

Submerged and Solid-State Fermentation of Spirulina with Lactic Acid Bacteria Strains: Antimicrobial Properties and the Formation of Bioactive Compounds of Protein Origin
 ,
,  ,
,  , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
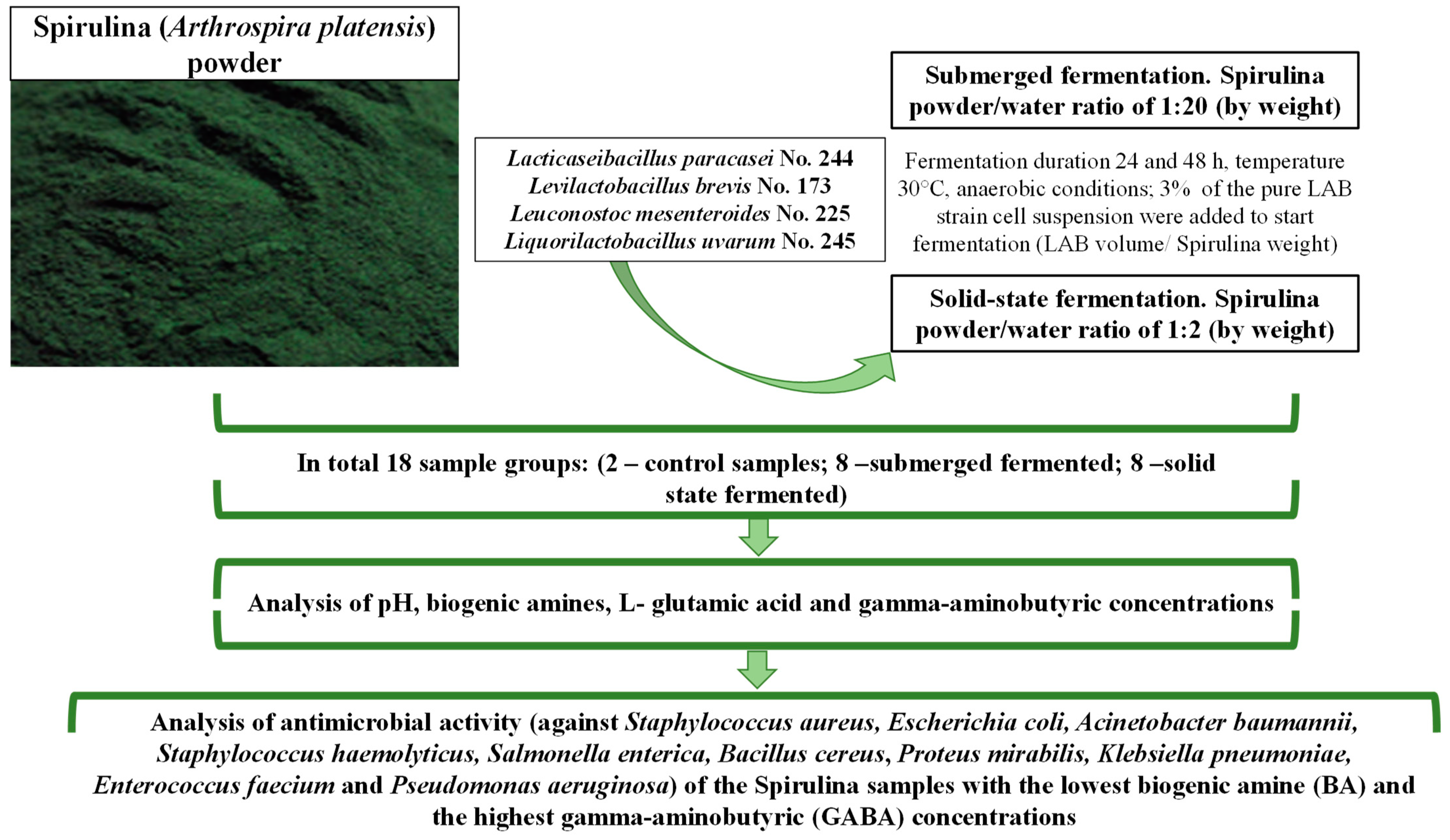
2.1. Spirulina, Microorganisms and Algae Fermentation Conditions
2.2. Evaluation of pH and Lactic acid Bacteria (LAB) Counts in the Spirulina Samples
2.3. Evaluation of the Concentration of L-Glutamic Acid (L-Glu) and Gamma-Aminobutyric (GABA) Acid in Spirulina Samples
2.4. Evaluation of the Concentration of Biogenic Amines (BAs) in Spirulina Samples
2.5. Evaluation of the Antimicrobial Activity of Spirulina Samples
2.6. Statistical Analysis
3. Results
3.1. Effectiveness of Submerged (SMF) and Solid-State (SSF) Fermentation of Spirulina
3.2. Evaluation of the Concentration of L-Glutamic (L-Glu) and Gamma-Aminobutyric (GABA) Acids in Spirulina Samples
3.3. Evaluation of the Concentrations of Biogenic Amines (BA) in Spirulina Samples
3.4. Antimicrobial Activity of Spirulina Samples
3.5. Relationship between the Formation of Bioactive Compounds of Proteinaceous Origin in Spirulina
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Chaiklahan, R.; Chirasuwan, N.; Srinorasing, T.; Attasat, S.; Nopharatana, A.; Bunnag, B. Enhanced Biomass and Phycocyanin Production of Arthrospira (Spirulina) Platensis by a Cultivation Management Strategy: Light Intensity and Cell Concentration. Bioresour. Technol. 2022, 343, 126077. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.A.V.; Freitas, B.C.B.; Rosa, G.M.; Moraes, L.; Morais, M.G.; Mitchell, B.G. Operational and Economic Aspects of Spirulina-Based Biorefinery. Bioresour. Technol. 2019, 292, 121946. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Moneim, A.-M.E.; Shehata, A.M.; Mohamed, N.G.; Elbaz, A.M.; Ibrahim, N.S. Synergistic Effect of Spirulina Platensis and Selenium Nanoparticles on Growth Performance, Serum Metabolites, Immune Responses, and Antioxidant Capacity of Heat-Stressed Broiler Chickens. Biol. Trace Elem. Res. 2022, 200, 768–779. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ramamoorthy, D.; Verma, D.K.; Kumar, A.; Kumar, N.; Kanak, K.R.; Marwein, B.M.; Mohan, K. Antioxidant and Phytonutrient Activities of Spirulina Platensis. Energy Nexus 2022, 6, 100070. [Google Scholar] [CrossRef]
- Guldas, M.; Ziyanok-Demirtas, S.; Sahan, Y.; Yildiz, E.; Gurbuz, O. Antioxidant and Anti-Diabetic Properties of Spirulina Platensis Produced in Turkey. Food Sci. Technol. 2020, 41, 615–625. [Google Scholar] [CrossRef]
- Montalvo, G.E.B.; Vandenberghe, L.P.D.S.; Soccol, V.T.; Carvalho, J.C.D.; Soccol, C.R. The Antihypertensive, Antimicrobial and Anticancer Peptides from Arthrospira with Therapeutic Potential: A Mini Review. Curr. Mol. Med. 2020, 20, 593–606. [Google Scholar] [CrossRef]
- Gogna, S.; Kaur, J.; Sharma, K.; Prasad, R.; Singh, J.; Bhadariya, V.; Kumar, P.; Jarial, S. Spirulina- An Edible Cyanobacterium with Potential Therapeutic Health Benefits and Toxicological Consequences. J. Am. Nutr. Assoc. 2022, 1–14. [Google Scholar] [CrossRef]
- Jung, F.; Krüger-Genge, A.; Waldeck, P.; Küpper, J.-H. Spirulina Platensis, a Super Food? J. Cell. Biotechnol. 2019, 5, 43–54. [Google Scholar] [CrossRef]
- Ovando, C.A.; Carvalho, J.C.D.; Vinícius de Melo Pereira, G.; Jacques, P.; Soccol, V.T.; Soccol, C.R. Functional Properties and Health Benefits of Bioactive Peptides Derived from Spirulina: A Review. Food Rev. Int. 2018, 34, 34–51. [Google Scholar] [CrossRef]
- Trotta, T.; Porro, C.; Cianciulli, A.; Panaro, M.A. Beneficial Effects of Spirulina Consumption on Brain Health. Nutrients 2022, 14, 676. [Google Scholar] [CrossRef]
- Castañeda-Ovando, A.; de Lourdes Pacheco-Hernández, M.; Páez-Hernández, M.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical Studies of Anthocyanins: A Review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- AlFadhly, N.K.Z.; Alhelfi, N.; Altemimi, A.B.; Verma, D.K.; Cacciola, F.; Narayanankutty, A. Trends and Technological Advancements in the Possible Food Applications of Spirulina and Their Health Benefits: A Review. Molecules 2022, 27, 5584. [Google Scholar] [CrossRef]
- Adebo, O.A.; Gabriela Medina-Meza, I. Impact of Fermentation on the Phenolic Compounds and Antioxidant Activity of Whole Cereal Grains: A Mini Review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cheng, H.; Liu, H.; Ma, R.; Ma, J.; Fang, H. Fermentation by Multiple Bacterial Strains Improves the Production of Bioactive Compounds and Antioxidant Activity of Goji Juice. Molecules 2019, 24, 3519. [Google Scholar] [CrossRef]
- Leonard, W.; Zhang, P.; Ying, D.; Adhikari, B.; Fang, Z. Fermentation Transforms the Phenolic Profiles and Bioactivities of Plant-Based Foods. Biotechnol. Adv. 2021, 49, 107763. [Google Scholar] [CrossRef]
- Sharma, R.; Garg, P.; Kumar, P.; Bhatia, S.K.; Kulshrestha, S. Microbial Fermentation and Its Role in Quality Improvement of Fermented Foods. Fermentation 2020, 6, 106. [Google Scholar] [CrossRef]
- D’Este, M.; Alvarado-Morales, M.; Angelidaki, I. Amino Acids Production Focusing on Fermentation Technologies—A Review. Biotechnol. Adv. 2018, 36, 14–25. [Google Scholar] [CrossRef]
- Mindt, M.; Walter, T.; Kugler, P.; Wendisch, V.F. Microbial Engineering for Production of N-Functionalized Amino Acids and Amines. Biotechnol. J. 2020, 15, 1900451. [Google Scholar] [CrossRef] [PubMed]
- Toe, C.J.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Abdul Rahim, R.; Idrus, Z. Extracellular Proteolytic Activity and Amino Acid Production by Lactic Acid Bacteria Isolated from Malaysian Foods. Int. J. Mol. Sci. 2019, 20, 1777. [Google Scholar] [CrossRef]
- Cui, Y.; Miao, K.; Niyaphorn, S.; Qu, X. Production of Gamma-Aminobutyric Acid from Lactic Acid Bacteria: A Systematic Review. Int. J. Mol. Sci. 2020, 21, 995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kieliszek, M.; Pobiega, K.; Piwowarek, K.; Kot, A.M. Characteristics of the Proteolytic Enzymes Produced by Lactic Acid Bacteria. Mol. Basel Switz. 2021, 26, 1858. [Google Scholar] [CrossRef]
- Hati, S.; Patel, N.; Mandal, S. Comparative Growth Behaviour and Biofunctionality of Lactic Acid Bacteria during Fermentation of Soy Milk and Bovine Milk. Probiotics Antimicrob. Proteins 2018, 10, 277–283. [Google Scholar] [CrossRef]
- Venegas-Ortega, M.G.; Flores-Gallegos, A.C.; Martínez-Hernández, J.L.; Aguilar, C.N.; Nevárez-Moorillón, G.V. Production of Bioactive Peptides from Lactic Acid Bacteria: A Sustainable Approach for Healthier Foods. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- García-Cano, I.; Rocha-Mendoza, D.; Kosmerl, E.; Zhang, L.; Jiménez-Flores, R. Technically Relevant Enzymes and Proteins Produced by LAB Suitable for Industrial and Biological Activity. Appl. Microbiol. Biotechnol. 2020, 104, 1401–1422. [Google Scholar] [CrossRef] [PubMed]
- Ngo, D.-H.; Vo, T.S. An Updated Review on Pharmaceutical Properties of Gamma-Aminobutyric Acid. Molecules 2019, 24, 2678. [Google Scholar] [CrossRef]
- Chua, J.-Y.; Koh, M.K.P.; Liu, S.-Q. Gamma-Aminobutyric Acid: A Bioactive Compound in Foods. Sprouted Grains; Feng, H., Nemzer, B., DeVries, J.W., Eds.; Woodhead Publishing and AACC International Press: Cambridge, MA, USA, 2019; pp. 25–54. [Google Scholar]
- Luo, H.; Liu, Z.; Xie, F.; Bilal, M.; Liu, L.; Yang, R.; Wang, Z. Microbial Production of Gamma-Aminobutyric Acid: Applications, State-of-the-Art Achievements, and Future Perspectives. Crit. Rev. Biotechnol. 2021, 41, 491–512. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, X.; Fu, J.; Wang, S.; Chen, Y.; Chang, K.; Li, H. Substrate Sustained Release-Based High Efficacy Biosynthesis of GABA by Lactobacillus Brevis NCL912. Microb. Cell Factories 2018, 17, 80. [Google Scholar] [CrossRef] [PubMed]
- Bartkiene, E.; Lele, V.; Ruzauskas, M.; Domig, K.J.; Starkute, V.; Zavistanaviciute, P.; Bartkevics, V.; Pugajeva, I.; Klupsaite, D.; Juodeikiene, G.; et al. Lactic Acid Bacteria Isolation from Spontaneous Sourdough and Their Characterization Including Antimicrobial and Antifungal Properties Evaluation. Microorganisms 2020, 8, 64. [Google Scholar] [CrossRef]
- Barbieri, F.; Montanari, C.; Gardini, F.; Tabanelli, G. Biogenic Amine Production by Lactic Acid Bacteria: A Review. Foods 2019, 8, 17. [Google Scholar] [CrossRef]
- Feddern, V.; Mazzuco, H.; Fonseca, F.N.; De Lima, G. A Review on Biogenic Amines in Food and Feed: Toxicological Aspects, Impact on Health and Control Measures. Anim. Prod. Sci. 2019, 59, 608–618. [Google Scholar] [CrossRef]
- Ruiz-Capillas, C.; Herrero, A.M. Impact of Biogenic Amines on Food Quality and Safety. Foods 2019, 8, 62. [Google Scholar] [CrossRef] [PubMed]
- Tabanelli, G. Biogenic Amines and Food Quality: Emerging Challenges and Public Health Concerns. Foods 2020, 9, 859. [Google Scholar] [CrossRef] [PubMed]
- Özogul, Y.; Özogul, F. Biogenic Amines Formation, Toxicity, Regulations in Food. In Biogenic Amines in Food: Analysis, Occurrence and Toxicity; Saad, B., Tofalo, R., Eds.; Royal Society of Chemistry: Cambridge, UK, 2019; pp. 1–17. [Google Scholar] [CrossRef]
- Ben-Gigirey, B.; Vieites Baaptista de Sousa, J.M.; Villa, T.G.; Barros-Velazquez, J. Histamine and Cadaverine Production by Bacteria Isolated from Fresh and Frozen Albacore (Thunnus Alalunga). J. Food Prot. 1999, 62, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.-F.; Xu, Y. Production of Tetramethylpyrazine by Batch Culture of Bacillus Subtilis with Optimal PH Control Strategy. J. Ind. Microbiol. Biotechnol. 2010, 37, 815–821. [Google Scholar] [CrossRef]
- Espirito-Santo, A.P.; Carlin, F.; Renard, C.M.G.C. Apple, Grape or Orange Juice: Which One Offers the Best Substrate for Lactobacilli Growth? — A Screening Study on Bacteria Viability, Superoxide Dismutase Activity, Folates Production and Hedonic Characteristics. Food Res. Int. 2015, 78, 352–360. [Google Scholar] [CrossRef]
- Valero-Cases, E.; Frutos, M.J. Development of Prebiotic Nectars and Juices as Potential Substrates for Lactobacillus Acidophilus: Special Reference to Physicochemical Characterization and Consumer Acceptability during Storage. LWT—Food Sci. Technol. 2017, 81, 136–143. [Google Scholar] [CrossRef]
- Chaiklahan, R.; Chirasuwan, N.; Triratana, P.; Loha, V.; Tia, S.; Bunnag, B. Polysaccharide Extraction from Spirulina Sp. and Its Antioxidant Capacity. Int. J. Biol. Macromol. 2013, 58, 73–78. [Google Scholar] [CrossRef]
- Niccolai, A.; Shannon, E.; Abu-Ghannam, N.; Biondi, N.; Rodolfi, L.; Tredici, M.R. Lactic Acid Fermentation of Arthrospira Platensis (Spirulina) Biomass for Probiotic-Based Products. J. Appl. Phycol. 2019, 31, 1077–1083. [Google Scholar] [CrossRef]
- Bao, J.; Zhang, X.; Zheng, J.-H.; Ren, D.-F.; Lu, J. Mixed Fermentation of Spirulina Platensis with Lactobacillus Plantarum and Bacillus Subtilis by Random-Centroid Optimization. Food Chem. 2018, 264, 64–72. [Google Scholar] [CrossRef]
- de Marco Castro, E.; Shannon, E.; Abu-Ghannam, N. Effect of Fermentation on Enhancing the Nutraceutical Properties of Arthrospira Platensis (Spirulina). Fermentation 2019, 5, 28. [Google Scholar] [CrossRef]
- Niccolai, A.; Bažec, K.; Rodolfi, L.; Biondi, N.; Zlatić, E.; Jamnik, P.; Tredici, M.R. Lactic Acid Fermentation of Arthrospira Platensis (Spirulina) in a Vegetal Soybean Drink for Developing New Functional Lactose-Free Beverages. Front. Microbiol. 2020, 11, 560684. [Google Scholar] [CrossRef] [PubMed]
- Kurt, H.; Isleten Hosoglu, M.; Guneser, O.; Karagul-Yuceer, Y. Influence of Different Bacteria Species in Chemical Composition and Sensory Properties of Fermented Spirulina. Food Chem. 2023, 400, 133994. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Jia, F.; Huang, J. The Variation of Two Extracellular Enzymes and Soybean Meal Bitterness during Solid-State Fermentation of Bacillus Subtilis. Grain Oil Sci. Technol. 2019, 2, 39–43. [Google Scholar] [CrossRef]
- Martelli, F.; Cirlini, M.; Lazzi, C.; Neviani, E.; Bernini, V. Solid-State Fermentation of Arthrospira Platensis to Implement New Food Products: Evaluation of Stabilization Treatments and Bacterial Growth on the Volatile Fraction. Foods 2021, 10, 67. [Google Scholar] [CrossRef]
- Moayedi, A.; Mora, L.; Aristoy, M.-C.; Hashemi, M.; Safari, M.; Toldrá, F. ACE-Inhibitory and Antioxidant Activities of Peptide Fragments Obtained from Tomato Processing By-Products Fermented Using Bacillus Subtilis: Effect of Amino Acid Composition and Peptides Molecular Mass Distribution. Appl. Biochem. Biotechnol. 2017, 181, 48–64. [Google Scholar] [CrossRef]
- Sano, C. History of Glutamate Production. Am. J. Clin. Nutr. 2009, 90, 728S–732S. [Google Scholar] [CrossRef]
- Vadopalas, L.; Ruzauskas, M.; Lele, V.; Starkute, V.; Zavistanaviciute, P.; Zokaityte, E.; Bartkevics, V.; Pugajeva, I.; Reinolds, I.; Badaras, S.; et al. Combination of Antimicrobial Starters for Feed Fermentation: Influence on Piglet Feces Microbiota and Health and Growth Performance, Including Mycotoxin Biotransformation in Vivo. Front. Vet. Sci. 2020, 7, 528990. [Google Scholar] [CrossRef] [PubMed]
- Leroy, F.; De Vuyst, L. Lactic Acid Bacteria as Functional Starter Cultures for the Food Fermentation Industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Tanous, C.; Chambellon, E.; Sepulchre, A.-M.; Yvon, M. The Gene Encoding the Glutamate Dehydrogenase in Lactococcus Lactis Is Part of a Remnant Tn3 Transposon Carried by a Large Plasmid. J. Bacteriol. 2005, 187, 5019–5022. [Google Scholar] [CrossRef] [Green Version]
- Zaunmüller, T.; Eichert, M.; Richter, H.; Unden, G. Variations in the Energy Metabolism of Biotechnologically Relevant Heterofermentative Lactic Acid Bacteria during Growth on Sugars and Organic Acids. Appl. Microbiol. Biotechnol. 2006, 72, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, Y.; Roberts, E.; Dietz Jr., G. W. A New Synaptosomal Biosynthetic Pathway of Glutamate and GABA from Ornithine and Its Negative Feedback Inhibition by GABA. J. Neurochem. 1982, 38, 1686–1694. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.G.; Withers, S.E.; Brechany, E.Y.; Banks, J.M. Glutamate Dehydrogenase Activity in Lactobacilli and the Use of Glutamate Dehydrogenase-Producing Adjunct Lactobacillus Spp. Cultures in the Manufacture of Cheddar Cheese. J. Appl. Microbiol. 2006, 101, 1062–1075. [Google Scholar] [CrossRef] [PubMed]
- Eggeling, L.; Bott, M. Handbook of Corynebacterium Glutamicum; CRC Press: Boca Raton, FL, USA, 2005; ISBN 0-429-12905-X. [Google Scholar]
- Zareian, M.; Ebrahimpour, A.; Bakar, F.A.; Mohamed, A.K.S.; Forghani, B.; Ab-Kadir, M.S.B.; Saari, N. A Glutamic Acid-Producing Lactic Acid Bacteria Isolated from Malaysian Fermented Foods. Int. J. Mol. Sci. 2012, 13, 5482–5497. [Google Scholar] [CrossRef] [PubMed]
- Jaichumjai, P.; Valyasevi, R.; Assavanig, A.; Kurdi, P. Isolation and Characterization of Acid-Sensitive Lactobacillus Plantarum with Application as Starter Culture for Nham Production. Food Microbiol. 2010, 27, 741–748. [Google Scholar] [CrossRef]
- Börmann, E.R.; Eikmanns, B.J.; Sahm, H. Molecular Analysis of the Corynebacterium Glutamicum Gdh Gene Encoding Glutamate Dehydrogenase. Mol. Microbiol. 1992, 6, 317–326. [Google Scholar] [CrossRef]
- Wu, Q.; Shah, N.P. High γ-Aminobutyric Acid Production from Lactic Acid Bacteria: Emphasis on Lactobacillus Brevis as a Functional Dairy Starter. Crit. Rev. Food Sci. Nutr. 2017, 57, 3661–3672. [Google Scholar] [CrossRef]
- Wu, J.-Y.; Matsuda, T.; Roberts, E. Purification and Characterization of Glutamate Decarboxylase from Mouse Brain. J. Biol. Chem. 1973, 248, 3029–3034. [Google Scholar] [CrossRef]
- Wu, Q.; Shah, N.P. Restoration of GABA Production Machinery in Lactobacillus Brevis by Accessible Carbohydrates, Anaerobiosis and Early Acidification. Food Microbiol. 2018, 69, 151–158. [Google Scholar] [CrossRef]
- SEO, M.-J.; NAM, Y.-D.; LEE, S.-Y.; PARK, S.-L.; YI, S.-H.; LIM, S.-I. Expression and Characterization of a Glutamate Decarboxylase from Lactobacillus Brevis 877G Producing γ-Aminobutyric Acid. Biosci. Biotechnol. Biochem. 2013, 77, 853–856. [Google Scholar] [CrossRef]
- Li, H.; Qiu, T.; Gao, D.; Cao, Y. Medium Optimization for Production of Gamma-Aminobutyric Acid by Lactobacillus Brevis NCL912. Amino Acids 2010, 38, 1439–1445. [Google Scholar] [CrossRef]
- Li, H.; Qiu, T.; Huang, G.; Cao, Y. Production of Gamma-Aminobutyric Acid by Lactobacillus Brevis NCL912 Using Fed-Batch Fermentation. Microb. Cell Factories 2010, 9, 85. [Google Scholar] [CrossRef] [PubMed]
- Park, K.-B.; Oh, S.-H. Cloning, Sequencing and Expression of a Novel Glutamate Decarboxylase Gene from a Newly Isolated Lactic Acid Bacterium, Lactobacillus Brevis OPK-3. Bioresour. Technol. 2007, 98, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Siragusa, S.; De Angelis, M.; Di Cagno, R.; Rizzello, C.G.; Coda, R.; Gobbetti, M. Synthesis of γ-Aminobutyric Acid by Lactic Acid Bacteria Isolated from a Variety of Italian Cheeses. Appl. Environ. Microbiol. 2007, 73, 7283–7290. [Google Scholar] [CrossRef] [PubMed]
- Diana, M.; Tres, A.; Quílez, J.; Llombart, M.; Rafecas, M. Spanish Cheese Screening and Selection of Lactic Acid Bacteria with High Gamma-Aminobutyric Acid Production. LWT—Food Sci. Technol. 2014, 56, 351–355. [Google Scholar] [CrossRef]
- Park, K.B.; Oh, S.H. Isolation and Characterization of Lactobacillus Buchneri Strains with High γ-Aminobutyric Acid Producing Capacity from Naturally Aged Cheese. Food Sci. Biotechnol. 2006. [Google Scholar]
- Cho, Y.R.; Chang, J.Y.; Chang, H.C. Production of Gamma-Aminobutyric Acid (GABA) by Lactobacillus Buchneri Isolated from Kimchi and Its Neuroprotective Effect on Neuronal Cells. J. Microbiol. Biotechnol. 2007, 17, 104–109. [Google Scholar]
- Gangaraju, D.; Murty, V.R.; Prapulla, S.G. Probiotic-Mediated Biotransformation of Monosodium Glutamate to γ-Aminobutyric Acid: Differential Production in Complex and Minimal Media and Kinetic Modelling. Ann. Microbiol. 2014, 64, 229–237. [Google Scholar] [CrossRef]
- Woraharn, S.; Lailerd, N.; Sivamaruthi, B.S.; Wangcharoen, W.; Sirisattha, S.; Peerajan, S.; Chaiyasut, C. Evaluation of Factors That Influence the L-Glutamic and γ-Aminobutyric Acid Production during Hericium Erinaceus Fermentation by Lactic Acid Bacteria. CyTA—J. Food 2016, 14, 47–54. [Google Scholar] [CrossRef]
- Lin, Q.; Li, D.; Qin, H. Molecular Cloning, Expression, and Immobilization of Glutamate Decarboxylase from Lactobacillus Fermentum YS2. Electron. J. Biotechnol. 2017, 27, 8–13. [Google Scholar] [CrossRef]
- Sun, T.; Zhao, S.; Wang, H.; Cai, C.; Chen, Y.; Zhang, H. ACE-Inhibitory Activity and Gamma-Aminobutyric Acid Content of Fermented Skim Milk by Lactobacillus Helveticus Isolated from Xinjiang Koumiss in China. Eur. Food Res. Technol. 2009, 228, 607–612. [Google Scholar] [CrossRef]
- Franciosi, E.; Carafa, I.; Nardin, T.; Schiavon, S.; Poznanski, E.; Cavazza, A.; Larcher, R.; Tuohy, K.M. Biodiversity and γ-Aminobutyric Acid Production by Lactic Acid Bacteria Isolated from Traditional Alpine Raw Cow’s Milk Cheeses. BioMed Res. Int. 2015, 2015, e625740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, K.; Jiang, Y.; Feng, X.; Li, L.; Dang, F.; Zhang, W.; Man, C. Transcriptomic Response to GABA-Producing Lactobacillus Plantarum CGMCC 1.2437T Induced by L-MSG. PLoS ONE 2018, 13, e0199021. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-Y.; Lü, F.-X.; Lu, Z.-X.; Bie, X.-M.; Jiao, Y.; Sun, L.-J.; Yu, B. Production of γ-Aminobutyric Acid by Streptococcus Salivarius Subsp. Thermophilus Y2 under Submerged Fermentation. Amino Acids 2008, 34, 473–478. [Google Scholar] [CrossRef]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-Aminobutyric Acid as a Bioactive Compound in Foods: A Review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Watanabe, M.; Maemura, K.; Kanbara, K.; Tamayama, T.; Hayasaki, H. GABA and GABA Receptors in the Central Nervous System and Other Organs. Int. Rev. Cytol. 2002, 213, 1–47. [Google Scholar]
- Sahab, N.R.M.; Subroto, E.; Balia, R.L.; Utama, G.L. γ-Aminobutyric Acid Found in Fermented Foods and Beverages: Current Trends. Heliyon 2020, 6, e05526. [Google Scholar] [CrossRef]
- Han, M.; Liao, W.; Wu, S.; Gong, X.; Bai, C. Use of Streptococcus Thermophilus for the in Situ Production of γ-Aminobutyric Acid-Enriched Fermented Milk. J. Dairy Sci. 2020, 103, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A. GABA-Modulating Bacteria of the Human Gut Microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Sanchart, C.; Watthanasakphuban, N.; Boonseng, O.; Nguyen, T.-H.; Haltrich, D.; Maneerat, S. Tuna Condensate as a Promising Low-Cost Substrate for Glutamic Acid and GABA Formation Using Candida Rugosa and Lactobacillus Futsaii. Process Biochem. 2018, 70, 29–35. [Google Scholar] [CrossRef]
- Zhang, S.; Xing, X.; Chu, Q.; Sun, S.; Wang, P. Impact of Co-Culture of Lactobacillus Plantarum and Oenococcus Oeni at Different Ratios on Malolactic Fermentation, Volatile and Sensory Characteristics of Mulberry Wine. LWT 2022, 169, 113995. [Google Scholar] [CrossRef]
- LIN, H.; TSAIG, J. Gamma Aminobutyric AcidproductioninblacksoybeanmilkbyLactobacillus BrevisFPA3709andtheantidepressanteffectofthefer Mentedproductonaforcedswimmingratmodel. ProcessBiochemistry 2013, 48, 559r568. [Google Scholar]
- Yokoyama, S.; Hiramatsu, J.; Hayakawa, K. Production of γ-Aminobutyric Acid from Alcohol Distillery Lees by Lactobacillus Brevis IFO-12005. J. Biosci. Bioeng. 2002, 93, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.S.; Cha, I.-T.; Roh, S.W.; Shin, H.-H.; Seo, M.-J. Enhanced Production of Gamma-Aminobutyric Acid by Optimizing Culture Conditions of Lactobacillus Brevis HYE1 Isolated from Kimchi, a Korean Fermented Food. J. Microbiol. Biotechnol. 2017, 27, 450–459. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-H.; Hsueh, Y.-H.; Kuo, J.-M.; Liu, S.-J. Characterization of a Potential Probiotic Lactobacillus Brevis RK03 and Efficient Production of γ-Aminobutyric Acid in Batch Fermentation. Int. J. Mol. Sci. 2018, 19, 143. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, T.; Tahara, M.; Ohshima, T. Mechanism of Gamma-Aminobutyric Acid (GABA) Production by a Lactic Acid Bacterium in Yogurt-Sake. Process Biochem. 2018, 74, 21–27. [Google Scholar] [CrossRef]
- Bakhiet, N.; Forney, F.W.; Stahly, D.P.; Daniels, L. Lysine Biosynthesis InMethanobacterium Thermoautotrophicum Is by the Diaminopimelic Acid Pathway. Curr. Microbiol. 1984, 10, 195–198. [Google Scholar] [CrossRef]
- Velasco, A.M.; Leguina, J.I.; Lazcano, A. Molecular Evolution of the Lysine Biosynthetic Pathways. J. Mol. Evol. 2002, 55, 445–449. [Google Scholar] [CrossRef]
- Ma, W.; Chen, K.; Li, Y.; Hao, N.; Wang, X.; Ouyang, P. Advances in Cadaverine Bacterial Production and Its Applications. Engineering 2017, 3, 308–317. [Google Scholar] [CrossRef]
- Samartzidou, H.; Mehrazin, M.; Xu, Z.; Benedik, M.J.; Delcour, A.H. Cadaverine Inhibition of Porin Plays a Role in Cell Survival at Acidic PH. J. Bacteriol. 2003, 185, 13–19. [Google Scholar] [CrossRef]
- Moreau, P.L. The Lysine Decarboxylase CadA Protects Escherichia Coli Starved of Phosphate against Fermentation Acids. J. Bacteriol. 2007, 189, 2249–2261. [Google Scholar] [CrossRef]
- Linares, D.M.; del Rio, B.; Redruello, B.; Ladero, V.; Martin, M.C.; Fernandez, M.; Ruas-Madiedo, P.; Alvarez, M.A. Comparative Analysis of the in Vitro Cytotoxicity of the Dietary Biogenic Amines Tyramine and Histamine. Food Chem. 2016, 197, 658–663. [Google Scholar] [CrossRef]
- del Rio, B.; Redruello, B.; Linares, D.M.; Ladero, V.; Fernandez, M.; Martin, M.C.; Ruas-Madiedo, P.; Alvarez, M.A. The Dietary Biogenic Amines Tyramine and Histamine Show Synergistic Toxicity towards Intestinal Cells in Culture. Food Chem. 2017, 218, 249–255. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No 2073/2005; Microbiological Criteria for Foodstuffs (Text with EEA Relevance). Online, 15 November 2005.
- FDA. FDA Fish and Fishery Products Hazards and Controls Guidance; US Department of Health and Human Services: Rockville, MD, USA, 2011.
- Shalaby, A.R. Significance of Biogenic Amines to Food Safety and Human Health. Food Res. Int. 1996, 29, 675–690. [Google Scholar] [CrossRef]
- Ladero, V.; Linares, D.M.; Pérez, M.; del Rio, B.; Fernández, M.; Alvarez, M.A.. Biogenic Amines in Dairy Products. In Microbial Toxins in Dairy Products; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 94–131. ISBN 978-1-118-82309-5. [Google Scholar]
- Kanjee, U.; Houry, W.A. Mechanisms of Acid Resistance in Escherichia Coli. Annu. Rev. Microbiol. 2013, 67, 65–81. [Google Scholar] [CrossRef]
- Perez, M.; Calles-Enríquez, M.; Nes, I.; Martin, M.C.; Fernandez, M.; Ladero, V.; Alvarez, M.A. Tyramine Biosynthesis Is Transcriptionally Induced at Low PH and Improves the Fitness of Enterococcus Faecalis in Acidic Environments. Appl. Microbiol. Biotechnol. 2015, 99, 3547–3558. [Google Scholar] [CrossRef]
- Diaz, M.; del Rio, B.; Ladero, V.; Redruello, B.; Fernández, M.; Martin, M.C.; Alvarez, M.A. Histamine Production in Lactobacillus Vaginalis Improves Cell Survival at Low PH by Counteracting the Acidification of the Cytosol. Int. J. Food Microbiol. 2020, 321, 108548. [Google Scholar] [CrossRef] [PubMed]
- Linares, D.M.; del Río, B.; Ladero, V.; Martínez, N.; Fernández, M.; Martín, M.C.; Alvarez, M.A. Factors Influencing Biogenic Amines Accumulation in Dairy Products. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M.; Pardo, I.; Ferrer, S. Regulation of Hdc Expression and HDC Activity by Enological Factors in Lactic Acid Bacteria. J. Appl. Microbiol. 2008, 105, 1544–1551. [Google Scholar] [CrossRef] [PubMed]
- McDonald, A.D.; Perkins, L.J.; Buller, A.R. Facile in Vitro Biocatalytic Production of Diverse Tryptamines. ChemBioChem 2019, 20, 1939–1944. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on Risk Based Control of Biogenic Amine Formation in Fermented Foods. EFSA J 2011, 9, 2393. [Google Scholar] [CrossRef]
- Wüst, N.; Rauscher-Gabernig, E.; Steinwider, J.; Bauer, F.; Paulsen, P. Risk Assessment of Dietary Exposure to Tryptamine for the Austrian Population. Food Addit. Contam. Part A 2017, 34, 404–420. [Google Scholar] [CrossRef]
- Rodriguez, M.B.R.; Carneiro, C.D.S.; Feijó, M.B.D.S.; Júnior, C.A.C.; Mano, S.B. Bioactive Amines: Aspects of Quality and Safety in Food. Food Nutr. Sci. 2014, 2014, 42092. [Google Scholar] [CrossRef]
- Stratton, J.E.; HUTKINS, R.W.; TAYLOR, S.L. Biogenic Amines in Cheese and Other Fermented Foods: A Review. J. Food Prot. 1991, 54, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, H.; Wechsler, D.; Irmler, S. Production of Putrescine and Cadaverine by Paucilactobacillus Wasatchensis. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Sudalaimani, S.; Esokkiya, A.; Hansda, S.; Suresh, C.; Tamilarasan, P.; Giribabu, K. Colorimetric Sensing of Putrescine and Cadaverine Using Ninhydrin as a Food Spoilage Detection Reagent. Food Anal. Methods 2020, 13, 629–636. [Google Scholar] [CrossRef]
- Zhang, G.; Loch, A.S.; Kistemaker, J.C.M.; Burn, P.L.; Shaw, P.E. Dicyanovinyl-Based Fluorescent Sensors for Dual Mechanism Amine Sensing. J. Mater. Chem. C 2020, 8, 13723–13732. [Google Scholar] [CrossRef]
- Santos, M.H.S. Biogenic Amines: Their Importance in Foods. Int. J. Food Microbiol. 1996, 29, 213–231. [Google Scholar] [CrossRef] [PubMed]
- Leroy, F.; De Vuyst, L. Bacteriocins of Lactic Acid Bacteria to Combat Undesirable Bacteria in Dairy Products. Aust. J. Dairy Technol. 2010, 65, 143–149. [Google Scholar]
- Andersen, G.; Marcinek, P.; Sulzinger, N.; Schieberle, P.; Krautwurst, D. Food Sources and Biomolecular Targets of Tyramine. Nutr. Rev. 2019, 77, 107–115. [Google Scholar] [CrossRef]
- Fernández, M.; Linares, D.M.; Rodríguez, A.; Alvarez, M.A. Factors Affecting Tyramine Production in Enterococcus Durans IPLA 655. Appl. Microbiol. Biotechnol. 2007, 73, 1400–1406. [Google Scholar] [CrossRef]
- Fernandez, M.; Linares, D.M.; Alvarez, M.A. Sequencing of the Tyrosine Decarboxylase Cluster of Lactococcus Lactis IPLA 655 and the Development of a PCR Method for Detecting Tyrosine Decarboxylating Lactic Acid Bacteria. J. Food Prot. 2004, 67, 2521–2529. [Google Scholar] [CrossRef]
- Coton, M.; Romano, A.; Spano, G.; Ziegler, K.; Vetrana, C.; Desmarais, C.; Lonvaud-Funel, A.; Lucas, P.; Coton, E. Occurrence of Biogenic Amine-Forming Lactic Acid Bacteria in Wine and Cider. Food Microbiol. 2010, 27, 1078–1085. [Google Scholar] [CrossRef] [PubMed]
- Bover-Cid, S.; Hugas, M.; Izquierdo-Pulido, M.; Vidal-Carou, M.C. Amino Acid-Decarboxylase Activity of Bacteria Isolated from Fermented Pork Sausages. Int. J. Food Microbiol. 2001, 66, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Bonetta, S.; Bonetta, S.; Carraro, E.; Coisson, J.D.; Travaglia, F.; Arlorio, M. Detection of Biogenic Amine Producer Bacteria in a Typical Italian Goat Cheese. J. Food Prot. 2008, 71, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Naila, A.; Flint, S.; Fletcher, G.; Bremer, P.; Meerdink, G. Control of Biogenic Amines in Food—Existing and Emerging Approaches. J. Food Sci. 2010, 75, R139–R150. [Google Scholar] [CrossRef]
- Marcobal, Á.; Martín-Álvarez, P.J.; Moreno-Arribas, M.V.; Muñoz, R. A Multifactorial Design for Studying Factors Influencing Growth and Tyramine Production of the Lactic Acid Bacteria Lactobacillus Brevis CECT 4669 and Enterococcus Faecium BIFI-58. Res. Microbiol. 2006, 157, 417–424. [Google Scholar] [CrossRef]
- Marcobal, A.; De Las Rivas, B.; Landete, J.M.; Tabera, L.; Muñoz, R. Tyramine and Phenylethylamine Biosynthesis by Food Bacteria. Crit. Rev. Food Sci. Nutr. 2012, 52, 448–467. [Google Scholar] [CrossRef]
- Cid, S.B.; Miguélez-Arrizado, M.J.; Becker, B.; Holzapfel, W.H.; Vidal-Carou, M.C. Amino Acid Decarboxylation by Lactobacillus Curvatus CTC273 Affected by the PH and Glucose Availability. Food Microbiol. 2008, 25, 269–277. [Google Scholar]
- CHANDER, H.; Batish, V.K.; Babu, S.; Singh, R.S. Factors Affecting Amine Production by a Selected Strain of Lactobacillus Bulgaricus. J. Food Sci. 1989, 54, 940–942. [Google Scholar] [CrossRef]
- Madeo, F.; Hofer, S.J.; Pendl, T.; Bauer, M.A.; Eisenberg, T.; Carmona-Gutierrez, D.; Kroemer, G. Nutritional Aspects of Spermidine. Annu. Rev. Nutr. 2020, 40, 135–159. [Google Scholar] [CrossRef]
- Muñoz-Esparza, N.C.; Latorre-Moratalla, M.L.; Comas-Basté, O.; Toro-Funes, N.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Polyamines in Food. Front. Nutr. 2019, 6, 108. [Google Scholar] [CrossRef]
- Zoumas-Morse, C.; Rock, C.L.; Quintana, E.L.; Neuhouser, M.L.; Gerner, E.W.; Meyskens Jr, F.L. Development of a Polyamine Database for Assessing Dietary Intake. J. Am. Diet. Assoc. 2007, 107, 1024–1027. [Google Scholar] [CrossRef]
- Yngve, A.; Ali, M.A.; Poortvliet, E.; Strömberg, R. Polyamines in Foods: Development of a Food Database. Food Nutr. Res. 2011. [Google Scholar]
- Eisenberg, T.; Abdellatif, M.; Schroeder, S.; Primessnig, U.; Stekovic, S.; Pendl, T.; Harger, A.; Schipke, J.; Zimmermann, A.; Schmidt, A. Cardioprotection and Lifespan Extension by the Natural Polyamine Spermidine. Nat. Med. 2016, 22, 1428–1438. [Google Scholar] [CrossRef]
- Eisenberg, T.; Knauer, H.; Schauer, A.; Büttner, S.; Ruckenstuhl, C.; Carmona-Gutierrez, D.; Ring, J.; Schroeder, S.; Magnes, C.; Antonacci, L. Induction of Autophagy by Spermidine Promotes Longevity. Nat. Cell Biol. 2009, 11, 1305–1314. [Google Scholar] [CrossRef]
- Madeo, F.; Carmona-Gutierrez, D.; Hofer, S.J.; Kroemer, G. Caloric Restriction Mimetics against Age-Associated Disease: Targets, Mechanisms, and Therapeutic Potential. Cell Metab. 2019, 29, 592–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeo, F.; Eisenberg, T.; Pietrocola, F.; Kroemer, G. Spermidine in Health and Disease. Science 2018, 359, eaan2788. [Google Scholar] [CrossRef] [PubMed]
- Soda, K.; Dobashi, Y.; Kano, Y.; Tsujinaka, S.; Konishi, F. Polyamine-Rich Food Decreases Age-Associated Pathology and Mortality in Aged Mice. Exp. Gerontol. 2009, 44, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Yue, F.; Li, W.; Zou, J.; Jiang, X.; Xu, G.; Huang, H.; Liu, L. Spermidine Prolongs Lifespan and Prevents Liver Fibrosis and Hepatocellular Carcinoma by Activating MAP1S-Mediated AutophagySpermidine Activates Autophagy and Suppresses HCC via MAP1S. Cancer Res. 2017, 77, 2938–2951. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yin, J.; Zhang, L.; Qi, C.-C.; Ma, Z.-L.; Gao, L.-P.; Wang, D.-G.; Jing, Y.-H. Spermidine Preconditioning Ameliorates Laurate-Induced Brain Injury by Maintaining Mitochondrial Stability. Neurol. Res. 2017, 39, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Kumar, P.; Deshmukh, R. Neuroprotective Potential of Spermidine against Rotenone Induced Parkinson’s Disease in Rats. Neurochem. Int. 2018, 116, 104–111. [Google Scholar] [CrossRef]
- Guo, X.; Harada, C.; Namekata, K.; Kimura, A.; Mitamura, Y.; Yoshida, H.; Matsumoto, Y.; Harada, T. Spermidine Alleviates Severity of Murine Experimental Autoimmune Encephalomyelitis. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2696–2703. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Pech, U.; Bhukel, A.; Fulterer, A.; Ender, A.; Mauermann, S.F.; Andlauer, T.F.; Antwi-Adjei, E.; Beuschel, C.; Thriene, K. Spermidine Suppresses Age-Associated Memory Impairment by Preventing Adverse Increase of Presynaptic Active Zone Size and Release. PLoS Biol. 2016, 14, e1002563. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Scheunemann, L.; Eisenberg, T.; Mertel, S.; Bhukel, A.; Koemans, T.S.; Kramer, J.M.; Liu, K.S.; Schroeder, S.; Stunnenberg, H.G. Restoring Polyamines Protects from Age-Induced Memory Impairment in an Autophagy-Dependent Manner. Nat. Neurosci. 2013, 16, 1453–1460. [Google Scholar] [CrossRef]
- Saaid, M.; Saad, B.; Hashim, N.H.; Ali, A.S.M.; Saleh, M.I. Determination of Biogenic Amines in Selected Malaysian Food. Food Chem. 2009, 113, 1356–1362. [Google Scholar] [CrossRef]
- Silva, C.M.; Glória, M.B.A. Bioactive Amines in Chicken Breast and Thigh after Slaughter and during Storage at 4±1 C and in Chicken-Based Meat Products. Food Chem. 2002, 78, 241–248. [Google Scholar] [CrossRef]
- Valsamaki, K.; Michaelidou, A.; Polychroniadou, A. Biogenic Amine Production in Feta Cheese. Food Chem. 2000, 71, 259–266. [Google Scholar] [CrossRef]
- Ziegler, W.; Hahn, M.; Wallnöfer, P.R. Changes in Biogenic Amine Contents during Processing of Several Plant Foods. Dtsch. Lebensm.-Rundsch. 1994, 90, 108–112. [Google Scholar]
- Okamoto, A.; Sugi, E.; Koizumi, Y.; Yanagida, F.; Udaka, S. Polyamine Content of Ordinary Foodstuffs and Various Fermented Foods. Biosci. Biotechnol. Biochem. 1997, 61, 1582–1584. [Google Scholar] [CrossRef]
- Novella-Rodríguez, S.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Biogenic Amines and Polyamines in Milks and Cheeses by Ion-Pair High Performance Liquid Chromatography. J. Agric. Food Chem. 2000, 48, 5117–5123. [Google Scholar] [CrossRef]
- Novella-Rodríguez, S.; Veciana-Nogués, M.T.; Izquierdo-Pulido, M.; Vidal-Carou, M.C. Distribution of Biogenic Amines and Polyamines in Cheese. J. Food Sci. 2003, 68, 750–756. [Google Scholar] [CrossRef]
- Nishimura, K.; Shiina, R.; Kashiwagi, K.; Igarashi, K. Decrease in Polyamines with Aging and Their Ingestion from Food and Drink. J. Biochem. (Tokyo) 2006, 139, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Nishibori, N.; Fujihara, S.; Akatuki, T. Amounts of Polyamines in Foods in Japan and Intake by Japanese. Food Chem. 2007, 100, 491–497. [Google Scholar] [CrossRef]
- Moret, S.; Smela, D.; Populin, T.; Conte, L.S. A Survey on Free Biogenic Amine Content of Fresh and Preserved Vegetables. Food Chem. 2005, 89, 355–361. [Google Scholar] [CrossRef]
- LoÈser, C. Polyamines in Human and Animal Milk. Br. J. Nutr. 2000, 84, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Lavizzari, T.; Veciana-Nogués, M.T.; Bover-Cid, S.; Mariné-Font, A.; Vidal-Carou, M.C. Improved Method for the Determination of Biogenic Amines and Polyamines in Vegetable Products by Ion-Pair High-Performance Liquid Chromatography. J. Chromatogr. A 2006, 1129, 67–72. [Google Scholar] [CrossRef]
- Larqué, E.; Sabater-Molina, M.; Zamora, S. Biological Significance of Dietary Polyamines. Nutrition 2007, 23, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Kozová, M.; Kalač, P.; Pelikánová, T. Contents of Biologically Active Polyamines in Chicken Meat, Liver, Heart and Skin after Slaughter and Their Changes during Meat Storage and Cooking. Food Chem. 2009, 116, 419–425. [Google Scholar] [CrossRef]
- Kalač, P.; Švecová, S.; Pelikánová, T. Levels of Biogenic Amines in Typical Vegetable Products. Food Chem. 2002, 77, 349–351. [Google Scholar] [CrossRef]
- Kalač, P.; Křížek, M.; Pelikánová, T.; Langová, M.; Veškrna, O. Contents of Polyamines in Selected Foods. Food Chem. 2005, 90, 561–564. [Google Scholar] [CrossRef]
- Kalac, P.; Krausová, P. A Review of Dietary Polyamines: Formation, Implications for Growth and Health and Occurrence in Foods. Food Chem. 2005, 90, 219–230. [Google Scholar] [CrossRef]
- Kalac, P. Recent Advances in the Research on Biological Roles of Dietary Polyamines in Man. J. Appl. Biomed. 2009, 7. [Google Scholar] [CrossRef]
- Eliassen, K.A.; Reistad, R.; Risøen, U.; Rønning, H.F. Dietary Polyamines. Food Chem. 2002, 78, 273–280. [Google Scholar] [CrossRef]
- Cipolla, B.G.; Havouis, R.; Moulinoux, J.-P. Polyamine Contents in Current Foods: A Basis for Polyamine Reduced Diet and a Study of Its Long Term Observance and Tolerance in Prostate Carcinoma Patients. Amino Acids 2007, 33, 203–212. [Google Scholar] [CrossRef]
- Binh, P.N.T.; Soda, K.; Maruyama, C.; Kawakami, M. Relationship between Food Polyamines and Gross Domestic Product in Association with Longevity in Asian Countries. Health (N. Y.) 2010, 2, 1390. [Google Scholar] [CrossRef]
- Bardócz, S.; Grant, G.; Brown, D.S.; Ralph, A.; Pusztai, A. Polyamines in Food—Implications for Growth and Health. J. Nutr. Biochem. 1993, 4, 66–71. [Google Scholar] [CrossRef]
- Bardócz, S.; Duguid, T.J.; Brown, D.S.; Grant, G.; Pusztai, A.; White, A.; Ralph, A. The Importance of Dietary Polyamines in Cell Regeneration and Growth. Br. J. Nutr. 1995, 73, 819–828. [Google Scholar] [CrossRef] [Green Version]
- Bardócz, S. Polyamines in Food and Their Consequences for Food Quality and Human Health. Trends Food Sci. Technol. 1995, 6, 341–346. [Google Scholar] [CrossRef]
- Metekia, W.A.; Ulusoy, B.H.; Habte-Tsion, H.-M. Spirulina Phenolic Compounds: Natural Food Additives with Antimicrobial Properties. Int. Food Res. J. 2021, 28. [Google Scholar] [CrossRef]
- Alshuniaber, M.A.; Krishnamoorthy, R.; AlQhtani, W.H. Antimicrobial Activity of Polyphenolic Compounds from Spirulina against Food-Borne Bacterial Pathogens. Saudi J. Biol. Sci. 2021, 28, 459–464. [Google Scholar] [CrossRef]
- Ould Bellahcen, T.; Cherki, M.; Sánchez, J.A.C.; Cherif, A.; EL Amrani, A. Chemical Composition and Antibacterial Activity of the Essential Oil of Spirulina Platensis from Morocco. J. Essent. Oil Bear. Plants 2019, 22, 1265–1276. [Google Scholar] [CrossRef]
- Kim, D.H.; Park, M.H.; Choi, Y.J.; Chung, K.W.; Park, C.H.; Jang, E.J.; An, H.J.; Yu, B.P.; Chung, H.Y. Molecular Study of Dietary Heptadecane for the Anti-Inflammatory Modulation of NF-KB in the Aged Kidney. PLoS ONE 2013, 8, e59316. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spirulina Samples | Fermentation | L-Glutamic Acid, mg/kg | Gamma-Aminobutyric Acid, mg/kg | |
|---|---|---|---|---|
| Duration, h | Conditions | |||
| Control (I) | - | 242 ± 14.8 f | 2.01 ± 0.18 a | |
| Control (II) | - | 2296 ± 59.3 k | 17.2 ± 0.20 c | |
| Lacticaseibacillus paracasei No. 244 | 24 h | SMF | 15.1 ± 1.30 a | 213 ± 15.0 l |
| 48 h | 41.7 ± 3.20 e | 287 ± 21.3 h | ||
| 24 h | SSF | 1784 ± 24.3 j | 2016 ± 46.5 j | |
| 48 h | 572 ± 21.2 i | 2396 ± 38.6 k | ||
| Levilactobacillus brevis No. 173 | 24 h | SMF | 357 ± 30.6 h | 12.0 ± 0.10 b |
| 48 h | 33.2 ± 2.10 d | 187 ± 10.3 g | ||
| 24 h | SSF | 3302 ± 44.9 n | 22.6 ± 1.32 d | |
| 48 h | 3841 ± 37.5 o | 58.8 ± 4.30 e | ||
| Leuconostoc mesenteroides No. 225 | 24 h | SMF | 23.4 ± 2.14 b | 170 ± 11.2 f,g |
| 48 h | 28.6 ± 2.12 c | 162 ± 12.1 f | ||
| 24 h | SSF | 2597 ± 51.8 l | 1264 ± 47.5 i | |
| 48 h | 3209 ± 43.0 n | 200 ± 15.0 g,h | ||
| Liquorilactobacillus uvarum No. 245 | 24 h | SMF | 356 ± 15.7 h | 53.6 ± 3.1 e |
| 48 h | 280 ± 11.0 g | 225 ± 11.2 l | ||
| 24 h | SSF | 2621 ± 58.4 l | 217 ± 8.9 l | |
| 48 h | 2908 ± 60.1 m | 165 ± 10.1 f | ||
| Spirulina Samples | Fermentation | Pathogenic and Opportunistic Bacterial Strain | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dura-tion, h | Condi-tions | Staphylococcus aureus | Escherichia coli | Acinetobacter baumannii | Staphylococcus haemolyticus | Salmonella enterica | Bacillus cereus | Proteus mirabilis | Klebsiella pneumoniae | Enterococcus faecium | Pseudomonas aeruginosa | |
| Diameter of the Inhibition Zone (DIZ), mm | ||||||||||||
| Control (I) | - | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Control (II) | - | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| No. 244 | 24 h | SMF | 12.0 ± 0.3 c | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 48 h | 10.1 ± 0.1 b | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| 24 h | SSF | 16.3 ± 0.2 e | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 48 h | 15.9 ± 0.3 e | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| No. 173 | 24 h | SMF | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 48 h | 13.1 ± 0.1 d | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| 24 h | SSF | 15.9 ± 0.3 e | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 48 h | 16.1 ± 0.2 e | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| No. 225 | 24 h | SMF | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 48 h | 12.2. ± 0.3 c | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| 24 h | SSF | 16.0 ± 0.4 e | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 48 h | 15.8 ± 0.3 e | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| No. 245 | 24 h | SMF | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 48 h | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| 24 h | SSF | 9.1 ± 0.3 a | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 48 h | 9.3 ± 0.4 b | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| pH | TRP | PUT | CAD | HIS | TYR | SPRMD | SPRM | GABA | LGlu | DIZ | LAB | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pH | r | 1 | −0.086 | −0.197 | 0.187 | 0.098 | 0.053 | −0.169 | −0.123 | 0.107 | −0.029 | 0.023 | −0.128 |
| p | 0.534 | 0.153 | 0.175 | 0.480 | 0.704 | 0.222 | 0.377 | 0.442 | 0.833 | 0.872 | 0.355 | ||
| TRP | r | −0.086 | 1 | 0.722 ** | 0.255 | 0.289 * | 0.314 * | 0.877 ** | 0.842 ** | 0.256 | 0.541** | −0.147 | −0.098 |
| p | 0.534 | 0.0001 | 0.062 | 0.034 | 0.021 | 0.0001 | 0.0001 | 0.061 | 0.0001 | 0.304 | 0.482 | ||
| PUT | r | −0.197 | 0.722 ** | 1 | 0.259 | 0.377 ** | 0.447 ** | 0.872 ** | 0.747 ** | 0.396 ** | 0.519 ** | 0.011 | 0.073 |
| p | 0.153 | 0.0001 | 0.059 | 0.005 | 0.001 | 0.0001 | 0.0001 | 0.003 | 0.0001 | 0.938 | 0.600 | ||
| CAD | r | 0.187 | 0.255 | 0.259 | 1 | 0.923 ** | 0.894 ** | 0.248 | 0.314 * | 0.531 ** | −0.124 | −0.142 | −0.282 * |
| p | 0.175 | 0.062 | 0.059 | 0.0001 | 0.0001 | 0.070 | 0.021 | 0.0001 | 0.372 | 0.322 | 0.039 | ||
| HIS | r | 0.098 | 0.289 * | 0.377 ** | 0.923 ** | 1 | 0.977 ** | 0.297 * | 0.306 * | 0.630 ** | −0.085 | −0.029 | −0.100 |
| p | 0.480 | 0.034 | 0.005 | 0.0001 | 0.0001 | 0.029 | 0.024 | 0.0001 | 0.539 | 0.840 | 0.472 | ||
| TYR | r | 0.053 | 0.314 * | 0.447 ** | 0.894 ** | 0.977 ** | 1 | 0.325 * | 0.310 * | 0.656 ** | −0.065 | 0.050 | −0.0045 |
| p | 0.704 | 0.021 | 0.001 | 0.0001 | 0.0001 | 0.016 | 0.023 | 0.0001 | 0.640 | 0.727 | 0.747 | ||
| SPRMD | r | −0.169 | 0.877 ** | 0.872 ** | 0.248 | 0.297 * | 0.325 * | 1 | 0.941 ** | 0.322 * | 0.627 ** | −0.125 | −0.181 |
| p | 0.222 | 0.0001 | 0.0001 | 0.070 | 0.029 | 0.016 | 0.0001 | 0.018 | 0.0001 | 0.383 | 0.191 | ||
| SPRM | r | −0.123 | 0.842 ** | 0.747 ** | 0.314 * | 0.30 6* | 0.310 * | 0.941 ** | 1 | 0.317 * | 0.572 ** | −0.133 | −0.347 * |
| p | 0.377 | 0.0001 | 0.0001 | 0.021 | 0.024 | 0.023 | 0.0001 | 0.019 | 0.0001 | 0.351 | 0.010 | ||
| GABA | r | 0.107 | 0.256 | 0.396 ** | 0.531 ** | 0.630 ** | 0.656 ** | 0.322 * | 0.317 * | 1 | 0.163 | 0.337 * | −0.055 |
| p | 0.442 | 0.061 | 0.003 | 0.0001 | 0.0001 | 0.0001 | 0.018 | 0.019 | 0.240 | 0.016 | 0.691 | ||
| LGlu | r | −0.029 | 0.541 ** | 0.519 ** | −0.124 | −0.085 | −0.065 | 0.627 ** | 0.572 ** | 0.163 | 1 | −0.099 | 0.007 |
| p | 0.833 | 0.0001 | 0.0001 | 0.372 | 0.539 | 0.640 | 0.0001 | 0.0001 | 0.240 | 0.489 | 0.960 | ||
| DIZ | r | 0.023 | −0.147 | 0.011 | −0.142 | −0.029 | 0.050 | −0.125 | −0.133 | 0.337 * | −0.099 | 1 | 0.027 |
| p | 0.872 | 0.304 | 0.938 | 0.322 | 0.840 | 0.727 | 0.383 | 0.351 | 0.016 | 0.489 | 0.853 | ||
| LAB count | r | −0.128 | −0.098 | 0.073 | −0.282 * | −0.100 | −0.045 | −0.181 | −0.347 * | −0.055 | 0.007 | 0.027 | 1 |
| p | 0.355 | 0.482 | 0.600 | 0.039 | 0.472 | 0.747 | 0.191 | 0.010 | 0.691 | 0.960 | 0.853 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tolpeznikaite, E.; Bartkevics, V.; Skrastina, A.; Pavlenko, R.; Ruzauskas, M.; Starkute, V.; Zokaityte, E.; Klupsaite, D.; Ruibys, R.; Rocha, J.M.; et al. Submerged and Solid-State Fermentation of Spirulina with Lactic Acid Bacteria Strains: Antimicrobial Properties and the Formation of Bioactive Compounds of Protein Origin. Biology 2023, 12, 248. https://doi.org/10.3390/biology12020248
Tolpeznikaite E, Bartkevics V, Skrastina A, Pavlenko R, Ruzauskas M, Starkute V, Zokaityte E, Klupsaite D, Ruibys R, Rocha JM, et al. Submerged and Solid-State Fermentation of Spirulina with Lactic Acid Bacteria Strains: Antimicrobial Properties and the Formation of Bioactive Compounds of Protein Origin. Biology. 2023; 12(2):248. https://doi.org/10.3390/biology12020248
Chicago/Turabian StyleTolpeznikaite, Ernesta, Vadims Bartkevics, Anna Skrastina, Romans Pavlenko, Modestas Ruzauskas, Vytaute Starkute, Egle Zokaityte, Dovile Klupsaite, Romas Ruibys, João Miguel Rocha, and et al. 2023. "Submerged and Solid-State Fermentation of Spirulina with Lactic Acid Bacteria Strains: Antimicrobial Properties and the Formation of Bioactive Compounds of Protein Origin" Biology 12, no. 2: 248. https://doi.org/10.3390/biology12020248