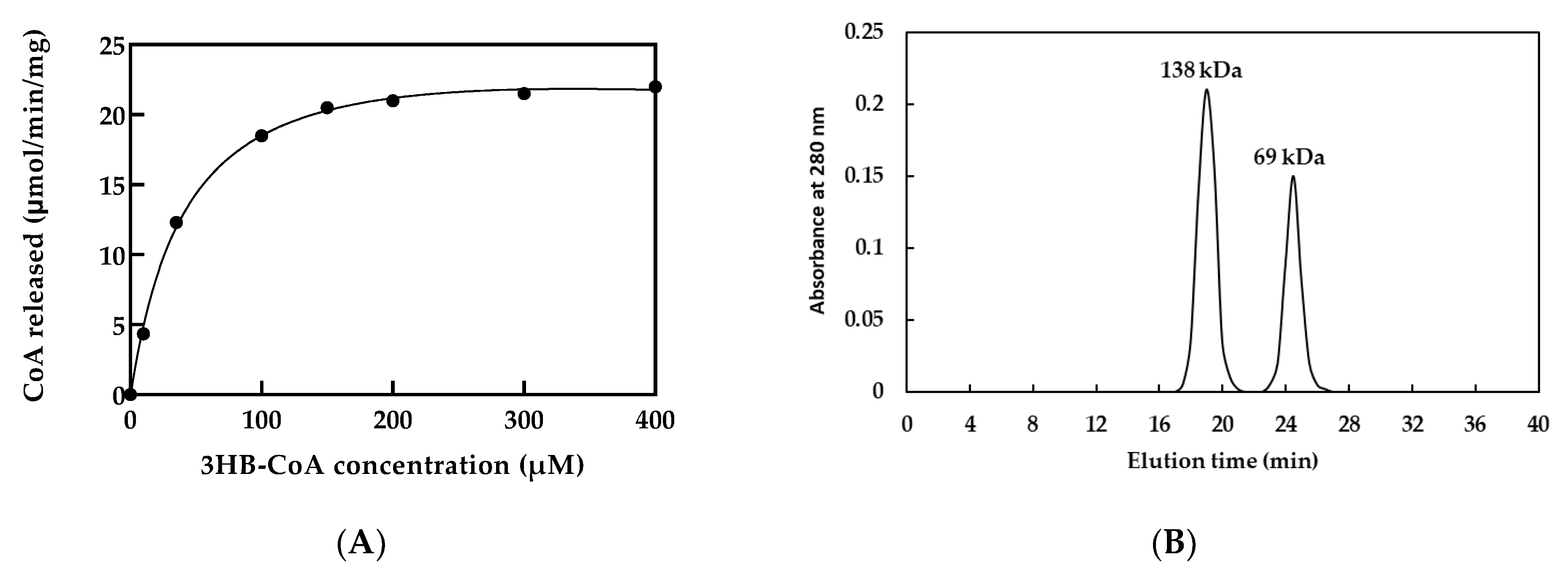
2.5. Determination of Steady-State Kinetic Constants of rPhaCAp
2.6. Production of Polyclonal Antibodies against rPhaCAp
2.7. Western Blot Analysis
2.8. Size-Exclusion Chromatography
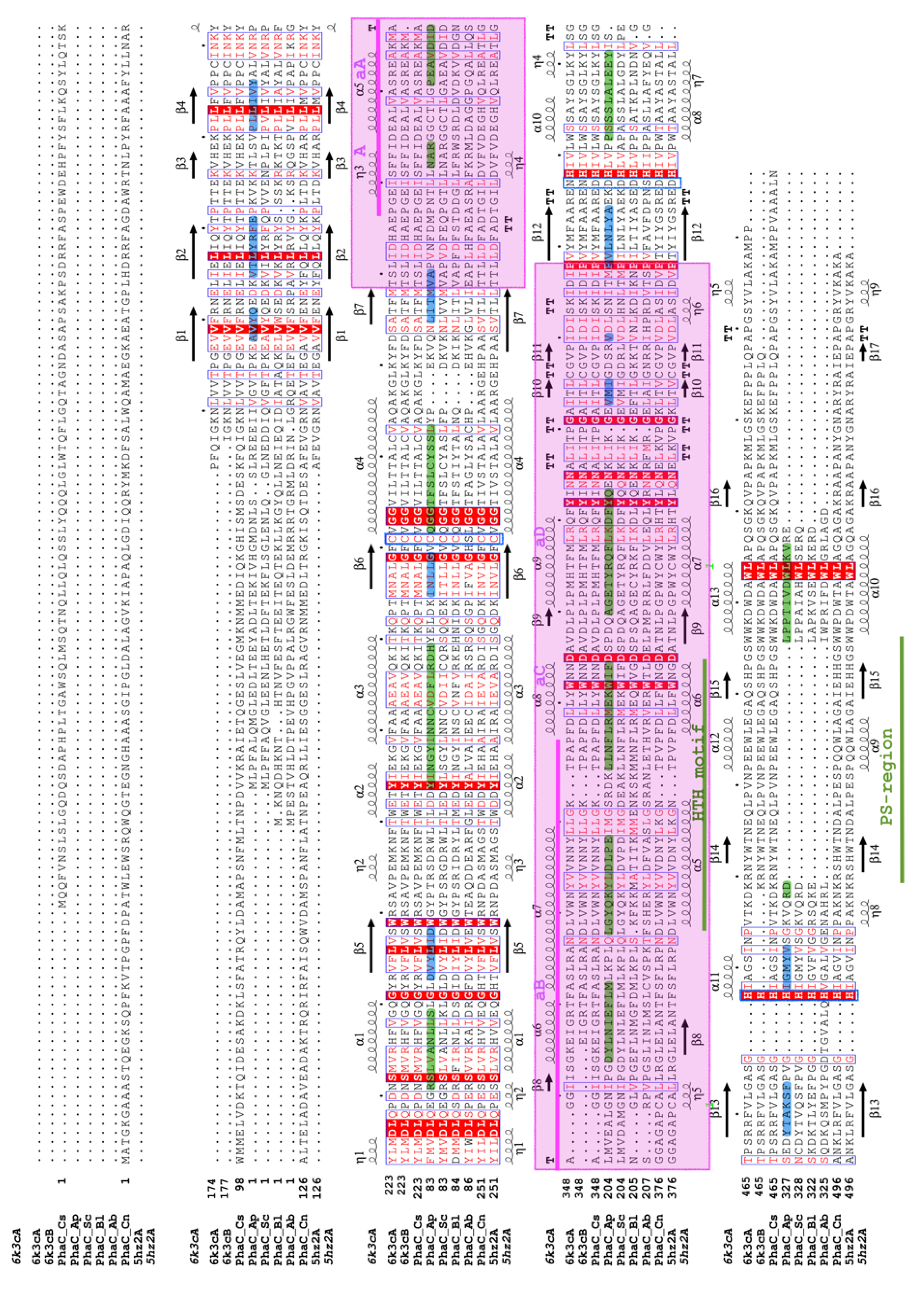
2.9. Sequence and Multiple Sequence Alignment Analysis
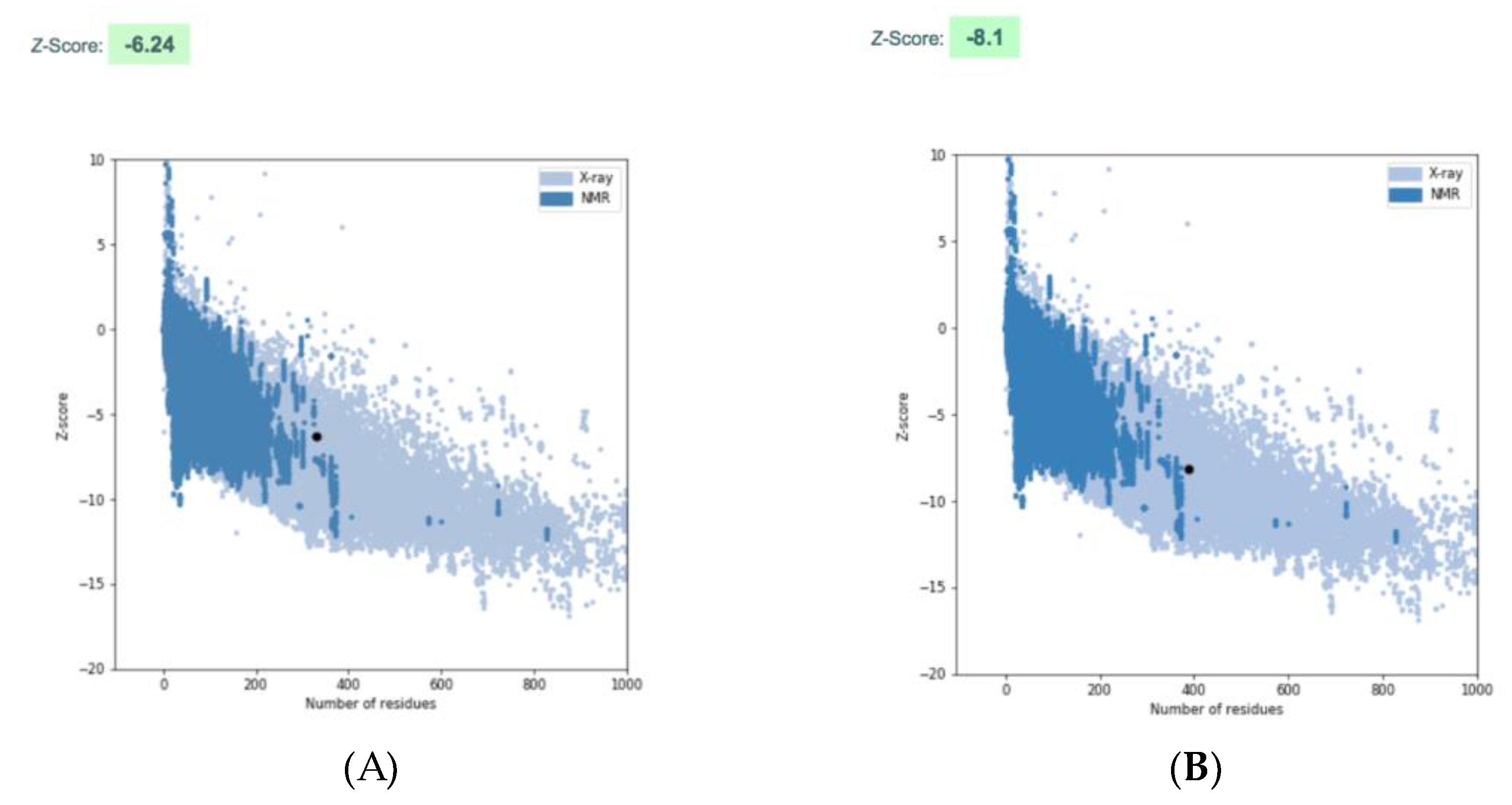
2.10. Homology Modelling
3. Results
3.1. Cloning, Expression, and Purification of rPhaCAp
3.2. PHA Synthase Activity and Kinetics
3.3. Size-Exclusion Chromatography
3.4. Sequence Analysis and Multiple Sequence Alignment for Homology Modeling
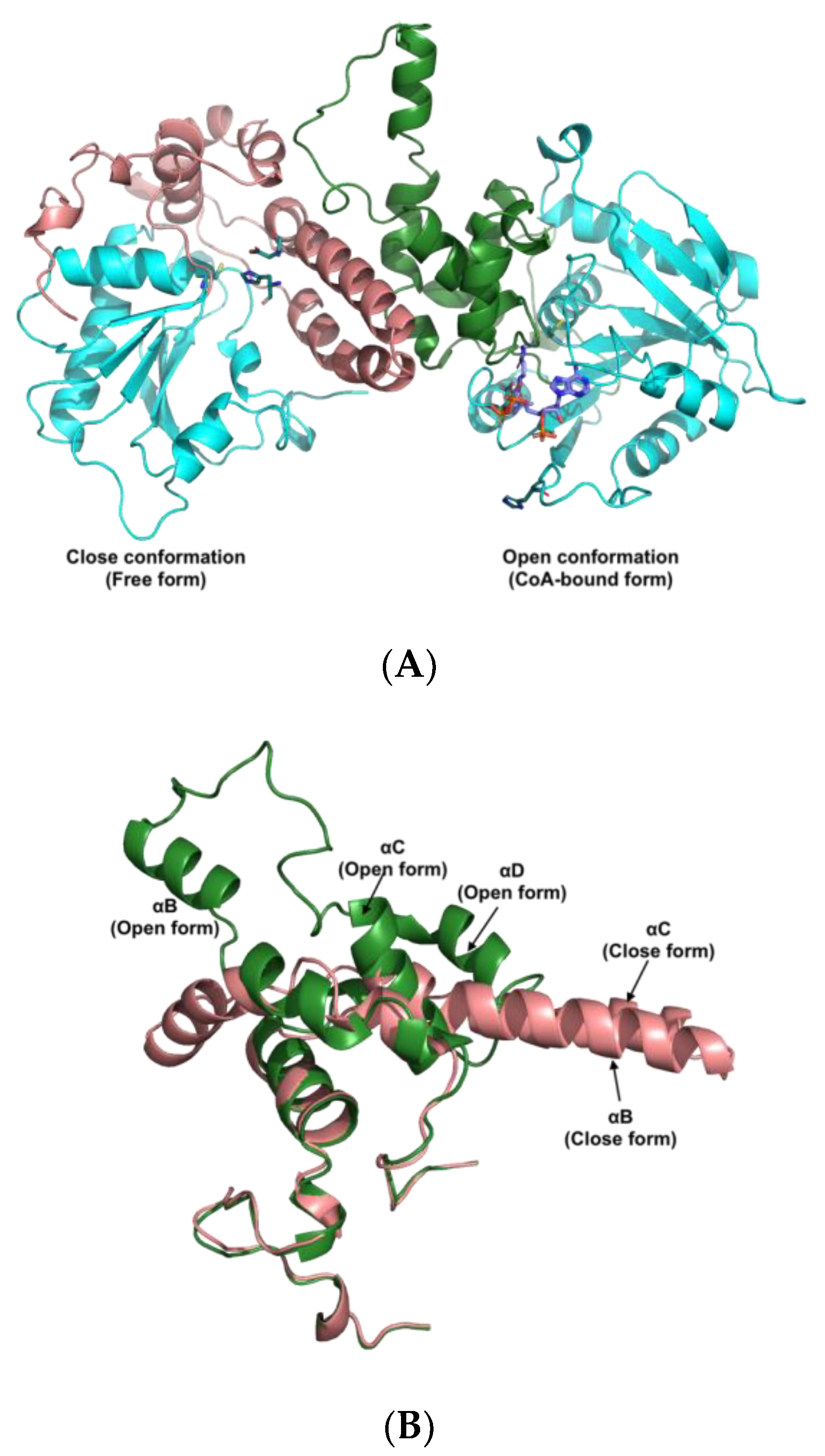
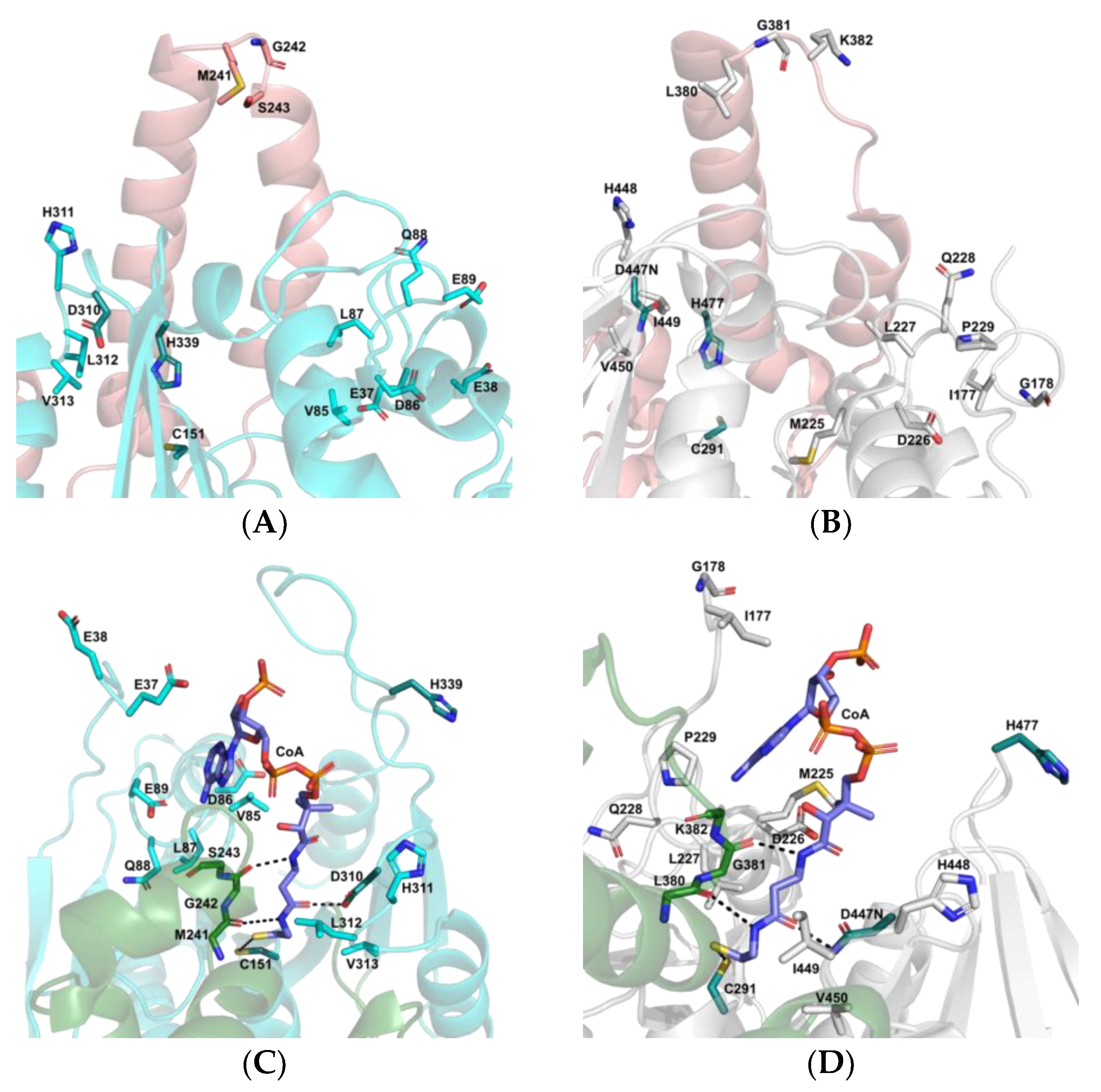
3.5. The Active Site and Catalytic Mechanism of PhaCAp
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agarwal, P.; Soni, R.; Kaur, P.; Madan, A.; Mishra, R.; Pandey, J.; Singh, S.; Singh, G. Cyanobacteria as a promising alternative for sustainable environment: Synthesis of biofuel and biodegradable plastics. Front. Microbiol. 2022, 13, 939347. [Google Scholar] [CrossRef] [PubMed]
- Price, S.; Kuzhiumparambil, U.; Pernice, M.; Ralph, P. Techno-economic analysis of cyanobacterial PHB bioplastic production. J. Environ. Chem. Eng. 2022, 10, 107502. [Google Scholar] [CrossRef]
- Alves, A.A.; Siqueira, E.C.; Barros, M.P.S.; Silva, P.E.C.; Houllou, L.M. Polyhydroxyalkanoates: A review of microbial production and technology application. Int. J. Environ. Sci. Technol. 2023, 20, 3409–3420. [Google Scholar] [CrossRef]
- Sharma, S.; Sharma, P.; Sharma, V.; Bajaj, B.K. Polyhydroxybutyrate as an eco-friendly alternative of synthetic plastics. In Environmental and Agricultural Microbiology: Applications for Sustainability; Mishra, B.B., Nayak, S.K., Mohapatra, S., Samantaray, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 101–149. [Google Scholar]
- Madbouly, S.A. 8 Bio-based polyhydroxyalkanoates blends and composites. In Biopolymers and Composites: Processing and Characterization; Madbouly, S.A., Zhang, C., Eds.; Boston: Berlin, Germany, 2021; pp. 235–254. [Google Scholar]
- Singh, A.K.; Sharma, L.; Mallick, N.; Mala, J. Progress and challenges in producing polyhydroxyalkanoate biopolymers from cyanobacteria. J. Appl. Phycol. 2017, 29, 1213–1232. [Google Scholar] [CrossRef]
- Wittenborn, E.C.; Jost, M.; Wei, Y.; Stubbe, J.; Drennan, C.L. Structure of the catalytic domain of the class I polyhydroxybutyrate synthase from Cupriavidus necator. J. Biol. Chem. 2016, 291, 25264–25277. [Google Scholar] [CrossRef] [PubMed]
- Chek, M.F.; Hiroe, A.; Hakoshima, T.; Sudesh, K.; Taguchi, S. PHA synthase (PhaC): Interpreting the functions of bioplastic-producing enzyme from a structural perspective. Appl. Microbiol. Biotechnol. 2019, 103, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Chek, M.F.; Kim, S.-Y.; Mori, T.; Arsad, H.; Samian, M.R.; Sudesh, K.; Hakoshima, T. Structure of polyhydroxyalkanoate (PHA) synthase PhaC from Chromobacterium sp. USM2, producing biodegradable plastics. Sci. Rep. 2017, 7, 5312. [Google Scholar] [CrossRef]
- Higuchi-Takeuchi, M.; Motoda, Y.; Kigawa, T.; Numata, K. Class I polyhydroxyalkanoate synthase from the purple photosynthetic bacterium Rhodovulum sulfidophilum predominantly exists as a functional dimer in the absence of a substrate. ACS Omega. 2017, 2, 5071–5078. [Google Scholar] [CrossRef]
- Teh, A.-H.; Chiam, N.-C.; Furusawa, G.; Sudesh, K. Modelling of polyhydroxyalkanoate synthase from Aquitalea sp. USM4 suggests a novel mechanism for polymer elongation. Int. J. Biol. Macromol. 2018, 119, 438–445. [Google Scholar]
- Tan, I.K.P.; Foong, C.P.; Tan, H.T.; Lim, H.; Zain, N.-A.A.; Tan, Y.C.; Hoh, C.C.; Sudesh, K. Polyhydroxyalkanoate (PHA) synthase genes and PHA-associated gene clusters in Pseudomonas spp. and Janthinobacterium spp. isolated from Antarctica. J. Biotechnol. 2020, 313, 18–28. [Google Scholar] [PubMed]
- Mezzolla, V.; D'Urso, O.F.; Poltronieri, P. Role of PhaC type I and type II enzymes during PHA biosynthesis. Polymers 2018, 10, 910. [Google Scholar] [CrossRef] [PubMed]
- Chek, M.F.; Kim, S.-Y.; Mori, T.; Tan, H.T.; Sudesh, K.; Hakoshima, T. Asymmetric open-closed dimer mechanism of polyhydroxyalkanoate synthase PhaC. iScience 2020, 23, 101084. [Google Scholar] [CrossRef] [PubMed]
- Markou, G. Overview of microalgal cultivation, biomass processing and application. In Handbook of Algal Science, Technology and Medicine; Konur, O., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 343–352. [Google Scholar]
- Deschoenmaeker, F.; Facchini, R.; Cabrera Pino, J.C.; Bayon-Vicente, G.; Sachdeva, N.; Flammang, P.; Wattiez, R. Nitrogen depletion in Arthrospira sp. PCC 8005, an ultrastructural point of view. J. Struct. Biol. 2016, 196, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Duangsri, C.; Mudtham, N.A.; Incharoensakdi, A.; Raksajit, W. Enhanced polyhydroxybutyrate (PHB) accumulation in heterotrophically grown Arthrospira platensis under nitrogen deprivation. J. Appl. Phycol. 2020, 32, 3645–3654. [Google Scholar] [CrossRef]
- Monshupanee, T.; Incharoensakdi, A. Enhanced accumulation of glycogen, lipids and polyhydroxybutyrate under optimal nutrients and light intensities in the cyanobacterium Synechocystis sp. PCC 6803. J. Appl. Microbiol. 2014, 116, 830–838. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Numata, K.; Motoda, Y.; Watanabe, S.; Osanai, T.; Kigawa, T. Co-expression of two polyhydroxyalkanoate synthase subunits from Synechocystis sp. PCC 6803 by cell-free synthesis and their specific activity for polymerization of 3-hydroxybutyryl-coenzyme A. Biochemistry 2015, 54, 1401–1407. [Google Scholar] [CrossRef]
- Johnson, M.S.; Lehtonen, J.V. Comparison of protein three-dimensional structures. In Bioinformatics, Sequence, Structure, and Databanks; Higgins, D., Taylor, W., Eds.; Oxford University Press: Oxford, UK, 2000; pp. 15–50. [Google Scholar]
- Lehtonen, J.V.; Still, D.-J.; Rantanen, V.-V.; Ekholm, J.; Björklund, D.; Iftikhar, Z.; Huhtala, M.; Repo, S.; Jussila, A.; Jaakkola, J.; et al. BODIL: A molecular modeling environment for structure-function analysis and drug design. J. Comput. Aided Mol. Des. 2004, 18, 401–419. [Google Scholar] [CrossRef]
- Drozdetskiy, A.; Cole, C.; Procter, J.; Barton, G.J. JPred4: A protein secondary structure prediction server. Nucleic Acids Res. 2015, 43, W389–W394. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef]
- Sippl, M.J. Recognition of errors in three-dimensional structures of proteins. Proteins 1993, 17, 355–362. [Google Scholar] [CrossRef]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef]
- GE Healthcare. Tagged proteins. In Handbook of Affinity Chromatography; General Electric Company: Uppsala, Sweden, 2016; Volume 2, pp. 9–291. [Google Scholar]
- Harada, K.; Kobayashi, S.; Oshima, K.; Yoshida, S.; Tsuge, T.; Sato, S. Engineering of Aeromonas caviae polyhydroxyalkanoate synthase through site-directed mutagenesis for enhanced polymerization of the 3-hydroxyhexanoate. Front. Bioeng. Biotechnol. 2021, 9, 627082. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Nambu, Y.; Mizuno, S.; Tsuge, T. In vivo and in vitro characterization of hydrophilic protein tag-fused Ralstonia eutropha polyhydroxyalkanoate synthase. Int. J. Biol. Macromol. 2019, 138, 379–385. [Google Scholar] [CrossRef]
- Wodzinska, J.; Snell, K.D.; Rhomberg, A.; Sinskey, A.J.; Biemann, K.; Stubbe, J. Polyhydroxybutyrate synthase: Evidence for covalent catalysis. J. Am. Chem. Soc. 1996, 118, 6319–6320. [Google Scholar] [CrossRef]
- Numata, K.; Motoda, Y.; Watanabe, S.; Tochio, N.; Kigawa, T.; Doi, Y. Active intermediates of polyhydroxyalkanoate synthase from Aeromonas caviae in polymerization reaction. Biomacromolecules 2012, 13, 3450–3455. [Google Scholar] [CrossRef] [PubMed]
- Jia, K.; Cao, R.; Hua, D.H.; Li, P. Study of class I and class III polyhydroxyalkanoate (PHA) synthases with substrates containing a modified side chain. Biomacromolecules 2016, 17, 1477–1485. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Kolvek, S.; Goodwin, S.; Lenz, R.W. Poly(hydroxyalkanoic acid) biosynthesis in Ectothiorhodospira shaposhnikovii: Characterization and reactivity of a type III PHA synthase. Biomacromolecules 2004, 5, 40–48. [Google Scholar] [CrossRef]
- Yuan, W.; Jia, Y.; Tian, J.; Snell, K.D.; Müh, U.; Sinskey, A.J.; Lambalot, R.H.; Walsh, C.T.; Stubbe, J. Class I and III polyhydroxyalkanoate synthases from Ralstonia eutropha and Allochromatium vinosum: Characterization and substrate specificity studies. Arch. Biochem. Biophys. 2001, 394, 87. [Google Scholar] [CrossRef]
- Zhang, S.; Yasuo, T.; Lenz, R.W.; Goodwin, S. Kinetic and mechanistic characterization of the polyhydroxybutyrate synthase from Ralstonia eutropha. Biomacromolecules 2000, 1, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Pu, N.; Wang, M.-R.; Li, Z.-J. Characterization of polyhydroxyalkanoate synthases from the marine bacterium Neptunomonas concharum JCM17730. J. Biotechnol. 2020, 319, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Agus, J.; Kahar, P.; Abe, H.; Doi, Y.; Tsuge, T. Molecular weight characterization of poly[(R-3-hydroxybutyrate] synthesized by genetically engineered strains of Escherichia coli. Polym. Degrad. Stab. 2006, 91, 1138–1146. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, C.; Cao, R.; Maurmann, L.; Li, P. Inhibitors of polyhydroxyalkanoate (PHA) synthases: Synthesis, molecular docking, and implications. Chem. BioChem. 2015, 16, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, Y.J.; Choi, S.Y.; Lee, S.Y.; Kim, K.J. Crystal structure of Ralstonia eutropha polyhydroxyalkanoate synthase C-terminal domain and reaction mechanisms. Biotechnol. J. 2017, 12, 1600648. [Google Scholar] [CrossRef]
- Chen, G.Q.; Jiang, X.R. Engineering bacteria for enhanced polyhydroxyalkanoates (PHA) biosynthesis. Synth. Syst. Biotechnol. 2017, 2, 192–197. [Google Scholar] [CrossRef]
- Tian, J.; Sinskey, A.J.; Stubbe, J. Detection of intermediates from the polymerization reaction catalyzed by a D302A mutant of class III polyhydroxyalkanoate (PHA) synthase. Biochemistry 2005, 44, 1495–1503. [Google Scholar] [CrossRef]
- Jia, Y.; Yuan, W.; Wodzinska, J.; Park, C.; Sinskey, A.J.; Stubbe, J. Mechanistic studies on class I polyhydroxybutyrate (PHB) synthase from Ralstonia eutropha: Class I and III synthases share a similar catalytic mechanism. Biochemistry 2001, 40, 1011–1019. [Google Scholar] [CrossRef]
- Stubbe, J.; Tian, J. Polyhydroxyalkanoate (PHA) homeostasis: The role of the PHA synthase. Nat. Prod. Rep. 2003, 20, 445–457. [Google Scholar] [CrossRef]
- Neoh, S.Z.; Chek, M.F.; Tan, H.T.; Linares-Pastén, J.A.; Nandakumar, A.; Hakoshima, T.; Sudesh, K. Polyhydroxyalkanoate synthase (PhaC): The key enzyme for biopolyester synthesis. Curr. Res. Biotechnol. 2022, 4, 87–101. [Google Scholar] [CrossRef]
- Müh, U.; Sinskey, A.J.; Kirby, D.P.; Lane, W.S.; Stubbe, J. PHA synthase from Chromatium vinosum: Cysteine 149 is involved in covalent catalysis. Biochemistry 1999, 38, 826–837. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conserved Residue | PhaCCn | PhaCCs | PhaCAp | Conserved Residue | PhaCCn | PhaCCs | PhaCAp |
|---|---|---|---|---|---|---|---|
| (This Study) | (This Study) | ||||||
| V | V211 | V183 | V43 | L | L316 | L288 | L148 |
| T | T212 | T184 | T44 | G | G317 | G289 | G149 |
| V | V217 | V189 | V49 | C | C319 | C291 | C151 |
| L | L225 | L197 | L57 | G | G321 | G293 | G153 |
| P | P230 | P202 | P62 | G | G322 | G294 | G154 |
| K | K234 | K206 | K66 | Y | Y406 | Y373 | Y234 |
| P | P239 | P211 | P71 | G | G414 | G381 | G242 |
| L | L240 | L212 | L72 | W | W425 | W392 | W256 |
| L | L241 | L213 | L73 | D | D428 | D395 | D259 |
| V | V243 | V215 | V75 | L | L441 | L408 | L272 |
| N | N248 | N220 | N80 | Y | Y445 | Y412 | Y276 |
| D | D254 | D226 | D86 | N | N448 | N415 | N279 |
| L | L255 | L227 | L87 | L | L450 | L417 | L280 |
| Q | Q256 | Q228 | Q88 | G | G454 | G421 | G284 |
| S | S260 | S232 | S92 | D | D464 | D431 | D294 |
| V | V262 | V234 | V94 | I | I468 | I435 | I298 |
| G | G269 | G241 | G101 | P | P471 | P438 | P301 |
| V | V272 | V244 | V104 | D | D480 | D447N | D310 |
| L | L274 | L246 | L106 | H | H481 | H448 | H311 |
| W | W277 | W249 | W109 | V | V483 | V450 | V313 |
| T | T288 | T260 | T120 | G | G507 | G476 | G338 |
| Y | Y292 | Y264 | Y124 | H | H508 | H477 | H339 |
| I | I293 | I265 | I125 | W | W554 | W523 | W359 |
| N | N314 | N286 | N146 | L | L555 | L524 | L360 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duangsri, C.; Salminen, T.A.; Alix, M.; Kaewmongkol, S.; Akrimajirachoote, N.; Khetkorn, W.; Jittapalapong, S.; Mäenpää, P.; Incharoensakdi, A.; Raksajit, W. Characterization and Homology Modeling of Catalytically Active Recombinant PhaCAp Protein from Arthrospira platensis. Biology 2023, 12, 751. https://doi.org/10.3390/biology12050751
Duangsri C, Salminen TA, Alix M, Kaewmongkol S, Akrimajirachoote N, Khetkorn W, Jittapalapong S, Mäenpää P, Incharoensakdi A, Raksajit W. Characterization and Homology Modeling of Catalytically Active Recombinant PhaCAp Protein from Arthrospira platensis. Biology. 2023; 12(5):751. https://doi.org/10.3390/biology12050751
Chicago/Turabian StyleDuangsri, Chanchanok, Tiina A. Salminen, Marion Alix, Sarawan Kaewmongkol, Nattaphong Akrimajirachoote, Wanthanee Khetkorn, Sathaporn Jittapalapong, Pirkko Mäenpää, Aran Incharoensakdi, and Wuttinun Raksajit. 2023. "Characterization and Homology Modeling of Catalytically Active Recombinant PhaCAp Protein from Arthrospira platensis" Biology 12, no. 5: 751. https://doi.org/10.3390/biology12050751