
Changes in Community Composition and Functional Traits of Bumblebees in an Alpine Ecosystem Relate to Climate Warming
 , , , , , , , , and
, , , , , , , , and
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
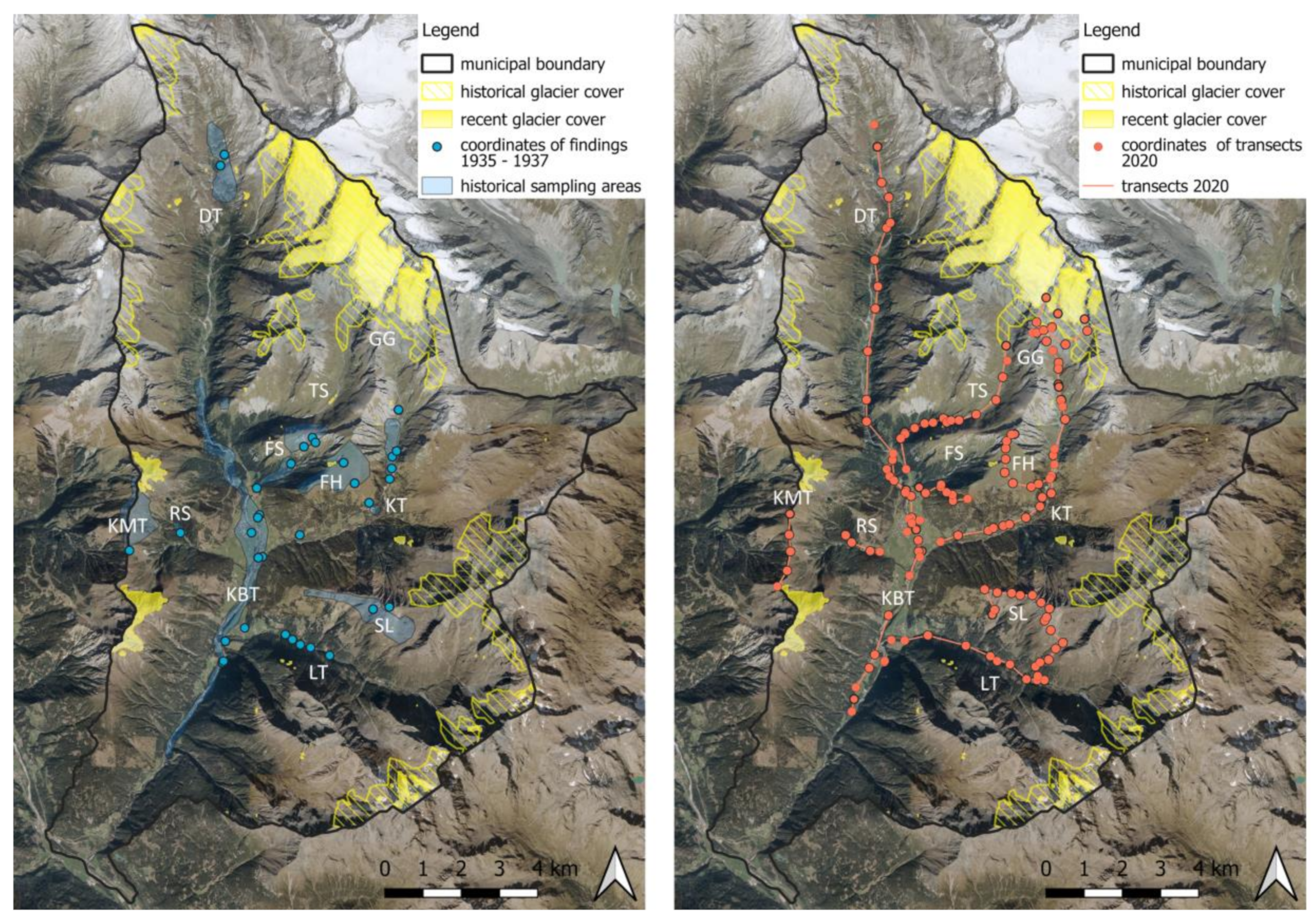
2.1. Study Area
2.2. Bumblebee Data Survey
2.3. Climate Data
2.4. Bumblebee Species Traits
2.5. Statistical Analyses
2.5.1. Processing of Data Records
2.5.2. Analyses on Community Composition
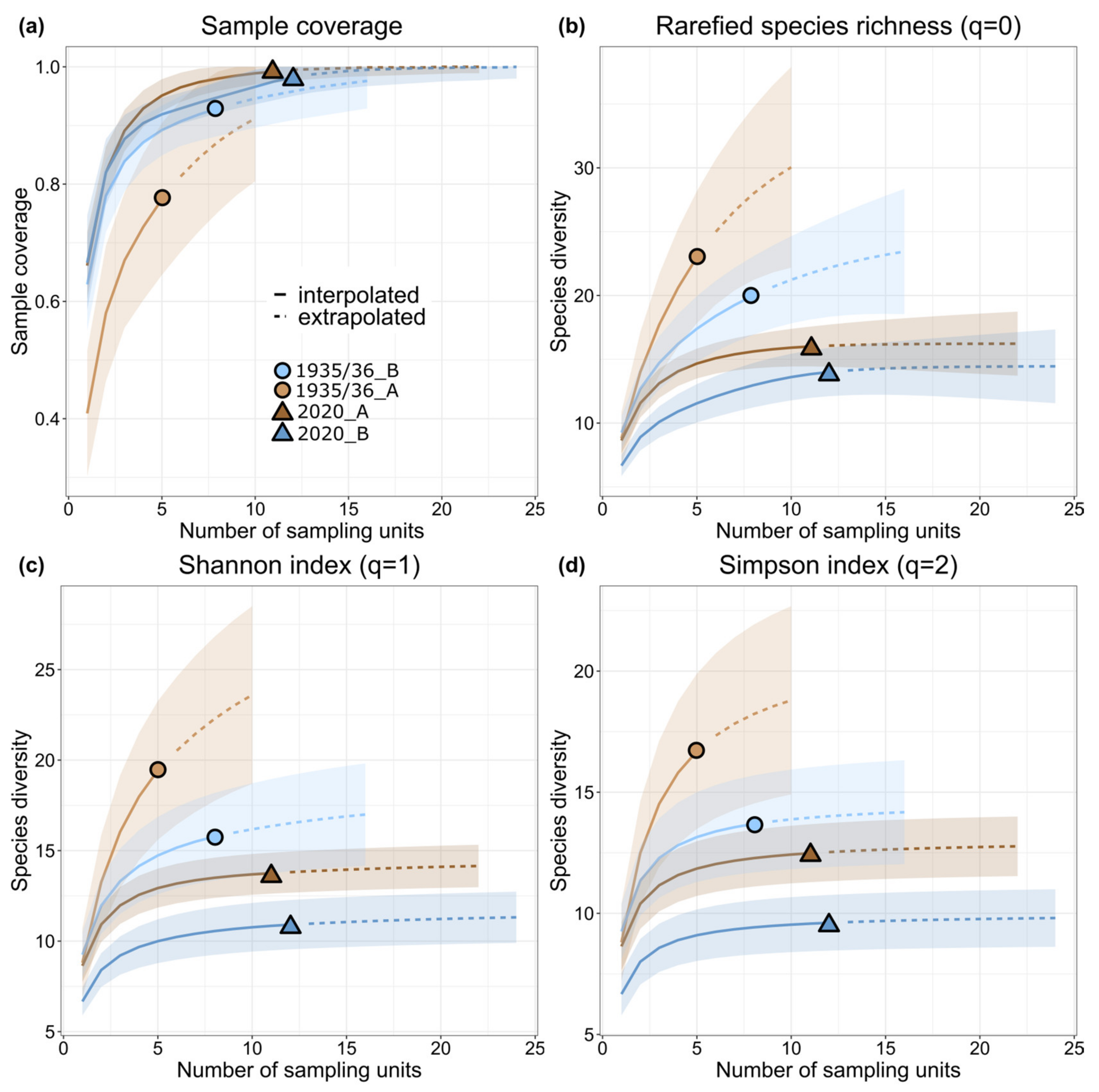
2.5.3. Species Diversity Metrics and Sample Coverage
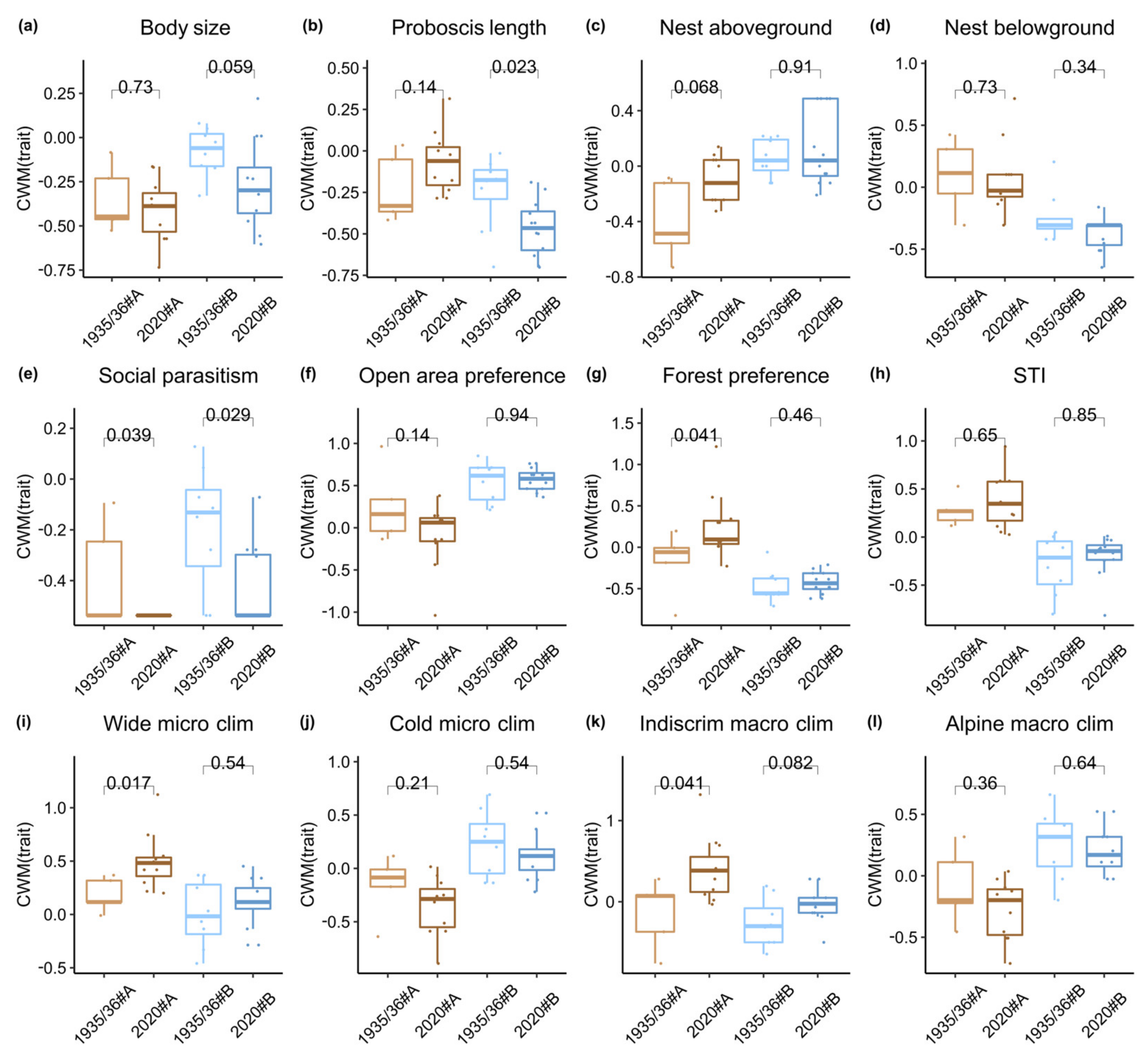
2.5.4. Functional Trait Analyses
3. Results
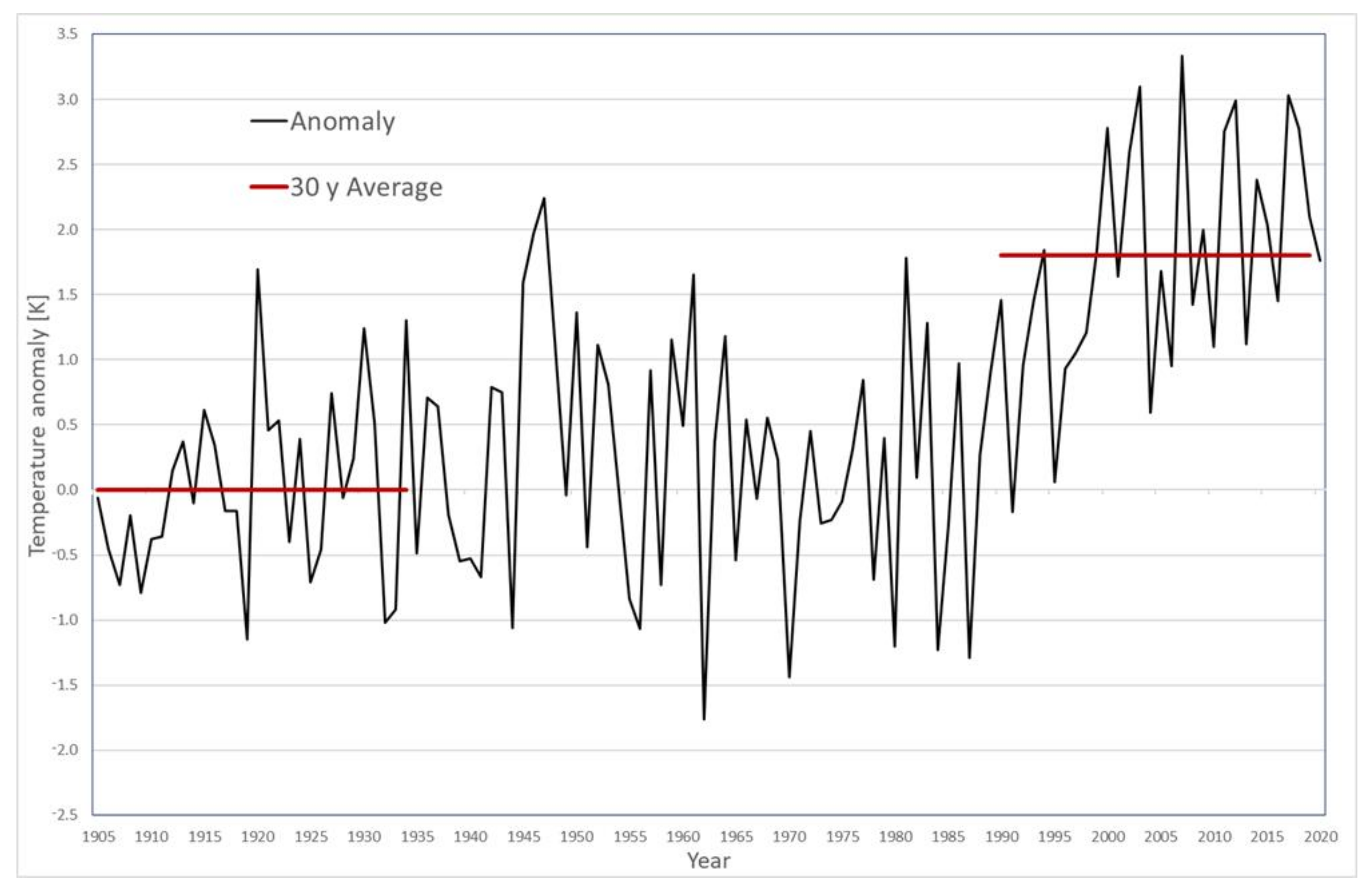
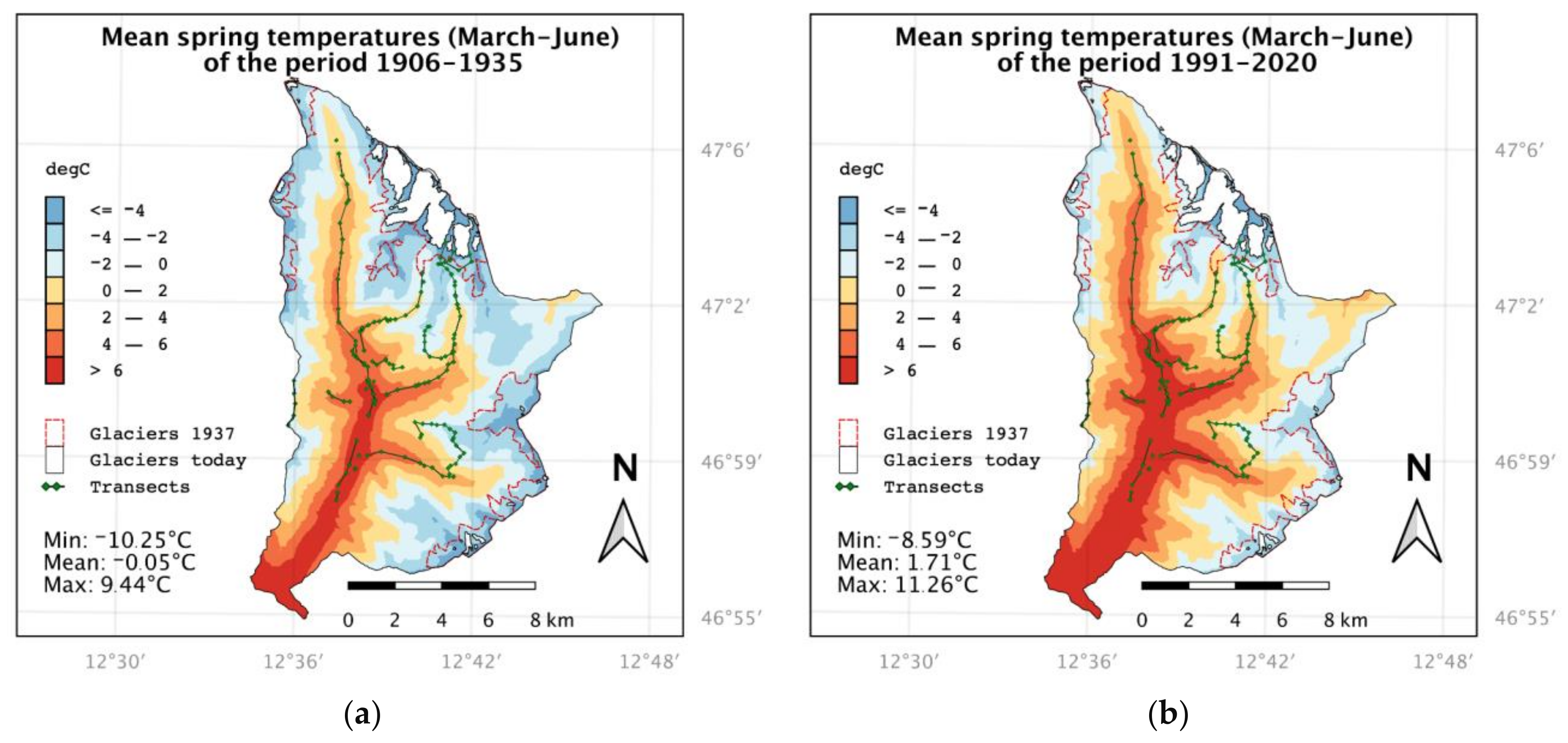
3.1. Climate Warming


3.2. Bumblebee Data
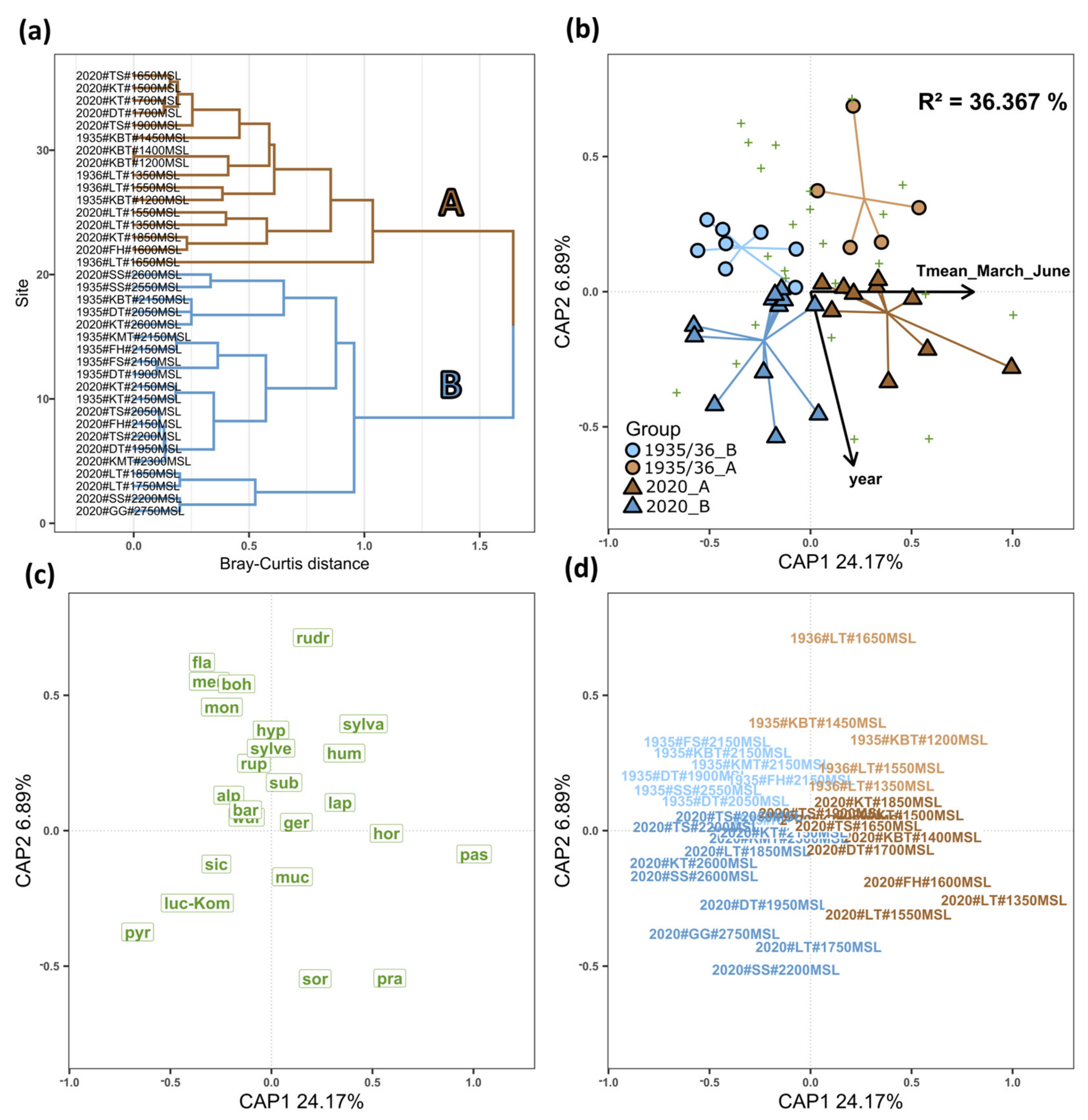
3.3. Cluster Analysis and Bumblebee Community Composition
3.4. Bumblebee Species Diversity
3.5. Distribution of Species Functional Traits
4. Discussion
4.1. Cluster Analysis and Bumblebee Community Composition
4.2. Bumble Species Diversity and Functionality
4.3. Community Changes in the Context of Range Shifts and Extinction Debts
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Body Size [mm] | Proboscis Length [mm] | Nest Aboveground | Nest Belowground | Social Parasitism | Open Area Preference | Forest Preference | STI [°C] | Wide Microclim | Cold Microclim | Indiscriminate Macroclim | Alpine Macroclim |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| alp | 26.0 | 8.469 | 1 | 1 | 0 | 1.0 | 0.0 | -0.3 | 0.0 | 1.0 | 0.0 | 1 |
| bar | 20.0 | NA | NA | NA | 1 | 1.0 | 0.0 | 8.6 | 1.0 | 0.0 | 1.0 | 0 |
| boh | 25.0 | NA | NA | NA | 1 | 0.0 | 0.0 | 6.9 | 1.0 | 0.0 | 1.0 | 0 |
| fla | 18.0 | NA | NA | NA | 1 | 1.0 | 0.0 | 0.6 | 0.0 | 1.0 | 0.0 | 1 |
| ger | 26.0 | 16.195 | 1 | 1 | 0 | 0.0 | 1.0 | 4.3 | 0.0 | 1.0 | 0.0 | 1 |
| hor | 20.0 | 12.764 | 1 | 1 | 0 | 0.0 | 1.0 | 8.2 | 1.0 | 0.0 | 1.0 | 0 |
| hum | 18.0 | 8.469 | 0 | 1 | 0 | 0.0 | 1.0 | 8.1 | 0.0 | 0.0 | 0.0 | 0 |
| hyp | 20.0 | 7.547 | 0 | 1 | 0 | 0.0 | 1.0 | 6.6 | 0.0 | 1.0 | 0.0 | 0 |
| jon | 18.0 | 7.790 | 1 | 1 | 0 | 0.0 | 0.0 | 4.4 | 0.0 | 1.0 | 0.0 | 0 |
| lap | 22.0 | 7.359 | 1 | 1 | 0 | 0.0 | 1.0 | 8.7 | 1.0 | 0.0 | 1.0 | 0 |
| luc-Kom | 20.5 | 7.369 | 1 | 0 | 0 | 0.5 | 0.5 | 7.2 | 0.5 | 0.5 | 0.5 | 0 |
| men | 20.0 | 11.688 | 1 | 0 | 0 | 1.0 | 0.0 | 4.2 | 0.0 | 1.0 | 0.0 | 1 |
| mon | 23.0 | 7.853 | 1 | 1 | 0 | 1.0 | 0.0 | 5.2 | 0.0 | 1.0 | 0.0 | 1 |
| muc | 20.0 | 9.375 | 1 | 0 | 0 | 1.0 | 0.0 | 4.8 | 0.0 | 1.0 | 0.0 | 1 |
| pas | 18.0 | 9.268 | 1 | 1 | 0 | 0.0 | 1.0 | 8.5 | 1.0 | 0.0 | 1.0 | 0 |
| pra | 17.0 | 7.674 | 0 | 1 | 0 | 0.0 | 1.0 | 8.0 | 1.0 | 0.0 | 1.0 | 0 |
| pyr | 20.0 | 7.969 | 1 | 1 | 0 | 1.0 | 0.0 | 3.8 | 0.0 | 1.0 | 0.0 | 1 |
| qua | 25.0 | NA | NA | NA | 1 | 1.0 | 0.0 | 5.2 | 0.0 | 1.0 | 0.0 | 1 |
| rudr | 18.0 | 9.160 | 0 | 1 | 0 | 1.0 | 0.0 | 7.7 | 1.0 | 0.0 | 0.0 | 0 |
| rup | 22.0 | NA | NA | NA | 1 | 0.0 | 1.0 | 7.8 | 1.0 | 0.0 | 1.0 | 0 |
| sic | 20.0 | 7.291 | 1 | 0 | 0 | 1.0 | 0.0 | 3.8 | 0.0 | 1.0 | 0.0 | 1 |
| sor | 17.0 | 7.700 | 1 | 0 | 0 | 1.0 | 0.0 | 5.5 | 1.0 | 0.0 | 1.0 | 0 |
| sub | 28.0 | 10.811 | 1 | 0 | 0 | 0.0 | 1.0 | 7.7 | 0.0 | 0.0 | 0.0 | 0 |
| sylva | 18.0 | 8.403 | 1 | 1 | 0 | 1.0 | 0.0 | 8.0 | 0.0 | 0.0 | 0.0 | 0 |
| sylve | 16.0 | NA | NA | NA | 1 | 0.0 | 1.0 | 7.5 | 1.0 | 0.0 | 1.0 | 0 |
| wur | 22.0 | 10.622 | 1 | 0 | 0 | 1.0 | 0.0 | 4.1 | 1.0 | 0.0 | 0.0 | 0 |
| Trait Name | Trait Type | Trait Description |
|---|---|---|
| Body size [mm] | Continuous | Mean worker body size in millimeters. Measurements were taken by Johann Neumayer from specimens in a private collection. |
| Proboscis length [mm] | Continuous | Mean proboscis length in millimeters. Measurements were taken by Johann Neumayer from specimens in a private collection. |
| Nest aboveground | Binary | Species build their nests aboveground (0 = no, 1 = yes). |
| Nest belowground | Binary | Species use existing holes belowground for nesting (0 = no, 1 = yes). |
| Social parasitism | Binary | Species occupy existing nests of other bumblebees for reproduction (0 = no, 1 = yes). |
| Open area preference | Binary | Species has preference for open areas such as pastures (0 = no, 1 = yes). |
| Forest preference | Binary | Species has preference for enclosed forest habitats (0 = no, 1 = yes). |
| STI [°C] | Continuous | Mean value of the mean annual temperatures in °C within a species distribution range (European level) according to Rasmont et al. [20]. |
| Wide microclim | Binary | Species has wide microclimatic niche adaptation (0 = no, 1 = yes). |
| Cold microclim | Binary | Species has cold microclimatic niche adaptation (0 = no, 1 = yes). |
| Indiscriminate macroclim | Binary | Species has indiscriminate macroclimatic niche adaptation (0 = no, 1 = yes). |
| Alpine macroclim | Binary | Species has alpine macroclimatic niche adaptation (0 = no, 1 = yes). |
| Species | 1935/36#A | 1935/36#B | 2020#A | 2020#B |
|---|---|---|---|---|
| alp | 0 | 1 | 0 | 1 |
| bar | 1 | 1 | 0 | 1 |
| boh | 1 | 1 | 0 | 0 |
| fla | 0 | 1 | 0 | 0 |
| ger | 1 | 0 | 1 | 0 |
| hor | 1 | 1 | 1 | 0 |
| hum | 1 | 0 | 1 | 0 |
| hyp | 1 | 1 | 1 | 0 |
| jon | 1 | 0 | 0 | 0 |
| lap | 1 | 1 | 1 | 0 |
| luc-Kom | 1 | 1 | 1 | 1 |
| men | 0 | 1 | 0 | 1 |
| mon | 1 | 1 | 1 | 1 |
| muc | 1 | 1 | 1 | 1 |
| pas | 1 | 0 | 1 | 1 |
| pra | 1 | 1 | 1 | 1 |
| pyr | 1 | 1 | 1 | 1 |
| qua | 1 | 1 | 0 | 0 |
| rudr | 1 | 1 | 1 | 1 |
| rup | 1 | 1 | 0 | 1 |
| sic | 1 | 1 | 1 | 1 |
| sor | 1 | 1 | 1 | 1 |
| sub | 1 | 0 | 0 | 0 |
| sylva | 1 | 0 | 1 | 0 |
| sylve | 1 | 1 | 0 | 0 |
| wur | 1 | 1 | 1 | 1 |
References
- Masson-Delmotte, V. Global Warming of 1.5 °C: An IPCC Special Report on Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in Context of Strengthening the Global Response to the Threat of Climate change, Sustainable Development, and Efforts to Eradicate Poverty; Cambridge University Press: Cambridge, UK, 2022; ISBN 9781009157940. [Google Scholar]
- Parmesan, C. Ecological and Evolutionary Responses to Recent Climate Change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef] [Green Version]
- EEA—European Environment Agency. Climate Change, Impacts and Vulnerability in Europe 2016: An Indicator-based Report; Publications Office of the European Union: Luxembourg, 2017; ISBN 9789292138356. [Google Scholar]
- Gehrig-Fasel, J.; Guisan, A.; Zimmermann, N.E. Tree line shifts in the Swiss Alps: Climate change or land abandonment? J. Veg. Sci. 2007, 18, 571–582. [Google Scholar] [CrossRef]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R.P. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Kudo, G. Vulnerability of phenological synchrony between plants and pollinators in an alpine ecosystem. Ecol. Res. 2014, 29, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Schenk, M.; Krauss, J.; Holzschuh, A. Desynchronizations in bee-plant interactions cause severe fitness losses in solitary bees. J. Anim. Ecol. 2018, 87, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Brunetti, M.; Lentini, G.; Maugeri, M.; Nanni, T.; Auer, I.; Böhm, R.; Schöner, W. Climate variability and change in the Greater Alpine Region over the last two centuries based on multi-variable analysis. Int. J. Climatol. 2009, 29, 2197–2225. [Google Scholar] [CrossRef]
- Lenoir, J.; Svenning, J.-C. Climate-related range shifts—A global multidimensional synthesis and new research directions. Ecography 2015, 38, 15–28. [Google Scholar] [CrossRef]
- Marshall, L.; Perdijk, F.; Dendoncker, N.; Kunin, W.; Roberts, S.; Biesmeijer, J.C. Bumblebees moving up: Shifts in elevation ranges in the Pyrenees over 115 years. Proc. Biol. Sci. 2020, 287, 20202201. [Google Scholar] [CrossRef]
- Wiens, J. Ecology and Conservation in a Changing World; 1. Auflage; John Wiley & Sons: New York, NY, USA, 2016; ISBN 9781118895108. [Google Scholar]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Menéndez, R.; Megías, A.G.; Hill, J.K.; Braschler, B.; Willis, S.G.; Collingham, Y.; Fox, R.; Roy, D.B.; Thomas, C.D. Species richness changes lag behind climate change. Proc. Biol. Sci. 2006, 273, 1465–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, P.H.; Osborne, J.L. Bumblebee vulnerability and conservation world-wide. Apidologie 2009, 40, 367–387. [Google Scholar] [CrossRef] [Green Version]
- Soroye, P.; Newbold, T.; Kerr, J. Climate change contributes to widespread declines among bumble bees across continents. Science 2020, 367, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Marshall, L.; Biesmeijer, J.C.; Rasmont, P.; Vereecken, N.J.; Dvorak, L.; Fitzpatrick, U.; Francis, F.; Neumayer, J.; Ødegaard, F.; Paukkunen, J.P.T.; et al. The interplay of climate and land use change affects the distribution of EU bumblebees. Glob. Change Biol. 2018, 24, 101–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, S.A.; Lozier, J.D.; Strange, J.P.; Koch, J.B.; Cordes, N.; Solter, L.F.; Griswold, T.L. Patterns of widespread decline in North American bumble bees. Proc. Natl. Acad. Sci. USA 2011, 108, 662–667. [Google Scholar] [CrossRef] [Green Version]
- Franzén, M.; Molander, M. How threatened are alpine environments? A cross taxonomic study. Biodivers Conserv. 2012, 21, 517–526. [Google Scholar] [CrossRef]
- Rasmont, P.; Franzen, M.; Lecocq, T.; Harpke, A.; Roberts, S.; Biesmeijer, K.; Castro, L.; Cederberg, B.; Dvorak, L.; Fitzpatrick, U.; et al. Climatic Risk and Distribution Atlas of European Bumblebees. BR 2015, 10, 1–236. [Google Scholar] [CrossRef]
- Amiet, F.; Krebs, A. Bienen Mitteleuropas: Gattungen, Lebensweise, Beobachtung; 3., Korrigierte, Auflage; Haupt Verlag: Bern, Switzerland, 2019; ISBN 9783258081045. [Google Scholar]
- Goulson, D. Bumblebees: Their Behaviour and Ecology; Oxford University Press: Oxford, UK, 2003; ISBN 0198526067. [Google Scholar]
- Casey, L.M.; Rebelo, H.; Rotheray, E.; Goulson, D. Evidence for habitat and climatic specializations driving the long-term distribution trends of UK and Irish bumblebees. Divers. Distrib. 2015, 21, 864–875. [Google Scholar] [CrossRef]
- Neumayer, J.; Paulus, H.F. Ökologie Alpiner Hummelgemeinschaften: Blütenbesuch, Ressourcenaufteilung und Energiehaushalt; Untersuchungen in den Ostalpen Österreichs; Biologiezentrum des Oberösterr; Landesmuseums: Linz, Austria, 1999. [Google Scholar]
- Hines, H.M. Historical biogeography, divergence times, and diversification patterns of bumble bees (Hymenoptera: Apidae: Bombus). Syst. Biol. 2008, 57, 58–75. [Google Scholar] [CrossRef]
- Martinet, B.; Lecocq, T.; Smet, J.; Rasmont, P. A protocol to assess insect resistance to heat waves, applied to bumblebees (Bombus Latreille, 1802). PLoS One 2015, 10, e0118591. [Google Scholar] [CrossRef] [Green Version]
- Heinrich, B. Bumblebee Economics; New ed.; Harvard University Press: Cambridge, MA, USA; London, UK, 2004; ISBN 0674016394. [Google Scholar]
- Iserbyt, S.; Rasmont, P. The effect of climatic variation on abundance and diversity of bumblebees: A ten years survey in a mountain hotspot. Ann. De La Société Entomol. De Fr. 2012, 48, 261–273. [Google Scholar] [CrossRef]
- Gagic, V.; Bartomeus, I.; Jonsson, T.; Taylor, A.; Winqvist, C.; Fischer, C.; Slade, E.M.; Steffan-Dewenter, I.; Emmerson, M.; Potts, S.G.; et al. Functional identity and diversity of animals predict ecosystem functioning better than species-based indices. Proc. Biol. Sci. 2015, 282, 20142620. [Google Scholar] [CrossRef] [Green Version]
- MacLean, S.A.; Beissinger, S.R. Species’ traits as predictors of range shifts under contemporary climate change: A review and meta-analysis. Glob. Chang. Biol. 2017, 23, 4094–4105. [Google Scholar] [CrossRef] [PubMed]
- Vogt, F.D.; Heinrich, B.; Dabolt, T.O.; McBath, H.L. Ovary development and colony founding in subarctic and temperate-zone bumblebee queens. Can. J. Zool. 1994, 72, 1551–1556. [Google Scholar] [CrossRef]
- Biella, P.; Bogliani, G.; Cornalba, M.; Manino, A.; Neumayer, J.; Porporato, M.; Rasmont, P.; Milanesi, P. Distribution patterns of the cold adapted bumblebee Bombus alpinus in the Alps and hints of an uphill shift (Insecta: Hymenoptera: Apidae). J. Insect Conserv. 2017, 21, 357–366. [Google Scholar] [CrossRef]
- Kerr, J.T.; Pindar, A.; Galpern, P.; Packer, L.; Potts, S.G.; Roberts, S.M.; Rasmont, P.; Schweiger, O.; Colla, S.R.; Richardson, L.L.; et al. Climate change impacts on bumblebees converge across continents. Science 2015, 349, 177–180. [Google Scholar] [CrossRef]
- Johnson, C.N. Species extinction and the relationship between distribution and abundance. Nature 1998, 394, 272–274. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araújo, M.B. Niche properties and geographical extent as predictors of species sensitivity to climate change. Glob. Ecol. Biogeogr. 2005, 14, 347–357. [Google Scholar] [CrossRef]
- Angert, A.L.; Crozier, L.G.; Rissler, L.J.; Gilman, S.E.; Tewksbury, J.J.; Chunco, A.J. Do species’ traits predict recent shifts at expanding range edges? Ecol. Lett. 2011, 14, 677–689. [Google Scholar] [CrossRef]
- Buckley, L.B.; Kingsolver, J.G. Functional and Phylogenetic Approaches to Forecasting Species’ Responses to Climate Change. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 205–226. [Google Scholar] [CrossRef] [Green Version]
- Devictor, V.; van Swaay, C.; Brereton, T.; Brotons, L.; Chamberlain, D.; Heliölä, J.; Herrando, S.; Julliard, R.; Kuussaari, M.; Lindström, Å.; et al. Differences in the climatic debts of birds and butterflies at a continental scale. Nat. Clim. Chang. 2012, 2, 121–124. [Google Scholar] [CrossRef] [Green Version]
- Fourcade, Y.; Åström, S.; Öckinger, E. Climate and land-cover change alter bumblebee species richness and community composition in subalpine areas. Biodivers. Conserv. 2019, 28, 639–653. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.J.; Gutíerrez, D.; Gutíerrez, J.; Monserrat, V.J. An elevational shift in butterfly species richness and composition accompanying recent climate change. Glob. Chang. Biol. 2007, 13, 1873–1887. [Google Scholar] [CrossRef]
- Kuussaari, M.; Bommarco, R.; Heikkinen, R.K.; Helm, A.; Krauss, J.; Lindborg, R.; Ockinger, E.; Pärtel, M.; Pino, J.; Rodà, F.; et al. Extinction debt: A challenge for biodiversity conservation. Trends Ecol. Evol. 2009, 24, 564–571. [Google Scholar] [CrossRef]
- Dullinger, S.; Essl, F.; Rabitsch, W.; Erb, K.-H.; Gingrich, S.; Haberl, H.; Hülber, K.; Jarosík, V.; Krausmann, F.; Kühn, I.; et al. Europe’s other debt crisis caused by the long legacy of future extinctions. Proc. Natl. Acad. Sci. USA 2013, 110, 7342–7347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maebe, K.; Hart, A.F.; Marshall, L.; Vandamme, P.; Vereecken, N.J.; Michez, D.; Smagghe, G. Bumblebee resilience to climate change, through plastic and adaptive responses. Glob. Chang. Biol. 2021, 27, 4223–4237. [Google Scholar] [CrossRef]
- Pittioni, B. Die Hummelfauna im Kalsbachtal in Ost-Tirol. Ein Beitrag zur Ökologie und Systematik der Hummeln Mitteleuropas; Festschrift für Prof. Dr. Embrik Strand III; Univer Latvijas Universitāte: Riga, Latvia, 1937. [Google Scholar]
- NHM. Pittioni Bee Collection. Available online: http.//pittioni.myspecies.info (accessed on 6 January 2023).
- Loken, A. Studies on Scandinavian Bumble Bees: (Hymenoptera, Apidae). Nor. J. Entomol. 1973, 20, 1–218. [Google Scholar]
- Nationalpark Hohe Tauern. Lebensräume im Nationalpark Hohe Tauern. Available online: https://hohetauern.at/de/natur/lebensraeume.html (accessed on 6 January 2023).
- Brugger, B. Biotopkartierung der Gemeinde Kals am Großglockner; Office of the Tyrolean Provincial Government, Department of Environmental Protection: Innsbruck, Austria, 2014. [Google Scholar]
- ZAMG. Klima—Klimaübersichten—Jahrbuch. Gesamtjahresauswertung Klima Kals am Großglockner 2020. Available online: https.//www.zamg.ac.at/cms/de/klima/klimauebersichten/jahrbuch (accessed on 6 January 2023).
- Boeckli, L.; Brenning, A.; Gruber, S.; Noetzli, J. Permafrost distribution in the European Alps: Calculation and evaluation of an index map and summary statistics. Cryosphere 2012, 6, 807–820. [Google Scholar] [CrossRef] [Green Version]
- Gokcezade, J.F.; Gereben-Krenn, B.-A.; Neumayer, J. Feldbestimmungsschlüssel für die Hummeln Deutschlands, Österreichs und der Schweiz; 2., durchgesehene Auflage; Quelle & Meyer Verlag: Wiebelsheim, Germany, 2018; ISBN 3494017875. [Google Scholar]
- Hiebl, J.; Frei, C. Daily temperature grids for Austria since 1961—Concept, creation and applicability. Appl. Clim. 2016, 124, 161–178. [Google Scholar] [CrossRef]
- Auer, I.; Böhm, R.; Jurkovic, A.; Lipa, W.; Orlik, A.; Potzmann, R.; Schöner, W.; Ungersböck, M.; Matulla, C.; Briffa, K.; et al. HISTALP—Historical instrumental climatological surface time series of the Greater Alpine Region. Int. J. Climatol. 2007, 27, 17–46. [Google Scholar] [CrossRef]
- Zhuang, J.; Dussin, R.; Huard, D.; Bourgault, P.; Banihirwe, A.; Raynaud, S.; Malevich, B.; Schupfner, M.; Levang, S.; Jüling, A.; et al. pangeo-data/xESMF; v0.7.0; Zenodo: Meyrin, Switzerland, 2022. [Google Scholar] [CrossRef]
- Gu, H.; Zong, Z.; Hung, K. A modified superconvergent patch recovery method and its application to large deformation problems. Finite Elem. Anal. Des. 2004, 40, 665–687. [Google Scholar] [CrossRef]
- ESMF. ESMF Reference Manual for Fortran. Available online: https://earthsystemmodeling.org/docs/release/ESMF_8_0_1/ESMF_refdoc/ESMF_refdoc.html. (accessed on 22 December 2022).
- Von Hagen, E.; Aichhorn, A. Hummeln: Bestimmen, Ansiedeln, Vermehren, Schützen; 6. neu bearb. Aufl.; Fauna-Verl.: Nottuln, Germany, 2014; ISBN 9783935980326. [Google Scholar]
- Wiesbauer, H. Wilde Bienen: Biologie, Lebensraumdynamik und Gefährdung. Artenporträts von über 470 Wildbienen Mitteleuropas; 2. Auflage; Verlag Eugen Ulmer: Stuttgart, Germany, 2020; ISBN 9783818611552. [Google Scholar]
- Westrich, P. Die Wildbienen Deutschlands; 2., aktualisierte Auflage; Ulmer: Stuttgart, Germany, 2019; ISBN 9783818610234. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 5 January 2023).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. JOSS 2019, 4, 1686. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 5 January 2023).
- Kassambara, A.; Mundt, F. factoextra: Extract and Visualize the Results of Multivariate Data Analyses. 2020. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 5 January 2023).
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: Interpolation and Extrapolation for Species Diversity. 2020. Available online: http://chao.stat.nthu.edu.tw/wordpress/software_download/ (accessed on 5 January 2023).
- Hill, M.O. Diversity and Evenness: A Unifying Notation and Its Consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sande, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology, R Package Version 1.0-12.1. 2014. Available online: https://cran.r-project.org/web/packages/FD/FD.pdf (accessed on 5 January 2023).
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Ricotta, C.; Moretti, M. CWM and Rao’s quadratic diversity: A unified framework for functional ecology. Oecologia 2011, 167, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Patil, I. Visualizations with statistical details: The ’ggstatsplot’ approach. JOSS 2021, 6, 3167. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS One 2017, 12, e0185809. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Le Féon, V.; Schermann-Legionnet, A.; Delettre, Y.; Aviron, S.; Billeter, R.; Bugter, R.; Hendrickx, F.; Burel, F. Intensification of agriculture, landscape composition and wild bee communities: A large scale study in four European countries. Agric. Ecosyst. Environ. 2010, 137, 143–150. [Google Scholar] [CrossRef]
- Kreisch, W.F.; Abl, M. Monitoring des Bergmähder-Ökosystems der “Pockhorner Wiesen” im Nationalpark Hohe Tauern (Kärnten). Landschaft im Wandel. Sauteria 2006, 14, 185–203. [Google Scholar]
- Iserbyt, S.; Vray, S.; Dendoncker, N.; Viart, S.; Rasmont, P. High-resolution distribution of bumblebees (Bombus spp.) in a mountain area marked by agricultural decline. Ann. De La Société Entomol. De Fr. 2015, 51, 375–391. [Google Scholar] [CrossRef]
- Tinner, W.; Kaltenrieder, P. Rapid responses of high-mountain vegetation to early Holocene environmental changes in the Swiss Alps. J Ecol. 2005, 93, 936–947. [Google Scholar] [CrossRef]
- Dirnböck, T.; Essl, F.; Rabitsch, W. Disproportional risk for habitat loss of high-altitude endemic species under climate change. Glob. Chang. Biol. 2011, 17, 990–996. [Google Scholar] [CrossRef]
- Herrera, J.M.; Ploquin, E.F.; Rasmont, P.; Obeso, J.R. Climatic niche breadth determines the response of bumblebees (Bombus spp.) to climate warming in mountain areas of the Northern Iberian Peninsula. J. Insect. Conserv. 2018, 22, 771–779. [Google Scholar] [CrossRef]
- Neumayer, J. Die Hummeln (Hymenoptera: Apidae, Bombus) des Nationalparks Thayatal, Salzburg. 2010. Available online: https://www.parcs.at/npt/pdf_public/2014/10366_20140725_060012_NeumayerJ.2010-DieHummelnHymenopteraApidaeBombusdesNationalparksThayatal-berarbeiteteFassung.pdf (accessed on 31 January 2023).
- Sheffield, C.S.; Pindar, A.; Packer, L.; Kevan, P.G. The potential of cleptoparasitic bees as indicator taxa for assessing bee communities. Apidologie 2013, 44, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Pradervand, J.-N.; Pellissier, L.; Randin, C.F.; Guisan, A. Functional homogenization of bumblebee communities in alpine landscapes under projected climate change. Clim. Chang. Responses 2014, 1, 1. [Google Scholar] [CrossRef] [Green Version]
- Bridle, J.R.; Vines, T.H. Limits to evolution at range margins: When and why does adaptation fail? Trends Ecol. Evol. 2007, 22, 140–147. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Grytnes, J.-A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E.; et al. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.A. Butterfly communities under threat. Science 2016, 353, 216–218. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scharnhorst, V.S.; Thierolf, K.; Neumayer, J.; Becsi, B.; Formayer, H.; Lanner, J.; Ockermüller, E.; Mirwald, A.; König, B.; Kriechbaum, M.; et al. Changes in Community Composition and Functional Traits of Bumblebees in an Alpine Ecosystem Relate to Climate Warming. Biology 2023, 12, 316. https://doi.org/10.3390/biology12020316
Scharnhorst VS, Thierolf K, Neumayer J, Becsi B, Formayer H, Lanner J, Ockermüller E, Mirwald A, König B, Kriechbaum M, et al. Changes in Community Composition and Functional Traits of Bumblebees in an Alpine Ecosystem Relate to Climate Warming. Biology. 2023; 12(2):316. https://doi.org/10.3390/biology12020316
Chicago/Turabian StyleScharnhorst, Victor Sebastian, Katharina Thierolf, Johann Neumayer, Benedikt Becsi, Herbert Formayer, Julia Lanner, Esther Ockermüller, Alina Mirwald, Barbara König, Monika Kriechbaum, and et al. 2023. "Changes in Community Composition and Functional Traits of Bumblebees in an Alpine Ecosystem Relate to Climate Warming" Biology 12, no. 2: 316. https://doi.org/10.3390/biology12020316