
Reduced Renal CSE/CBS/H2S Contributes to the Progress of Lupus Nephritis
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Tissues
2.2. Pristane-Induced Lupus Mice
2.3. MRL/lpr Mice
2.4. R848-Induced Lupus Mice
2.5. HE Staining, Immunohistochemical (IHC) Staining, and Immunofluorescence (IF) Staining
2.6. Microarray Data and Identification of Differentially Expressed Genes (DEGs)
2.7. Functional Enrichment and Protein–Protein Interaction (PPI) Analysis
2.8. ROC Analysis and Profile Infiltrating Immune Cells with ImmucellAI in the Kidney of LN
2.9. Real-Time qPCR Analysis
2.10. Statistical Analysis
3. Results
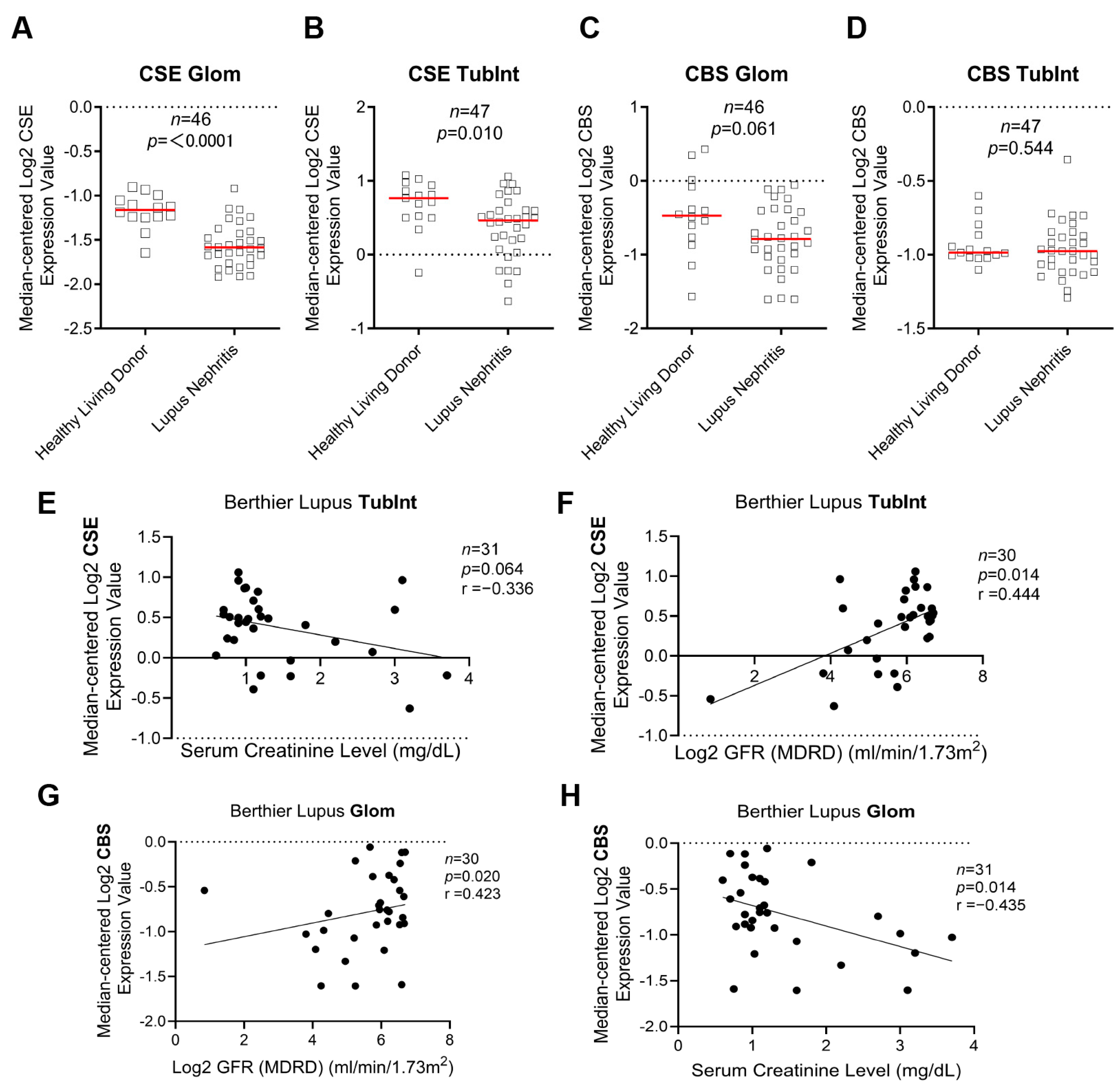
3.1. CSE and CBS Are Downregulated in Renal Tissues of Patients with LN and Are Associated with LN’s Prognosis
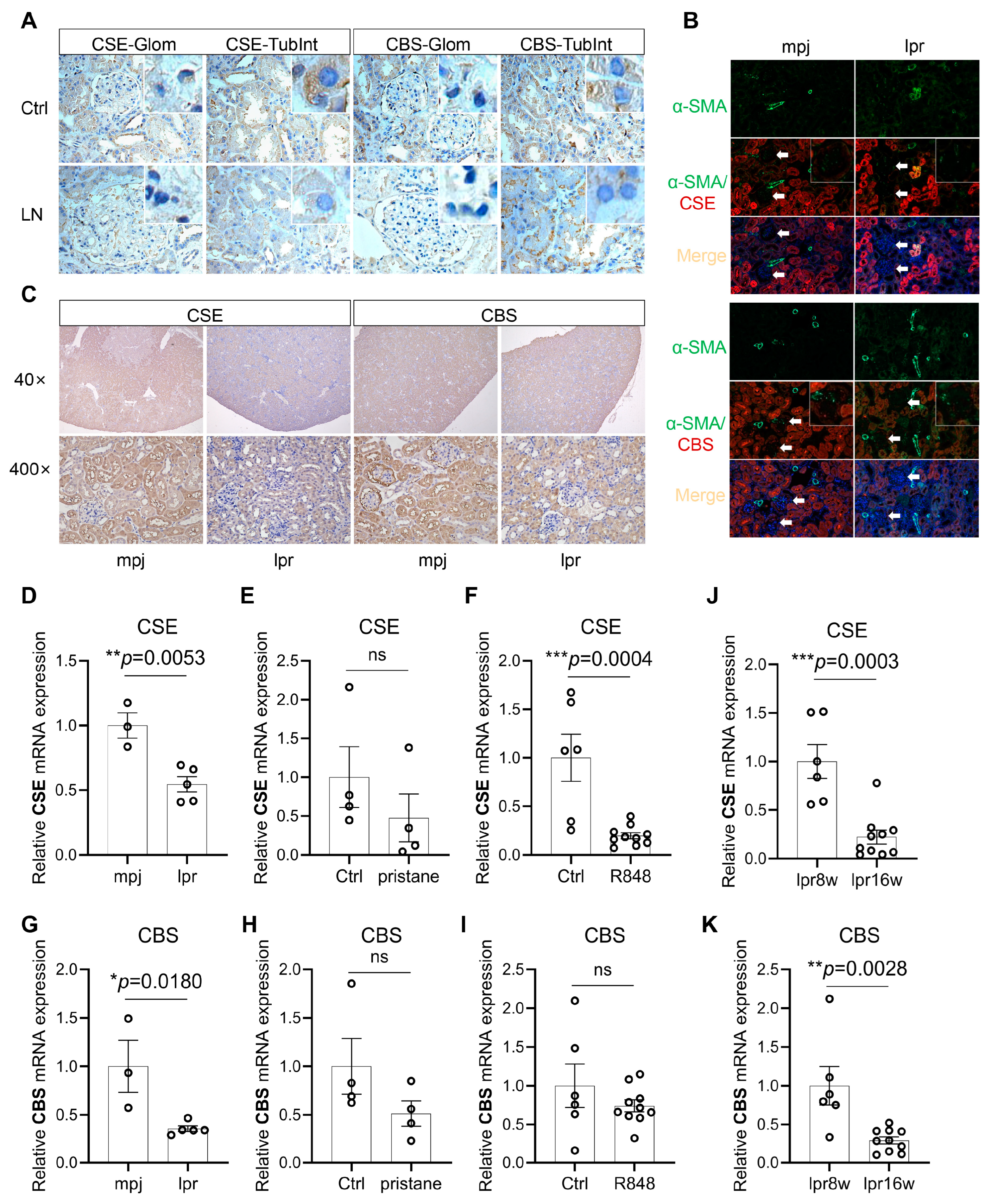
3.2. CSE and CBS Levels Are Reduced in the Renal Tissue of LN Patients and Lupus Mice
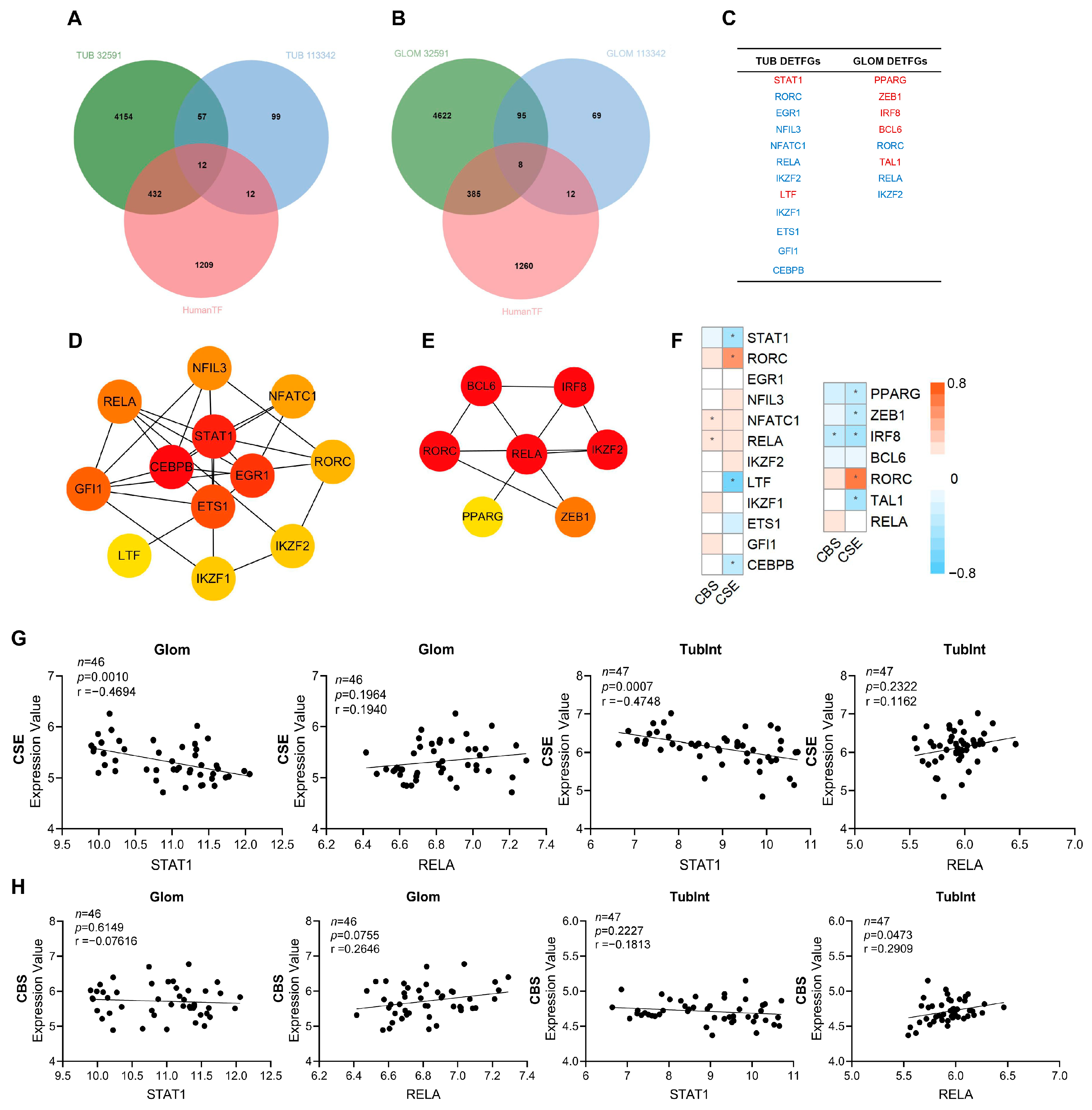
3.3. RELA (p65) and STAT1 Are Hub TFs in the Kidney of LN Patients and CSE and CBS Levels Are Correlated with STAT1 and RELA Levels
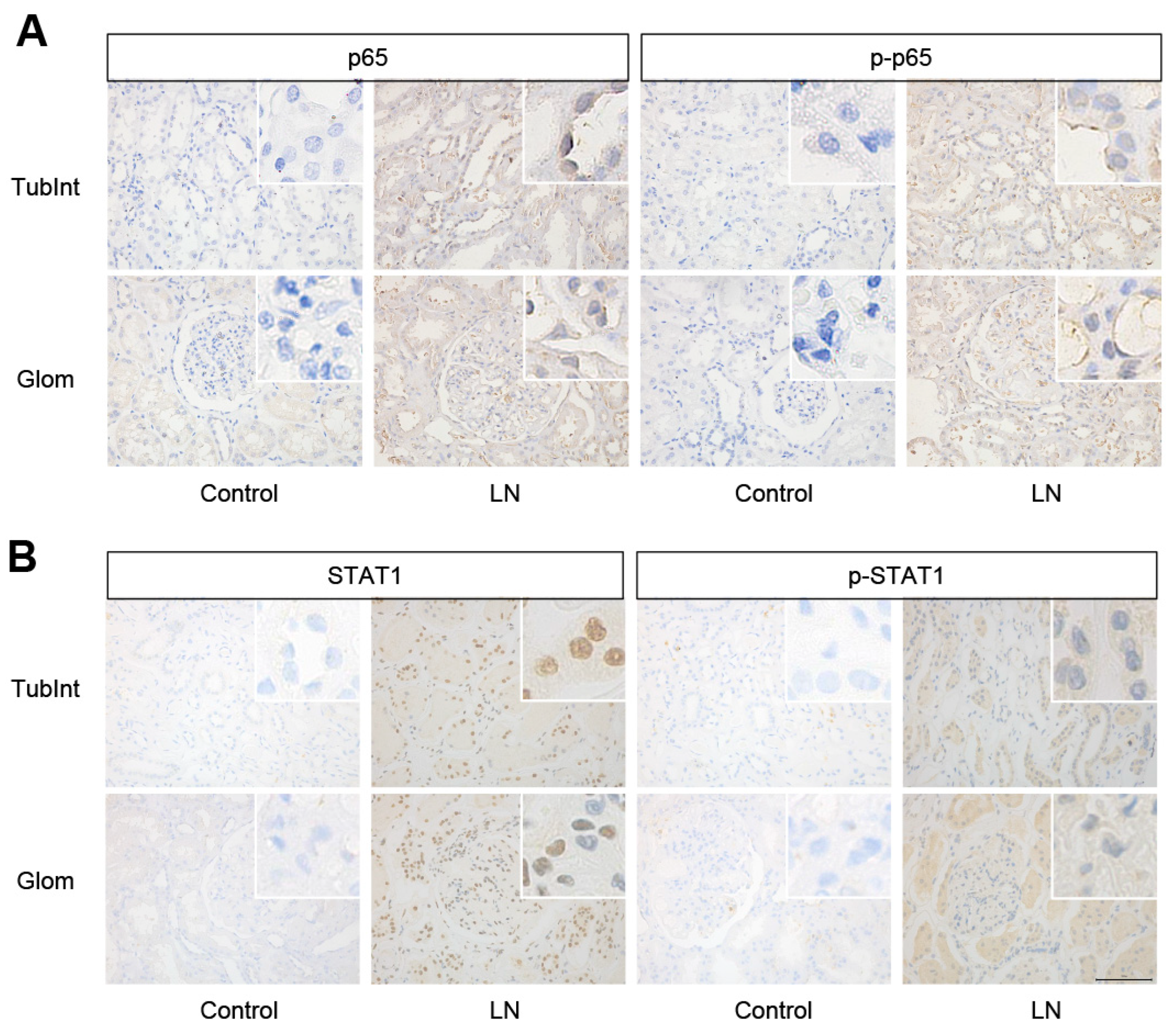
3.4. RELA (p65) and STAT1 Were Increased in the Kidney Tissue of LN Patients
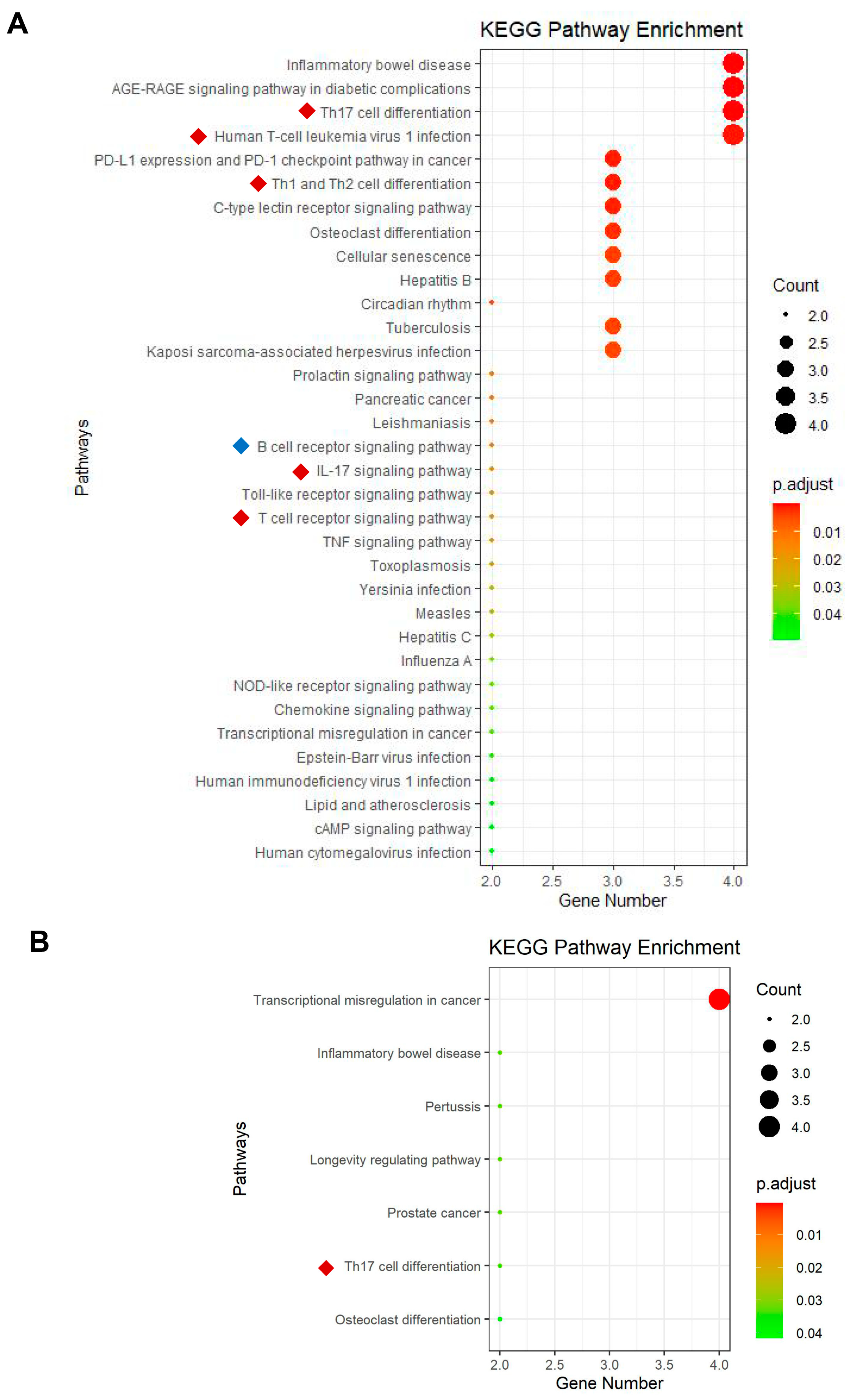
3.5. Key TFs in the Renal Tissue of Patients with LN Are Associated with T-Cell Activation and Function
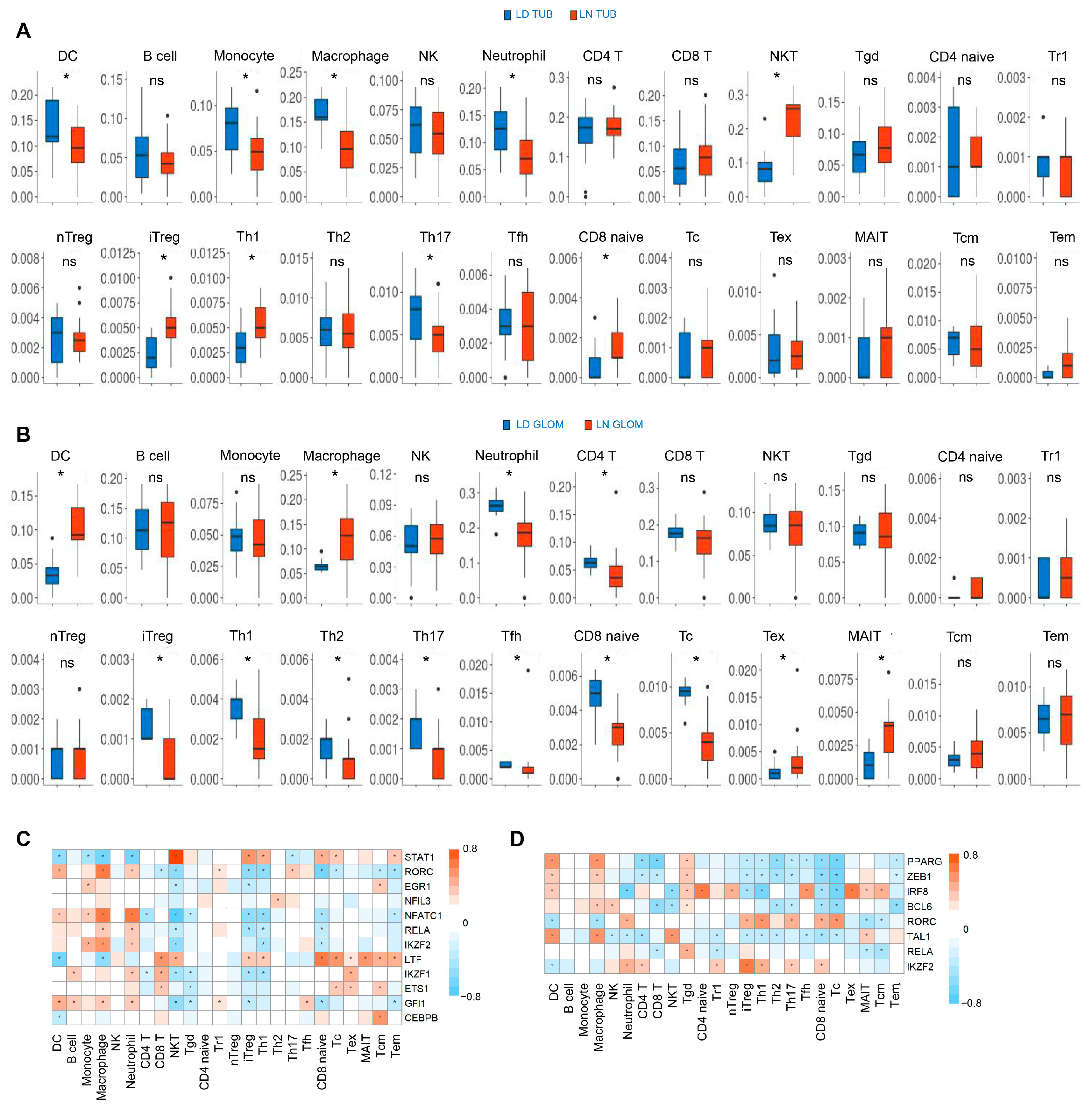
3.6. The Subtypes of Immune Cell Subtypes Are Differentially Regulated in the Renal Tissues of Patients with LN
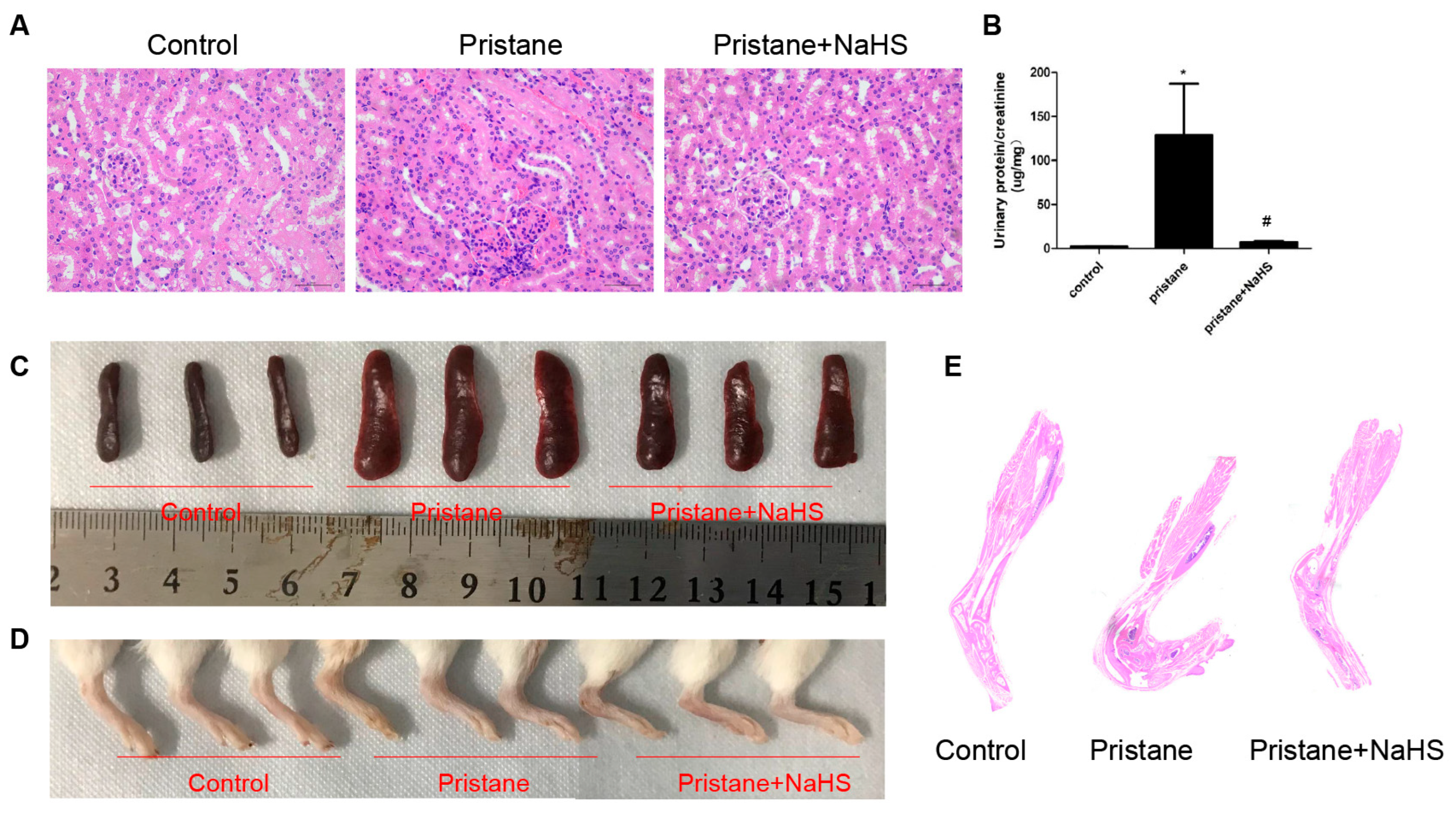
3.7. H2S Donor Alleviates Renal Injury, Hypersplenotrophy, and Arthritis in Pristane-Induced Lupus Mice
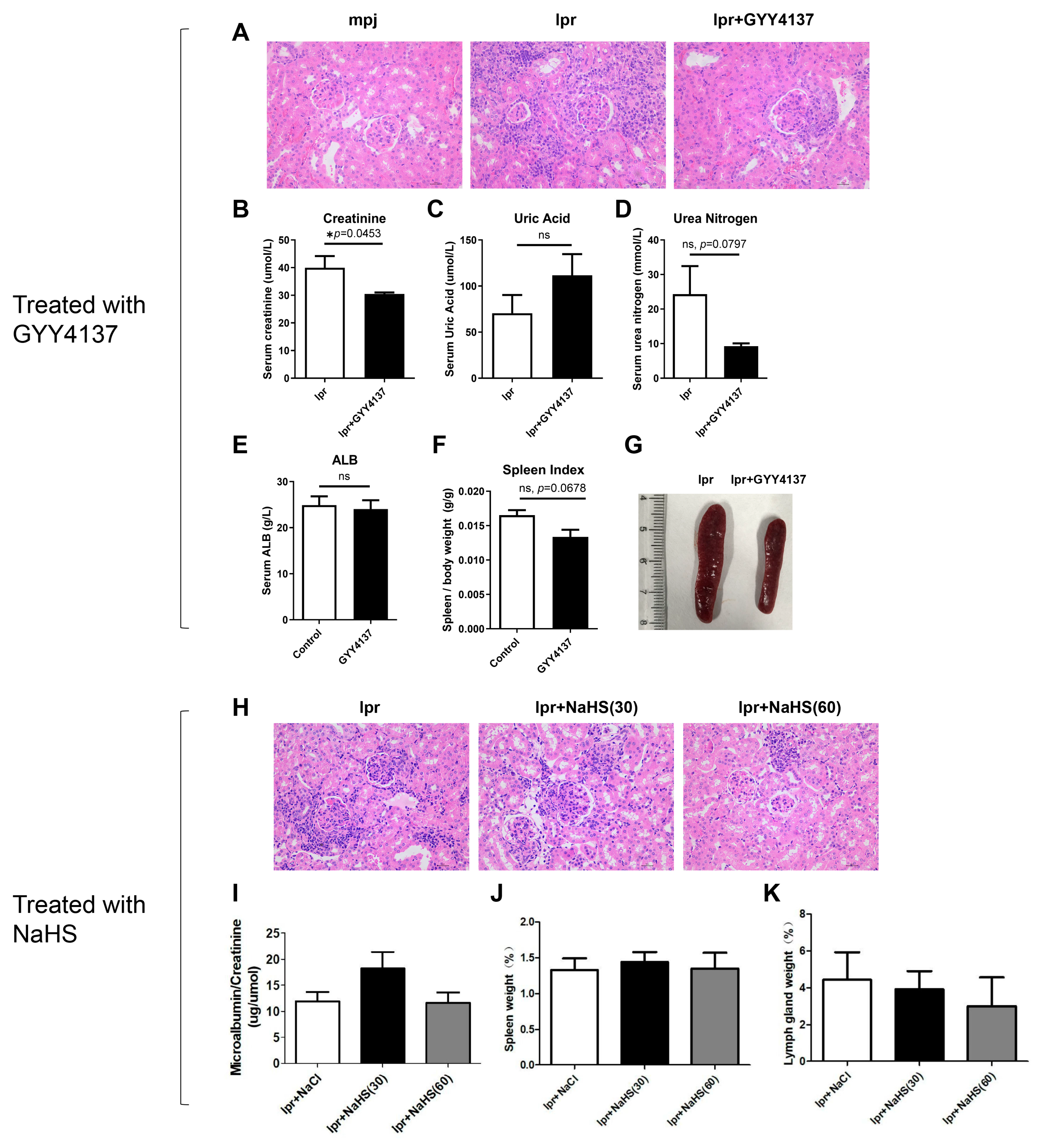
3.8. H2S Donor GYY4137 Attenuates Renal Injury and Reverses Reduced CSE and CBS Expression in MRL/lpr Mice
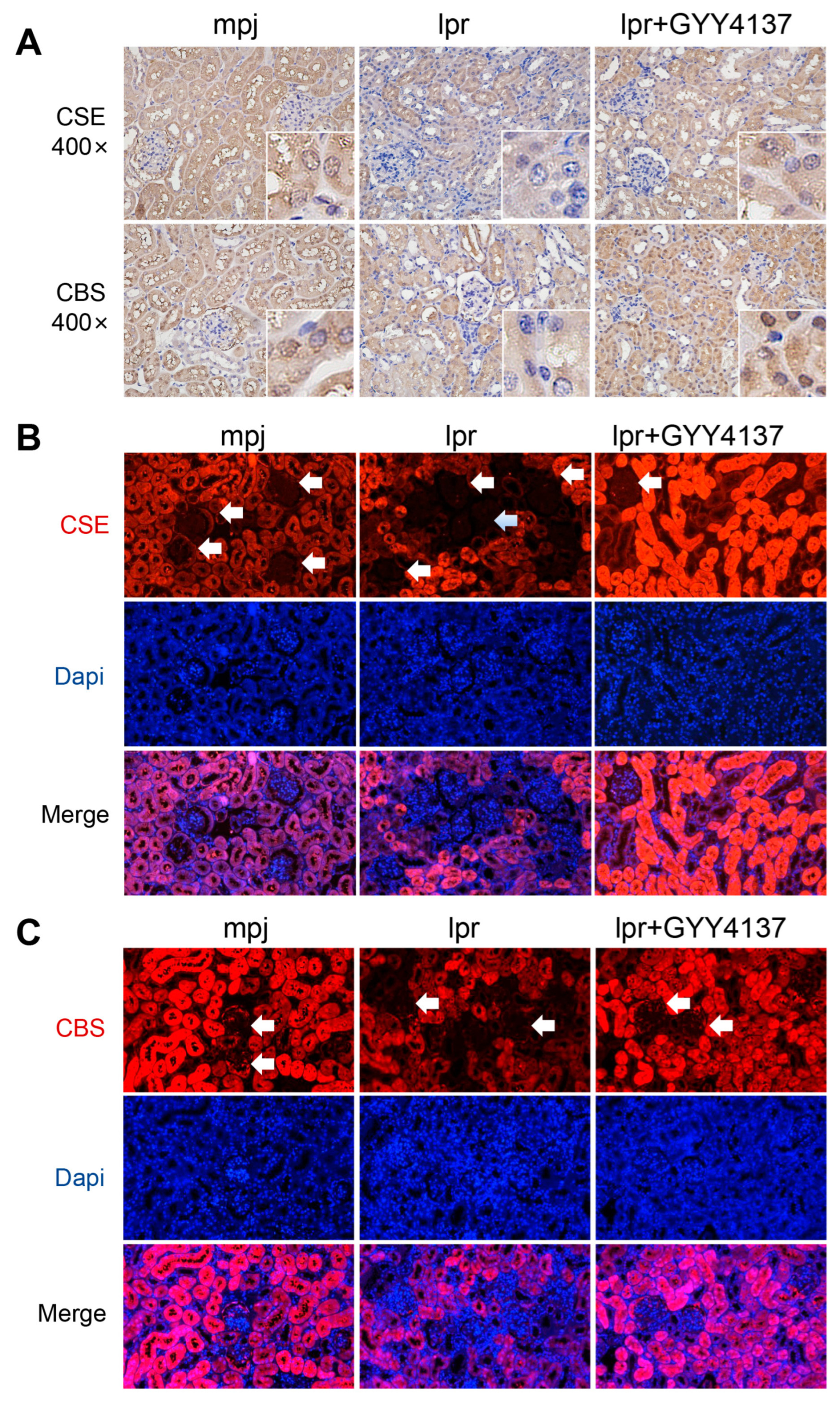
3.9. H2S Donor Recovers Low Expression of CSE and CBS
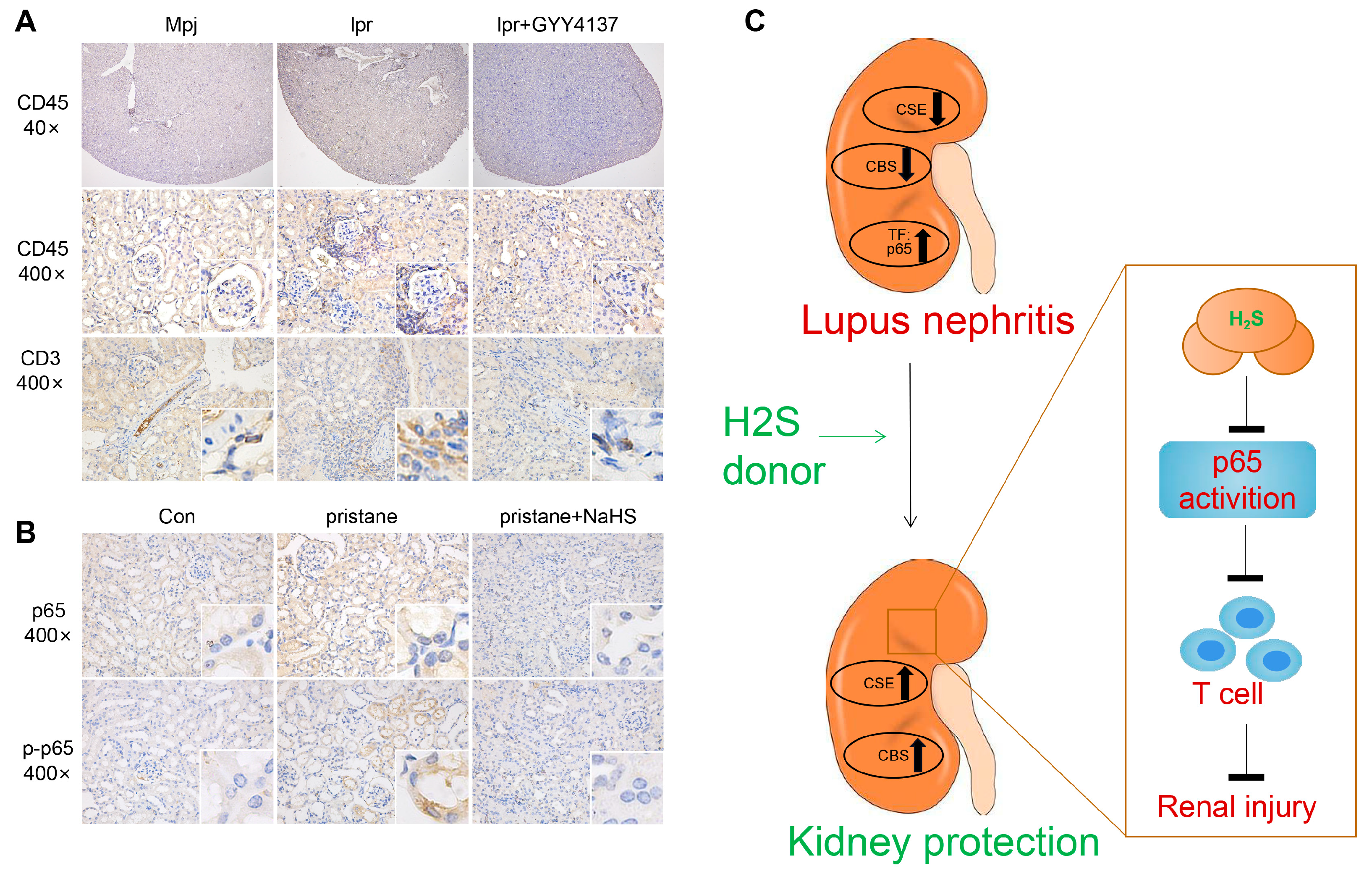
3.10. H2S Donors Inhibit p65 and p-p65 Expression and Reduce Inflammatory Cell Infiltration in the Kidney of Lupus Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, F.; Haas, M.; Glassock, R.; Zhao, M.-H. Redefining lupus nephritis: Clinical implications of pathophysiologic subtypes. Nat. Rev. Nephrol. 2017, 13, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Li, P.; Dang, X.; Zhang, X.; Mao, Y.; Chen, X. Lupus nephritis: New progress in diagnosis and treatment. J. Autoimmun. 2022, 132, 102871. [Google Scholar] [CrossRef] [PubMed]
- Almaani, S.; Meara, A.; Rovin, B.H. Update on Lupus Nephritis. Clin. J. Am. Soc. Nephrol. 2017, 12, 825–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, F.; Chaudhary, K. Lupus Nephritis: A Treatment Update. Curr. Clin. Pharmacol. 2018, 13, 4–13. [Google Scholar] [CrossRef]
- Davidson, A.; Aranow, C.; Mackay, M. Lupus nephritis: Challenges and progress. Curr. Opin. Rheumatol. 2019, 31, 682–688. [Google Scholar] [CrossRef]
- Lech, M.; Anders, H.-J. The pathogenesis of lupus nephritis. J. Am. Soc. Nephrol. 2013, 24, 1357–1366. [Google Scholar] [CrossRef] [Green Version]
- Chang, A.; Clark, M.-R.; Ko, K. Cellular aspects of the pathogenesis of lupus nephritis. Curr. Opin. Rheumatol. 2021, 33, 197–204. [Google Scholar] [CrossRef]
- Chen, C.-Q.; Xin, H.; Zhu, Y.-Z. Hydrogen sulfide: Third gaseous transmitter, but with great pharmacological potential. Acta Pharmacol. Sin. 2007, 28, 1709–1716. [Google Scholar] [CrossRef] [Green Version]
- Faro, M.L.L.; Fox, B.; Whatmore, J.-L.; Winyard, P.G.; Whiteman, M. Hydrogen sulfide and nitric oxide interactions in inflammation. Nitric Oxide 2014, 41, 38–47. [Google Scholar] [CrossRef]
- Sun, H.-J.; Wu, Z.-Y.; Nie, X.-W.; Bian, J.-S. Role of Endothelial Dysfunction in Cardiovascular Diseases: The Link Between Inflammation and Hydrogen Sulfide. Front. Pharmacol. 2019, 10, 1568. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Mao, J.; Zhu, Y. New Therapeutic Approaches Using Hydrogen Sulfide Donors in Inflammation and Immune Response. Antioxid. Redox Signal. 2021, 35, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, M.; Gaddam, R.-R. Hydrogen Sulfide in Inflammation: A Novel Mediator and Therapeutic Target. Antioxid. Redox Signal. 2021, 34, 1368–1377. [Google Scholar] [CrossRef] [PubMed]
- Han, S.-J.; Kim, J.-I.; Park, J.-W.; Park, K.M. Hydrogen sulfide accelerates the recovery of kidney tubules after renal ischemia/reperfusion injury. Nephrol. Dial. Transplant. 2015, 30, 1497–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahangarpour, A.; Fard, A.A.; Gharibnaseri, M.-K.; Jalali, T.; Rashidi, I. Hydrogen sulfide ameliorates the kidney dysfunction and damage in cisplatin-induced nephrotoxicity in rat. Vet. Res. Forum. 2014, 5, 121–127. [Google Scholar]
- Lee, G.; Hosgood, S.-A.; Patel, M.-S.; Nicholson, M.L. Hydrogen sulphide as a novel therapy to ameliorate cyclosporine nephrotoxicity. J. Surg. Res. 2015, 197, 419–426. [Google Scholar] [CrossRef]
- Otunctemur, A.; Ozbek, E.; Dursun, M.; Sahin, S.; Besiroglu, H.; Ozsoy, O.D.; Cekmen, M.; Somay, A.; Ozbay, N. Protective effect of hydrogen sulfide on gentamicin-induced renal injury. Ren. Fail. 2014, 36, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Lobel, L.; Cao, Y.-G.; Fenn, K.; Glickman, J.N.; Garrett, W.S. Diet posttranslationally modifies the mouse gut microbial proteome to modulate renal function. Science 2020, 369, 1518–1524. [Google Scholar] [CrossRef]
- Wang, Y.; Xing, Q.-Q.; Tu, J.-K.; Tang, W.-B.; Yuan, X.-N.; Xie, Y.-Y.; Wang, W.; Peng, Z.-Z.; Huang, L.; Xu, H.; et al. Involvement of hydrogen sulfide in the progression of renal fibrosis. Chin. Med. J. 2019, 132, 2872–2880. [Google Scholar] [CrossRef]
- Sato, S.; Zhang, X.-K.; Temmoku, J.; Fujita, Y.; Matsuoka, N.; Yashiro-Furuya, M.; Asano, T.; Kobayashi, H.; Watanabe, H.; Migita, K. Ets Family Transcription Factor Fli-1 Promotes Leukocyte Recruitment and Production of IL-17A in the MRL/Lpr Mouse Model of Lupus Nephritis. Cells 2020, 9, 714. [Google Scholar] [CrossRef] [Green Version]
- Menke, J.; Bork, T.; Kutska, B.; Byrne, K.T.; Blanfeld, M.; Relle, M.; Kelley, V.R.; Schwarting, A. Targeting transcription factor Stat4 uncovers a role for interleukin-18 in the pathogenesis of severe lupus nephritis in mice. Kidney Int. 2011, 79, 452–463. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Oates, J.-C.; Helke, K.-L.; Gilkeson, G.S.; Zhang, X.K. Camptothecin and Topotecan, Inhibitors of Transcription Factor Fli-1 and Topoisomerase, Markedly Ameliorate Lupus Nephritis in (NZB x NZW)F1 Mice and Reduce the Production of Inflammatory Mediators in Human Renal Cells. Arthritis Rheumatol. 2021, 73, 1478–1488. [Google Scholar] [CrossRef]
- Shelton, L.-M.; Park, B.-K.; Copple, I.-M. Role of Nrf2 in protection against acute kidney injury. Kidney Int. 2013, 84, 1090–1095. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, Y.; Wang, W.; Dai, Y.; Ning, C.; Luo, R.; Sun, K.; Glover, L.; Grenz, A.; Sun, H.; et al. Elevated ecto-5’-nucleotidase-mediated increased renal adenosine signaling via A2B adenosine receptor contributes to chronic hypertension. Circ. Res. 2013, 112, 1466–1478. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhu, X.; Wang, X.; Peng, Y.; Du, J.; Yin, H.; Yang, H.; Ni, X.; Zhang, W. H2S alleviates renal injury and fibrosis in response to unilateral ureteral obstruction by regulating macrophage infiltration via inhibition of NLRP3 signaling. Exp. Cell Res. 2020, 387, 111779. [Google Scholar] [CrossRef] [PubMed]
- Yokogawa, M.; Takaishi, M.; Nakajima, K.; Kamijima, R.; Fujimoto, C.; Kataoka, S.; Terada, Y.; Sano, S. Epicutaneous application of toll-like receptor 7 agonists leads to systemic autoimmunity in wild-type mice: A new model of systemic Lupus erythematosus. Arthritis Rheumatol. 2014, 66, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, Y.; Peng, Y.; Huang, T.; Xia, F.; Yang, T.; Duan, Q.; Zhang, W. Bromodomain-containing protein 4 contributes to renal fibrosis through the induction of epithelial-mesenchymal transition. Exp. Cell Res. 2019, 383, 111507. [Google Scholar] [CrossRef]
- Berthier, C.-C.; Bethunaickan, R.; Gonzalez-Rivera, T.; Nair, V.; Ramanujam, M.; Zhang, W.; Bottinger, E.P.; Segerer, S.; Lindenmeyer, M.; Cohen, C.D.; et al. Cross-species transcriptional network analysis defines shared inflammatory responses in murine and human lupus nephritis. J. Immunol. 2012, 189, 988–1001. [Google Scholar] [CrossRef] [Green Version]
- Mejia-Vilet, J.-M.; Parikh, S.-V.; Song, H.; Fadda, P.; Shapiro, J.P.; Ayoub, I.; Yu, L.; Zhang, J.; Uribe-Uribe, N.; Rovin, B.H. Immune gene expression in kidney biopsies of lupus nephritis patients at diagnosis and at renal flare. Nephrol. Dial. Transplant. 2019, 34, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.-R.; Zhang, Q.; Lei, Q.; Luo, M.; Xie, G.Y.; Wang, H.; Guo, A.Y. ImmuCellAI: A Unique Method for Comprehensive T-Cell Subsets Abundance Prediction and its Application in Cancer Immunotherapy. Adv. Sci. 2020, 7, 1902880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngowi, E.-E.; Sarfraz, M.; Afzal, A.; Khan, N.H.; Khattak, S.; Zhang, X.; Li, T.; Duan, S.-F.; Ji, X.-Y.; Wu, D.-D. Roles of Hydrogen Sulfide Donors in Common Kidney Diseases. Front. Pharmacol. 2020, 11, 564281. [Google Scholar] [CrossRef]
- Han, Y.; Zeng, F.; Tan, G.; Yang, C.; Tang, H.; Luo, Y.; Feng, J.; Xiong, H.; Guo, Q. Hydrogen sulfide inhibits abnormal proliferation of lymphocytes via AKT/GSK3beta signal pathway in systemic lupus erythematosus patients. Cell Physiol. Biochem. 2013, 31, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Qin, M.; Jia, W.; Huang, Z.; Li, Z.; Yang, D.; Huang, M.; Xiao, C.; Long, F.; Mao, J.; et al. Cystathionine-gamma-lyase ameliorates the histone demethylase JMJD3-mediated autoimmune response in rheumatoid arthritis. Cell Mol. Immunol. 2019, 16, 694–705. [Google Scholar] [CrossRef] [PubMed]
- Zhi, L.; Ang, A.-D.; Zhang, H.; Moore, P.K.; Bhatia, M. Hydrogen sulfide induces the synthesis of proinflammatory cytokines in human monocyte cell line U937 via the ERK-NF-kappaB pathway. J. Leukoc. Biol. 2007, 81, 1322–1332. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wang, M.-J.; Jin, S.; Bai, Y.-D.; Hou, C.-L.; Ma, F.-F.; Li, X.-H.; Zhu, Y.-C. The H2S Donor NaHS Changes the Expression Pattern of H2S-Producing Enzymes after Myocardial Infarction. Oxidative Med. Cell Longev. 2016, 2016, 6492469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, K.; Liu, Y.; Zhu, Q.; Liu, C.H.; Duan, J.L.; Tan, B.K.; Zhu, Y.Z. H2S donor, S-propargyl-cysteine, increases CSE in SGC-7901 and cancer-induced mice: Evidence for a novel anti-cancer effect of endogenous H2S? PLoS ONE 2011, 6, e20525. [Google Scholar] [CrossRef] [PubMed]
- Anders, H.-J.; Saxena, R.; Zhao, M.-H.; Ioannis, P.; Salmon, J.E.; Mohan, C. Lupus nephritis. Nat. Rev. Dis. Prim. 2020, 6, 7. [Google Scholar] [CrossRef]
- Chen, P.-M.; Tsokos, G.-C. T Cell Abnormalities in the Pathogenesis of Systemic Lupus Erythematosus: An Update. Curr. Rheumatol. Rep. 2021, 23, 12. [Google Scholar] [CrossRef]
- Katsuyama, T.; Tsokos, G.-C.; Moulton, V.-R. Aberrant T Cell Signaling and Subsets in Systemic Lupus Erythematosus. Front. Immunol. 2018, 9, 1088. [Google Scholar] [CrossRef] [Green Version]
- Ohl, K.; Tenbrock, K. Regulatory T cells in systemic lupus erythematosus. Eur. J. Immunol. 2015, 45, 344–355. [Google Scholar] [CrossRef]
- Shin, M.-S.; Lee, N.; Kang, I. Effector T-cell subsets in systemic lupus erythematosus: Update focusing on Th17 cells. Curr. Opin. Rheumatol. 2011, 23, 444–448. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Huang, S.; Yuan, X.; Liang, J.; Xu, R.; Yao, G.; Feng, X.; Sun, L. The regulation of the Treg/Th17 balance by mesenchymal stem cells in human systemic lupus erythematosus. Cell. Mol. Immunol. 2017, 14, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Zoller, E.-E.; Weirauch, M.-T.; Wu, Z.; Namjou, B.; Williams, A.H.; Ziegler, J.T.; Comeau, M.E.; Marion, M.C.; Glenn, S.B.; et al. Lupus Risk Variant Increases pSTAT1 Binding and Decreases ETS1 Expression. Am. J. Hum. Genet. 2015, 96, 731–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Tang, D.; Yin, L.; Dai, Y. New insights for regulatory T cell in lupus nephritis. Autoimmun. Rev. 2022, 21, 103134. [Google Scholar] [CrossRef] [PubMed]
- Krebs, C.F.; Schmidt, T.; Riedel, J.-H.; Panzer, U. T helper type 17 cells in immune-mediated glomerular disease. Nat. Rev. Nephrol. 2017, 13, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Arazi, A.; Rao, D.A.; Berthier, C.C.; Davidson, A.; Liu, Y.; Hoover, P.J.; Chicoine, A.; Eisenhaure, T.M.; Jonsson, A.H.; Li, S.; et al. The immune cell landscape in kidneys of patients with lupus nephritis. Nat. Immunol. 2019, 20, 902–914. [Google Scholar] [CrossRef]
- Der, E.; Ranabothu, S.; Suryawanshi, H.; Akat, K.M.; Clancy, R.; Morozov, P.; Kustagi, M.; Czuppa, M.; Izmirly, P.; Belmont, H.M.; et al. Single cell RNA sequencing to dissect the molecular heterogeneity in lupus nephritis. JCI Insight 2017, 2, e93009. [Google Scholar] [CrossRef] [Green Version]
- Toubiana, J.; Okada, S.; Hiller, J.; Oleastro, M.; Gomez, M.L.; Becerra, J.C.A.; Ouachée-Chardin, M.; Fouyssac, F.; Girisha, K.; Etzioni, A.; et al. Heterozygous STAT1 gain-of-function mutations underlie an unexpectedly broad clinical phenotype. Blood 2016, 127, 3154–3164. [Google Scholar] [CrossRef]
- Lee, W.; Kim, H.-S.; Baek, S.-Y.; Lee, G.R. Transcription factor IRF8 controls Th1-like regulatory T-cell function. Cell. Mol. Immunol. 2016, 13, 785–794. [Google Scholar] [CrossRef] [Green Version]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Lin, T.; He, Y.; Zhou, Y.; Peng, Y.; Zhang, W.; Ni, X. Reduced Renal CSE/CBS/H2S Contributes to the Progress of Lupus Nephritis. Biology 2023, 12, 318. https://doi.org/10.3390/biology12020318
Wang X, Lin T, He Y, Zhou Y, Peng Y, Zhang W, Ni X. Reduced Renal CSE/CBS/H2S Contributes to the Progress of Lupus Nephritis. Biology. 2023; 12(2):318. https://doi.org/10.3390/biology12020318
Chicago/Turabian StyleWang, Xuan, Tao Lin, Yifei He, Yueyuan Zhou, Yi Peng, Weiru Zhang, and Xin Ni. 2023. "Reduced Renal CSE/CBS/H2S Contributes to the Progress of Lupus Nephritis" Biology 12, no. 2: 318. https://doi.org/10.3390/biology12020318