
Anticancer Drugs Paclitaxel, Carboplatin, Doxorubicin, and Cyclophosphamide Alter the Biophysical Characteristics of Red Blood Cells, In Vitro
 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of RBC’s Suspensions and Their Treatment
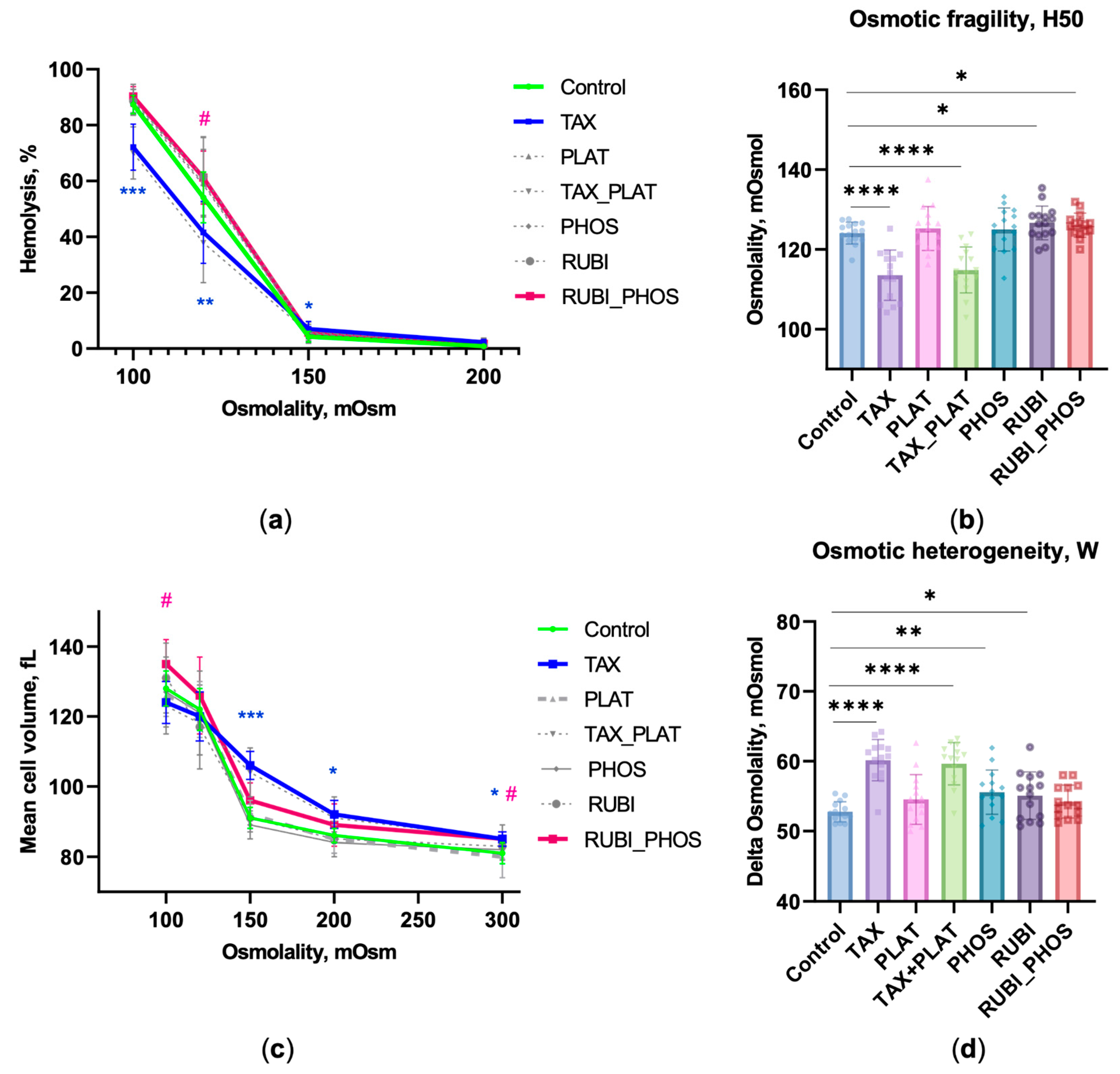
2.2. Osmotic Fragility Test
- H50, mOsmol—the buffer osmolality, at which 50% of cells were lysed.
- W, mOsmol—the distribution width of the osmotic resistance curve, at which 90% and 10% lysis occurs (W = H90–H10, mOsmol). It is a characteristic of the heterogeneity of the RBCs’ pool.
- MCVosm, fL—the hydrodynamic cell volume versus the buffer osmolality. The MCVosm curve was normalised to the MCV value at 300 mOsmol measured by the hematological analyser.
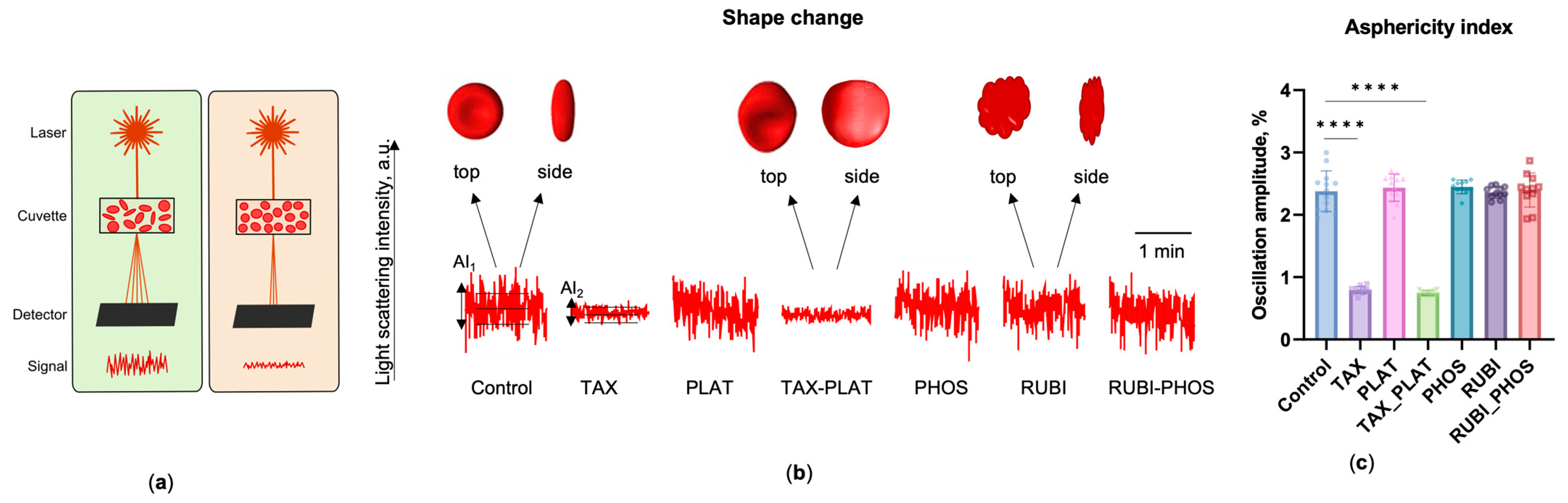
- The asphericity index, AI, %, which is the normalised amplitude of the SLI oscillations of the RBCs in the buffer with physiological osmolality (300 mOsmol). This index is proportional to the shape asymmetry of the cells and can be used to distinguish the normal discoid and spherical shape of RBCs.
2.3. Flow Cytometry Analysis
2.4. Confocal Microscopy
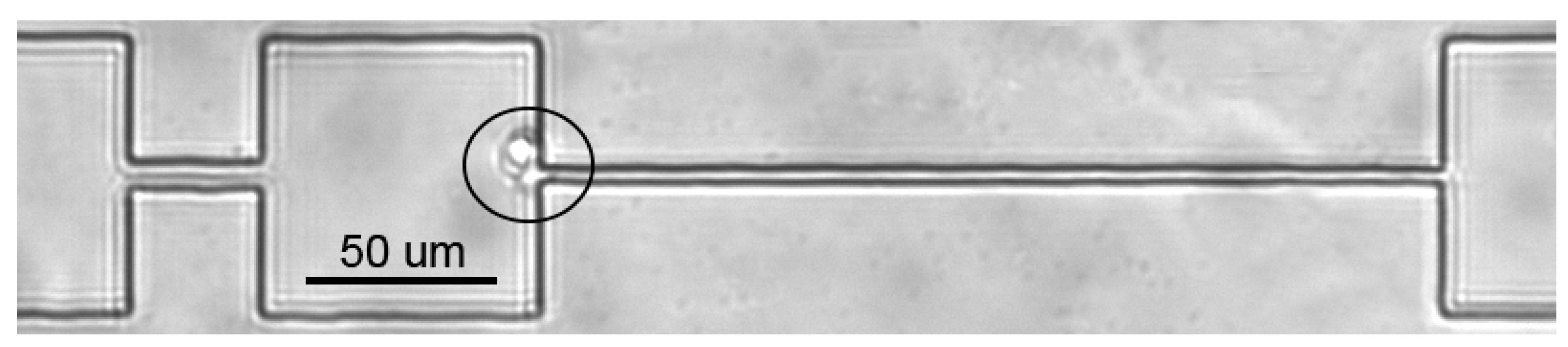
2.5. Microfluidic Analysis
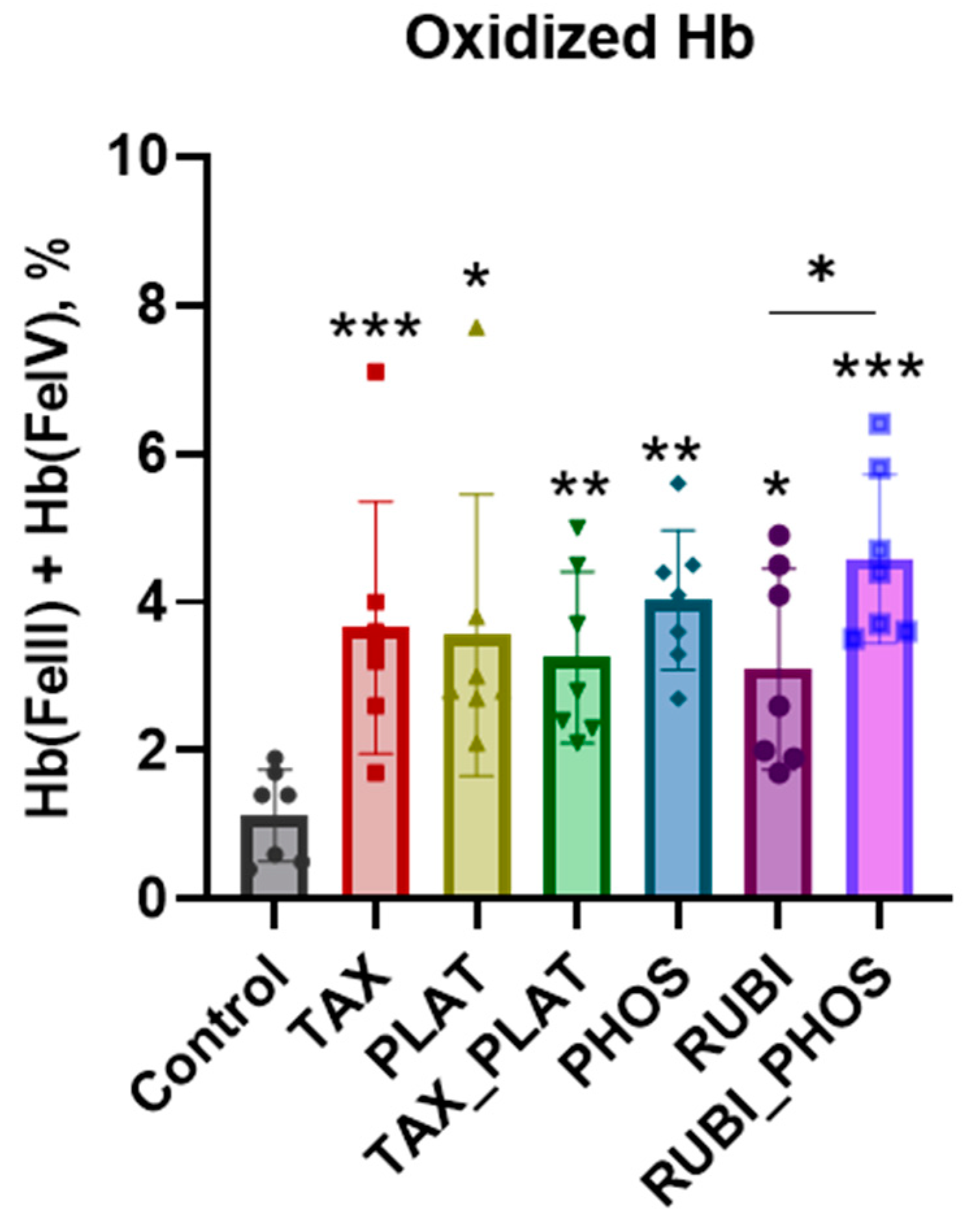
2.6. Free Hemoglobin and Hemoglobin Species Calculation
2.7. Statistics
3. Results
3.1. Osmotic Fragility Test Based on Laser Diffractometry
3.1.1. OFT: CT Drugs Impair RBCs’ Osmotic Resistance
3.1.2. OFT: CT Drugs Compromise the Ability of RBCs to Maintain Hydrodynamic Volume
3.1.3. OFT: CT Drugs Increase the Heterogeneity of the RBC Population
3.1.4. OFT: TAX and Its Combinations Disrupts the Discoid Shape of RBCs
3.2. CT Drugs Change the Hematological and Morphometric Characteristics of RBCs
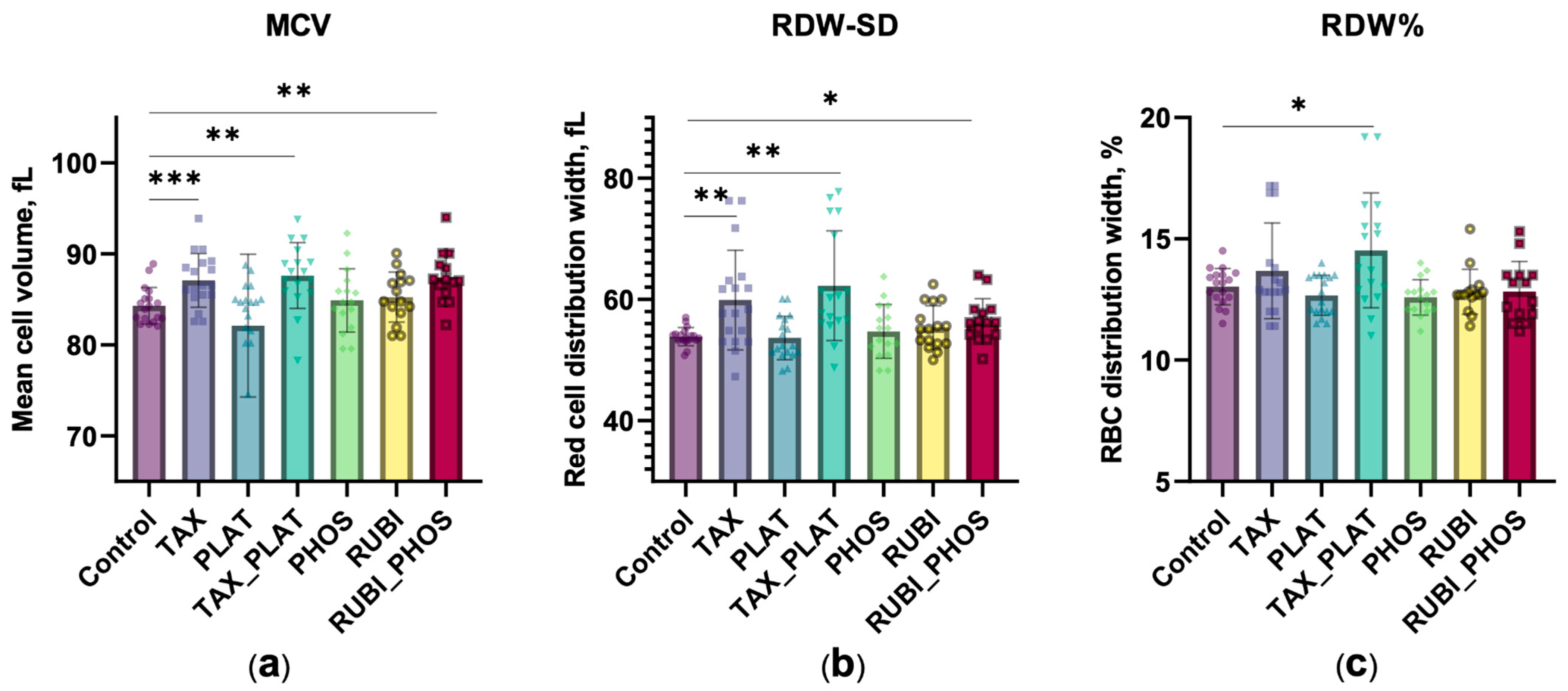
3.2.1. Hematological Analysis: CT Drugs Cause Increased Cell Volume and Population Heterogeneity
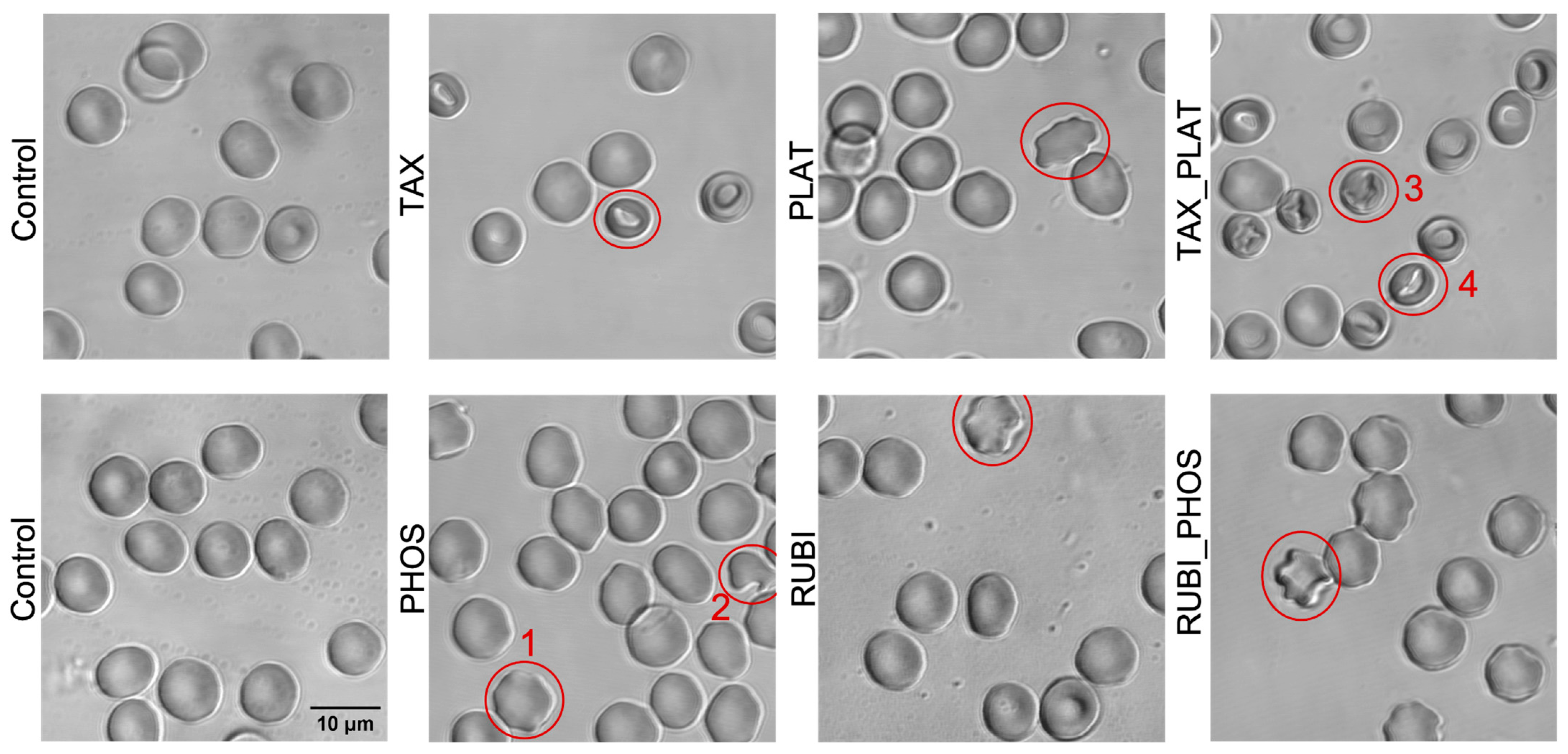
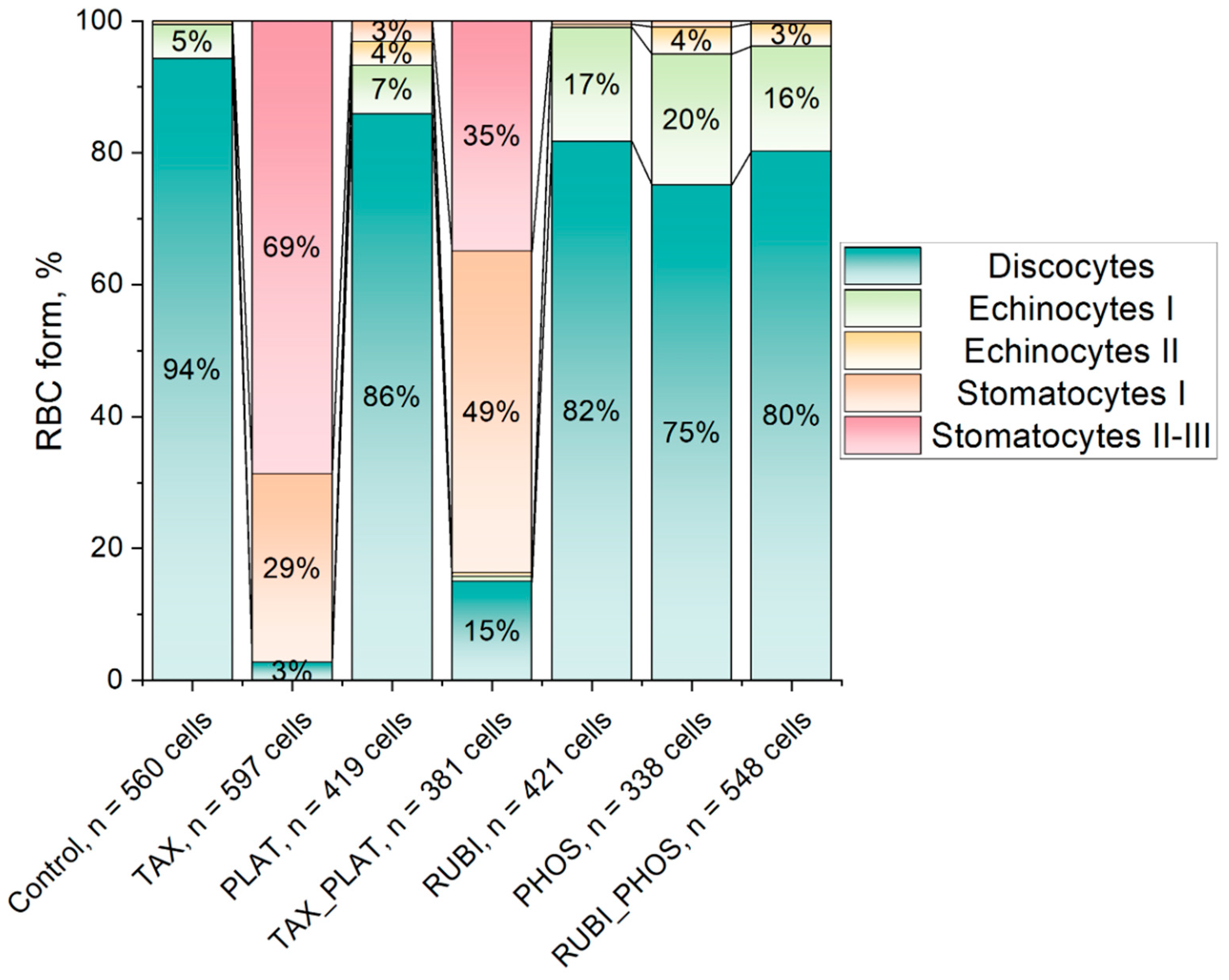
3.2.2. Confocal Microscopy: CT Drugs Change the Morphology of RBCs
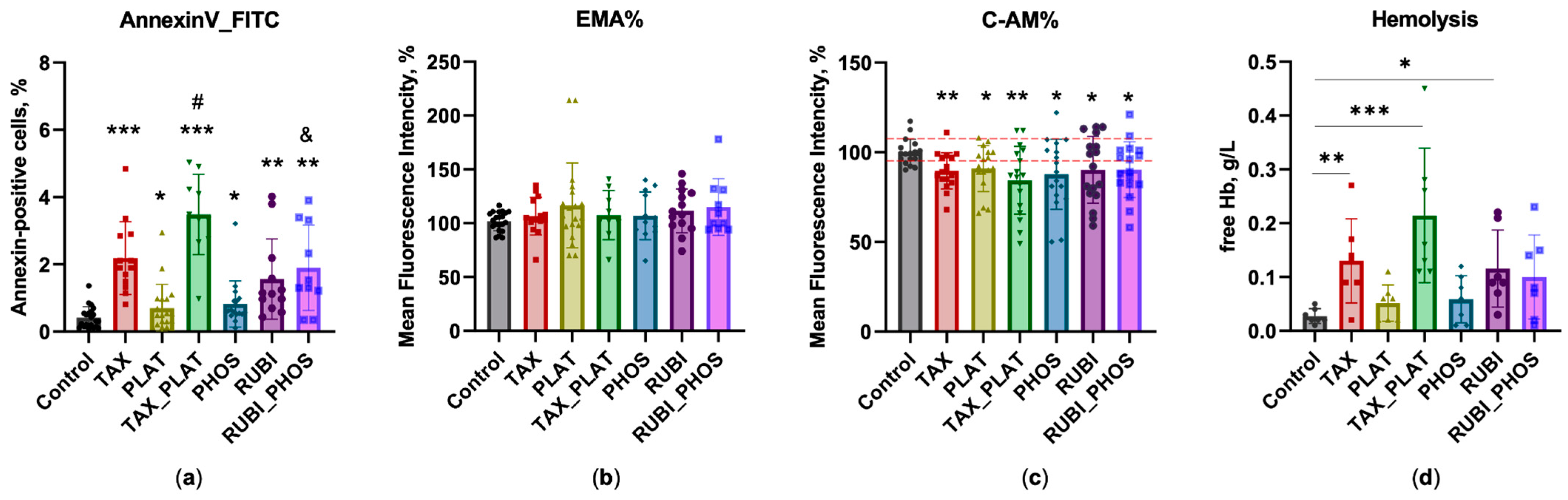
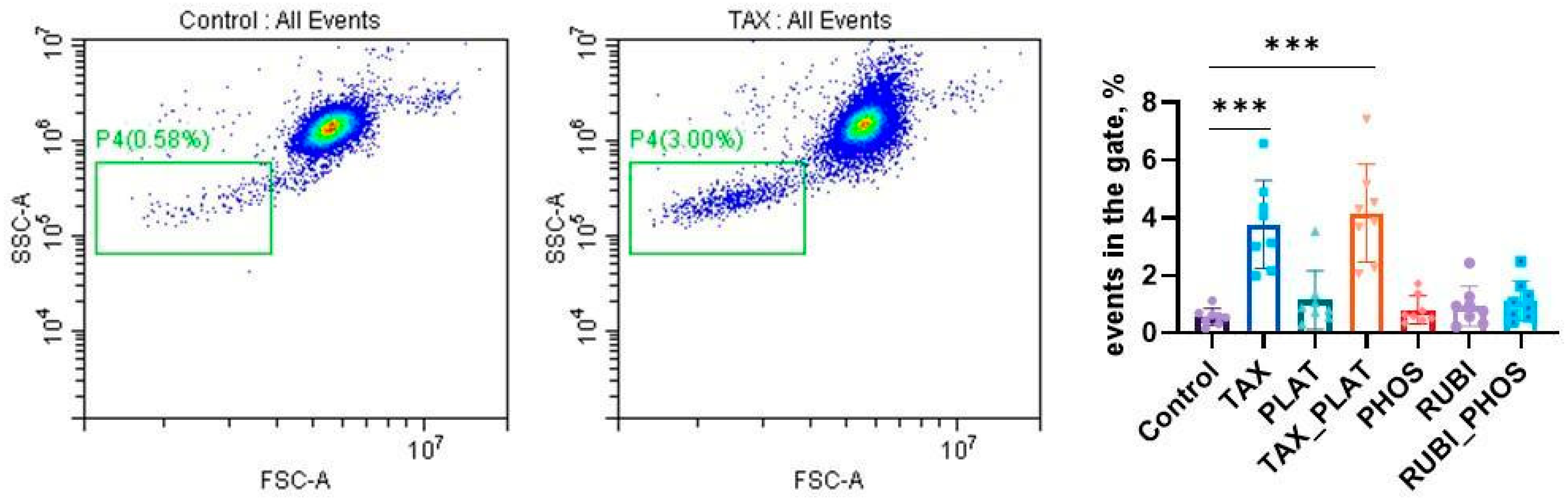
3.3. Flow Cytometry: Assessment of Membrane Transformation and Viability of RBCs
3.3.1. Annexin-Positive Cells Test
3.3.2. EMA Test: Action of Anticancer Drugs Does Not Cause Clustering of Band3 Membrane Complexes
3.3.3. Calcein-AM Vitality Test
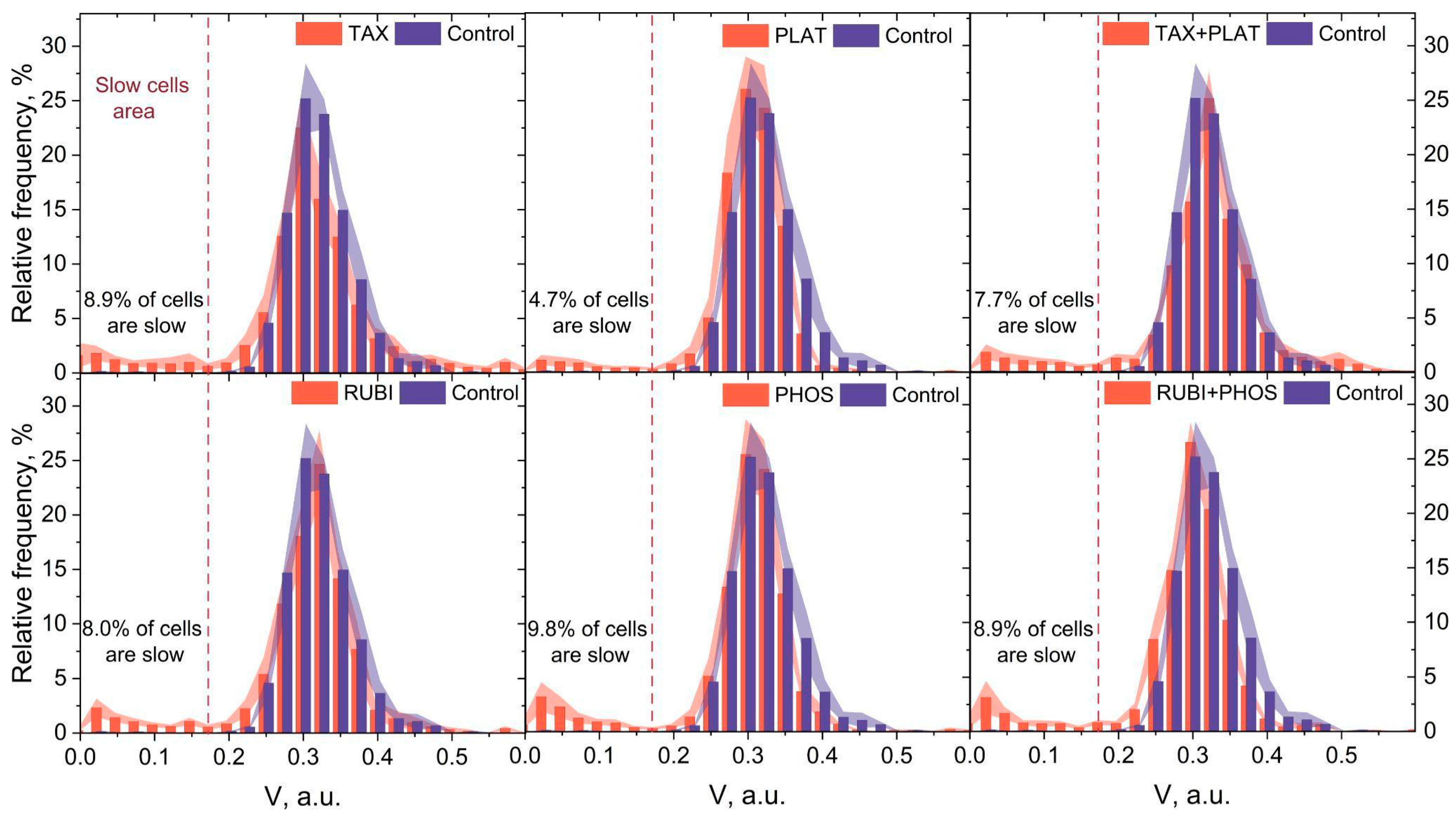
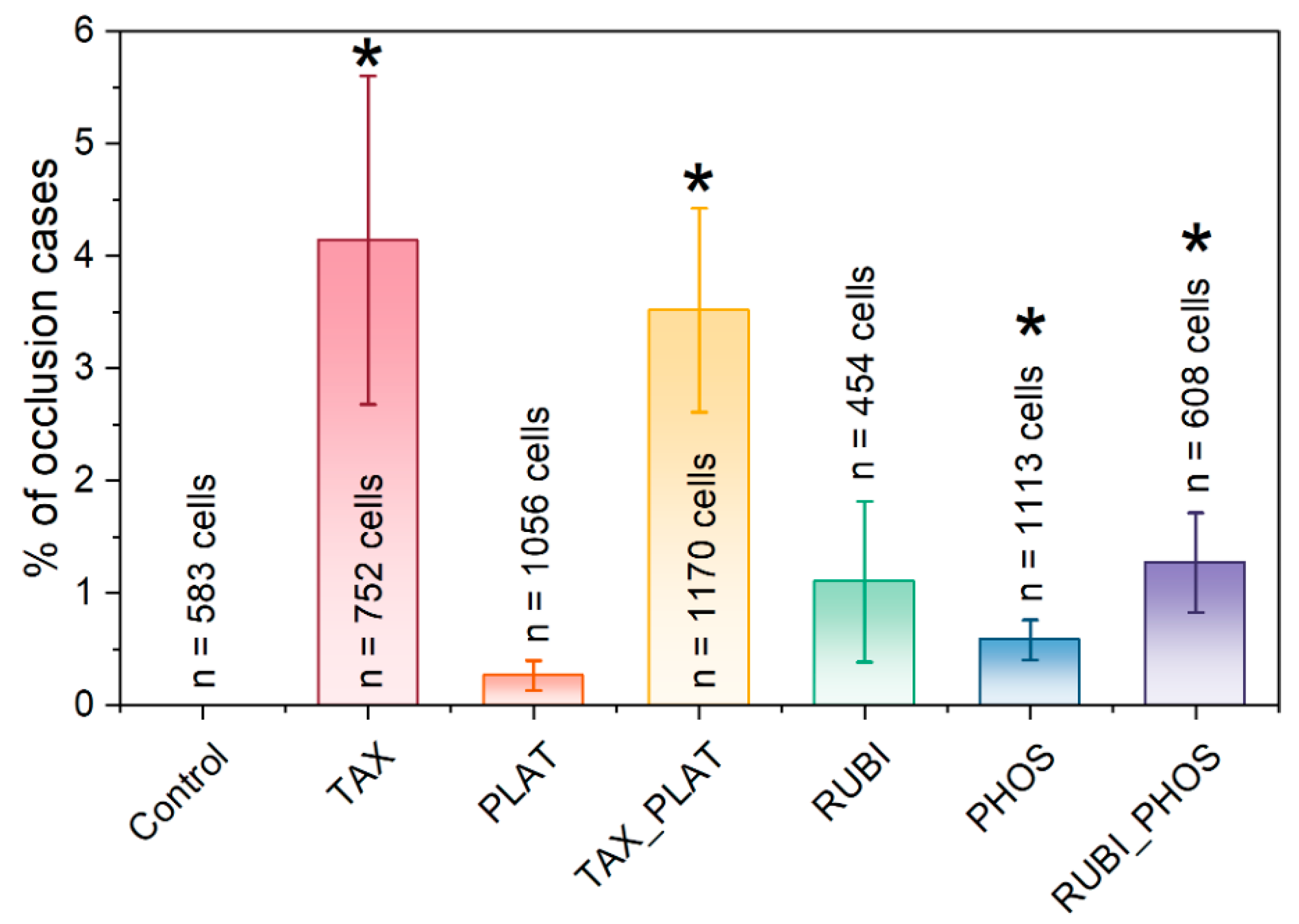
3.4. Microfluidic Analysis: CT Drugs Degrade RBCs’ Ability to Pass through the Microchannels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A





Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | TAX | PLAT | TAX_PLAT | PHOS | RUBI | RUBI_PHOS | |
|---|---|---|---|---|---|---|---|
| Hematology analysis | |||||||
| MCV, fL n = 15 | 83.8 ± 3.1 | 87.1 ± 2.9 p ≤ 0.0321 | 82.7 ± 7.6 p ≤ 0.416 | 87.0 ± 4.7 p ≤ 0.0106 | 84.9 ± 3.5 p ≤ 0.326 | 85.2 ± 2.8 p ≤ 0.156 | 86.7 ± 2.3 p ≤ 0.0345 |
| RDW-SD, fL n = 15 | 53.9 ± 1.5 | 59.9 ± 8.2 p ≤ 0.007 | 53.7 ± 3.6 p ≤ 0.824 | 62.3 ± 9.0 p ≤ 0.0001 | 54.7 ± 4.4 p ≤ 0.465 | 55.4 ± 3.6 p ≤ 0.140 | 56.5 ± 3.6 p ≤ 0.047 |
| RDW-CV, % n = 15 | 13.0 ± 0.8 | 13.7 ± 2.0 p ≤ 0.208 | 12.7 ± 2.4 p ≤ 0.178 | 14.5 ± 0.7 p ≤ 0.023 | 12.6 ± 0.9 p ≤ 0.090 | 12.8 ± 0.9 p ≤ 0.487 | 12.9 ± 1.2 p ≤ 0.592 |
| Osmotic Fragility Test | |||||||
| H50, mOsmol n = 15 | 123.9 ± 2.9 | 119.5 ± 3.9 p ≤ 0.001 | 124.6 ± 4.4 p ≤ 0.521 | 115.1 ± 5.7 p ≤ 0.000 | 124.6 ± 5.1 p ≤ 0.610 | 125.9 ± 3.5 p ≤ 0.037 | 126.1 ± 3.0 p ≤ 0.054 |
| W, mOsmol n = 15 | 52.8 ± 1.5 | 60.2 ± 2.9 p ≤ 0.000 | 54.3 ± 3.7 p ≤ 0.341 | 59.7 ± 3.0 p ≤ 0.000 | 55.7 ± 3.0 p ≤ 0.0345 | 54.7 ± 3.7 p ≤ 0.0492 | 55.2 ± 4.4 p ≤ 0.0324 |
| Asphericity Index, a.u. n = 15 | 2.36 ± 0.32 | 0.79 ± 0.06 p ≤ 0.000 | 2.37 ± 0.33 p ≤ 0.967 | 0.76 ± 0.04 p ≤ 0.000 | 2.39 ± 0.23 p ≤ 0.817 | 2.32 ± 0.17 p ≤ 0.628 | 2.34 ± 0.32 p ≤ 0.868 |
| Hemolysis, % 100 mOsmol. n = 15 | 86.5 ± 3.1 | 72.1 ± 8.3 p ≤ 0.000 | 89.2 ± 5.4 p ≤ 0.753 | 70.0 ± 9.4 p ≤ 0.000 | 88.2 ± 4.7 p ≤ 0.938 | 88.6 ± 4.2 p ≤ 0.879 | 90.6 ± 3.1 p ≤ 0.0352 |
| Spectrophotometric analysis | |||||||
| Free Hb, % n = 7 | 0.03 ± 0.01 | 0.13 ± 0.08 p ≤ 0.0208 | 0.05 ± 0.03 p ≤ 0.09 | 0.21 ± 0.13 p ≤ 0.001 | 0.06 ± 0.04 p ≤ 0.10 | 0.12 ± 0.07 p ≤ 0.02 | 0.10 ± 0.08 p ≤ 0.0493 |
| Microfluidic analysis | |||||||
| Slow cells, % | 0.45 ± 0.20 | 8.9 ± 3.29 | 4.7 ± 1.88 | 7.7 ± 2.39 | 9.8 ± 3.76 | 8.0 ± 2.10 | 8.9 ± 3.30 |
| Number of analysed cells | 5386 | 4379 | 6389 | 5188 | 5010 | 5420 | 4936 |
| Occlusions, % n = 3 | 0 ± 0 | 4.14 ± 1.46 p ≤ 0.0472 | 0.26539 ± 0.13 p ≤ 0.1193 | 3.51702 ± 0.91 p ≤ 0.0179 | 1.10036 ± 0.71 p ≤ 0.0305 | 0.58389 ± 0.18 p ≤ 0.1747 | 1.27221 ± 0.44 p ≤ 0.0452 |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Abbas, Z.; Rehman, S. An Overview of Cancer Treatment Modalities. In Neoplasm; Shahzad, H.N., Ed.; InTech: Rang-Du-Fliers, France, 2018; ISBN 978-1-78923-777-1. [Google Scholar]
- Jordan, M.A.; Wilson, L. Microtubules as a Target for Anticancer Drugs. Nat. Rev. Cancer 2004, 4, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Kelland, L. The Resurgence of Platinum-Based Cancer Chemotherapy. Nat. Rev. Cancer 2007, 7, 573–584. [Google Scholar] [CrossRef]
- Minotti, G.; Menna, P.; Salvatorelli, E.; Cairo, G.; Gianni, L. Anthracyclines: Molecular Advances and Pharmacologic Developments in Antitumor Activity and Cardiotoxicity. Pharmacol. Rev. 2004, 56, 185–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, R.; Necco, A.; Vailati, G.; Milzani, A. Dose-Dependence of Doxorubicin Effect on Actin Assembly in Vitro. Exp. Mol. Pathol. 1988, 49, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Alves, A.C.; Ribeiro, D.; Nunes, C.; Reis, S. Biophysics in Cancer: The Relevance of Drug-Membrane Interaction Studies. Biochim. Biophys. Acta (BBA)—Biomembr. 2016, 1858, 2231–2244. [Google Scholar] [CrossRef]
- Shinohara, K.; Tanaka, K.R. The Effects of Adriamycin (Doxorubicin HCL) on Human Red Blood Cells. Hemoglobin 1980, 4, 735–745. [Google Scholar] [CrossRef]
- Goodman, S.R.; Daescu, O.; Kakhniashvili, D.G.; Zivanic, M. The Proteomics and Interactomics of Human Erythrocytes. Exp. Biol. Med. 2013, 238, 509–518. [Google Scholar] [CrossRef]
- Amaiden, M.R.; Monesterolo, N.E.; Santander, V.S.; Campetelli, A.N.; Arce, C.A.; Pie, J.; Hope, S.I.; Vatta, M.S.; Casale, C.H. Involvement of Membrane Tubulin in Erythrocyte Deformability and Blood Pressure. J. Hypertens. 2012, 30, 1414–1422. [Google Scholar] [CrossRef]
- Chavez, J.D.; Keller, A.; Zhou, B.; Tian, R.; Bruce, J.E. Cellular Interactome Dynamics during Paclitaxel Treatment. Cell Rep. 2019, 29, 2371–2383.e5. [Google Scholar] [CrossRef] [Green Version]
- Ashrafuzzaman, M.; Tseng, C.-Y.; Tuszynski, J.A. Regulation of Channel Function Due to Physical Energetic Coupling with a Lipid Bilayer. Biochem. Biophys. Res. Commun. 2014, 445, 463–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashrafuzzaman, M.; Khan, Z.; Alqarni, A.; Alanazi, M.; Alam, M.S. Cell Surface Binding and Lipid Interactions behind Chemotherapy-Drug-Induced Ion Pore Formation in Membranes. Membranes 2021, 11, 501. [Google Scholar] [CrossRef] [PubMed]
- Harper, B.W.; Krause-Heuer, A.M.; Grant, M.P.; Manohar, M.; Garbutcheon-Singh, K.B.; Aldrich-Wright, J.R. Advances in Platinum Chemotherapeutics. Chemistry 2010, 16, 7064–7077. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.; Lee, S.; Kim, H.; Kang, H.; Youn, H.; Jo, S.; Youn, B.; Kim, H.Y. Revisiting Platinum-Based Anticancer Drugs to Overcome Gliomas. IJMS 2021, 22, 5111. [Google Scholar] [CrossRef] [PubMed]
- Marković, S.D.; Djačić, D.S.; Cvetković, D.M.; Obradović, A.D.; Žižić, J.B.; Ognjanović, B.I.; Štajn, A.Š. Effects of Acute In Vivo Cisplatin and Selenium Treatment on Hematological and Oxidative Stress Parameters in Red Blood Cells of Rats. Biol. Trace. Elem. Res. 2011, 142, 660–670. [Google Scholar] [CrossRef] [PubMed]
- Tolan, D.; Gandin, V.; Morrison, L.; El-Nahas, A.; Marzano, C.; Montagner, D.; Erxleben, A. Oxidative Stress Induced by Pt(IV) Pro-Drugs Based on the Cisplatin Scaffold and Indole Carboxylic Acids in Axial Position. Sci. Rep. 2016, 6, 29367. [Google Scholar] [CrossRef]
- Suwalsky, M.; Hernández, P.; Villena, F.; Sotomayor, C.P. The Anticancer Drug Cisplatin Interacts with the Human Erythrocyte Membrane. Z. Für Nat. C 2000, 55, 461–466. [Google Scholar] [CrossRef]
- Lü, C.; Yu, H.; Hou, J.; Zhou, J. Increased Procoagulant Activity of Red Blood Cells in the Presence of Cisplatin. Chin. Med. J. 2008, 121, 1775–1780. [Google Scholar] [CrossRef]
- Cavallasca, J.A.; Costa, C.A.; del Maliandi, M.R.; Contini, L.E.; Fernandez de Carrera, E.; Musuruana, J.L. Severe Infections in Patients With Autoimmune Diseases Treated With Cyclophosphamide. Reumatol. Clínica (Engl. Ed.) 2015, 11, 221–223. [Google Scholar] [CrossRef]
- Mills, K.A.; Chess-Williams, R.; McDermott, C. Novel Insights into the Mechanism of Cyclophosphamide-Induced Bladder Toxicity: Chloroacetaldehyde’s Contribution to Urothelial Dysfunction in Vitro. Arch. Toxicol. 2019, 93, 3291–3303. [Google Scholar] [CrossRef]
- Akamo, A.J.; Akinloye, D.I.; Ugbaja, R.N.; Adeleye, O.O.; Dosumu, O.A.; Eteng, O.E.; Antiya, M.C.; Amah, G.; Ajayi, O.A.; Faseun, S.O. Naringin Prevents Cyclophosphamide-Induced Erythrocytotoxicity in Rats by Abrogating Oxidative Stress. Toxicol. Rep. 2021, 8, 1803–1813. [Google Scholar] [CrossRef]
- Xu, H.; Xu, L.; Page, J.; Cannavale, K.; Sattayapiwat, O.; Rodriguez, R.; Chao, C. Incidence of Anemia in Patients Diagnosed with Solid Tumors Receiving Chemotherapy, 2010–2013. CLEP 2016, 8, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mark, M.; Walter, R.; Meredith, D.O.; Reinhart, W.H. Commercial Taxane Formulations Induce Stomatocytosis and Increase Blood Viscosity: Taxanes and Blood Rheology. Br. J. Pharmacol. 2001, 134, 1207–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vader, P.; Fens, M.H.; Sachini, N.; van Oirschot, B.A.; Andringa, G.; Egberts, A.C.; Gaillard, C.A.; Rasmussen, J.T.; van Wijk, R.; van Solinge, W.W.; et al. Taxol ® -Induced Phosphatidylserine Exposure and Microvesicle Formation in Red Blood Cells Is Mediated by Its Vehicle Cremophor ® EL. Nanomedicine 2013, 8, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Bryer, E.; Henry, D. Chemotherapy-Induced Anemia: Etiology, Pathophysiology, and Implications for Contemporary Practice. IJCTM 2018, 6, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, G.M.; Becker, P.S.; Blinder, M.; Cella, D.; Chanan-Khan, A.; Cleeland, C.; Coccia, P.F.; Djulbegovic, B.; Gilreath, J.A.; Kraut, E.H.; et al. Cancer- and Chemotherapy-Induced Anemia. J. Natl. Compr. Canc. Netw. 2012, 10, 628–653. [Google Scholar] [CrossRef] [Green Version]
- Tomaiuolo, G. Biomechanical Properties of Red Blood Cells in Health and Disease towards Microfluidics. Biomicrofluidics 2014, 8, 51501. [Google Scholar] [CrossRef] [Green Version]
- Aich, A.; Lamarre, Y.; Sacomani, D.P.; Kashima, S.; Covas, D.T.; de la Torre, L.G. Microfluidics in Sickle Cell Disease Research: State of the Art and a Perspective Beyond the Flow Problem. Front. Mol. Biosci. 2021, 7, 558982. [Google Scholar] [CrossRef]
- Barshtein, G.; Pajic-Lijakovic, I.; Gural, A. Deformability of Stored Red Blood Cells. Front. Physiol. 2021, 12, 722896. [Google Scholar] [CrossRef]
- Besedina, N.A.; Skverchinskaya, E.A.; Shmakov, S.V.; Ivanov, A.S.; Mindukshev, I.V.; Bukatin, A.S. Persistent Red Blood Cells Retain Their Ability to Move in Microcapillaries under High Levels of Oxidative Stress. Commun. Biol. 2022, 5, 659. [Google Scholar] [CrossRef]
- Zhu, S.; Jiang, F.; Han, Y.; Xiang, N.; Ni, Z. Microfluidics for Label-Free Sorting of Rare Circulating Tumor Cells. Analyst 2020, 145, 7103–7124. [Google Scholar] [CrossRef] [PubMed]
- Gholizadeh, S.; Shehata Draz, M.; Zarghooni, M.; Sanati-Nezhad, A.; Ghavami, S.; Shafiee, H.; Akbari, M. Microfluidic Approaches for Isolation, Detection, and Characterization of Extracellular Vesicles: Current Status and Future Directions. Biosens. Bioelectron. 2017, 91, 588–605. [Google Scholar] [CrossRef] [Green Version]
- Mokhtari, R.B.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination Therapy in Combating Cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobbe, J.G.G.; Hardeman, M.R. Red Blood Cell Aggregation as Measured with the LORCA. Int. J. Artif. Organs 2006, 29, 641–642. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.R.; Mohandas, N.; Shohet, S.B. Osmotic Gradient Ektacytometry: Comprehensive Characterization of Red Cell Volume and Surface Maintenance. Blood 1983, 61, 899–910. [Google Scholar] [CrossRef] [Green Version]
- Walski, T.; Chludzińska, L.; Komorowska, M.; Witkiewicz, W. Individual Osmotic Fragility Distribution: A New Parameter for Determination of the Osmotic Properties of Human Red Blood Cells. BioMed. Res. Int. 2014, 2014, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Sudnitsyna, J.; Skverchinskaya, E.; Dobrylko, I.; Nikitina, E.; Gambaryan, S.; Mindukshev, I. Microvesicle Formation Induced by Oxidative Stress in Human Erythrocytes. Antioxidants 2020, 9, 929. [Google Scholar] [CrossRef]
- Mindukshev, I.; Gambaryan, S.; Kehrer, L.; Schuetz, C.; Kobsar, A.; Rukoyatkina, N.; Nikolaev, V.O.; Krivchenko, A.; Watson, S.P.; Walter, U.; et al. Low Angle Light Scattering Analysis: A Novel Quantitative Method for Functional Characterization of Human and Murine Platelet Receptors. Clin. Chem. Lab. Med. 2012, 50, 1253–1262. [Google Scholar] [CrossRef]
- Nobre, C.S.; Silva, J.A.; Jácomo, R.H.; Nery, L.F.A.; Barra, G.B. Flow Cytometric Analysis of Erythrocytes Osmotic Fragility in Hereditary Spherocytosis: A Case-Controlled Study Evaluating the Best Anticoagulant, Sample Pre-Treatment and NaCl Concentration for Reliable Screening of This Red Blood Cell Membrane Disorder. Cytometry 2018, 94, 910–917. [Google Scholar] [CrossRef] [Green Version]
- Besedina, N.A.; Skverchinskaya, E.A.; Ivanov, A.S.; Kotlyar, K.P.; Morozov, I.A.; Filatov, N.A.; Mindukshev, I.V.; Bukatin, A.S. Microfluidic Characterization of Red Blood Cells Microcirculation under Oxidative Stress. Cells 2021, 10, 3552. [Google Scholar] [CrossRef]
- Besedina, N.A.; Skverchinskaya, E.A.; Shmakov, S.V.; Ivanov, A.S.; Mindukshev, I.V.; Bukatin, A.S. Matlab Scripts for Calculation of Cell Velocity in Microfluidic Channel. Zenodo 2022. [Google Scholar] [CrossRef]
- Tarasev, M.; Chakraborty, S.; Alfano, K. RBC Mechanical Fragility as a Direct Blood Quality Metric to Supplement Storage Time. Mil. Med. 2015, 180, 150–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanias, T.; Acker, J.P. Mechanism of Hemoglobin-Induced Cellular Injury in Desiccated Red Blood Cells. Free. Radic. Biol. Med. 2010, 49, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Orbach, A.; Zelig, O.; Yedgar, S.; Barshtein, G. Biophysical and Biochemical Markers of Red Blood Cell Fragility. Transfus. Med. Hemother. 2017, 44, 183–187. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, N.; Kiss, F.; Miszti-Blasius, K. Interpretation of Osmotic Gradient Ektacytometry (Osmoscan) Data: A Comparative Study for Methodological Standards. Scand. J. Clin. Lab. Investig. 2015, 75, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Huisjes, R.; Bogdanova, A.; van Solinge, W.W.; Schiffelers, R.M.; Kaestner, L.; van Wijk, R. Squeezing for Life—Properties of Red Blood Cell Deformability. Front. Physiol. 2018, 9, 656. [Google Scholar] [CrossRef] [Green Version]
- Bessis, M. Red cell shapes. An illustrated classification and its rationale. In Red Cell Shape; Bessis, M., Weed, R.I., Leblond, P.F., Eds.; Springer: Berlin/Heidelberg, Germany, 1973; pp. 1–25. ISBN 978-3-642-88064-3. [Google Scholar]
- Mrowietz, C.; Hiebl, B.; Franke, R.P.; Park, J.-W.; Jung, F. Reversibility of Echinocyte Formation after Contact of Erythrocytes with Various Radiographic Contrast Media. Clin. Hemorheol. Microcirc. 2008, 39, 281–286. [Google Scholar] [CrossRef]
- Wong, P. An Explanation of the Reversal of Erythrocyte Echinocytosis by Incubation and Storage by Serum Albumin. Clin. Hemorheol. Microcirc. 2018, 68, 383–389. [Google Scholar] [CrossRef]
- Thiagarajan, P.; Parker, C.J.; Prchal, J.T. How Do Red Blood Cells Die? Front. Physiol. 2021, 12, 655393. [Google Scholar] [CrossRef]
- Arashiki, N.; Kimata, N.; Manno, S.; Mohandas, N.; Takakuwa, Y. Membrane Peroxidation and Methemoglobin Formation Are Both Necessary for Band 3 Clustering: Mechanistic Insights into Human Erythrocyte Senescence. Biochemistry 2013, 52, 5760–5769. [Google Scholar] [CrossRef] [Green Version]
- Gottlieb, Y.; Topaz, O.; Cohen, L.A.; Yakov, L.D.; Haber, T.; Morgenstern, A.; Weiss, A.; Chait Berman, K.; Fibach, E.; Meyron-Holtz, E.G. Physiologically Aged Red Blood Cells Undergo Erythrophagocytosis in Vivo but Not in Vitro. Haematologica 2012, 97, 994–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, L.J. A Band 3-Based Macrocomplex of Integral and Peripheral Proteins in the RBC Membrane. Blood 2003, 101, 4180–4188. [Google Scholar] [CrossRef] [PubMed]
- Burger, P.; de Korte, D.; van den Berg, T.K.; van Bruggen, R. CD47 in Erythrocyte Ageing and Clearance—the Dutch Point of View. Transfus. Med. Hemother. 2012, 39, 348–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantaleo, A.; Ferru, E.; Pau, M.C.; Khadjavi, A.; Mandili, G.; Mattè, A.; Spano, A.; De Franceschi, L.; Pippia, P.; Turrini, F. Band 3 Erythrocyte Membrane Protein Acts as Redox Stress Sensor Leading to Its Phosphorylation by p 72 Syk. Oxidative Med. Cell. Longev. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigra, A.D.; Santander, V.S.; Dircio-Maldonado, R.; Amaiden, M.R.; Monesterolo, N.E.; Flores-Guzmán, P.; Muhlberger, T.; Rivelli, J.F.; Campetelli, A.N.; Mayani, H.; et al. Tubulin Is Retained throughout the Human Hematopoietic/Erythroid Cell Differentiation Process and Plays a Structural Role in Sedimentable Fraction of Mature Erythrocytes. Int. J. Biochem. Cell Biol. 2017, 91, 29–36. [Google Scholar] [CrossRef]
- Gokhin, D.S.; Fowler, V.M. Feisty Filaments: Actin Dynamics in the Red Blood Cell Membrane Skeleton. Curr. Opin. Hematol. 2016, 23, 206–214. [Google Scholar] [CrossRef] [Green Version]
- Barvitenko, N.; Aslam, M.; Lawen, A.; Saldanha, C.; Skverchinskaya, E.; Uras, G.; Manca, A.; Pantaleo, A. Two Motors and One Spring: Hypothetic Roles of Non-Muscle Myosin II and Submembrane Actin-Based Cytoskeleton in Cell Volume Sensing. IJMS 2021, 22, 7967. [Google Scholar] [CrossRef]
- Dasari, S.; Bernard Tchounwou, P. Cisplatin in Cancer Therapy: Molecular Mechanisms of Action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [Green Version]
- Tikhomirova, I.A.; Muravyov, A.V.; Petrochenko, E.P.; Kislov, N.V.; Cheporov, S.V.; Peganova, E.V. Alteration of Red Blood Cell Microrheology by Anti-Tumor Chemotherapy Drugs. Biochem. Mosc. Suppl. Ser. A 2016, 10, 135–141. [Google Scholar] [CrossRef]
- Wagstaff, A.J.; Ward, A.; Benfield, P.; Heel, R.C. Carboplatin: A Preliminary Review of Its Pharmacodynamic and Pharmacokinetic Properties and Therapeutic Efficacy in the Treatment of Cancer. Drugs 1989, 37, 162–190. [Google Scholar] [CrossRef]
- Kuck, L.; McNamee, A.P.; Simmonds, M.J. Impact of Small Fractions of Abnormal Erythrocytes on Blood Rheology. Microvasc. Res. 2022, 139, 104261. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skverchinskaya, E.; Levdarovich, N.; Ivanov, A.; Mindukshev, I.; Bukatin, A. Anticancer Drugs Paclitaxel, Carboplatin, Doxorubicin, and Cyclophosphamide Alter the Biophysical Characteristics of Red Blood Cells, In Vitro. Biology 2023, 12, 230. https://doi.org/10.3390/biology12020230
Skverchinskaya E, Levdarovich N, Ivanov A, Mindukshev I, Bukatin A. Anticancer Drugs Paclitaxel, Carboplatin, Doxorubicin, and Cyclophosphamide Alter the Biophysical Characteristics of Red Blood Cells, In Vitro. Biology. 2023; 12(2):230. https://doi.org/10.3390/biology12020230
Chicago/Turabian StyleSkverchinskaya, Elisaveta, Nadezhda Levdarovich, Alexander Ivanov, Igor Mindukshev, and Anton Bukatin. 2023. "Anticancer Drugs Paclitaxel, Carboplatin, Doxorubicin, and Cyclophosphamide Alter the Biophysical Characteristics of Red Blood Cells, In Vitro" Biology 12, no. 2: 230. https://doi.org/10.3390/biology12020230