
Causal Effects of Modifiable Behaviors on Prostate Cancer in Europeans and East Asians: A Comprehensive Mendelian Randomization Study

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
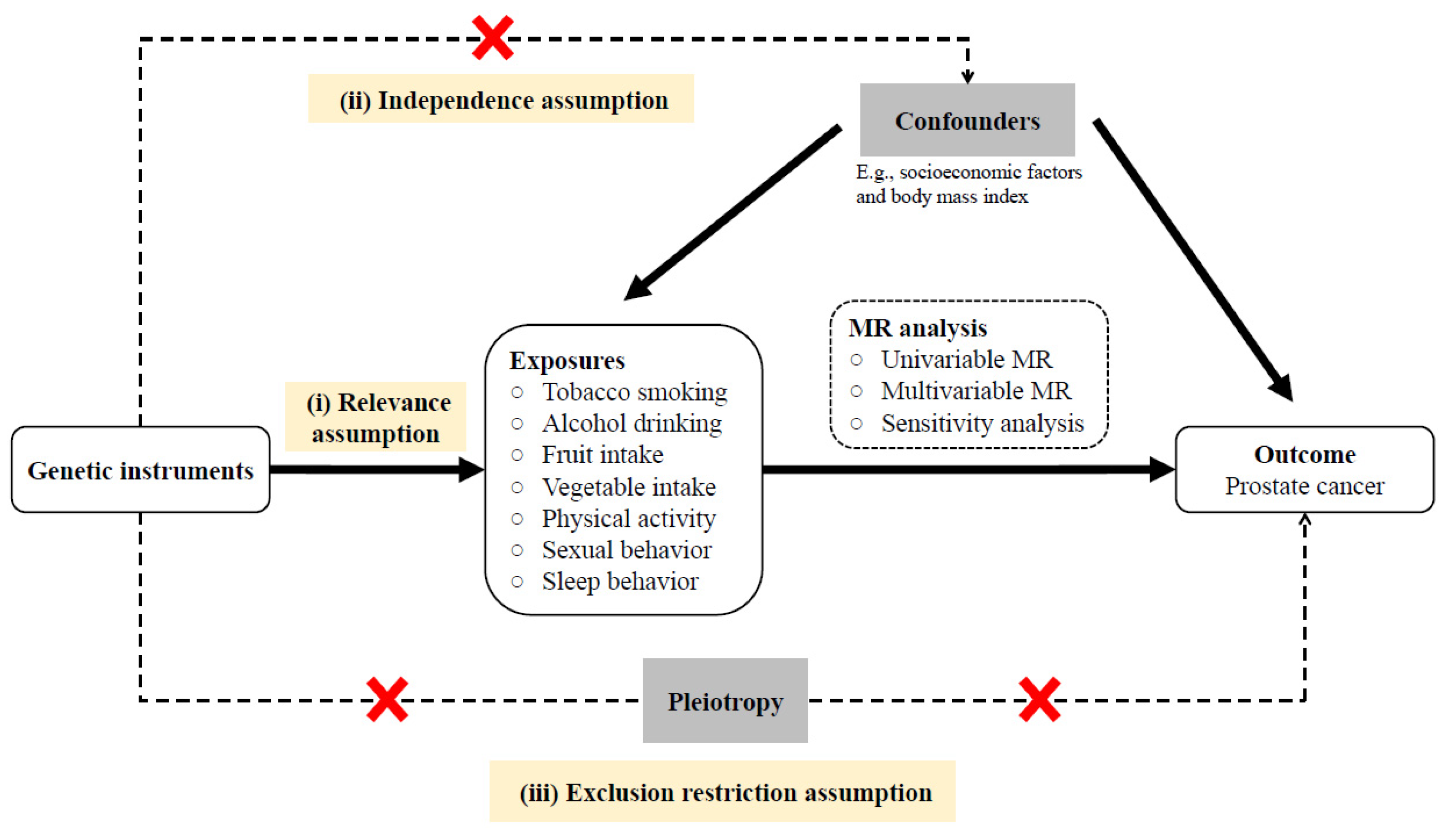
2. Materials and Methods
2.1. Selection of Genetic Variants
2.2. Outcome Traits
2.3. Statistical Analysis
3. Results
3.1. Baseline Characteristics
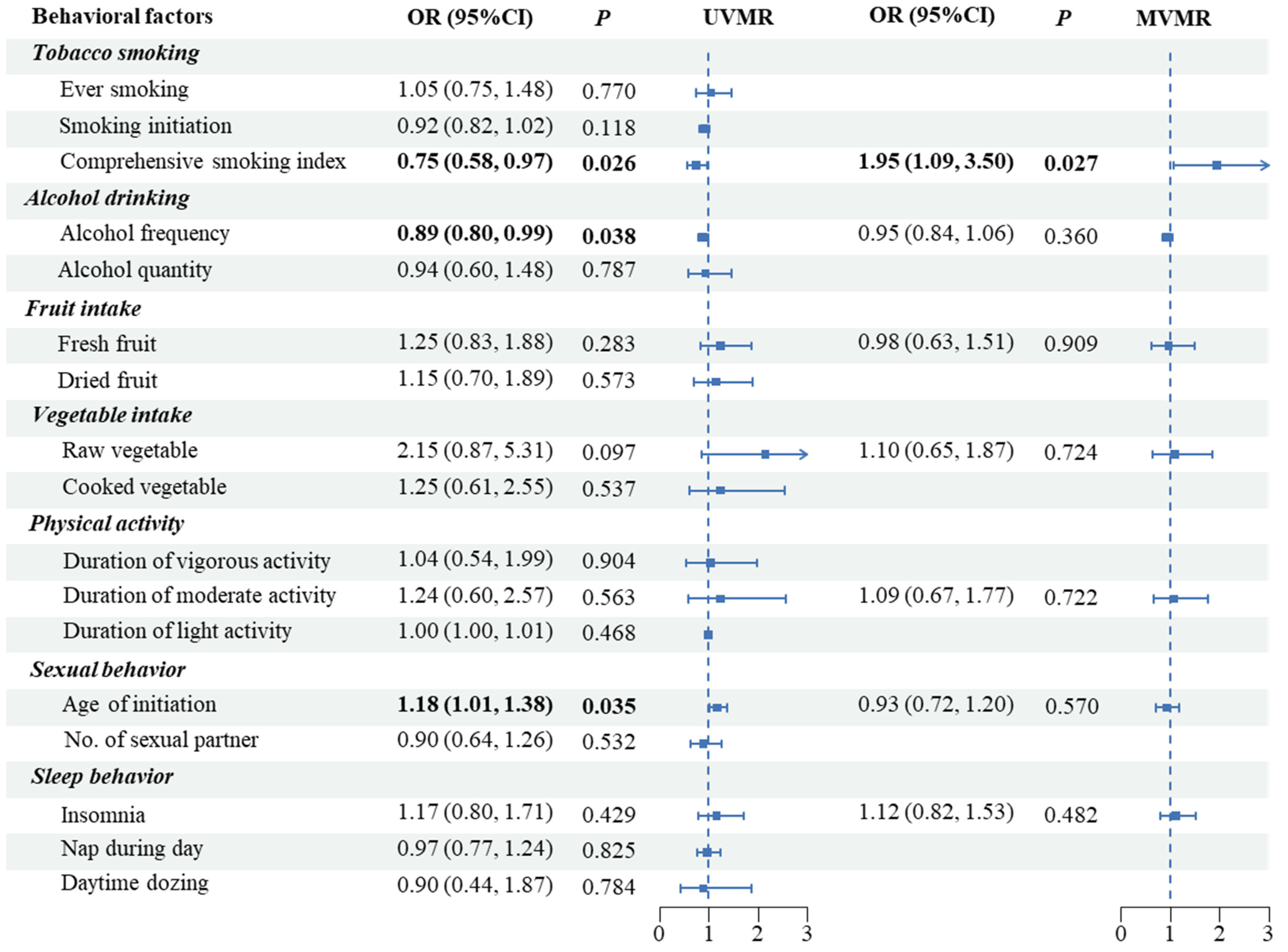
3.2. MR Analyses in Europeans
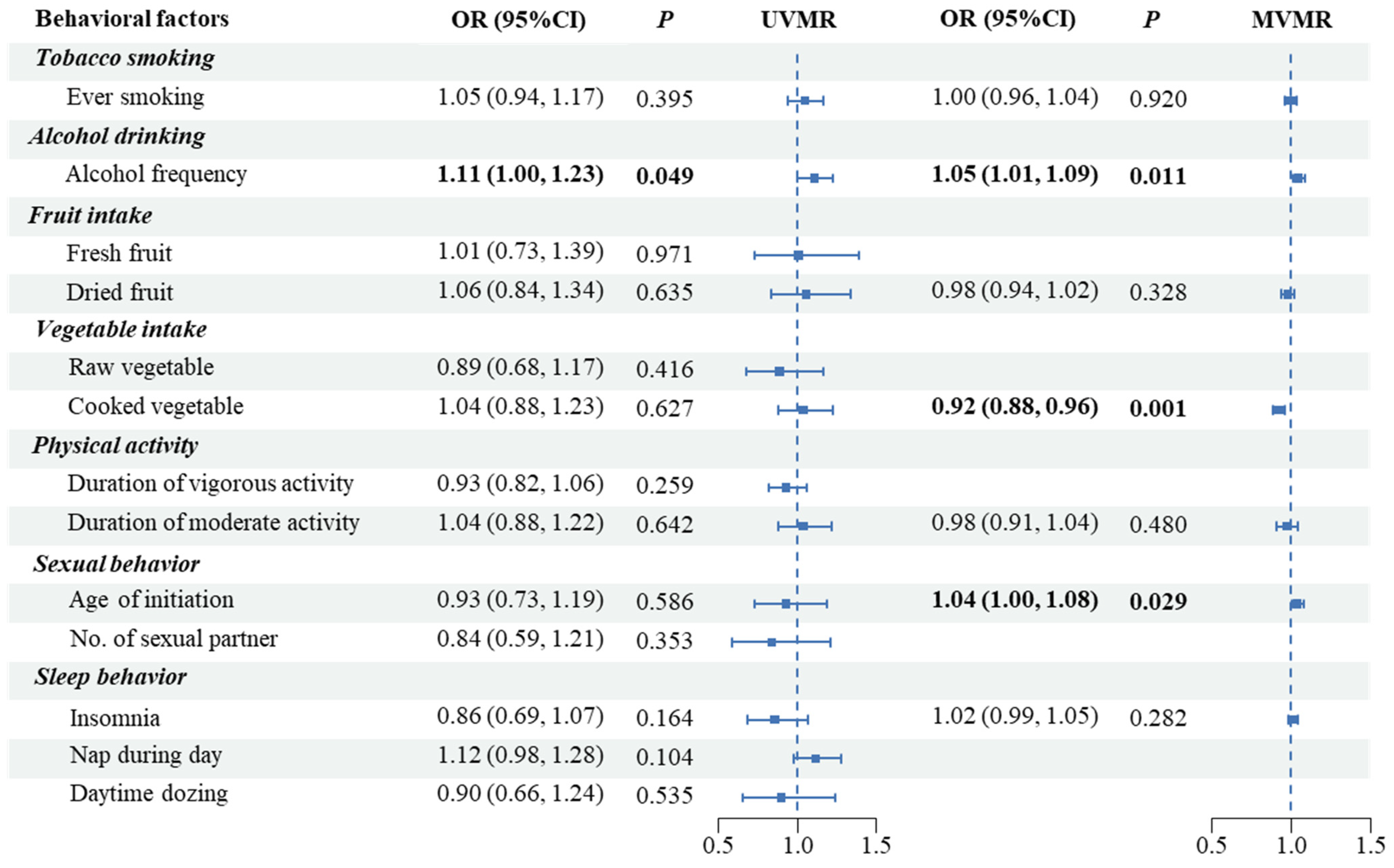
3.3. MR Analyses in East Asians
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Cantarero-Prieto, D.; Lera, J.; Lanza-Leon, P.; Barreda-Gutierrez, M.; Guillem-Porta, V.; Castelo-Branco, L.; Martin-Moreno, J.M. The Economic Burden of Localized Prostate Cancer and Insights Derived from Cost-Effectiveness Studies of the Different Treatments. Cancers 2022, 14, 4088. [Google Scholar] [CrossRef] [PubMed]
- Ferro, M.; Lucarelli, G.; de Cobelli, O.; Del Giudice, F.; Musi, G.; Mistretta, F.A.; Luzzago, S.; Busetto, G.M.; Buonerba, C.; Sciarra, A.; et al. The emerging landscape of tumor marker panels for the identification of aggressive prostate cancer: The perspective through bibliometric analysis of an Italian translational working group in uro-oncology. Minerva Urol. Nephrol. 2021, 73, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Brookman-May, S.D.; Campi, R.; Henriquez, J.D.S.; Klatte, T.; Langenhuijsen, J.F.; Brausi, M.; Linares-Espinos, E.; Volpe, A.; Marszalek, M.; Akdogan, B.; et al. Latest Evidence on the Impact of Smoking, Sports, and Sexual Activity as Modifiable Lifestyle Risk Factors for Prostate Cancer Incidence, Recurrence, and Progression: A Systematic Review of the Literature by the European Association of Urology Section of Oncological Urology (ESOU). Eur. Urol. Focus 2019, 5, 756–787. [Google Scholar] [CrossRef] [PubMed]
- Islami, F.; Moreira, D.M.; Boffetta, P.; Freedland, S.J. A systematic review and meta-analysis of tobacco use and prostate cancer mortality and incidence in prospective cohort studies. Eur. Urol. 2014, 66, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Khil, H.; Lee, D.H.; Keum, N.; Giovannucci, E.L. Alcohol Consumption and the Risk of Prostate Cancer: A Dose-Response Meta-Analysis. Nutrients 2020, 12, 2188. [Google Scholar] [CrossRef]
- Cirne, F.; Kappel, C.; Zhou, S.; Mukherjee, S.D.; Dehghan, M.; Petropoulos, J.A.; Leong, D.P. Modifiable risk factors for prostate cancer in low- and lower-middle-income countries: A systematic review and meta-analysis. Prostate Cancer Prostatic Dis. 2022, 25, 453–462. [Google Scholar] [CrossRef]
- Kazmi, N.; Haycock, P.; Tsilidis, K.; Lynch, B.M.; Truong, T.; Practical Consortium, C.B.C.P.; Martin, R.M.; Lewis, S.J. Appraising causal relationships of dietary, nutritional and physical-activity exposures with overall and aggressive prostate cancer: Two-sample Mendelian-randomization study based on 79 148 prostate-cancer cases and 61 106 controls. Int. J. Epidemiol. 2020, 49, 587–596. [Google Scholar] [CrossRef]
- Huang, J.; Huang, D.; Ruan, X.; Huang, J.; Xu, D.; Heavey, S.; Olivier, J.; Na, R. Association between cannabis use with urological cancers: A population-based cohort study and a mendelian randomization study in the UK biobank. Cancer Med. 2022, 12, 3468–3476. [Google Scholar] [CrossRef]
- Larsson, S.C.; Carter, P.; Kar, S.; Vithayathil, M.; Mason, A.M.; Michaelsson, K.; Burgess, S. Smoking, alcohol consumption, and cancer: A mendelian randomisation study in UK Biobank and international genetic consortia participants. PLoS Med. 2020, 17, e1003178. [Google Scholar] [CrossRef]
- Liu, M.; Jiang, Y.; Wedow, R.; Li, Y.; Brazel, D.M.; Chen, F.; Datta, G.; Davila-Velderrain, J.; McGuire, D.; Tian, C.; et al. Association studies of up to 1.2 million individuals yield new insights into the genetic etiology of tobacco and alcohol use. Nat. Genet. 2019, 51, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Kensler, K.H.; Hu, Z.; Zhang, Y.; Zhang, T.; Jiang, J.; Xu, M.; Pan, Y.; Long, M.; Montone, K.T.; et al. Integrative comparison of the genomic and transcriptomic landscape between prostate cancer patients of predominantly African or European genetic ancestry. PLoS Genet. 2020, 16, e1008641. [Google Scholar] [CrossRef]
- Xu, J.; Mo, Z.; Ye, D.; Wang, M.; Liu, F.; Jin, G.; Xu, C.; Wang, X.; Shao, Q.; Chen, Z.; et al. Genome-wide association study in Chinese men identifies two new prostate cancer risk loci at 9q31.2 and 19q13.4. Nat. Genet. 2012, 44, 1231–1235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Huang, D.; Jiang, G.; Chen, S.; Ruan, X.; Chen, H.; Huang, J.; Liu, A.; Zhang, W.; Lin, X.; et al. Genome-Wide 3’-UTR Single Nucleotide Polymorphism Association Study Identifies Significant Prostate Cancer Risk-Associated Functional Loci at 8p21.2 in Chinese Population. Adv. Sci. 2022, 9, e2201420. [Google Scholar] [CrossRef]
- Skrivankova, V.W.; Richmond, R.C.; Woolf, B.A.R.; Yarmolinsky, J.; Davies, N.M.; Swanson, S.A.; VanderWeele, T.J.; Higgins, J.P.T.; Timpson, N.J.; Dimou, N.; et al. Strengthening the Reporting of Observational Studies in Epidemiology Using Mendelian Randomization: The STROBE-MR Statement. JAMA 2021, 326, 1614–1621. [Google Scholar] [CrossRef]
- Schumacher, F.R.; Al Olama, A.A.; Berndt, S.I.; Benlloch, S.; Ahmed, M.; Saunders, E.J.; Dadaev, T.; Leongamornlert, D.; Anokian, E.; Cieza-Borrella, C.; et al. Association analyses of more than 140,000 men identify 63 new prostate cancer susceptibility loci. Nat. Genet. 2018, 50, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.; Butterworth, A.; Thompson, S.G. Mendelian randomization analysis with multiple genetic variants using summarized data. Genet. Epidemiol. 2013, 37, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, E. Multivariable Mendelian Randomization and Mediation. Cold Spring Harb. Perspect. Med. 2021, 11, a038984. [Google Scholar] [CrossRef]
- Secretan, B.; Straif, K.; Baan, R.; Grosse, Y.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Freeman, C.; Galichet, L.; et al. A review of human carcinogens--Part E: Tobacco, areca nut, alcohol, coal smoke, and salted fish. Lancet Oncol. 2009, 10, 1033–1034. [Google Scholar] [CrossRef]
- Jochems, S.H.J.; Fritz, J.; Haggstrom, C.; Jarvholm, B.; Stattin, P.; Stocks, T. Smoking and Risk of Prostate Cancer and Prostate Cancer Death: A Pooled Study. Eur. Urol. 2022, 83, 422–431. [Google Scholar] [CrossRef]
- Littlejohns, T.J.; Travis, R.C.; Key, T.J.; Allen, N.E. Lifestyle factors and prostate-specific antigen (PSA) testing in UK Biobank: Implications for epidemiological research. Cancer Epidemiol. 2016, 45, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Wootton, R.E.; Richmond, R.C.; Stuijfzand, B.G.; Lawn, R.B.; Sallis, H.M.; Taylor, G.M.J.; Hemani, G.; Jones, H.J.; Zammit, S.; Davey Smith, G.; et al. Evidence for causal effects of lifetime smoking on risk for depression and schizophrenia: A Mendelian randomisation study. Psychol. Med. 2020, 50, 2435–2443. [Google Scholar] [CrossRef]
- Sawada, N. Risk and preventive factors for prostate cancer in Japan: The Japan Public Health Center-based prospective (JPHC) study. J. Epidemiol. 2017, 27, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Macke, A.J.; Petrosyan, A. Alcohol and Prostate Cancer: Time to Draw Conclusions. Biomolecules 2022, 12, 375. [Google Scholar] [CrossRef]
- Montel, R.A.; Munoz-Zuluaga, C.; Stiles, K.M.; Crystal, R.G. Can gene therapy be used to prevent cancer? Gene therapy for aldehyde dehydrogenase 2 deficiency. Cancer Gene Ther. 2022, 29, 889–896. [Google Scholar] [CrossRef]
- Resnick, M.J.; Lacchetti, C.; Bergman, J.; Hauke, R.J.; Hoffman, K.E.; Kungel, T.M.; Morgans, A.K.; Penson, D.F. Prostate cancer survivorship care guideline: American Society of Clinical Oncology Clinical Practice Guideline endorsement. J. Clin. Oncol. 2015, 33, 1078–1085. [Google Scholar] [CrossRef]
- Yan, H.; Cui, X.; Zhang, P.; Li, R. Fruit and Vegetable Consumption and the Risk of Prostate Cancer: A Systematic Review and Meta-Analysis. Nutr. Cancer 2022, 74, 1235–1242. [Google Scholar] [CrossRef]
- Parsons, J.K.; Zahrieh, D.; Mohler, J.L.; Paskett, E.; Hansel, D.E.; Kibel, A.S.; Liu, H.; Seisler, D.K.; Natarajan, L.; White, M.; et al. Effect of a Behavioral Intervention to Increase Vegetable Consumption on Cancer Progression Among Men with Early-Stage Prostate Cancer: The MEAL Randomized Clinical Trial. JAMA 2020, 323, 140–148. [Google Scholar] [CrossRef]
- Miglio, C.; Chiavaro, E.; Visconti, A.; Fogliano, V.; Pellegrini, N. Effects of different cooking methods on nutritional and physicochemical characteristics of selected vegetables. J. Agric. Food Chem. 2008, 56, 139–147. [Google Scholar] [CrossRef]
- Jian, Z.; Ye, D.; Chen, Y.; Li, H.; Wang, K. Sexual Activity and Risk of Prostate Cancer: A Dose-Response Meta-Analysis. J. Sex Med. 2018, 15, 1300–1309. [Google Scholar] [CrossRef] [PubMed]
- Rider, J.R.; Wilson, K.M.; Sinnott, J.A.; Kelly, R.S.; Mucci, L.A.; Giovannucci, E.L. Ejaculation Frequency and Risk of Prostate Cancer: Updated Results with an Additional Decade of Follow-up. Eur. Urol. 2016, 70, 974–982. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Phenotype | GWAS ID | Total Population | European Discovery | European Replication | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SNPs (n) | F-Stat | R2 (%) | Power (%) | SNPs (n) | F-Stat | R2 (%) | Power (%) | |||
| Tobacco smoking | ||||||||||
| Ever smoking | ukb-b-20261 | 461,066 | 79 | 9.29 | 0.16 | 39 | 80 | 9.27 | 0.16 | 10 |
| Smoking initiation | ieu-b-4877 | 632,802 | 92 | 172.17 | 2.44 | >99 | 87 | 173.00 | 2.32 | 68 |
| Lifetime smoking index | N/A | 462,690 | 108 | 20.27 | 0.47 | 82 | 120 | 19.65 | 0.51 | 21 |
| Alcohol drinking | ||||||||||
| Alcohol frequency | ukb-b-5779 | 462,346 | 87 | 113.08 | 2.08 | >99 | 97 | 130.57 | 2.40 | 70 |
| Alcohol quantity | ieu-b-73 | 335,394 | 33 | 32.94 | 0.31 | 65 | 31 | 31.34 | 0.30 | 14 |
| Fruit intake | ||||||||||
| Fresh fruit | ukb-b-3881 | 446,462 | 49 | 15.60 | 0.17 | 41 | 53 | 16.65 | 0.18 | 10 |
| Dried fruit | ukb-b-16576 | 421,764 | 36 | 24.08 | 0.21 | 49 | 39 | 26.64 | 0.23 | 12 |
| Vegetable intake | ||||||||||
| Raw vegetables | ukb-b-1996 | 435,435 | 20 | 16.52 | 0.08 | 21 | 22 | 18.85 | 0.09 | 8 |
| Cooked vegetables | ukb-b-8089 | 448,651 | 15 | 16.93 | 0.07 | 20 | 16 | 24.03 | 0.07 | 7 |
| Physical activity | ||||||||||
| Duration of vigorous activity | ukb-b-13932 | 251,501 | 3 | 3.47 | <0.01 | 5 | 3 | 3.47 | <0.01 | 5 |
| Duration of moderate activity | ukb-b-2346 | 343,827 | 3 | 7.07 | <0.01 | 6 | 3 | 10.91 | <0.01 | 5 |
| Duration of light activity | ukb-b-8865 | 64,949 | 3 | 2.28 | 0.01 | 5 | 3 | 2.28 | 0.01 | 5 |
| Sexual behavior | ||||||||||
| Age of initiation | ebi-a-GCST90000047 | 397,338 | 167 | 16.70 | 0.70 | 94 | 169 | 16.85 | 0.70 | 27 |
| No. of sexual partners | ukb-b-4256 | 378,882 | 59 | 25.70 | 0.39 | 75 | 59 | 25.41 | 0.39 | 17 |
| Sleep behavior | ||||||||||
| Insomnia | ukb-b-3957 | 462,341 | 36 | 8.64 | 0.13 | 33 | 40 | 12.98 | 0.20 | 11 |
| Nap during day | ebi-a-GCST011494 | 452,633 | 96 | 15.20 | 0.32 | 67 | 95 | 15.88 | 0.34 | 15 |
| Daytime dozing | ukb-b-5776 | 460,913 | 26 | 10.58 | 0.06 | 18 | 28 | 10.39 | 0.06 | 7 |
| Phenotype | GWAS ID | Total Population | East Asian Discovery | East Asian Replication | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SNPs (n) | F-Stat | R2 (%) | Power (%) | SNPs (n) | F-Stat | R2 (%) | Power (%) | |||
| Tobacco smoking | ||||||||||
| Ever smoking | ukb-e-20160_EAS | 2649 | 10 | 6.17 | 2.29 | 28 | 7 | 6.91 | 1.80 | 49 |
| Alcohol drinking | ||||||||||
| Alcohol frequency | ukb-e-1558_EAS | 2658 | 6 | 18.96 | 4.11 | 46 | 4 | 20.12 | 2.94 | 70 |
| Fruit intake | ||||||||||
| Fresh fruit | ukb-e-1309_EAS | 2513 | 6 | 3.41 | 0.81 | 13 | 5 | 3.85 | 0.76 | 24 |
| Dried fruit | ukb-e-1319_EAS | 2219 | 11 | 7.23 | 3.48 | 40 | 9 | 6.69 | 2.65 | 66 |
| Vegetables intake | ||||||||||
| Raw vegetables | ukb-e-1299_EAS | 2365 | 3 | 2.83 | 0.36 | 9 | 3 | 2.83 | 0.36 | 14 |
| Cooked vegetables | ukb-e-1289_EAS | 2499 | 19 | 10.67 | 7.56 | 71 | 19 | 10.67 | 7.56 | 98 |
| Physical activity | ||||||||||
| Duration of vigorous activity | ukb-e-914_EAS | 1373 | 4 | 5.13 | 1.48 | 20 | 4 | 5.13 | 1.48 | 42 |
| Duration of moderate activity | ukb-e-894_EAS | 1853 | 3 | 3.19 | 0.52 | 10 | 3 | 3.19 | 0.52 | 18 |
| Sexual behavior | ||||||||||
| Age of initiation | ukb-e-2139_EAS | 1711 | 4 | 5.65 | 1.31 | 18 | 4 | 5.65 | 1.31 | 38 |
| No. of sexual partners | ukb-e-2149_EAS | 1559 | 5 | 9.27 | 2.90 | 34 | 5 | 9.27 | 2.90 | 69 |
| Sleep behavior | ||||||||||
| Insomnia | ukb-e-1200_EAS | 2654 | 18 | 6.93 | 4.03 | 45 | 16 | 5.88 | 3.45 | 77 |
| Nap during day | ukb-e-1190_EAS | 2606 | 14 | 6.67 | 3.24 | 38 | 13 | 6.36 | 3.09 | 72 |
| Daytime dozing | ukb-e-1220_EAS | 2582 | 16 | 7.11 | 4.25 | 47 | 16 | 7.11 | 4.25 | 85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhan, Y.; Ruan, X.; Wang, P.; Huang, D.; Huang, J.; Huang, J.; Chun, T.T.S.; Ho, B.S.-H.; Ng, A.T.-L.; Tsu, J.H.-L.; et al. Causal Effects of Modifiable Behaviors on Prostate Cancer in Europeans and East Asians: A Comprehensive Mendelian Randomization Study. Biology 2023, 12, 673. https://doi.org/10.3390/biology12050673
Zhan Y, Ruan X, Wang P, Huang D, Huang J, Huang J, Chun TTS, Ho BS-H, Ng AT-L, Tsu JH-L, et al. Causal Effects of Modifiable Behaviors on Prostate Cancer in Europeans and East Asians: A Comprehensive Mendelian Randomization Study. Biology. 2023; 12(5):673. https://doi.org/10.3390/biology12050673
Chicago/Turabian StyleZhan, Yongle, Xiaohao Ruan, Pei Wang, Da Huang, Jingyi Huang, Jinlun Huang, Tsun Tsun Stacia Chun, Brian Sze-Ho Ho, Ada Tsui-Lin Ng, James Hok-Leung Tsu, and et al. 2023. "Causal Effects of Modifiable Behaviors on Prostate Cancer in Europeans and East Asians: A Comprehensive Mendelian Randomization Study" Biology 12, no. 5: 673. https://doi.org/10.3390/biology12050673