
Ingestion of Soybean Sprouts Containing a HASPIN Inhibitor Improves Condition in a Mouse Model of Alzheimer’s Disease
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. In Vitro Kinase Assay
2.3. Hippocampal Slice Experiment
2.4. Western Blot Analysis
2.5. Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
2.6. Transfection
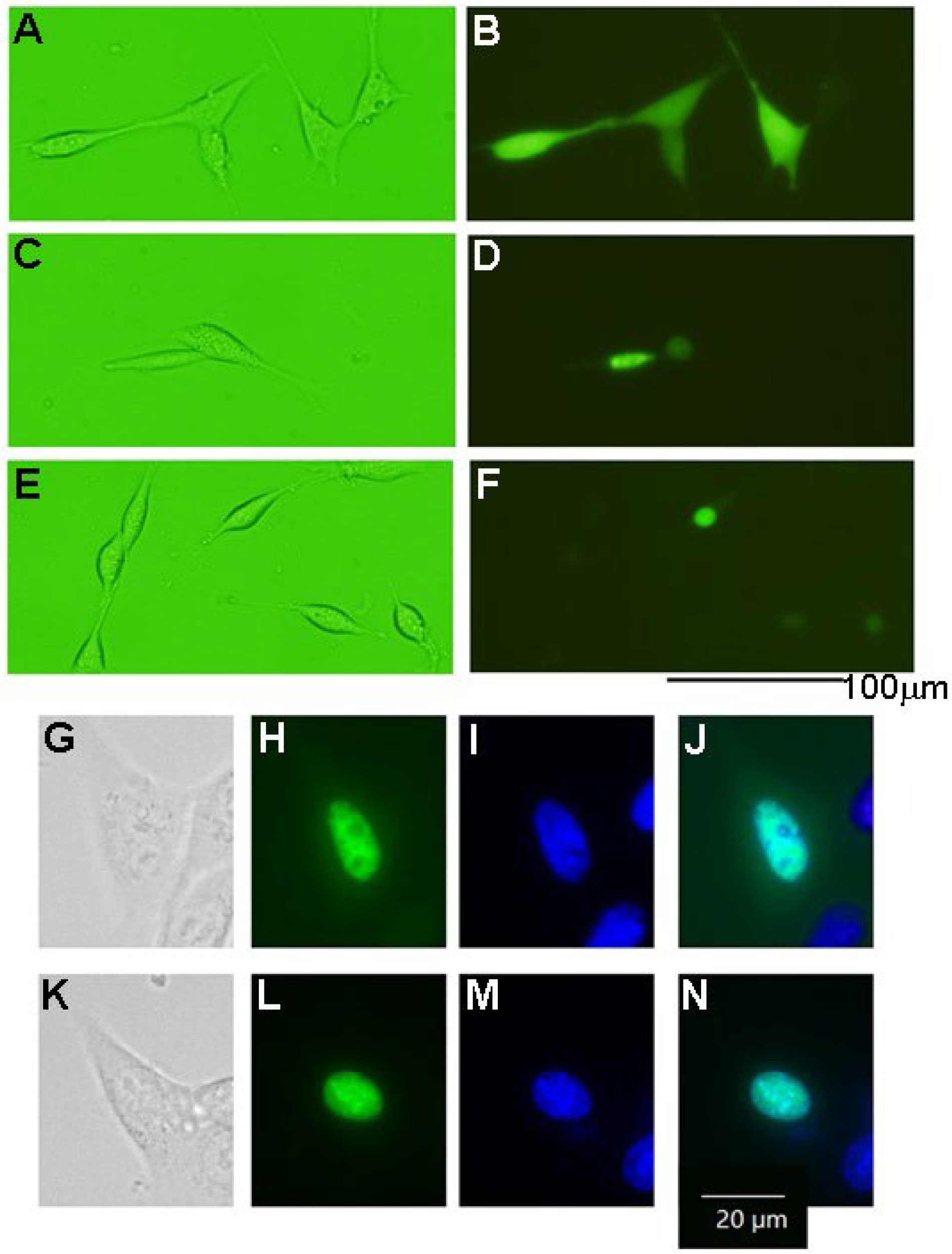
2.7. Microscopic Observations
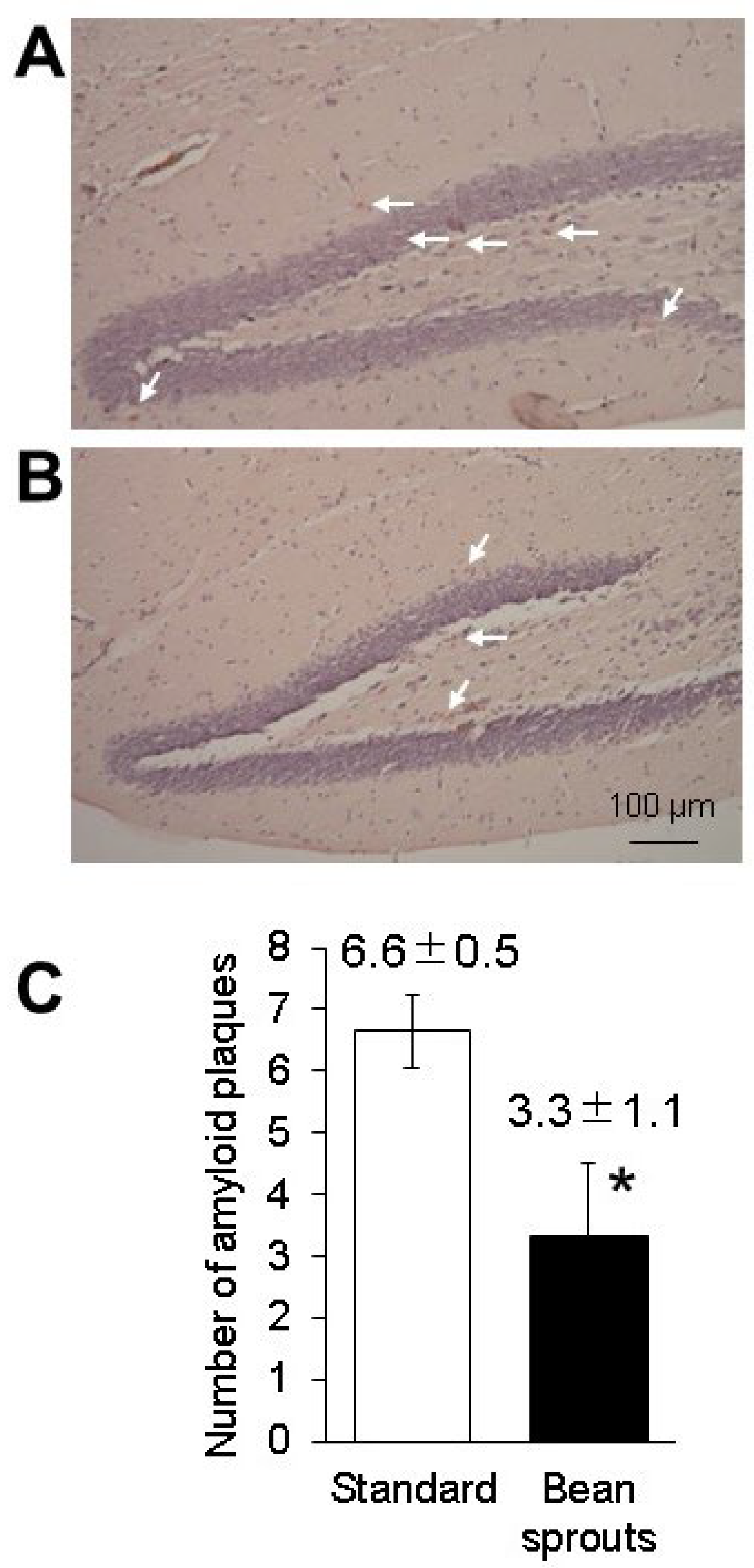
2.8. Congo Red Staining
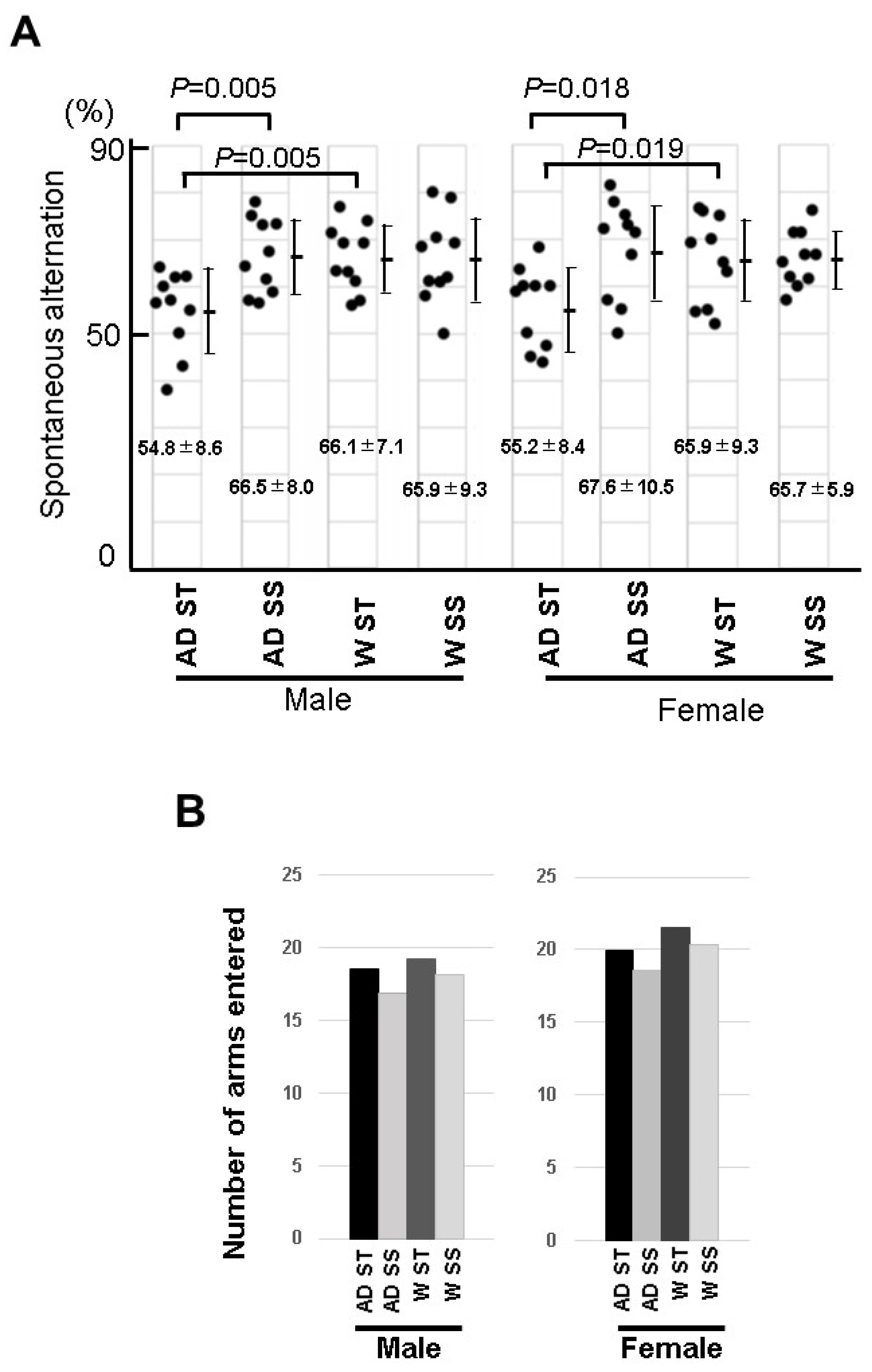
2.9. Y-Maze Test
2.10. Statistical Analysis
3. Results
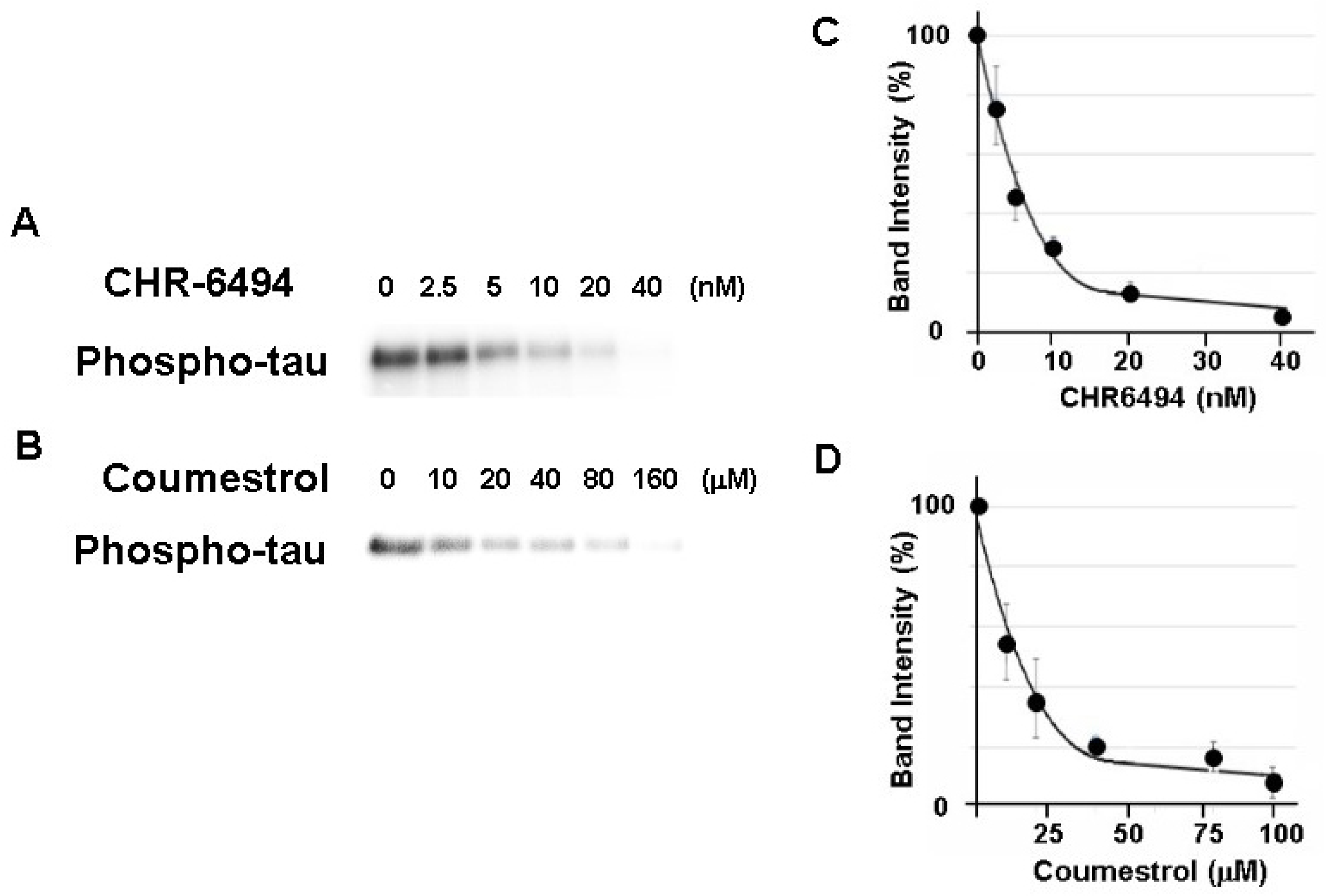
3.1. Phosphorylation of the Tau Protein by HASPIN
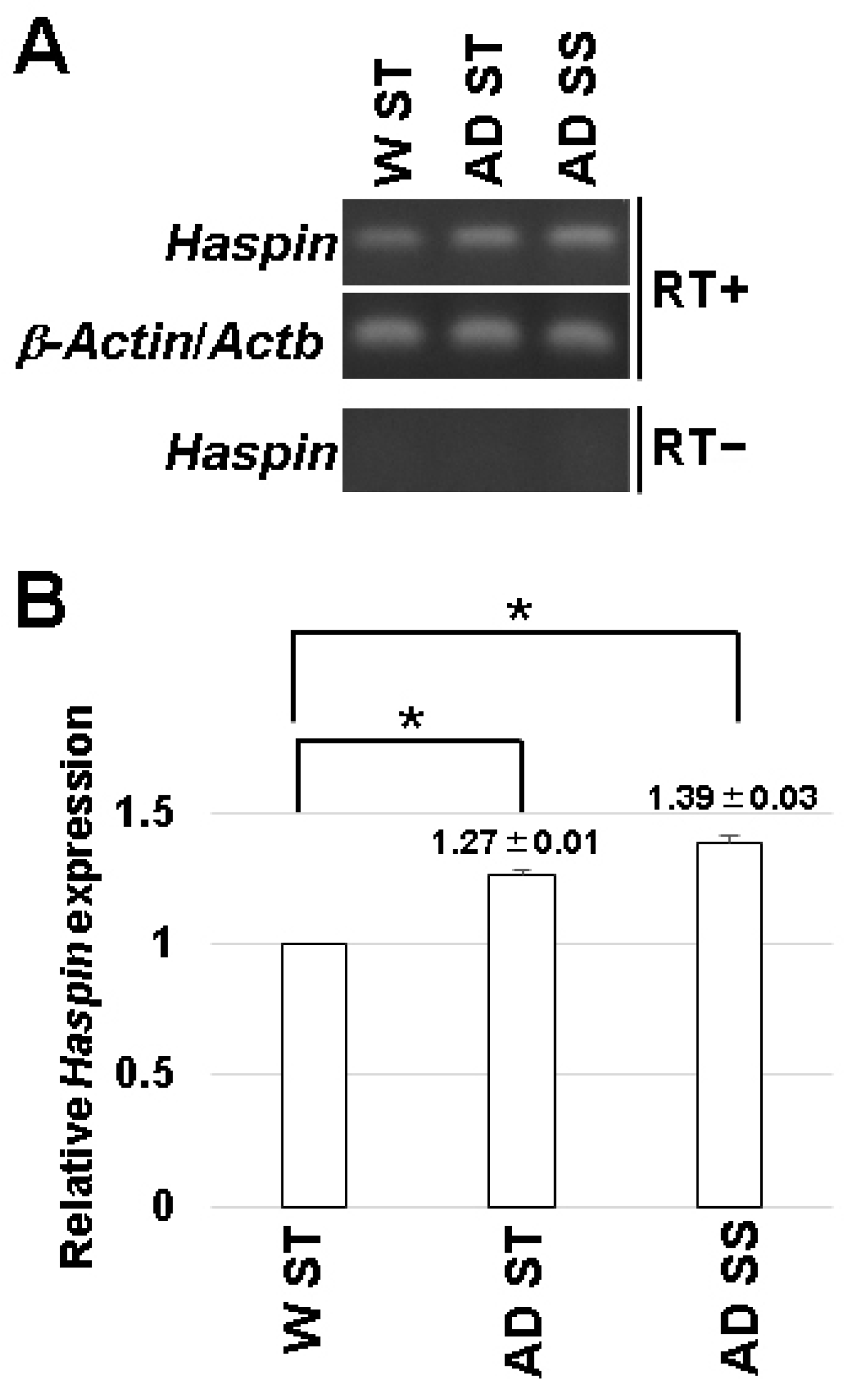
3.2. Expression of HASPIN in the Hippocampus
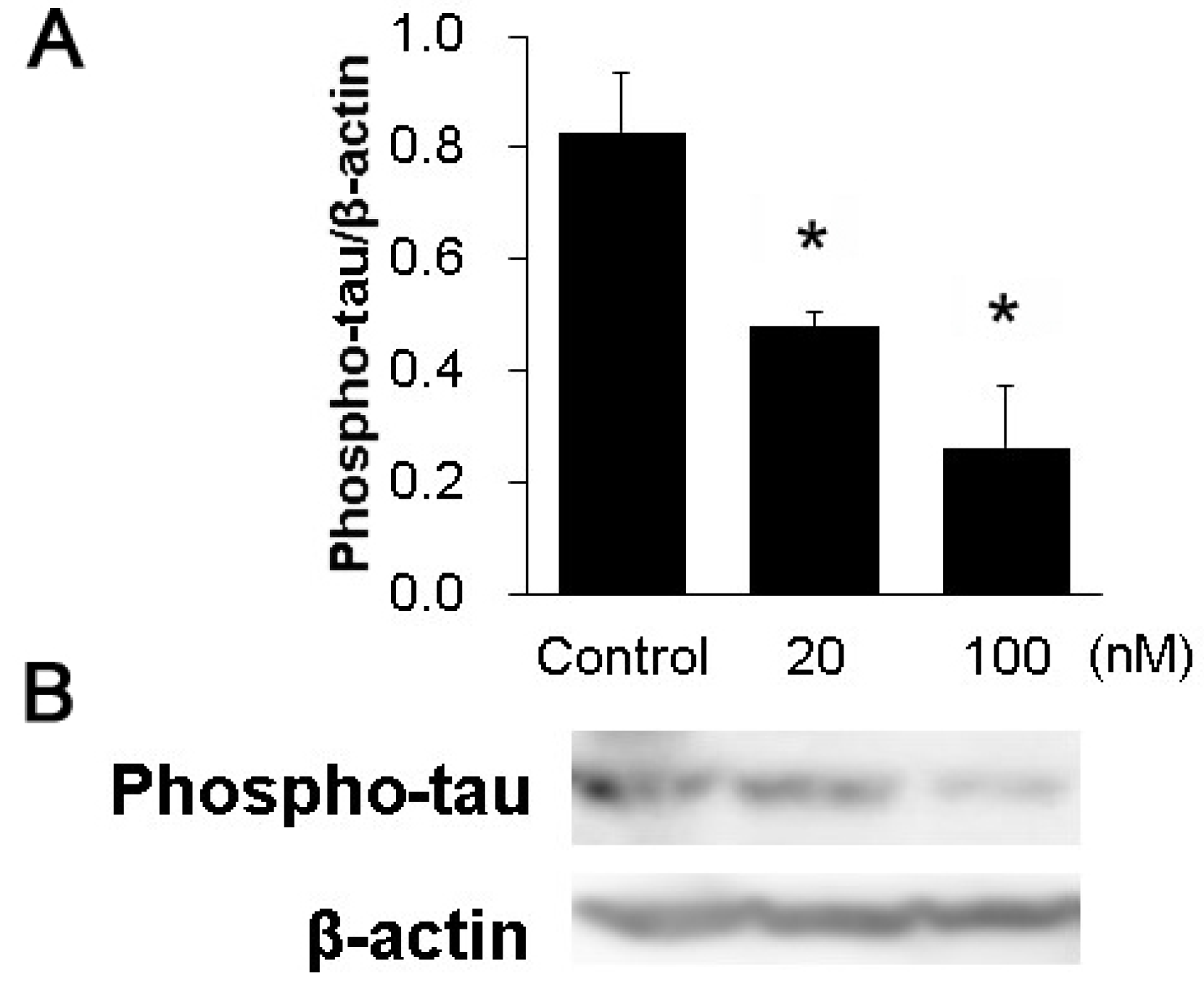
3.3. The Phosphorylation of Tau in the Hippocampal Slices
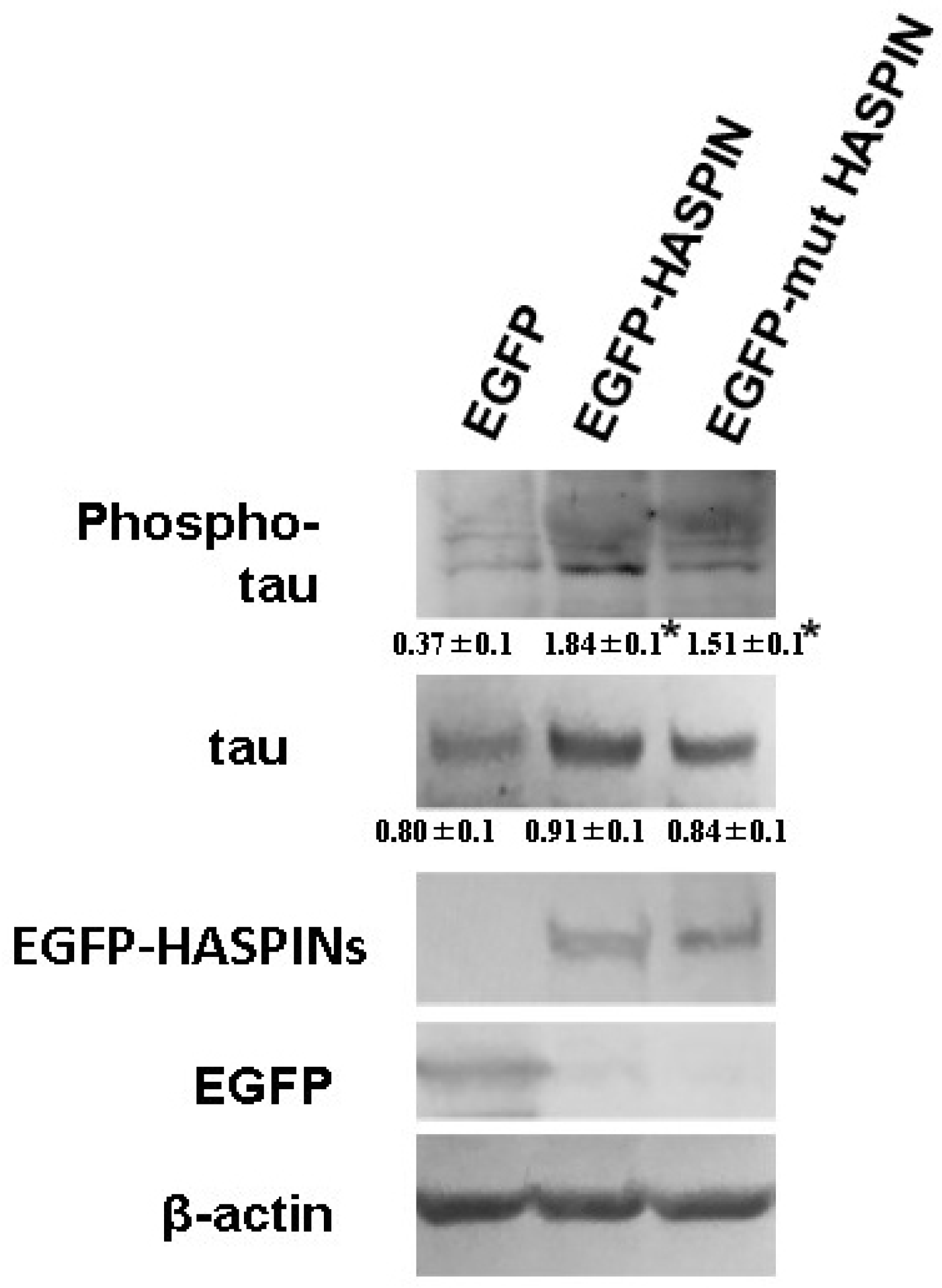
3.4. The Phosphorylation of Tau Protein in HT-22 Cells
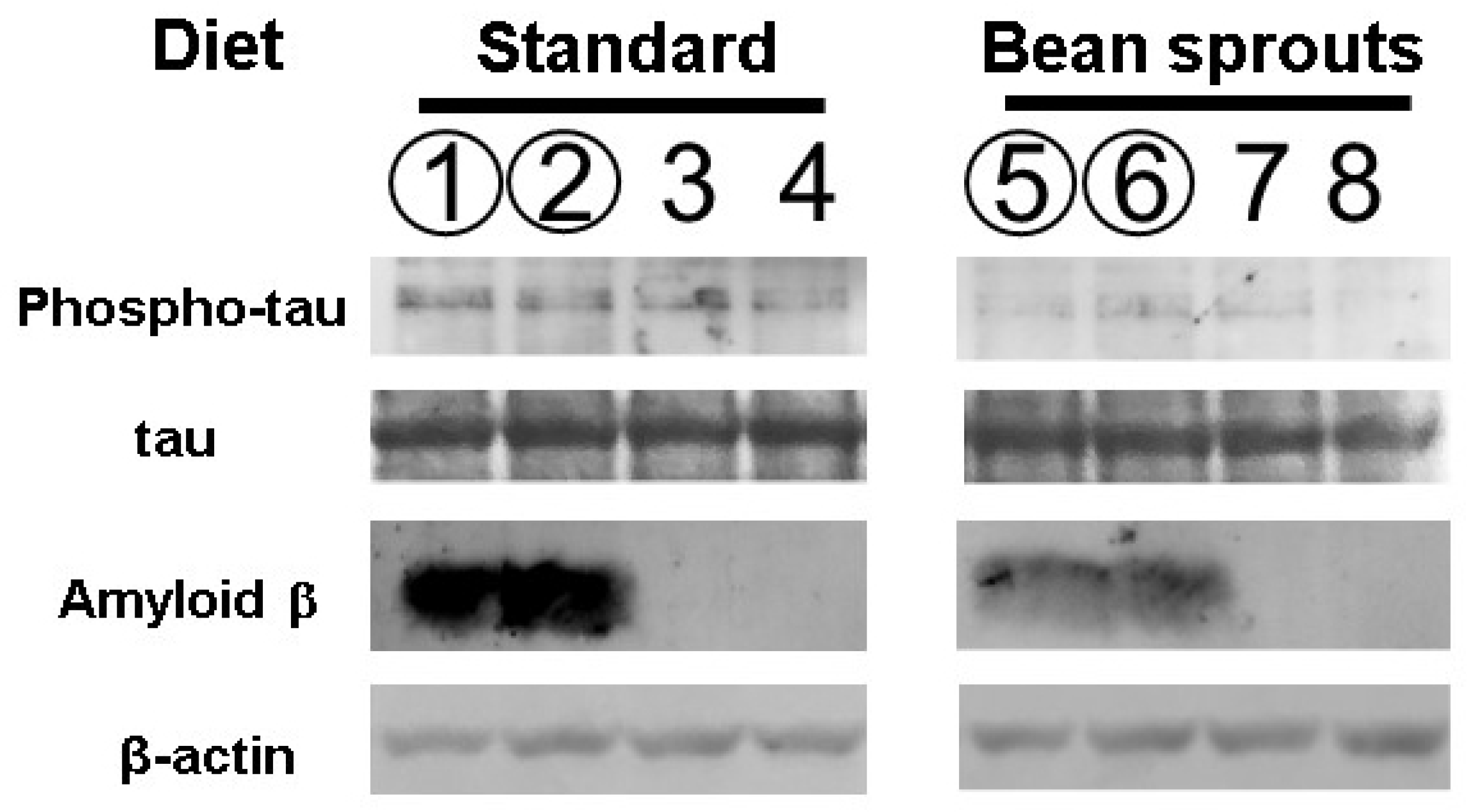
3.5. Effect of the Diet Containing Soybean Sprouts on Tau Protein Phosphorylation in Mice
3.6. Effect of the Diet Containing Soybean Sprouts on Amyloid β Protein in Mice
3.7. Behavioral Experiment of the Mice Fed with Soybean Sprouts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tanaka, H.; Yoshimura, Y.; Nozaki, M.; Yomogida, K.; Tsuchida, J.; Tosaka, Y.; Habu, T.; Nakanishi, T.; Okada, M.; Nojima, H.; et al. Identification and characterization of a haploid germ cell-specific nuclear protein kinase (Haspin) in spermatid nuclei and its effects on somatic cells. J. Biol. Chem. 1999, 274, 17049–17057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, J.M. The Haspin gene: Location in an intron of the integrin alphaE gene, associated transcription of an integrin alphaE-derived RNA and expression in diploid as well as haploid cells. Gene 2001, 267, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Iguchi, N.; Nakamura, Y.; Kohroki, J.; de Carvalho, C.E.; Nishimune, Y. Cloning and characterization of human haspin gene encoding haploid germ cell-specific nuclear protein kinase. Mol. Hum. Reprod. 2001, 7, 211–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, J.M. Structure, function and evolution of haspin and haspin-related proteins, a distinctive group of eukaryotic protein kinases. Cell Mol. Life Sci. 2003, 60, 446–462. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Sultan, S.; Taylor, S.S.; Higgins, J.M. The kinase haspin is required for mitotic histone H3 Thr 3 phosphorylation and normal metaphase chromosome alignment. Genes Dev. 2005, 19, 472–488. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Dai, J.; Daum, J.R.; Niedzialkowska, E.; Banerjee, B.; Stukenberg, P.T.; Gorbsky, G.J.; Higgins, J.M. Histone H3 Thr-3 phosphorylation by Haspin positions Aurora B at centromeres in mitosis. Science 2010, 330, 231–235. [Google Scholar] [CrossRef] [Green Version]
- Hada, M.; Kim, J.; Inoue, E.; Fukuda, Y.; Tanaka, H.; Watanabe, Y.; Okada, Y. TH2A is phosphorylated at meiotic centromere by Haspin. Chromosoma 2017, 126, 769–780. [Google Scholar] [CrossRef]
- Maeda, N.; Tsuchida, J.; Nishimune, Y.; Tanaka, H. Analysis of Ser/Thr Kinase HASPIN-Interacting Proteins in the Spermatids. Int. J. Mol. Sci. 2022, 23, 9060. [Google Scholar] [CrossRef]
- Tanaka, H.; Tokuhiro, K. A Haspin promoter element induces tissue-specific methylation of a transcription region and the regulation of gene expression in mouse ova. Cell J. 2022, 24, 552–554. [Google Scholar] [CrossRef]
- Quadri, R.; Sertic, S.; Muzi-Falconi, M. Roles and regulation of Haspin kinase and its impact on carcinogenesis. Cell Signal. 2022, 93, 110303. [Google Scholar] [CrossRef]
- Han, X.; Kuang, T.; Ren, Y.; Lu, Z.; Liao, Q.; Chen, W. Haspin knockdown can inhibit progression and development of pancreatic cancer in vitro and vivo. Exp. Cell. Res. 2019, 385, 111605. [Google Scholar] [CrossRef]
- Shimada, M.; Goshima, T.; Matsuo, H.; Johmura, Y.; Haruta, M.; Murata, K.; Tanaka, H.; Ikawa, M.; Nakanishi, K.; Nakanishi, M. Essential role of autoactivation circuitry on Aurora B-mediated H2AX-pS121 in mitosis. Nat. Commun. 2016, 7, 12059. [Google Scholar] [CrossRef] [Green Version]
- Nelson, P.T.; Alafuzoff, I.; Bigio, E.H.; Bouras, C.; Braak, H.; Cairns, N.J.; Castellani, R.J.; Crain, B.J.; Davies, P.; Tredici, D.K.; et al. Correlation of Alzheimer disease neuropathologic changes with cognitive status: A review of the literature. Neuropathol. Exp. Neurol. 2012, 71, 362–381. [Google Scholar] [CrossRef]
- Ohta, T.; Uto, T.; Tanaka, H. Effective methods for increasing coumestrol in soybean sprouts. PLoS ONE 2021, 16, e0260147. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Gillardon, F.; Skutella, T.; Uhlmann, E.; Holsboer, F.; Zimmermann, M.; Behl, C. Activation of c-Fos contributes to amyloid beta-peptide-induced neurotoxicity. Brain Res. 1996, 706, 169–172. [Google Scholar] [CrossRef]
- Oakley, H.; Cole, S.L.; Logan, S.; Maus, E.; Shao, P.; Craft, J.; Guillozet-Bongaarts, A.; Ohno, M.; Disterhoft, J.; Van Eldik, L.; et al. Intraneuronal beta-amyloid aggregates, neurodegeneration, and neuron loss in transgenic mice with five familial Alzheimer’s disease mutations: Potential factors in amyloid plaque formation. J. Neurosci. 2006, 26, 10129–10140. [Google Scholar] [CrossRef] [Green Version]
- Huertas, D.; Soler, M.; Moreto, J.; Villanueva, A.; Martinez, A.; Vidal, A.; Charlton, M.; Moffat, D.; Patel, S.; McDermott, J.; et al. Antitumor activity of a small-molecule inhibitor of the histone kinase Haspin. Oncogene 2012, 31, 1408–1418. [Google Scholar] [CrossRef]
- Kim, J.E.; Lee, S.Y.; Jang, M.; Choi, H.K.; Kim, J.H.; Chen, H.; Lim, T.G.; Dong, Z.; Lee, K.W. Coumestrol Epigenetically Suppresses Cancer Cell Proliferation: Coumestrol Is a Natural Haspin Kinase Inhibitor. Int. J. Mol. Sci. 2017, 18, 2228. [Google Scholar] [CrossRef]
- Creighton, S.D.; Mendell, A.L.; Palmer, D.; Kalisch, B.E.; MacLusky, N.J.; Prado, V.F.; Prado, M.A.M.; Winters, B.D. Dissociable cognitive impairments in two strains of transgenic Alzheimer’s disease mice revealed by a battery of object-based tests. Sci. Rep. 2019, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Montero, G.; Arriagada, F.; Günther, G.; Bollo, S.; Mura, F.; Berríos, E.; Morales, J. Phytoestrogen coumestrol: Antioxidant capacity and its loading in albumin nanoparticles. Int. J. Pharm. 2019, 562, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Seong, S.H.; Kim, B.R.; Cho, M.L.; Kim, T.S.; Im, S.; Han, S.; Jeong, J.W.; Jung, H.A.; Choi, J.S. Phytoestrogen Coumestrol Selectively Inhibits Monoamine Oxidase-A and Amyloid β Self-Aggregation. Nutrients 2022, 14, 3822. [Google Scholar] [CrossRef] [PubMed]
- Baumann, K.; Mandelkow, E.M.; Biernat, J.; Piwnica-Worms, H.; Mandelkow, E. Abnormal Alzheimer-like phosphorylation of tau-protein by cyclin-dependent kinases cdk2 and cdk5. FEBS Lett. 1993, 336, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wang, Y.; Zhuang, X.; Luan, F.; Zhao, C.; Cordeiro, M.N.D.S. Interaction of Coumarin Phytoestrogens with ERα and ERβ: A Molecular Dynamics Simulation Study. Molecules 2020, 25, 1165. [Google Scholar] [CrossRef] [Green Version]
- Blomquist, C.H.; Lima, P.H.; Hotchkiss, J.R. Inhibition of 3alpha-hydroxysteroid dehydrogenase (3alpha-HSD) activity of human lung microsomes by genistein, daidzein, coumestrol and C(18)-, C(19)- and C(21)-hydroxysteroids and ketosteroids. Steroids 2005, 70, 507–514. [Google Scholar] [CrossRef]
- Jeon, H.Y.; Seo, D.B.; Shin, H.J.; Lee, S.J. Effect of Aspergillus oryzae-challenged germination on soybean isoflavone content and antioxidant activity. J. Agric. Food Chem. 2012, 60, 2807–2814. [Google Scholar] [CrossRef]
- Koirala, P.; Seong, S.H.; Jung, H.A.; Choi, J.S. Comparative Evaluation of the Antioxidant and Anti-Alzheimer’s Disease Potential of Coumestrol and Puerarol Isolated from Pueraria lobata Using Molecular Modeling Studies. Molecules 2018, 23, 785. [Google Scholar] [CrossRef] [Green Version]
- Seo, D.B.; Jeong, H.W.; Lee, S.J.; Lee, S.J. Coumestrol induces mitochondrial biogenesis by activating Sirt1 in cultured skeletal muscle cells. J. Agric. Food Chem. 2014, 62, 4298–4305. [Google Scholar] [CrossRef]
- Kim, S.N.; Ahn, S.Y.; Song, H.D.; Kwon, H.J.; Saha, A.; Son, Y.; Cho, Y.K.; Jung, Y.S.; Jeong, H.W.; Lee, Y.H. Antiobesity effec;ts of coumestrol through expansion and activation of brown adipose tissue metabolism. J. Nutr. Biochem. 2020, 76, 108300. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Reference (Product Code) |
|---|---|
| anti-tau-mouse IgG | Abcam (ab80579) |
| anti-phosphorylated tau-rabbit IgG | Cell Signaling Technology (#29957) |
| anti-β-amyloid rabbit IgG | Cell Signaling Technology (#51374) |
| anti-EGFP monoclonal antibody | Tanaka et al. [1] |
| anti-β-actin mouse IgG | Proteintech (HRP-60008) |
| anti-rabbit IgG HRP-linked whole Ab (from donkey) | Amersham Biosciences (NA934V) |
| rabbit anti-rat IgG HRP-secondly antibody | DAKO (P0450) |
| anti-rabbit IgG HRP-linked secondary antibody | Cell Signaling Technology (#7074) |
| anti-mouse IgG HRP-secondly antibody | Cell Signaling Technology (#7076) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, H.; Matsushita, H.; Tokuhiro, K.; Fukunari, A.; Ando, Y. Ingestion of Soybean Sprouts Containing a HASPIN Inhibitor Improves Condition in a Mouse Model of Alzheimer’s Disease. Biology 2023, 12, 320. https://doi.org/10.3390/biology12020320
Tanaka H, Matsushita H, Tokuhiro K, Fukunari A, Ando Y. Ingestion of Soybean Sprouts Containing a HASPIN Inhibitor Improves Condition in a Mouse Model of Alzheimer’s Disease. Biology. 2023; 12(2):320. https://doi.org/10.3390/biology12020320
Chicago/Turabian StyleTanaka, Hiromitsu, Hiroaki Matsushita, Keizo Tokuhiro, Atsushi Fukunari, and Yukio Ando. 2023. "Ingestion of Soybean Sprouts Containing a HASPIN Inhibitor Improves Condition in a Mouse Model of Alzheimer’s Disease" Biology 12, no. 2: 320. https://doi.org/10.3390/biology12020320