
Me, Myself and My Insula: An Oasis in the Forefront of Self-Consciousness

{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
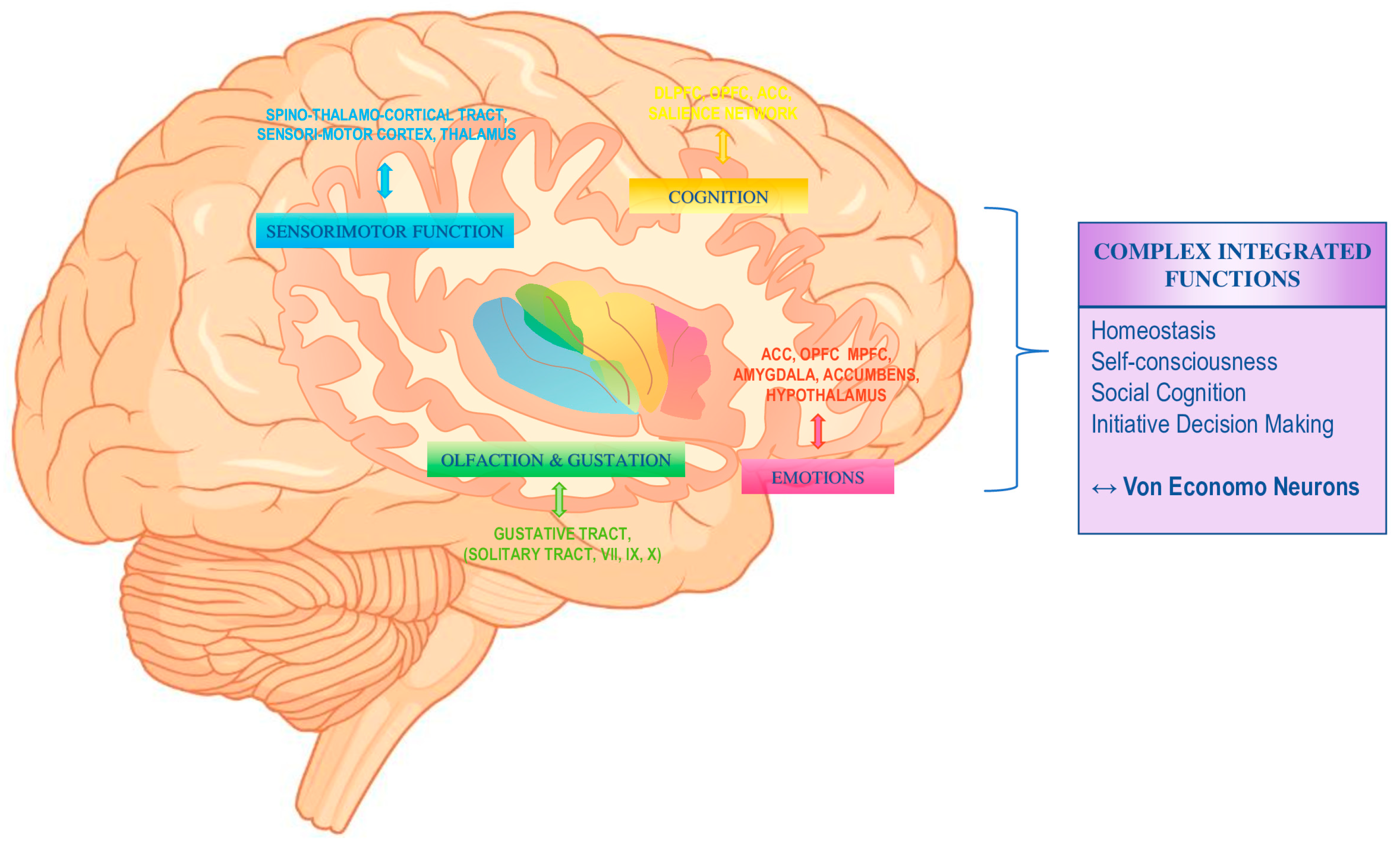
2. The Insula
2.1. General Anatomy and Functions of the Insula
2.2. From Viscera to Insular Cortex
2.3. From Interoception to Awareness
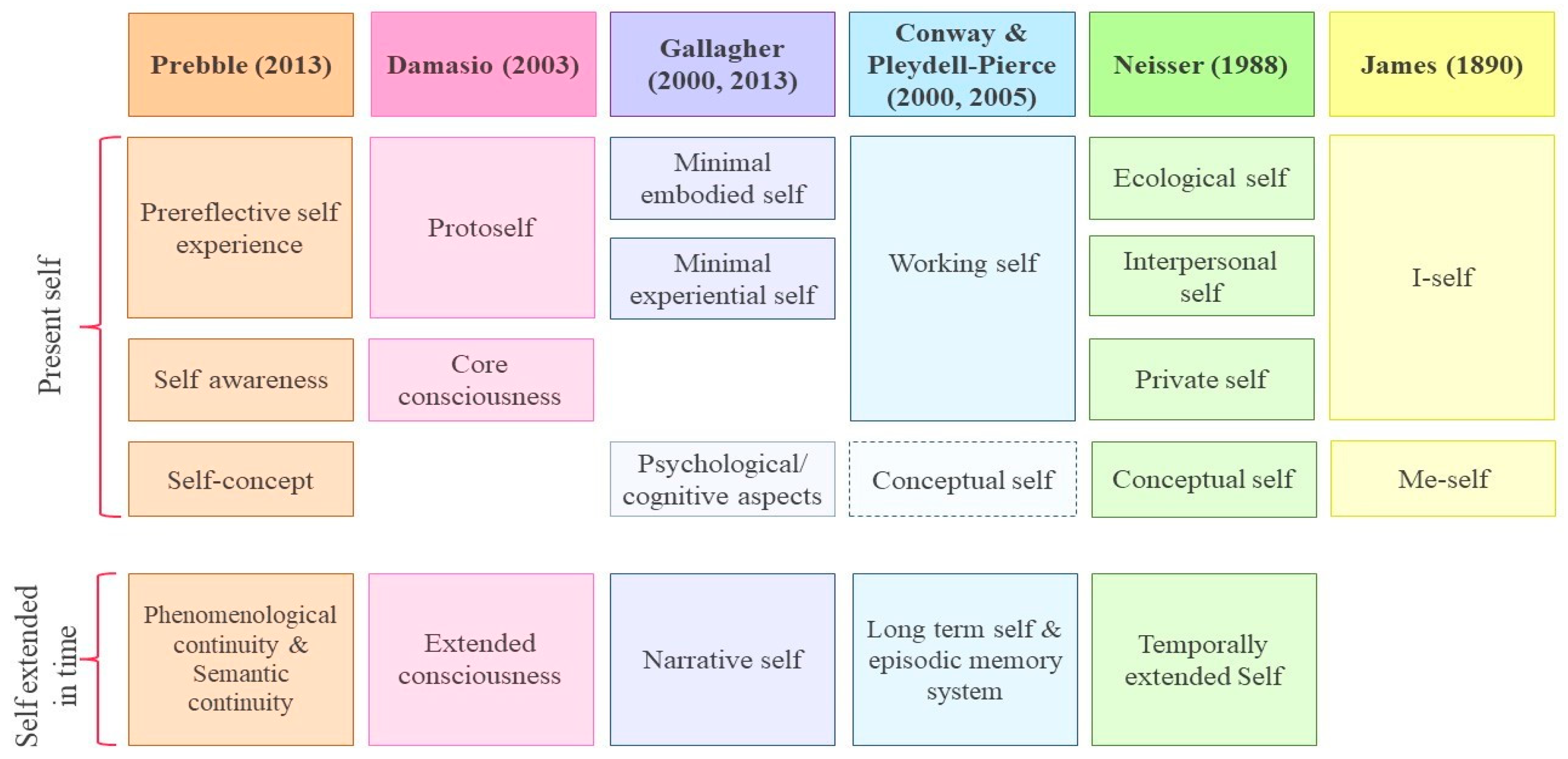
3. The Self
3.1. The Present Self
3.1.1. The Subjective Sense of Self
3.1.2. The Self-Concept
3.2. The Self Extended in Time, or Autobiographical Memories
4. Insula and the Self
4.1. Insula and the Subjective Sense of Self
4.1.1. Subjective Sense of Self in Healthy Insula
4.1.2. Subjective Sense of Self in Damaged Insula
4.2. Insula and the Self-Concept
4.2.1. Self-Concept in Healthy Insula
4.2.2. Self-Concept in Damaged Insula
4.3. Insula and the Self Extended in Time
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Frewen, P.; Schroeter, M.L.; Riva, G.; Cipresso, P.; Fairfield, B.; Padulo, C.; Kemp, A.H.; Palaniyappan, L.; Owolabi, M.; Kusi-Mensah, K.; et al. Neuroimaging the consciousness of self: Review, and conceptual-methodological framework. Neurosci. Biobehav. Rev. 2020, 112, 164–212. [Google Scholar] [CrossRef]
- Uddin, L.Q.; Iacoboni, M.; Lange, C.; Keenan, J.P. The self and social cognition: The role of cortical midline structures and mirror neurons. Trends Cogn. Sci. 2007, 11, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Wicker, B.; Ruby, P.; Royet, J.P.; Fonlupt, P. A relation between rest and the self in the brain? Brain Res. Brain Res. Rev. 2003, 43, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Rosenblum, K.; Meiri, N.; Dudai, Y. Taste memory: The role of protein synthesis in gustatory cortex. Behav. Neural Biol. 1993, 59, 49–56. [Google Scholar] [CrossRef]
- Maffei, A.; Haley, M.; Fontanini, A. Neural processing of gustatory information in insular circuits. Curr. Opin. Neurobiol. 2012, 22, 709–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, A.D. How do you feel–now? The anterior insula and human awareness. Nat. Rev. Neurosci. 2009, 10, 59–70. [Google Scholar] [CrossRef]
- Kurth, F.; Zilles, K.; Fox, P.T.; Laird, A.R.; Eickhoff, S.B. A link between the systems: Functional differentiation and integration within the human insula revealed by meta-analysis. Brain Struct. Funct. 2010, 214, 519–534. [Google Scholar] [CrossRef]
- Morel, A.; Gallay, M.N.; Baechler, A.; Wyss, M.; Gallay, D.S. The human insula: Architectonic organization and postmortem MRI registration. Neuroscience 2013, 236, 117–135. [Google Scholar] [CrossRef] [Green Version]
- Craig, A.D. Topographically organized projection to posterior insular cortex from the posterior portion of the ventral medial nucleus in the long-tailed macaque monkey. J. Comp. Neurol. 2014, 522, 36–63. [Google Scholar] [CrossRef] [Green Version]
- Stephani, C.; Fernandez-Baca Vaca, G.; Maciunas, R.; Koubeissi, M.; Luders, H.O. Functional neuroanatomy of the insular lobe. Brain Struct. Funct. 2011, 216, 137–149. [Google Scholar] [CrossRef] [Green Version]
- Calder, A.J.; Keane, J.; Manes, F.; Antoun, N.; Young, A.W. Impaired recognition and experience of disgust following brain injury. Nat. Neurosci. 2000, 3, 1077–1078. [Google Scholar] [CrossRef]
- Fan, J.; Gu, X.; Liu, X.; Guise, K.G.; Park, Y.; Martin, L.; de Marchena, A.; Tang, C.Y.; Minzenberg, M.J.; Hof, P.R. Involvement of the anterior cingulate and frontoinsular cortices in rapid processing of salient facial emotional information. Neuroimage 2011, 54, 2539–2546. [Google Scholar] [CrossRef] [Green Version]
- Picard, F.; Scavarda, D.; Bartolomei, F. Induction of a sense of bliss by electrical stimulation of the anterior insula. Cortex 2013, 49, 2935–2937. [Google Scholar] [CrossRef] [PubMed]
- Seeley, W.W.; Menon, V.; Schatzberg, A.F.; Keller, J.; Glover, G.H.; Kenna, H.; Reiss, A.L.; Greicius, M.D. Dissociable intrinsic connectivity networks for salience processing and executive control. J. Neurosci. 2007, 27, 2349–2356. [Google Scholar] [CrossRef] [Green Version]
- Allman, J.M.; Watson, K.K.; Tetreault, N.A.; Hakeem, A.Y. Intuition and autism: A possible role for Von Economo neurons. Trends Cogn. Sci. 2005, 9, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Butti, C.; Sherwood, C.C.; Hakeem, A.Y.; Allman, J.M.; Hof, P.R. Total number and volume of Von Economo neurons in the cerebral cortex of cetaceans. J. Comp. Neurol. 2009, 515, 243–259. [Google Scholar] [CrossRef]
- Hakeem, A.Y.; Sherwood, C.C.; Bonar, C.J.; Butti, C.; Hof, P.R.; Allman, J.M. Von Economo neurons in the elephant brain. Anat. Rec. 2009, 292, 242–248. [Google Scholar] [CrossRef]
- Raghanti, M.A.; Spurlock, L.B.; Treichler, F.R.; Weigel, S.E.; Stimmelmayr, R.; Butti, C.; Thewissen, J.G.M.H.; Hof, P.R. An analysis of von Economo neurons in the cerebral cortex of cetaceans, artiodactyls, and perissodactyls. Brain Struct. Funct. 2015, 220, 2303–2314. [Google Scholar] [CrossRef]
- Ghaziri, J.; Tucholka, A.; Girard, G.; Houde, J.-C.; Boucher, O.; Gilbert, G.; Descoteaux, M.; Lippé, S.; Rainville, P.; Nguyen, D.K. The Corticocortical Structural Connectivity of the Human Insula. Cereb. Cortex 2017, 27, 1216–1228. [Google Scholar] [CrossRef] [Green Version]
- Cerliani, L.; Thomas, R.M.; Jbabdi, S.; Siero, J.C.; Nanetti, L.; Crippa, A.; Gazzola, V.; D’Arceuil, H.; Keysers, C. Probabilistic tractography recovers a rostrocaudal trajectory of connectivity variability in the human insular cortex. Hum. Brain Mapp. 2012, 33, 2005–2034. [Google Scholar] [CrossRef] [PubMed]
- Cloutman, L.L.; Binney, R.J.; Drakesmith, M.; Parker, G.J.; Lambon Ralph, M.A. The variation of function across the human insula mirrors its patterns of structural connectivity: Evidence from in vivo probabilistic tractography. Neuroimage 2012, 59, 3514–3521. [Google Scholar] [CrossRef]
- Ghaziri, J.; Tucholka, A.; Girard, G.; Boucher, O.; Houde, J.-C.; Descoteaux, M.; Obaid, S.; Gilbert, G.; Rouleau, I.; Nguyen, D.K. Subcortical structural connectivity of insular subregions. Sci. Rep. 2018, 8, 8596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddin, L.Q.; Nomi, J.S.; Hebert-Seropian, B.; Ghaziri, J.; Boucher, O. Structure and Function of the Human Insula. J. Clin. Neurophysiol. 2017, 34, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Evrard, H.C. The Organization of the Primate Insular Cortex. Front. Neuroanat. 2019, 13, 43. [Google Scholar] [CrossRef] [Green Version]
- Fischer, D.B.; Boes, A.D.; Demertzi, A.; Evrard, H.C.; Laureys, S.; Edlow, B.L.; Liu, H.; Saper, C.B.; Pascual-Leone, A.; Fox, M.D.; et al. A human brain network derived from coma-causing brainstem lesions. Neurology 2016, 87, 2427–2434. [Google Scholar] [CrossRef] [Green Version]
- Loewy, A.D.; Spyer, K.M. (Eds.) Vagal preganglionic neurons. In Central Regulation of Autonomic Functions; Oxford University Press: New York, NY, USA, 1990; pp. 68–87. [Google Scholar]
- Shipley, M.T. Insular cortex projection to the nucleus of the solitary tract and brainstem visceromotor regions in the mouse. Brain Res. Bull. 1982, 8, 139–148. [Google Scholar] [CrossRef]
- Bates, E.; Wilson, S.M.; Saygin, A.P.; Dick, F.; Sereno, M.I.; Knight, R.T.; Dronkers, N.F. Voxel-based lesion-symptom mapping. Nat. Neurosci. 2003, 6, 448–450. [Google Scholar] [CrossRef]
- Dronkers, N.F. A new brain region for coordinating speech articulation. Nature 1996, 384, 159–161. [Google Scholar] [CrossRef]
- Clark, L.; Bechara, A.; Damasio, H.; Aitken, M.R.; Sahakian, B.J.; Robbins, T.W. Differential effects of insular and ventromedial prefrontal cortex lesions on risky decision-making. Brain 2008, 131, 1311–1322. [Google Scholar] [CrossRef] [Green Version]
- Augustine, J.R. Circuitry and functional aspects of the insular lobe in primates including humans. Brain Res. Brain Res. Rev. 1996, 22, 229–244. [Google Scholar] [CrossRef]
- Karnath, H.O.; Baier, B.; Nagele, T. Awareness of the functioning of one’s own limbs mediated by the insular cortex? J. Neurosci. 2005, 25, 7134–7138. [Google Scholar] [CrossRef] [Green Version]
- Manes, F.; Paradiso, S.; Robinson, R.G. Neuropsychiatric effects of insular stroke. J. Nerv. Ment. Dis. 1999, 187, 707–712. [Google Scholar] [CrossRef]
- Craig, A.D. Interoceptive cortex in the posterior insula: Comment on Garcia-Larrea et al. Brain 2011, 134, e166. [Google Scholar] [CrossRef] [PubMed]
- Penfield, W.; Faulk, M.E., Jr. The insula; further observations on its function. Brain 1955, 78, 445–470. [Google Scholar] [CrossRef]
- Isnard, J.; Guenot, M.; Sindou, M.; Mauguiere, F. Clinical manifestations of insular lobe seizures: A stereo-electroencephalographic study. Epilepsia 2004, 45, 1079–1090. [Google Scholar] [CrossRef]
- Tayah, T.; Savard, M.; Desbiens, R.; Nguyen, D.K. Ictal bradycardia and asystole in an adult with a focal left insular lesion. Clin. Neurol. Neurosurg. 2013, 115, 1885–1887. [Google Scholar] [CrossRef]
- Craig, A.D. Interoception and emotion: A neuroanatomical perspective. In Handbook of Emotions; Lewis, M., Haviland-Jones, J.M., Barrett, L.F., Eds.; Guilford Press: New York, NY, USA, 2008; pp. 272–292. [Google Scholar]
- Craig, A.D. The sentient self. Brain Struct. Funct. 2010, 214, 563–577. [Google Scholar] [CrossRef]
- Craig, A.D. How do you feel? Interoception: The sense of the physiological condition of the body. Nat. Rev. Neurosci. 2002, 3, 655–666. [Google Scholar] [CrossRef] [PubMed]
- MacIver, M.B.; Tanelian, D.L. Activation of C fibers by metabolic perturbations associated with tourniquet ischemia. Anesthesiology 1992, 76, 617–623. [Google Scholar] [CrossRef]
- Weber, E.H. Tastsinn und Gemeingefühl. In Handwörterbuch des Physiologie mit Rücksicht auf Physiologische Pathologie; Wagner, R., Ed.; Vieweg: Braunschweig, Germany, 1846; pp. 481–555. [Google Scholar]
- Craig, A.D. Interoception: The sense of the physiological condition of the body. Curr. Opin. Neurobiol. 2003, 13, 500–505. [Google Scholar] [CrossRef]
- Kuehn, E.; Mueller, K.; Lohmann, G.; Schuetz-Bosbach, S. Interoceptive awareness changes the posterior insula functional connectivity profile. Brain Struct. Funct. 2016, 221, 1555–1571. [Google Scholar] [CrossRef]
- Harrison, N.A.; Gray, M.A.; Gianaros, P.J.; Critchley, H.D. The embodiment of emotional feelings in the brain. J. Neurosci. 2010, 30, 12878–12884. [Google Scholar] [CrossRef] [Green Version]
- Critchley, H.D.; Wiens, S.; Rotshtein, P.; Ohman, A.; Dolan, R.J. Neural systems supporting interoceptive awareness. Nat. Neurosci. 2004, 7, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Simmons, W.K.; Avery, J.A.; Barcalow, J.C.; Bodurka, J.; Drevets, W.C.; Bellgowan, P. Keeping the body in mind: Insula functional organization and functional connectivity integrate interoceptive, exteroceptive, and emotional awareness. Hum. Brain Mapp. 2013, 34, 2944–2958. [Google Scholar] [CrossRef]
- Damasio, A. The Feeling of What Happens: Body, Emotion and the Making of Consciousness; Harcourt: Orlando, FL, USA, 1999. [Google Scholar]
- Seth, A.K. Interoceptive inference, emotion, and the embodied self. Trends Cogn. Sci. 2013, 17, 565–573. [Google Scholar] [CrossRef]
- Berlucchi, G.; Aglioti, S.M. The body in the brain revisited. Exp. Brain Res. 2010, 200, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Blanke, O. Multisensory brain mechanisms of bodily self-consciousness. Nat. Rev. Neurosci. 2012, 13, 556–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanke, O.; Slater, M.; Serino, A. Behavioral, Neural, and Computational Principles of Bodily Self-Consciousness. Neuron 2015, 88, 145–166. [Google Scholar] [CrossRef] [Green Version]
- Legrand, D. The bodily self: The sensori-motor roots of prereflexive self-consciousness. Phenomenol. Cogn. Sci. 2006, 5, 89–118. [Google Scholar] [CrossRef]
- Legrand, D. Subjective and physical dimensions of bodily self-consciousness, and their dis-integration in anorexia nervosa. Neuropsychologia 2010, 48, 726–737. [Google Scholar] [CrossRef]
- Bechara, A.; Damasio, H.; Damasio, A.R. Emotion, decision making and the orbitofrontal cortex. Cereb. Cortex 2000, 10, 295–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benkelfat, C.; Bradwejn, J.; Meyer, E.; Ellenbogen, M.; Milot, S.; Gjedde, A.; Evans, A. Functional neuroanatomy of CCK4-induced anxiety in normal healthy volunteers. Am. J. Psychiatry 1995, 152, 1180–1184. [Google Scholar] [CrossRef] [PubMed]
- Blood, A.J.; Zatorre, R.J. Intensely pleasurable responses to music correlate with activity in brain regions implicated in reward and emotion. Proc. Natl. Acad. Sci. USA 2001, 98, 11818–11823. [Google Scholar] [CrossRef] [Green Version]
- Lindquist, K.A.; Wager, T.D.; Kober, H.; Bliss-Moreau, E.; Barrett, L.F. The brain basis of emotion: A meta-analytic review. Behav. Brain Sci. 2012, 35, 121–143. [Google Scholar] [CrossRef] [Green Version]
- Mayberg, H.S.; Liotti, M.; Brannan, S.K.; McGinnis, S.; Mahurin, R.K.; Jerabek, P.A.; Silva, J.A.; Tekell, J.L.; Martin, C.C.; Lancaster, J.L.; et al. Reciprocal limbic-cortical function and negative mood: Converging PET findings in depression and normal sadness. Am. J. Psychiatry 1999, 156, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Ploghaus, A.; Tracey, I.; Gati, J.S.; Clare, S.; Menon, R.S.; Matthews, P.M.; Rawlins, J.N.P. Dissociating pain from its anticipation in the human brain. Science 1999, 284, 1979–1981. [Google Scholar] [CrossRef] [Green Version]
- Reiman, E.M. The application of positron emission tomography to the study of normal and pathologic emotions. J. Clin. Psychiatry 1997, 58 (Suppl. 16), 4–12. [Google Scholar]
- James, W. The Principles of Psychology; Henry Holt and Company: New York, NY, USA, 1890. [Google Scholar]
- Barrett, L.F.; Quigley, K.S.; Bliss-Moreau, E.; Aronson, K.R. Interoceptive sensitivity and self-reports of emotional experience. J. Pers. Soc. Psychol. 2004, 87, 684–697. [Google Scholar] [CrossRef] [Green Version]
- Damasio, A. Descartes’ Error: Emotion, Reason, and the Human Brain; G.P. Putnam: Putnam, NY, USA, 1993. [Google Scholar]
- Prebble, S.C.; Addis, D.R.; Tippett, L.J. Autobiographical memory and sense of self. Psychol. Bull. 2013, 139, 815–840. [Google Scholar] [CrossRef]
- Damasio, A. Feelings of emotion and the self. Ann. N. Y. Acad. Sci. 2003, 1001, 253–261. [Google Scholar] [CrossRef]
- Harré, R. The Singular Self: An Introduction to the Psychology of Personhood; Sage: Washington, DC, USA, 1998. [Google Scholar]
- Neisser, U. Five kinds of self-knowledge. Philos. Psychol. 1988, 1, 35–59. [Google Scholar] [CrossRef]
- Gallagher, I.I. Philosophical conceptions of the self: Implications for cognitive science. Trends Cogn. Sci. 2000, 4, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, S. A Pattern Theory of Self. Front. Hum. Neurosci. 2013, 7, 443. [Google Scholar] [CrossRef] [Green Version]
- Singer, J.A. Seeing one’s self: Locating narrative memory in a framework of personality. J. Pers. 1995, 63, 429–457. [Google Scholar] [CrossRef]
- Lewis, M. Ways of knowing: Objective self-aareness or consciousness. Dev. Rev. 1991, 11, 231–243. [Google Scholar] [CrossRef]
- Martin, J. Real perspectival selves. Theory Psychol. 2005, 15, 207–224. [Google Scholar] [CrossRef]
- Morin, A.; Everett, J. Inner speech as a mediator of self-awareness, self-consciousness, and self-knowledge: An hypothesis. New Ideas Psychol. 1990, 8, 337–356. [Google Scholar] [CrossRef] [Green Version]
- Carver, C.S. Self-awareness. In Handbook of Self and Identity; Leary, M.R., Tangney, J.P., Eds.; Guilford Press: New York, NY, USA, 2012. [Google Scholar]
- Christoff, K.; Cosmelli, D.; Legrand, D.; Thompson, E. Specifying the self for cognitive neuroscience. Trends Cogn. Sci. 2011, 15, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Vandekerckhove, M. Memory, autonoetic consciousness and the self: Consciousness as a continuum of stages. Self Identity 2009, 8, 4–23. [Google Scholar] [CrossRef]
- Legrand, D. Pre-reflective self-as-subject from experiential and empirical perspectives. Conscious. Cogn. 2007, 16, 583–599. [Google Scholar] [CrossRef] [PubMed]
- Zahavi, A. Is group selection necessary to explain social adaptations in microorganisms? Heredity 2005, 94, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Parvizi, J.; Damasio, A. Consciousness and the brainstem. Cognition 2001, 79, 135–160. [Google Scholar] [CrossRef]
- Keenan, J.P.; Rubio, J.; Racioppi, C.; Johnson, A.; Barnacz, A. The right hemisphere and the dark side of consciousness. Cortex 2005, 41, 695–704. [Google Scholar] [CrossRef]
- Leary, M.R.; Tangney, J.P. Handbook of Self and Identity; Guilford: New York, NY, USA, 2012. [Google Scholar]
- Stuss, D.T.; Levine, B. Adult clinical neuropsychology: Lessons from studies of the frontal lobes. Annu. Rev. Psychol. 2002, 53, 401–433. [Google Scholar] [CrossRef] [PubMed]
- Duval, S.; Wicklund, R.A. Effects of objective self-awareness on attribution of causality. J. Exp. Soc. Psychol. 1973, 9, 17–31. [Google Scholar] [CrossRef]
- Gallup, G.G., Jr. Self-awareness and the emergence of mind in primates. Am. J. Primatol. 1982, 2, 237–248. [Google Scholar] [CrossRef]
- Scheier, M.F. Self-awareness, self-consciousness, and angry aggression. J. Pers. 1976, 44, 627–644. [Google Scholar] [CrossRef]
- Schooler, J.W. Re-representing consciousness: Dissociations between experience and meta-consciousness. Trends Cogn. Sci. 2002, 6, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Stuss, D.T.; Picton, T.W.; Alexander, M.P. Consciousness, self-awareness, and the frontal lobes. In The Frontal Lobes and Neuropsychiatric Illness; Salloway, S.P., Malloy, P.F., Duffy, J.D., Eds.; American Psychiatric: Arlington, VA, USA, 2001. [Google Scholar]
- Northoff, G.; Heinzel, A.; Greck, M.; Bermpohl, F.; Dobrowolny, H.; Panksepp, J. Self-referential processing in our brain—A meta-analysis of imaging studies on the self. Neuroimage 2006, 31, 440–457. [Google Scholar] [CrossRef]
- Conway, M.A. Memory and the self. J. Mem. Lang. 2005, 53, 594–628. [Google Scholar] [CrossRef]
- Conway, M.A.; Singer, J.A.; Tagini, A. The self and autobiographical memory: Correspondence and coherence. Soc. Cogn. 2004, 22, 491–529. [Google Scholar] [CrossRef] [Green Version]
- Addis, D.R.; McIntosh, A.R.; Moscovitch, M.; Crawley, A.P.; McAndrews, M.P. Characterizing spatial and temporal features of autobiographical memory retrieval networks: A partial least squares approach. Neuroimage 2004, 23, 1460–1471. [Google Scholar] [CrossRef] [PubMed]
- Addis, D.R.; Tippett, L.J. The contributions of autobiographical memory to the content and continuity of self: A social-cognitive neuro-science approach. In Self-Continuity: Individual and Collective Perspectives; Fabio, S., Ed.; Psychology Press: New York, NY, USA, 2008; pp. 71–84. [Google Scholar]
- Kihlstrom, J.F.; Beer, J.S.; Klein, S.B. Self and identity as memory. In Handbook of Self and Identity; Leary, M.R., Tangney, J.P., Eds.; Guilford Press: New York, NY, USA, 2003; pp. 68–90. [Google Scholar]
- Klein, S.B.; Loftus, J. The mental representation of trait and autobiographical knowledge about the self. In Advances in Social Cognition; Srull, T.K., Wyer, R.S., Eds.; Erlbaum: Hillsdale, NJ, USA, 1993; Volume 5. [Google Scholar]
- Rathbone, C.J.; Moulin, C.J.; Conway, M.A. Self-centered memories: The reminiscence bump and the self. Mem. Cognit. 2008, 36, 1403–1414. [Google Scholar] [CrossRef]
- Wilson, A.E.; Ross, M. The identity function of autobiographical memory: Time is on our side. Memory 2003, 11, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Damasio, A.; Meyer, K. Consciousness: An Overview of the Phenomenon and of Its Possible Neural Basis. In The Neurology of Consciousness; Academic Press: Cambridge, MA, USA, 2009; pp. 3–14. [Google Scholar] [CrossRef]
- Demiray, B.; Gulgoz, S.; Bluck, S. Examining the life story account of the reminiscence bump: Why we remember more from young adulthood. Memory 2009, 17, 708–723. [Google Scholar] [CrossRef]
- Gluck, J.; Bluck, S. Looking back across the life span: A life story account of the reminiscence bump. Mem. Cognit. 2007, 35, 1928–1939. [Google Scholar] [CrossRef] [Green Version]
- Janssen, S.M.; Murre, J.M. Reminiscence bump in autobiographical memory: Unexplained by novelty, emotionality, valence, or importance of personal events. Q. J. Exp. Psychol. 2008, 61, 1847–1860. [Google Scholar] [CrossRef]
- Kihlstrom, J.F.; Klein, S.B. Self-knowledge and self-awareness. Ann. N. Y. Acad. Sci. 1997, 818, 4–17. [Google Scholar] [CrossRef]
- Klein, S.B.; Cosmides, L.; Costabile, K.A.; Mei, L. Is there something special about the self? A neuropsychological case study. J. Res. Personal. 2002, 36, 490–506. [Google Scholar] [CrossRef]
- Klein, S.B.; Lax, M.L. The unanticipated resilience of trait self-knowledge in the face of neural damage. Memory 2010, 18, 918–948. [Google Scholar] [CrossRef]
- Klein, S.B.; Loftus, J.; Kihlstrom, J.F. Self-knowledge of an amnesic patient: Toward a neuropsychology of personality and social psychology. J. Exp. Psychol. Gen. 1996, 125, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Rathbone, C.J.; Moulin, C.J.; Conway, M.A. Autobiographical memory and amnesia: Using conceptual knowledge to ground the self. Neurocase 2009, 15, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Tulving, E. Self-knowledge of an amnesic individual is represented abstractly. In Advances in Social Cognition; Srull, T.K., Wyer, R.S., Eds.; Erlbaum: Hillsdale, NJ, USA, 1993; Volume 5. [Google Scholar]
- Tanweer, T.; Rathbone, C.J.; Souchay, C. Autobiographical memory, autonoetic consciousness, and identity in Asperger syndrome. Neuropsychologia 2010, 48, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Brewer, W.F. What is autobiographical memory? In Autobiographical Memory; Rubin, D.C., Ed.; Cambridge University Press: New York, NY, USA, 1986. [Google Scholar]
- Martinelli, P.; Sperduti, M.; Piolino, P. Neural substrates of the self-memory system: New insights from a meta-analysis. Hum. Brain Mapp. 2013, 34, 1515–1529. [Google Scholar] [CrossRef]
- Modinos, G.; Ormel, J.; Aleman, A. Activation of anterior insula during self-reflection. PLoS ONE 2009, 4, 4618. [Google Scholar] [CrossRef] [Green Version]
- Craik, F.I.; Moroz, T.M.; Moscovitch, M.; Stuss, D.T.; Winocur, G.; Tulving, E.; Kapur, S. In search of the self: A positron emission tomography study. Psychol. Sci. 1999, 10, 26–34. [Google Scholar] [CrossRef]
- D’Argembeau, A.; Feyers, D.; Majerus, S.; Collette, F.; Van der Linden, M.; Maquet, P.; Salmon, E. Self-reflection across time: Cortical midline structures differentiate between present and past selves. Soc. Cogn. Affect. Neurosci. 2008, 3, 244–252. [Google Scholar] [CrossRef] [Green Version]
- D’Argembeau, A.; Ruby, P.; Collette, F.; Degueldre, C.; Balteau, E.; Luxen, A.; Maquet, P.; Salmon, E. Distinct regions of the medial prefrontal cortex are associated with self-referential processing and perspective taking. J. Cogn. Neurosci. 2007, 19, 935–944. [Google Scholar] [CrossRef]
- D’Argembeau, A.; Linden, M. Phenomenal characteristics associated with projecting oneself back into the past and forward into the future: Influence of valence and temporal distance. Conscious. Cogn. 2004, 13, 844–858. [Google Scholar] [CrossRef] [Green Version]
- D’Argembeau, A.; Linden, M. Influence of emotion on memory for temporal information. Emotion 2005, 5, 503–507. [Google Scholar] [CrossRef] [Green Version]
- Fossati, P.; Hevenor, S.J.; Lepage, M.; Graham, S.J.; Grady, C.; Keightley, M.L.; Craik, F.; Mayberg, H. Distributed self in episodic memory: Neural correlates of successful retrieval of self-encoded positive and negative personality traits. Neuroimage 2004, 22, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- Gutchess, A.H.; Kensinger, E.A.; Schacter, D.L. Aging, self-referencing, and medial prefrontal cortex. Soc. Neurosci. 2007, 2, 117–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heatherton, T.F.; Wyland, C.L.; Macrae, C.N.; Demos, K.E.; Denny, B.T.; Kelley, W.M. Medial prefrontal activity differentiates self from close others. Soc. Cogn. Affect. Neurosci. 2006, 1, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, A.C.; Macrae, C.N.; Mitchell, J.P. Repetition suppression of ventromedial prefrontal activity during judgments of self and others. Proc. Natl. Acad. Sci. USA 2008, 105, 4507–4512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, H.C.; Luber, B.; Crupain, M.; Keenan, J.P.; Nowak, M.; Kjaer, T.W.; Sackeim, H.A.; Lisanby, S.H. Parietal cortex and representation of the mental Self. Proc. Natl. Acad. Sci. USA 2004, 101, 6827–6832. [Google Scholar] [CrossRef] [Green Version]
- Seger, C.A.; Stone, M.; Keenan, J.P. Cortical Activations during judgments about the self and an other person. Neuropsychologia 2004, 42, 1168–1177. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.J.; Schaer, M.; Debbane, M. Degrees of separation: A quantitative neuroimaging meta-analysis investigating self-specificity and shared neural activation between self- and other-reflection. Neurosci. Biobehav. Rev. 2012, 36, 1043–1059. [Google Scholar] [CrossRef]
- Meer, L.; Costafreda, S.; Aleman, A.; David, A. Self-reflection and the brain: A theoretical review and meta-analysis of neuroimaging studies with implications for schizophrenia. Neurosci. Biobehav. Rev. 2010, 34, 935–946. [Google Scholar] [CrossRef]
- Tulving, E. Memory and consciousness. Can. Psychol. 1985, 26, 1–12. [Google Scholar] [CrossRef]
- Tulving, E. Episodic memory: From mind to brain. Annu. Rev. Psychol. 2002, 53, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Tulving, E. Episodic memory and autonoesis: Uniquely human? In The Missing Link in Cognition: Origins of Self-Reflective Consciousness; Terrace, H.S., Metcalfe, J., Eds.; Oxford University Press: New York, NY, USA, 2005; pp. 3–56. [Google Scholar]
- Levine, B.; Svoboda, E.; Hay, J.F.; Winocur, G.; Moscovitch, M. Aging and autobiographical memory: Dissociating episodic from semantic retrieval. Psychol. Aging 2002, 17, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Markowitsch, H.J. Autonoetic consciousness. In The Self in Neuroscience and Psychiatry; Kircher, T., David, A., Eds.; Cambridge University Press: New York, NY, USA, 2003; pp. 180–196. [Google Scholar]
- Butterworth, G. Origins of Self-Perception in Infancy. Psychol. Inq. 1992, 3, 103–111. [Google Scholar] [CrossRef]
- Howe, M.L.; Courage, M.L. The emergence and early development of autobiographical memory. Psychol. Rev. 1997, 104, 499–523. [Google Scholar] [CrossRef] [PubMed]
- Irish, M.; Addis, D.R.; Hodges, J.R.; Piguet, O. Considering the role of semantic memory in episodic future thinking: Evidence from semantic dementia. Brain 2012, 135, 2178–2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAdams, D.P. Personal narratives and the life story. In Handbook of Personality: Theory and Research; John, O.P., Robins, R.W., Pervin, L.A., Eds.; Guilford Press: New York, NY, USA, 2008; pp. 242–262. [Google Scholar]
- Pasupathi, M.; Mansour, E.; Brubaker, J.R. Developing a Life Story: Constructing Relations between Self and Experience in Autobiographical Narratives. Hum. Dev. 2007, 50, 85–110. [Google Scholar] [CrossRef]
- Botzung, A.; Denkova, E.; Manning, L. Experiencing past and future personal events: Functional neuroimaging evidence on the neural bases of mental time travel. Brain Cogn. 2008, 66, 202–212. [Google Scholar] [CrossRef]
- Cabeza, R.; Prince, S.E.; Daselaar, S.M.; Greenberg, D.L.; Budde, M.; Dolcos, F.; LaBar, K.S.; Rubin, D.C. Brain activity during episodic retrieval of autobiographical and laboratory events: An fMRI study using a novel photo paradigm. J. Cogn. Neurosci. 2004, 16, 1583–1594. [Google Scholar] [CrossRef] [Green Version]
- Gilboa, A. Autobiographical and episodic memory–one and the same? Evidence from prefrontal activation in neuroimaging studies. Neuropsychologia 2004, 42, 1336–1349. [Google Scholar] [CrossRef]
- Levine, B. Autobiographical memory and the self in time: Brain lesion effects, functional neuroanatomy, and lifespan development. Brain Cogn. 2004, 55, 54–68. [Google Scholar] [CrossRef]
- Viard, A.; Piolino, P.; Desgranges, B.; Chételat, G.; Lebreton, K.; Landeau, B.; Young, A.; De La Sayette, V.; Eustache, F. Hippocampal activation for autobiographical memories over the entire lifetime in healthy aged subjects: An fMRI study. Cereb. Cortex 2007, 17, 2453–2467. [Google Scholar] [CrossRef] [Green Version]
- Conway, M.A.; Turk, D.J.; Miller, S.L.; Logan, J.; Nebes, R.D.; Meltzer, C.C.; Becker, J.T. A positron emission tomography (PET) study of autobiographical memory retrieval. Memory 1999, 7, 679–702. [Google Scholar] [CrossRef] [PubMed]
- Gilboa, A.; Ramirez, J.; Kohler, S.; Westmacott, R.; Black, S.E.; Moscovitch, M. Retrieval of autobiographical memory in Alzheimer’s disease: Relation to volumes of medial temporal lobe and other structures. Hippocampus 2005, 15, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Maguire, E.A.; Frith, C.D. Aging affects the engagement of the hippocampus during autobiographical memory retrieval. Brain 2003, 126, 1511–1523. [Google Scholar] [CrossRef] [Green Version]
- Philippi, N.; Noblet, V.; Botzung, A.; Després, O.; Renard, F.; Sfikas, G.; Cretin, B.; Kremer, S.; Manning, L.; Blanc, F. MRI-based volumetry correlates of autobiographical memory in Alzheimer’s disease. PLoS ONE 2012, 7, 46200. [Google Scholar] [CrossRef] [Green Version]
- Philippi, N.; Rousseau, F.; Noblet, V.; Botzung, A.; Després, O.; Cretin, B.; Kremer, S.; Blanc, F.; Manning, L. Different Temporal Patterns of Specific and General Autobiographical Memories across the Lifespan in Alzheimer’s Disease. Behav. Neurol. 2015, 2015, 963460. [Google Scholar] [CrossRef]
- Piolino, P.; Desgranges, B.; Benali, K.; Eustache, F. Episodic and semantic remote autobiographical memory in ageing. Memory 2002, 10, 239–257. [Google Scholar] [CrossRef]
- Ring, C.; Brener, J. Heartbeat counting is unrelated to heartbeat detection: A comparison of methods to quantify interoception. Psychophysiology 2018, 55, e13084. [Google Scholar] [CrossRef] [Green Version]
- Schandry, R. Heart beat perception and emotional experience. Psychophysiology 1981, 18, 483–488. [Google Scholar] [CrossRef]
- Schulz, S.M. Neural correlates of heart-focused interoception: A functional magnetic resonance imaging meta-analysis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20160018. [Google Scholar] [CrossRef] [Green Version]
- Stephan, E.; Pardo, J.V.; Faris, P.L.; Hartman, B.K.; Kim, S.W.; Ivanov, E.H.; Daughters, R.S.; Costello, P.A.; Goodale, R.L. Functional neuroimaging of gastric distention. J. Gastrointest. Surg. 2003, 7, 740–749. [Google Scholar] [CrossRef]
- Tsakiris, M.; Hesse, M.D.; Boy, C.; Haggard, P.; Fink, G.R. Neural signatures of body ownership: A sensory network for bodily self-consciousness. Cereb. Cortex 2007, 17, 2235–2244. [Google Scholar] [CrossRef]
- Farrer, C.; Franck, N.; Georgieff, N.; Frith, C.D.; Decety, J.; Jeannerod, M. Modulating the experience of agency: A positron emission tomography study. NeuroImage 2003, 18, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Devue, C.; Collette, F.; Balteau, E.; Degueldre, C.; Luxen, A.; Maquet, P.; Brédart, S. Here I am: The cortical correlates of visual self-recognition. Brain Res. 2007, 1143, 169–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kircher, T.T.; Senior, C.; Phillips, M.L.; Benson, P.J.; Bullmore, E.T.; Brammer, M.; Simmons, A.; Williams, S.C.; Bartels, M.; David, A.S. Towards a functional neuroanatomy of self processing: Effects of faces and words. Brain Res. Cogn. Brain Res. 2000, 10, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Gusnard, D.A.; Akbudak, E.; Shulman, G.L.; Raichle, M.E. Medial prefrontal cortex and self-referential mental activity: Relation to a default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 4259–4264. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M.L.; Gregory, L.J.; Cullen, S.; Cohen, S.; Ng, V.; Andrew, C.; Giampietro, V.; Bullmore, E.; Zelaya, F.; Amaro, E.; et al. The effect of negative emotional context on neural and behavioural responses to oesophageal stimulation. Brain 2003, 126, 669–684. [Google Scholar] [CrossRef] [Green Version]
- Crossley, N.A.; Scott, J.; Ellison-Wright, I.; Mechelli, A. Neuroimaging distinction between neurological and psychiatric disorders. Br. J. Psychiatry 2015, 207, 429–434. [Google Scholar] [CrossRef] [Green Version]
- Goodkind, M.; Eickhoff, S.B.; Oathes, D.; Jiang, Y.; Chang, A.; Jones-Hagata, L.B.; Ortega, B.N.; Zaiko, Y.V.; Roach, E.L.; Korgaonkar, M.; et al. Identification of a common neurobiological substrate for mental illness. JAMA Psychiatry 2015, 72, 305–315. [Google Scholar] [CrossRef]
- Kaye, W.H.; Wierenga, C.E.; Bischoff-Grethe, A.; Berner, L.A.; Ely, A.V.; Bailer, U.F.; Paulus, M.P.; Fudge, J.L. Neural Insensitivity to the Effects of Hunger in Women Remitted From Anorexia Nervosa. AJP Am. J. Psychiatry 2020, 177, 601–610. [Google Scholar] [CrossRef]
- Naqvi, N.H.; Rudrauf, D.; Damasio, H.; Bechara, A. Damage to the Insula Disrupts Addiction to Cigarette Smoking. Science 2007, 315, 531–534. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, N.H.; Bechara, A. The hidden island of addiction: The insula. Trends Neurosci. 2009, 32, 56–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borgwardt, S.J.; Riecher-Rössler, A.; Dazzan, P.; Chitnis, X.; Aston, J.; Drewe, M.; Gschwandtner, U.; Haller, S.; Pflüger, M.; Rechsteiner, E.; et al. Regional gray matter volume abnormalities in the at risk mental state. Biol. Psychiatry 2007, 61, 1148–1156. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Wood, S.J.; Yung, A.R.; Phillips, L.J.; Soulsby, B.; McGorry, P.D.; Tanino, R.; Zhou, S.-Y.; Suzuki, M.; Velakoulis, D.; et al. Insular cortex gray matter changes in individuals at ultra-high-risk of developing psychosis. Schizophr. Res. 2009, 111, 94–102. [Google Scholar] [CrossRef]
- Duggal, H.S.; Muddasani, S.; Keshavan, M.S. Insular volumes in first-episode schizophrenia: Gender effect. Schizophr. Res. 2005, 73, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Shapleske, J.; Rossell, S.L.; Chitnis, X.A.; Suckling, J.; Simmons, A.; Bullmore, E.T.; Woodruff, P.W.; David, A.S. A computational morphometric MRI study of schizophrenia: Effects of hallucinations. Cereb. Cortex 2002, 12, 1331–1341. [Google Scholar] [CrossRef] [Green Version]
- Sigmundsson, T.; Suckling, J.; Maier, M.; Williams, S.; Bullmore, E.; Greenwood, K.; Fukuda, R.; Ron, M.A.; Toone, B.K. Structural Abnormalities in Frontal, Temporal, and Limbic Regions and Interconnecting White Matter Tracts in Schizophrenic Patients With Prominent Negative Symptoms. AJP Am. J. Psychiatry 2001, 158, 234–243. [Google Scholar] [CrossRef] [Green Version]
- Wylie, K.P.; Tregellas, J.R. The role of the insula in schizophrenia. Schizophr. Res. 2010, 123, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baier, B.; Karnath, H.O. Tight link between our sense of limb ownership and self-awareness of actions. Stroke 2008, 39, 486–488. [Google Scholar] [CrossRef] [Green Version]
- Bediou, B.; Franck, N.; Saoud, M.; Baudouin, J.-Y.; Tiberghien, G.; Daléry, J.; D’Amato, T. Effects of emotion and identity on facial affect processing in schizophrenia. Psychiatry Res. 2005, 133, 149–157. [Google Scholar] [CrossRef]
- Carr, L.; Iacoboni, M.; Dubeau, M.C.; Mazziotta, J.C.; Lenzi, G.L. Neural mechanisms of empathy in humans: A relay from neural systems for imitation to limbic areas. Proc. Natl. Acad. Sci. USA 2003, 100, 5497–5502. [Google Scholar] [CrossRef] [Green Version]
- Schneider, F.; Gur, R.C.; Koch, K.; Backes, V.; Amunts, K.; Shah, N.J.; Bilker, W.; Gur, R.E.; Habel, U. Impairment in the specificity of emotion processing in schizophrenia. Am. J. Psychiatry 2006, 163, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Fathy, Y.Y.; Hoogers, S.E.; Berendse, H.W.; van der Werf, Y.D.; Visser, P.J.; de Jong, F.J.; van de Berg, W.D. Differential insular cortex sub-regional atrophy in neurodegenerative diseases: A systematic review and meta-analysis. Brain Imaging Behav. 2020, 14, 2799–2816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Cordero, I.; Sedeño, L.; de la Fuente, L.; Slachevsky, A.; Forno, G.; Klein, F.; Lillo, P.; Ferrari, J.; Rodriguez, C.; Bustin, J.; et al. Feeling, learning from and being aware of inner states: Interoceptive dimensions in neurodegeneration and stroke. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20160006. [Google Scholar] [CrossRef] [Green Version]
- Santangelo, G.; Vitale, C.; Baiano, C.; D’Iorio, A.; Longo, K.; Barone, P.; Amboni, M.; Conson, M. Interoceptive processing deficit: A behavioral marker for subtyping Parkinson’s disease. Park. Relat. Disord. 2018, 53, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Philippi, N.; Roquet, D.; Ben Malek, H.; Noblet, V.; Botzung, A.; Cretin, B.; Blanc, F. Henry, where have you lost your Self? Cortex 2017, 95, 37–50. [Google Scholar] [CrossRef]
- Cosentino, S.; Brickman, A.M.; Griffith, E.; Habeck, C.; Cines, S.; Farrell, M.; Shaked, D.; Huey, E.D.; Briner, T.; Stern, Y. The right insula contributes to memory awareness in cognitively diverse older adults. Neuropsychologia 2015, 75, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Seeley, W.W. Anterior insula degeneration in frontotemporal dementia. Brain Struct. Funct. 2010, 214, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Silani, G.; Bird, G.; Brindley, R.; Singer, T.; Frith, C.; Frith, U. Levels of emotional awareness and autism: An fMRI study. Soc. Neurosci. 2008, 3, 97–112. [Google Scholar] [CrossRef] [Green Version]
- Hogeveen, J.; Bird, G.; Chau, A.; Krueger, F.; Grafman, J. Acquired alexithymia following damage to the anterior insula. Neuropsychologia 2016, 82, 142–148. [Google Scholar] [CrossRef] [Green Version]
- Hornberger, M.; Yew, B.; Gilardoni, S.; Mioshi, E.; Gleichgerrcht, E.; Manes, F.; Hodges, J.R. Ventromedial-frontopolar prefrontal cortex atrophy correlates with insight loss in frontotemporal dementia and Alzheimer’s disease. Hum. Brain Mapp. 2014, 35, 616–626. [Google Scholar] [CrossRef]
- Farrer, C.; Frith, C.D. Experiencing oneself vs another person as being the cause of an action: The neural correlates of the experience of agency. Neuroimage 2002, 15, 596–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruby, P.; Decety, J. Effect of subjective perspective taking during simulation of action: A PET investigation of agency. Nat. Neurosci. 2001, 4, 546–550. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Matsuura, M.; Koeda, M.; Yahata, N.; Suhara, T.; Kato, M.; Okubo, Y. Brain activations during judgments of positive self-conscious emotion and positive basic emotion: Pride and joy. Cereb. Cortex 2008, 18, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Finlayson-Short, L.; Davey, C.G.; Harrison, B.J. Neural correlates of integrated self and social processing. Soc. Cogn. Affect. Neurosci. 2020, 15, 941–949. [Google Scholar] [CrossRef]
- Perini, I.; Gustafsson, P.A.; Hamilton, J.P.; Kampe, R.; Zetterqvist, M.; Heilig, M. The salience of self, not social pain, is encoded by dorsal anterior cingulate and insula. Sci. Rep. 2018, 8, 6165. [Google Scholar] [CrossRef] [Green Version]
- Suharaab, T.; Yasunoab, F.; Sudoab, Y.; Yamamotoabc, M.; Inoueab, M.; Okuboabc, Y.; Suzukiab, K. Dopamine D2 receptors in the insular cortex and the personality trait of novelty seeking. Neuroimage 2001, 13, 891–895. [Google Scholar] [CrossRef]
- Johnson, D.L.; Wiebe, J.S.; Gold, S.M.; Andreasen, N.C.; Hichwa, R.D.; Watkins, G.L.; Ponto, L.L.B. Cerebral blood flow and personality: A positron emission tomography study. Am. J. Psychiatry 1999, 156, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.D. Self-esteem and clarity of the self-concept. J. Pers. Soc. Psychol. 1990, 59, 538–549. [Google Scholar] [CrossRef]
- Campbell, J.D.; Trapnell, P.D.; Heine, S.J.; Katz, I.M.; Lavallee, L.F.; Lehman, D.R. Self-concept clarity: Measurement, personality correlates, and cultural boundaries. J. Pers. Soc. Psychol. 1996, 70, 141–156. [Google Scholar] [CrossRef]
- Cicero, D.C.; Martin, E.A.; Becker, T.M.; Kerns, J.G. Decreased Self-Concept Clarity in People with Schizophrenia. J. Nerv. Ment. Dis. 2016, 204, 142–147. [Google Scholar] [CrossRef] [Green Version]
- Uddin, L.Q.; Menon, V. The anterior insula in autism: Under-connected and under-examined. Neurosci. Biobehav. Rev. 2009, 33, 1198–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capps, L.; Sigman, M.; Yirmiya, N. Self-competence and emotional understanding in high-functioning children with autism. Dev. Psychopathol. 1995, 7, 137–149. [Google Scholar] [CrossRef]
- Williamson, S.; Craig, J.; Slinger, R. Exploring the relationship between measures of self-esteem and psychological adjustment among adolescents with Asperger Syndrome. Autism 2008, 12, 391–402. [Google Scholar] [CrossRef]
- Naylor, E.; Clare, L. Awareness of memory functioning, autobiographical memory and identity in early-stage dementia. Neuropsychol. Rehabil. 2008, 18, 590–606. [Google Scholar] [CrossRef] [PubMed]
- Tisserand, A.; Noblet, V.; Botzung, A.; Blanc, F.; Philippi, N. Who am I with my Lewy bodies? A self-concept study: Neuropsychiatry and behavioral neurology/Dementia. Alzheimer’s Dement. 2020, 16, e042898. [Google Scholar] [CrossRef]
- Philippi, N.; Noblet, V.; Hamdaoui, M.; Soulier, D.; Botzung, A.; Ehrhard, E.; Cretin, B.; Blanc, F. The insula, a grey matter of tastes: A volumetric MRI study in dementia with Lewy bodies. Alzheimers Res. Ther. 2020, 12, 79. [Google Scholar] [CrossRef]
- Soulier, D.; Philippi, N.; Marescaux, C.; Wolff, V. Interoceptive, intuitive, and emotional dysfunction after right insular infarction: Three manifestations of the same syndrome? Rev. Neuropsychol. 2019, 11, 227–235. [Google Scholar]
- Svoboda, E.; McKinnon, M.C.; Levine, B. The functional neuroanatomy of autobiographical memory: A meta-analysis. Neuropsychologia 2006, 44, 2189–2208. [Google Scholar] [CrossRef] [Green Version]
- Berna, F.; Potheegadoo, J.; Aouadi, I.; Ricarte, J.J.; Allé, M.C.; Coutelle, R.; Boyer, L.; Cuervo-Lombard, C.V.; Danion, J.-M. A Meta-Analysis of Autobiographical Memory Studies in Schizophrenia Spectrum Disorder. Schizophr. Bull. 2016, 42, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Wantzen, P.; Boursette, A.; Zante, E.; Mioche, J.; Eustache, F.; Guénolé, F.; Baleyte, J.-M.; Guillery-Girard, B. Autobiographical Memory and Social Identity in Autism: Preliminary Results of Social Positioning and Cognitive Intervention. Front. Psychol. 2021, 12, 641765. [Google Scholar] [CrossRef]
- Roquet, D.; Noblet, V.; Anthony, P.; Philippi, N.; Demuynck, C.; Cretin, B.; Martin-Hunyadi, C.; de Sousa, P.L.; Blanc, F. Insular atrophy at the prodromal stage of dementia with Lewy bodies: A VBM DARTEL study. Sci. Rep. 2017, 7, 9437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, F.; Colloby, S.J.; Philippi, N.; De Pétigny, X.; Jung, B.; Demuynck, C.; Phillipps, C.; Anthony, P.; Thomas, A.; Bing, F.; et al. Cortical Thickness in Dementia with Lewy Bodies and Alzheimer’s Disease: A Comparison of Prodromal and Dementia Stages. PLoS ONE 2015, 10, e0127396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehrlach, D.A.; Dolensek, N.; Klein, A.S.; Chowdhury, R.R.; Matthys, A.; Junghänel, M.; Gaitanos, T.N.; Podgornik, A.; Black, T.D.; Vaka, N.R.; et al. Aversive state processing in the posterior insular cortex. Nat. Neurosci. 2019, 22, 1424–1437. [Google Scholar] [CrossRef] [PubMed]
- Livneh, I.; Moshitch-Moshkovitz, S.; Amariglio, N.; Rechavi, G.; Dominissini, D. The m6A epitranscriptome: Transcriptome plasticity in brain development and function. Nat. Rev. Neurosci. 2020, 21, 36–51. [Google Scholar] [CrossRef]
- Vincis, R.; Chen, K.; Czarnecki, L.; Chen, J.; Fontanini, A. Dynamic Representation of Taste-Related Decisions in the Gustatory Insular Cortex of Mice. Curr. Biol. 2020, 30, 1834–1844.e5. [Google Scholar] [CrossRef]
- Yiannakas, A.; Kolatt Chandran, S.; Kayyal, H.; Gould, N.; Khamaisy, M.; Rosenblum, K. Parvalbumin interneuron inhibition onto anterior insula neurons projecting to the basolateral amygdala drives aversive taste memory retrieval. Curr. Biol. 2021, 31, 2770–2784.e6. [Google Scholar] [CrossRef]
- Schiff, H.C.; Kogan, J.F.; Isaac, M.; Czarnecki, L.A.; Fontanini, A.; Maffei, A. Experience-dependent plasticity of gustatory insular cortex circuits and taste preferences. Sci. Adv. 2023, 9, eade6561. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tisserand, A.; Philippi, N.; Botzung, A.; Blanc, F. Me, Myself and My Insula: An Oasis in the Forefront of Self-Consciousness. Biology 2023, 12, 599. https://doi.org/10.3390/biology12040599
Tisserand A, Philippi N, Botzung A, Blanc F. Me, Myself and My Insula: An Oasis in the Forefront of Self-Consciousness. Biology. 2023; 12(4):599. https://doi.org/10.3390/biology12040599
Chicago/Turabian StyleTisserand, Alice, Nathalie Philippi, Anne Botzung, and Frédéric Blanc. 2023. "Me, Myself and My Insula: An Oasis in the Forefront of Self-Consciousness" Biology 12, no. 4: 599. https://doi.org/10.3390/biology12040599