
Characterization and Expression Analysis of Four Cadmium-Tolerance-Associated Genes of Avicennia marina (Forsk.)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Cadmium Treatment
2.2. Identification of the Full-Length cDNA Sequences
2.3. Feature Prediction of Deduced Proteins
2.4. Quantitative Reverse Transcription Real-Time PCR (RT-qPCR)
2.5. Statistical Analysis
3. Results
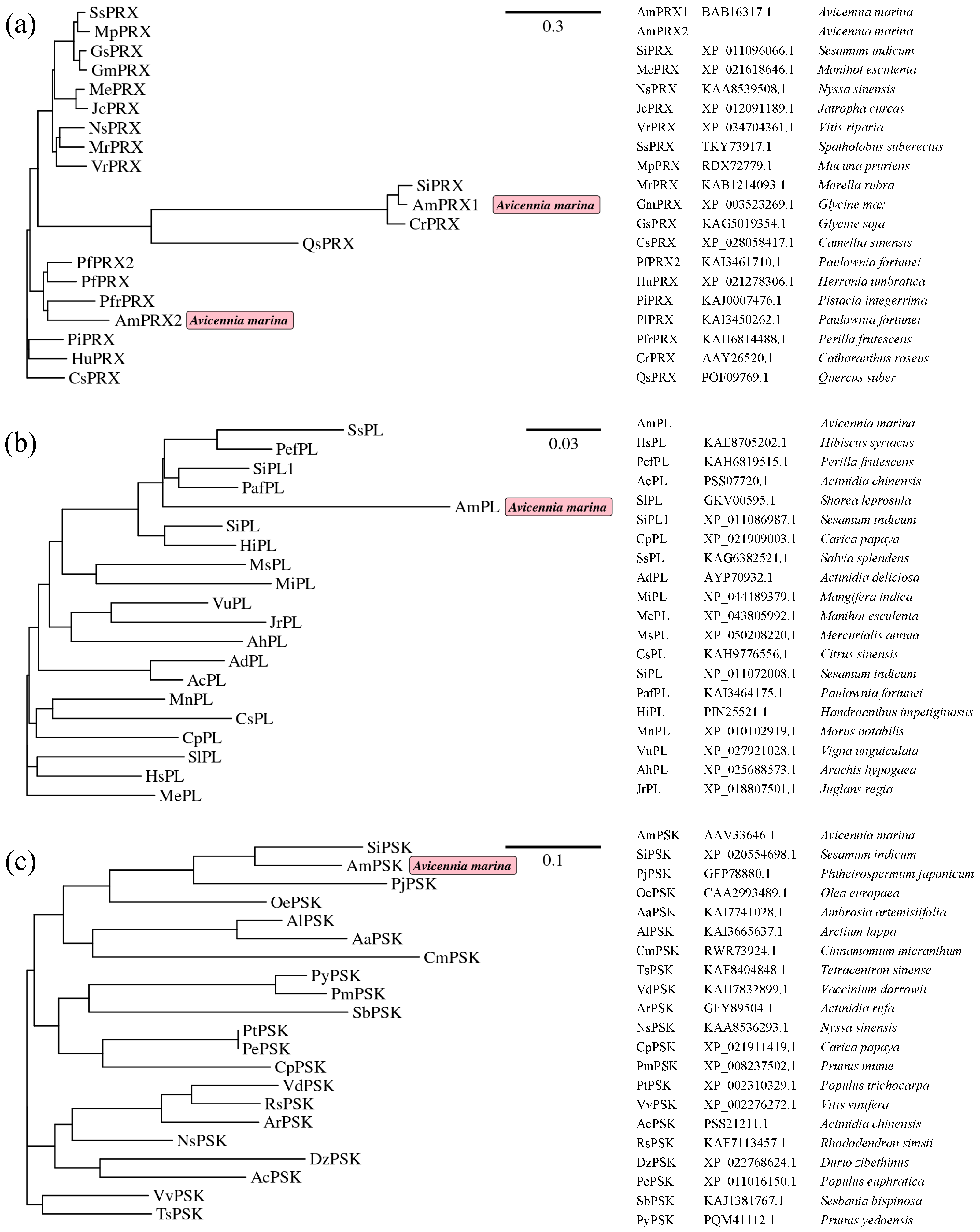
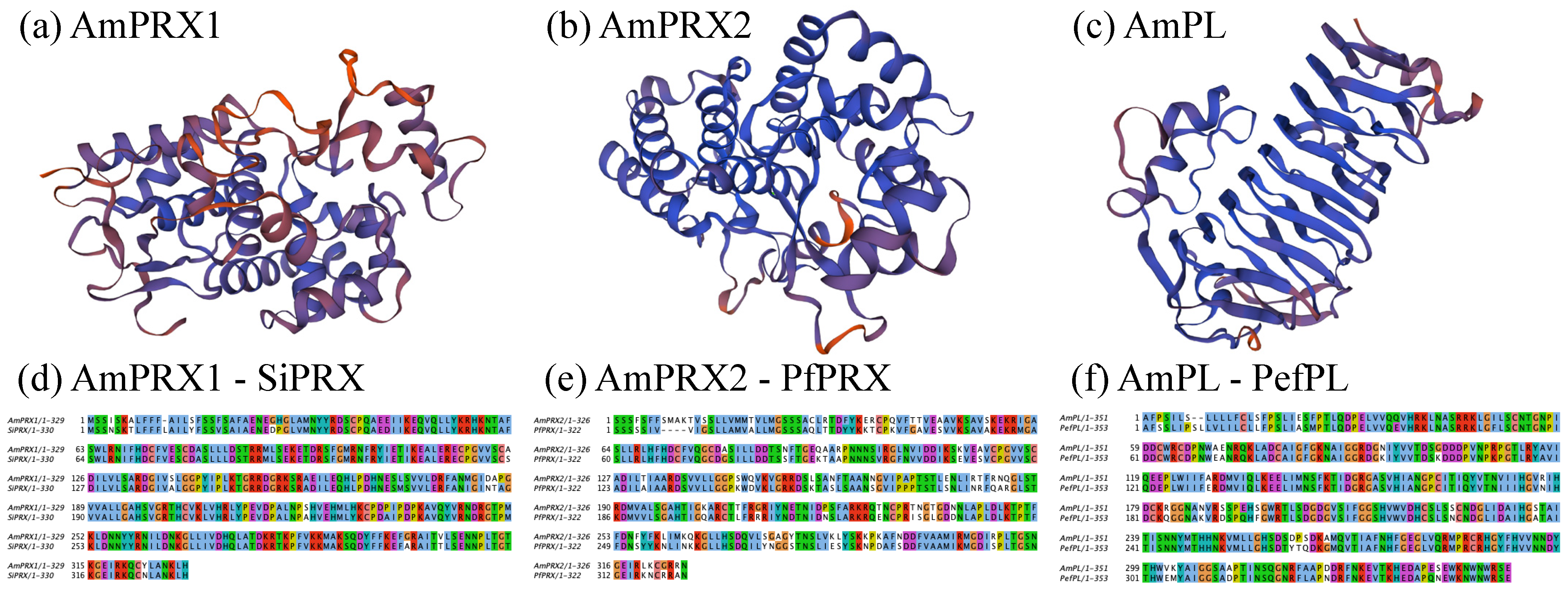
3.1. Structural Characterization of AmPRX1, AmPRX2, AmPL, and AmPSK cDNA Clones
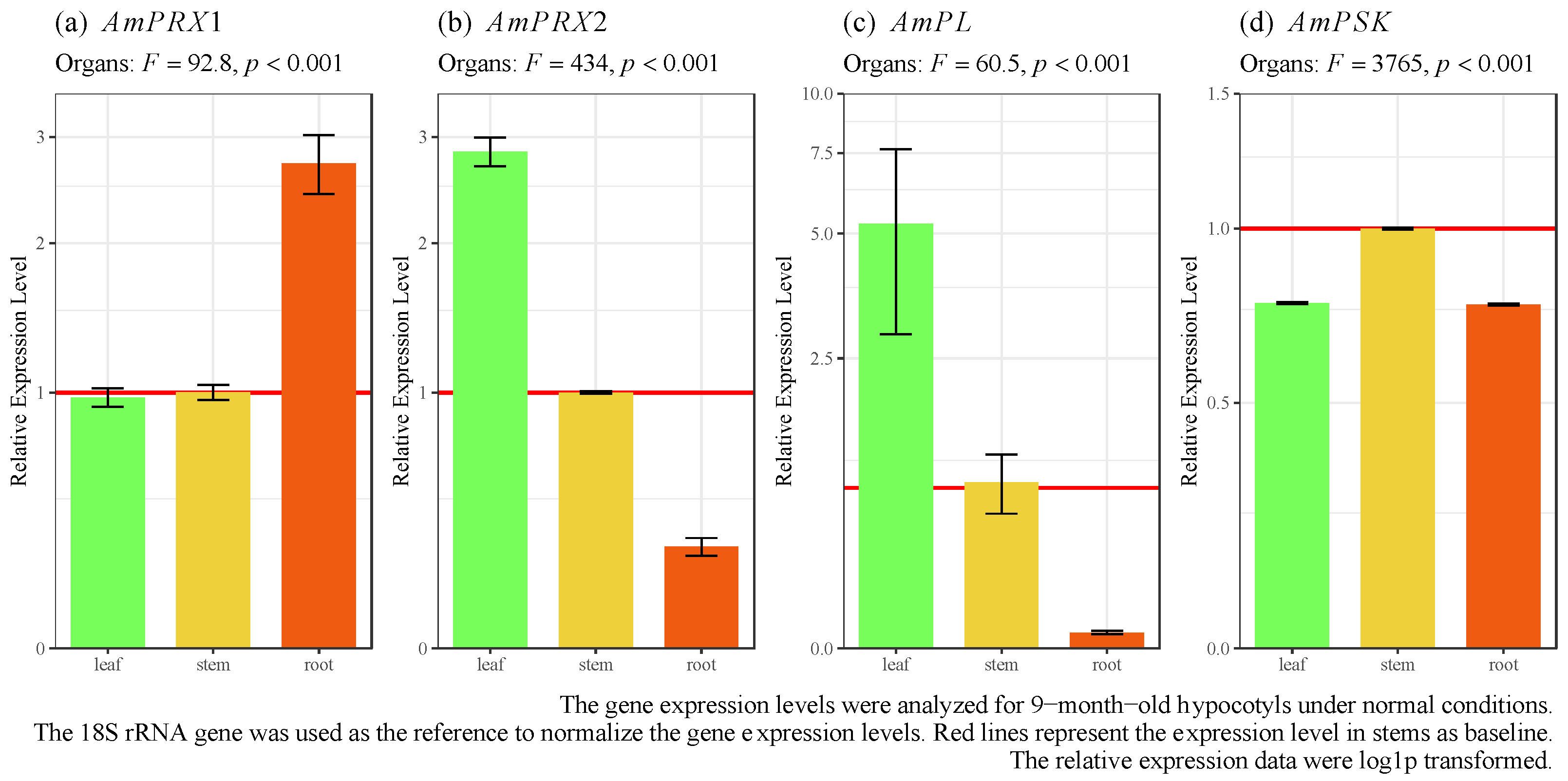
3.2. Spatial Expression of AmPRX1, AmPRX2, AmPL, and AmPSK Genes in Avicennia marina
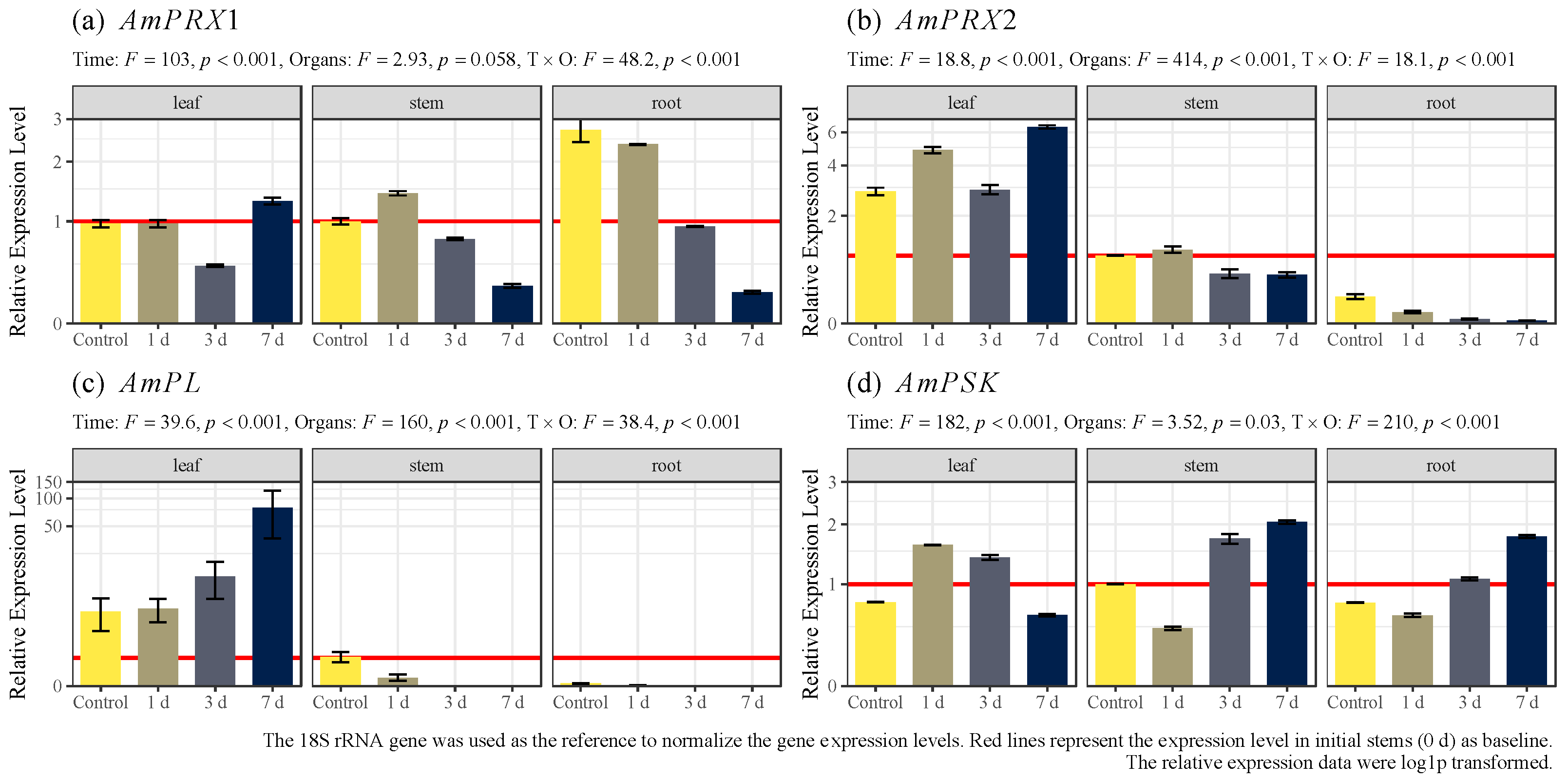
3.3. Expression Levels of AmPRX1, AmPRX2, AmPL, and AmPSK Genes in Avicennia marina under Cadmium Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tomlinson, P.B. The Botany of Mangroves; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar]
- Ghyoumi, R.; Ebrahimi, E.; Mousavi, S.M. Dynamics of Mangrove Forest Distribution Changes in Iran. J. Water Clim. Chang. 2022, 13, 2479–2489. [Google Scholar] [CrossRef]
- Duke, N.C.; Meynecke, J.-O.; Dittmann, S.; Ellison, A.M.; Anger, K.; Berger, U.; Cannicci, S.; Diele, K.; Ewel, K.C.; Field, C.D.; et al. A World without Mangroves? Science 2007, 317, 41–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nath, B.; Birch, G.; Chaudhuri, P. Trace Metal Biogeochemistry in Mangrove Ecosystems: A Comparative Assessment of Acidified (by Acid Sulfate Soils) and Non-Acidified Sites. Sci. Total Environ. 2013, 463–464, 667–674. [Google Scholar] [CrossRef]
- MacFarlane, G.; Burchett, M. Photosynthetic Pigments and Peroxidase Activity as Indicators of Heavy Metal Stress in the Grey Mangrove, Avicennia marina (Forsk.) Vierh. Mar. Pollut. Bull. 2001, 42, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.-Y.; Wang, Y.-S. Expression and Characterization Analysis of Type 2 Metallothionein from Grey Mangrove Species (Avicennia marina) in Response to Metal Stress. Aquat. Toxicol. 2010, 99, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Mendoza, D.; Moreno, A.Q.; Zapata-Perez, O. Coordinated Responses of Phytochelatin Synthase and Metallothionein Genes in Black Mangrove, Avicennia germinans, Exposed to Cadmium and Copper. Aquat. Toxicol. 2007, 83, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Zhang, Y.; Liu, B.; Liu, J.; Ye, J.; Yan, C. Rhizodegradation Gradients of Phenanthrene and Pyrene in Sediment of Mangrove (Kandelia candel (L.) Druce). J. Hazard. Mater. 2011, 196, 263–269. [Google Scholar] [CrossRef]
- Billah, M.M.; Kokushi, E.; Uno, S. Distribution, Geochemical Speciation, and Bioavailable Potencies of Cadmium, Copper, Lead, and Zinc in Sediments from Urban Coastal Environment in Osaka Bay, Japan. Water Air Soil Pollut. 2019, 230, 157. [Google Scholar] [CrossRef]
- Lu, H.; Yan, C.; Liu, J. Low-Molecular-Weight Organic Acids Exuded by Mangrove (Kandelia candel (L.) Druce) Roots and Their Effect on Cadmium Species Change in the Rhizosphere. Environ. Exp. Bot. 2007, 61, 159–166. [Google Scholar] [CrossRef]
- Sarath, N.G.; Puthur, J.T. Heavy Metal Pollution Assessment in a Mangrove Ecosystem Scheduled as a Community Reserve. Wetl. Ecol. Manag. 2021, 29, 719–730. [Google Scholar] [CrossRef]
- Huang, G.-Y.; Wang, Y.-S. Physiological and Biochemical Responses in the Leaves of Two Mangrove Plant Seedlings (Kandelia candel and Bruguiera gymnorrhiza) Exposed to Multiple Heavy Metals. J. Hazard. Mater. 2010, 182, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Tu, Y.; Wang, H.; Wang, Z.; Li, Y.; Chai, L.; Zhang, W.; Lin, Z. Environmental Behavior, Human Health Effect and Pollution Control of Heavy Metal(Loid)s Towards Full Life Cycle Processes. Eco-Environ. Health 2022, 1, 229–243. [Google Scholar] [CrossRef]
- Li, J.; Liu, J.; Lu, H.; Jia, H.; Yu, J.; Hong, H.; Yan, C. Influence of the Phenols on the Biogeochemical Behavior of Cadmium in the Mangrove Sediment. Chemosphere 2016, 144, 2206–2213. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Wu, S.; Wang, Q.; Qian, L.; Lu, H.; Liu, J.; Lin, H.-J.; Zhang, J.; Xu, W.-B.; Yan, C. Trace Metal Pollution Risk Assessment in Urban Mangrove Patches: Potential Linkage with the Spectral Characteristics of Chromophoric Dissolved Organic Matter. Environ. Pollut. 2021, 272, 115996. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.; Lu, H.; Liu, J.; Yu, J.; Wang, Q.; Li, J.; Yang, J.; Hong, H.; Yan, C. SODs Involved in the Hormone Mediated Regulation of H2O2 Content in Kandelia obovata Root Tissues under Cadmium Stress. Environ. Pollut. 2020, 256, 113272. [Google Scholar] [CrossRef]
- Imchen, M.; Kumavath, R.; Barh, D.; Vaz, A.; Góes-Neto, A.; Tiwari, S.; Ghosh, P.; Wattam, A.R.; Azevedo, V. Comparative Mangrove Metagenome Reveals Global Prevalence of Heavy Metals and Antibiotic Resistome across Different Ecosystems. Sci. Rep. 2018, 8, 11187. [Google Scholar] [CrossRef] [Green Version]
- Duke, N. A Systematic Revision of the Mangrove Genus Avicennia (Avicenniaceae) in Australasia*. Aust. Syst. Bot. 1991, 4, 299. [Google Scholar] [CrossRef]
- MacFarlane, G. Leaf Biochemical Parameters in Avicennia marina (Forsk.) Vierh as Potential Biomarkers of Heavy Metal Stress in Estuarine Ecosystems. Mar. Pollut. Bull. 2002, 44, 244–256. [Google Scholar] [CrossRef]
- Kathiresan, K.; Bingham, B.L. Biology of Mangroves and Mangrove Ecosystems. Adv. Mar. Biol. 2001, 40, 81–251. [Google Scholar] [CrossRef]
- MacFarlane, G.; Burchett, M. Cellular Distribution of Copper, Lead and Zinc in the Grey Mangrove, Avicennia marina (Forsk.) Vierh. Aquat. Bot. 2000, 68, 45–59. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C.; Schat, H. Molecular Mechanisms of Metal Hyperaccumulation in Plants. New Phytol. 2009, 181, 759–776. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-Y.; Bovet, L.; Maeshima, M.; Martinoia, E.; Lee, Y. The ABC Transporter AtPDR8 Is a Cadmium Extrusion Pump Conferring Heavy Metal Resistance. Plant J. 2007, 50, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S. Molecular Mechanisms of Plant Metal Tolerance and Homeostasis. Planta 2001, 212, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Song, W.-Y.; Martinoia, E.; Lee, J.; Kim, D.; Kim, D.-Y.; Vogt, E.; Shim, D.; Choi, K.S.; Hwang, I.; Lee, Y. A Novel Family of Cys-Rich Membrane Proteins Mediates Cadmium Resistance in Arabidopsis. Plant Physiol. 2004, 135, 1027–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Goldsbrough, P.B. Functional Homologs of Fungal Metallothionein Genes from Arabidopsis. Plant Cell 1994, 6, 875–884. [Google Scholar] [CrossRef]
- Cobbett, C.S. Phytochelatin Biosynthesis and Function in Heavy-Metal Detoxification. Curr. Opin. Plant Biol. 2000, 3, 211–216. [Google Scholar] [CrossRef]
- Passardi, F.; Penel, C.; Dunand, C. Performing the Paradoxical: How Plant Peroxidases Modify the Cell Wall. Trends Plant Sci. 2004, 9, 534–540. [Google Scholar] [CrossRef]
- Bonifacio, A.; Martins, M.O.; Ribeiro, C.W.; Fontenele, A.V.; Carvalho, F.E.L.; Margis-Pinheiro, M.; Silveira, J.A.G. Role of Peroxidases in the Compensation of Cytosolic Ascorbate Peroxidase Knockdown in Rice Plants under Abiotic Stress. Plant Cell Environ. 2011, 34, 1705–1722. [Google Scholar] [CrossRef]
- Anjum, N.A.; Ahmad, I.; Mohmood, I.; Pacheco, M.; Duarte, A.C.; Pereira, E.; Umar, S.; Ahmad, A.; Khan, N.A.; Iqbal, M.; et al. Modulation of Glutathione and Its Related Enzymes in Plants’ Responses to Toxic Metals and Metalloids—A Review. Environ. Exp. Bot. 2011, 75, 307–324. [Google Scholar] [CrossRef]
- Hu, L.; Liang, W.; Yin, C.; Cui, X.; Zong, J.; Wang, X.; Hu, J.; Zhang, D. Rice MADS3 Regulates ROS Homeostasis during Late Anther Development. Plant Cell 2011, 23, 515–533. [Google Scholar] [CrossRef]
- Dixit, P.; Mukherjee, P.K.; Ramachandran, V.; Eapen, S. Glutathione Transferase from Trichoderma Virens Enhances Cadmium Tolerance without Enhancing Its Accumulation in Transgenic Nicotiana Tabacum. PLoS ONE 2011, 6, e16360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Jiang, Y. Identification of Differentially Expressed Genes under Drought Stress in Perennial Ryegrass. Physiol. Plant. 2010, 139, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Collins, F.S.; Patrinos, A.; Jordan, E.; Chakravarti, A.; Gesteland, R.; Walters, L. New Goals for the U.S. Human Genome Project: 1998–2003. Science 1998, 282, 682–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Yu, J.; Hong, H.; Liu, J.; Lu, H.; Yan, C. Identification of Heavy Metal Pollutant Tolerance-Associated Genes in Avicennia marina (Forsk.) by Suppression Subtractive Hybridization. Mar. Pollut. Bull. 2017, 119, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lu, H.; Liu, J.; Hong, H.; Yan, C. The Influence of Flavonoid Amendment on the Absorption of Cadmium in Avicennia marina Roots. Ecotoxicol. Environ. Saf. 2015, 120, 1–6. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil; California Agricultural Experiment Station: Oakland, CA, USA, 1950; Available online: https://www.researchgate.net/file.PostFileLoader.html?id=54aefd7ed4c118b6358b45db&assetKey=AS%3A273668901408776%401442259158553 (accessed on 1 December 2022).
- Dai, M.; Lu, H.; Liu, W.; Jia, H.; Hong, H.; Liu, J.; Yan, C. Phosphorus Mediation of Cadmium Stress in Two Mangrove Seedlings Avicennia marina and Kandelia obovata Differing in Cadmium Accumulation. Ecotoxicol. Environ. Saf. 2017, 139, 272–279. [Google Scholar] [CrossRef]
- Yang, G.; Zhou, R.; Tang, T.; Shi, S. Simple and Efficient Isolation of High-Quality Total RNA from Hibiscus Tiliaceus, a Mangrove Associate and Its Relatives. Prep. Biochem. Biotechnol. 2008, 38, 257–264. [Google Scholar] [CrossRef]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-Coffee: A Novel Method for Fast and Accurate Multiple Sequence Alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tanaka, S.; Ikeda, K.; Ono, M.; Miyasaka, H. Isolation of Several Anti-Stress Genes from a Mangrove Plant Avicennia marina. World J. Microbiol. Biotechnol. 2002, 18, 801–804. [Google Scholar] [CrossRef]
- Mehta, P.A.; Sivaprakash, K.; Parani, M.; Venkataraman, G.; Parida, A.K. Generation and Analysis of Expressed Sequence Tags from the Salt-Tolerant Mangrove Species Avicennia marina (Forsk) Vierh. Theor. Appl. Genet. 2005, 110, 416–424. [Google Scholar] [CrossRef] [PubMed]
- de Albuquerque, T.L.; Silva, J.D.S.; de Macedo, A.C.; Gonçalves, L.R.B.; Rocha, M.V.P. Biotechnological Strategies for the Lignin-Based Biorefinery Valorization. In Reference Module in Chemistry, Molecular Sciences and Chemical Engineering; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Hiraga, S.; Sasaki, K.; Ito, H.; Ohashi, Y.; Matsui, H. A Large Family of Class III Plant Peroxidases. Plant Cell Physiol. 2001, 42, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. Oxidative Stress, Antioxidants and Stress Tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Lagrimini, L.M.; Rothstein, S. Tissue Specificity of Tobacco Peroxidase Isozymes and Their Induction by Wounding and Tobacco Mosaic Virus Infection. Plant Physiol. 1987, 84, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.T. We Be Jammin’: An Update on Pectin Biosynthesis, Trafficking and Dynamics. J. Exp. Bot. 2016, 67, 495–502. [Google Scholar] [CrossRef] [Green Version]
- Sénéchal, F.; Wattier, C.; Rustérucci, C.; Pelloux, J. Homogalacturonan-Modifying Enzymes: Structure, Expression, and Roles in Plants. J. Exp. Bot. 2014, 65, 5125–5160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Yeats, T.H.; Uluisik, S.; Rose, J.K.C.; Seymour, G.B. Fruit Softening: Revisiting the Role of Pectin. Trends Plant Sci. 2018, 23, 302–310. [Google Scholar] [CrossRef]
- Konno, H.; Nakashima, S.; Katoh, K. Metal-Tolerant Moss Scopelophila Cataractae Accumulates Copper in the Cell Wall Pectin of the Protonema. J. Plant Physiol. 2010, 167, 358–364. [Google Scholar] [CrossRef]
- Carpita, N.C.; Gibeaut, D.M. Structural Models of Primary Cell Walls in Flowering Plants: Consistency of Molecular Structure with the Physical Properties of the Walls during Growth. Plant J. 1993, 3, 1–30. [Google Scholar] [CrossRef]
- Holzwart, E.; Huerta, A.I.; Glöckner, N.; Garnelo Gómez, B.; Wanke, F.; Augustin, S.; Askani, J.C.; Schürholz, A.-K.; Harter, K.; Wolf, S. BRI1 Controls Vascular Cell Fate in the Arabidopsis Root through RLP44 and Phytosulfokine Signaling. Proc. Natl. Acad. Sci. USA 2018, 115, 11838–11843. [Google Scholar] [CrossRef]
- Sauter, M. Phytosulfokine Peptide Signalling. J. Exp. Bot. 2015, 66, 5161–5169. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, Y.; Ogawa, M.; Morita, A.; Sakagami, Y. An LRR Receptor Kinase Involved in Perception of a Peptide Plant Hormone, Phytosulfokine. Science 2002, 296, 1470–1472. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Liu, J.; Liu, W.; Lu, H.; Jia, H.; Hong, H.; Yan, C. Phosphorus Effects on Radial Oxygen Loss, Root Porosity and Iron Plaque in Two Mangrove Seedlings under Cadmium Stress. Mar. Pollut. Bull. 2017, 119, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Liu, W.; Hong, H.; Lu, H.; Liu, J.; Jia, H.; Yan, C. Exogenous Phosphorus Enhances Cadmium Tolerance by Affecting Cell Wall Polysaccharides in Two Mangrove Seedlings Avicennia marina (Forsk.) Vierh and Kandelia Obovata (S., L.) Yong Differing in Cadmium Accumulation. Mar. Pollut. Bull. 2018, 126, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Honda, M.; Oki, S.; Kimura, R.; Harada, A.; Maehara, K.; Tanaka, K.; Meno, C.; Ohkawa, Y. High-Depth Spatial Transcriptome Analysis by Photo-Isolation Chemistry. Nat. Commun. 2021, 12, 4416. [Google Scholar] [CrossRef]
- Leng, Z.; Wu, Y.; Li, J.; Nie, Z.; Jia, H.; Yan, C.; Hong, H.; Wang, X.; Du, D. Phenolic Root Exudates Enhance Avicennia marina Tolerance to Cadmium under the Mediation of Functional Bacteria in Mangrove Sediments. Mar. Pollut. Bull. 2022, 185, 114227. [Google Scholar] [CrossRef]
- Pan, C.; Lu, H.; Yang, C.; Wang, L.; Chen, J.; Yan, C. Comparative Transcriptome Analysis Reveals Different Functions of Kandelia obovata Superoxide Dismutases in Regulation of Cadmium Translocation. Sci. Total Environ. 2021, 771, 144922. [Google Scholar] [CrossRef]
- Garcia, J.S.; Dalmolin, Â.C.; Cortez, P.A.; Barbeira, P.S.; Mangabeira, P.A.O.; França, M.G.C. Short-Term Cadmium Exposure Induces Gas Exchanges, Morphological and Ultrastructural Disturbances in Mangrove Avicennia schaueriana Young Plants. Mar. Pollut. Bull. 2018, 131, 122–129. [Google Scholar] [CrossRef]
- Zhang, Q.; Yan, C.; Liu, J.; Lu, H.; Duan, H.; Du, J.; Wang, W. Silicon Alleviation of Cadmium Toxicity in Mangrove (Avicennia marina) in Relation to Cadmium Compartmentation. J. Plant Growth Regul. 2014, 33, 233–242. [Google Scholar] [CrossRef]
- Pan, C.; Lu, H.; Yu, J.; Liu, J.; Liu, Y.; Yan, C. Identification of Cadmium-Responsive Kandelia obovata SOD Family Genes and Response to Cd Toxicity. Environ. Exp. Bot. 2019, 162, 230–238. [Google Scholar] [CrossRef]
- Kumar, S.; Asif, M.H.; Chakrabarty, D.; Tripathi, R.D.; Dubey, R.S.; Trivedi, P.K. Expression of a Rice Lambda Class of Glutathione S-Transferase, OsGSTL2, in Arabidopsis Provides Tolerance to Heavy Metal and Other Abiotic Stresses. J. Hazard. Mater. 2013, 248–249, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Sun, J.; Du, L.; Liu, X. Comparative Transcriptome Analysis of Cadmium Responses in Solanum nigrum and Solanum torvum. New Phytol. 2012, 196, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, C.; Du, J.; Liu, H.; Cui, Y.; Zhang, Y.; He, Y.; Wang, Y.; Chu, C.; Feng, Z.; et al. Co-Overexpression FIT with AtbHLH38 or AtbHLH39 in Arabidopsis-Enhanced Cadmium Tolerance via Increased Cadmium Sequestration in Roots and Improved Iron Homeostasis of Shoots. Plant Physiol. 2012, 158, 790–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, J.; Matamoros, M.A.; Naya, L.; James, E.K.; Rouhier, N.; Sato, S.; Tabata, S.; Becana, M. The Glutathione Peroxidase Gene Family of Lotus japonicus: Characterization of Genomic Clones, Expression Analyses and Immunolocalization in Legumes. New Phytol. 2009, 181, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Dunajska-Ordak, K.; Skorupa-Kłaput, M.; Kurnik, K.; Tretyn, A.; Tyburski, J. Cloning and Expression Analysis of a Gene Encoding for Ascorbate Peroxidase and Responsive to Salt Stress in Beet (Beta vulgaris). Plant Mol. Biol. Rep. 2014, 32, 162–175. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, J.C.; Yan, C.L.; Du, D.L.; Lu, H.L. The Alleviation Effect of Iron on Cadmium Phytotoxicity in Mangrove A. marina. Alleviation Effect of Iron on Cadmium Phytotoxicity in Mangrove Avicennia marina (Forsk.) Vierh. Chemosphere 2019, 226, 413–420. [Google Scholar] [CrossRef]
- Zhu, C.-Q.; Ghoto, K.; Gao, G.-F.; Chen, J.; Hu, W.-J.; Qiao, F.; Liu, J.-Y.; Zheng, H.-L. Trace Metals Complexation Behavior with Root Exudates Induced by Salinity from a Mangrove Plant Avicennia marina (Forsk.) Vierh. Bioremediat. J. 2019, 23, 82–93. [Google Scholar] [CrossRef]
- Yan, Z.; Li, X.; Chen, J.; Tam, N.F.-Y. Combined Toxicity of Cadmium and Copper in Avicennia marina Seedlings and the Regulation of Exogenous Jasmonic Acid. Ecotoxicol. Environ. Saf. 2015, 113, 124–132. [Google Scholar] [CrossRef]
- Wang, L.-Y.; Wang, Y.-S.; Zhang, J.-P.; Gu, J.-D. Molecular Cloning of Class III Chitinase Gene from Avicennia marina and Its Expression Analysis in Response to Cadmium and Lead Stress. Ecotoxicology 2015, 24, 1697–1704. [Google Scholar] [CrossRef]
- Zhang, Q.; Yan, C.; Liu, J.; Lu, H.; Wang, W.; Du, J.; Duan, H. Silicon Alleviates Cadmium Toxicity in Avicennia marina (Forsk.) Vierh. Seedlings in Relation to Root Anatomy and Radial Oxygen Loss. Mar. Pollut. Bull. 2013, 76, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.-L.; Wang, Y.-S.; Cheng, H.; Sun, C.-C.; Wu, P.; Wang, L.-Y.; Fei, J. Characterization and Expression Analysis of Three CBF/DREB1 Transcriptional Factor Genes from Mangrove Avicennia marina. Aquat. Toxicol. 2013, 140–141, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Raju, K.A.; Ramakrishna, C. The Effects of Heavy Metals on the Anatomical Structures of Avicennia marina (Forssk.) Vierh. Braz. J. Bot. 2021, 44, 439–447. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Full Length | 5′ UTR | 3′ UTR | ORF | Predicted MW | Theoretical pI | Cloned by | Accession Number |
|---|---|---|---|---|---|---|---|---|
| (bp) | (bp) | (bp) | (bp) | (kDa) | ||||
| AmPRX1 | 1345 | 48 | 301 | 996 | 37.60 | 8.41 | [42] | AB049589.1 |
| AmPRX2 | 1404 | 70 | 341 | 993 | 36.05 | 9.58 | This study | OQ160797 |
| AmPL | 1545 | 393 | 1152 | 42.85 | 6.92 | This study | OQ160798 | |
| AmPSK | 727 | 162 | 331 | 234 | 55.43 | 5.06 | [43] | AY639950.1 |
| Interface | Key Adaptive Feature | Mechanism | Reference |
|---|---|---|---|
| sediment-root | rhizosphere microbiome | phytoavailability | [59] |
| root | anatomical structures | stress-regulated genes; metal translocation | [60] |
| root | gene expression; phytohormone | stress-regulated genes | [16] |
| root-leaf | gene expression; enzyme activity | stress-regulated genes | [63] |
| sediment-root | iron plaque | metal segregation | [69] |
| sediment-root | root exudates | phytoavailability | [70] |
| root | enriched substances | cellular structure | [57] |
| root-leaf | anatomical structures | metal translocation | [61] |
| leaf | enriched substances | detoxification | [38] |
| sediment-root | metal speciation | phytoavailability | [14] |
| sediment-root | root exudates | phytoavailability | [36] |
| leaf | gene expression; enzyme activity | stress-regulated genes; detoxification | [71] |
| leaf | gene expression | stress-regulated genes | [72] |
| sediment-root | anatomical structures | phytoavailability | [73] |
| root-leaf | subcellular distribution | metal translocation | [62] |
| leaf | gene expression | stress-regulated genes | [74] |
| leaf | gene expression | stress-regulated genes | [6] |
| stem-leaf | anatomical structures | metal translocation | [75] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Zhang, J.; Hong, H. Characterization and Expression Analysis of Four Cadmium-Tolerance-Associated Genes of Avicennia marina (Forsk.). Biology 2023, 12, 216. https://doi.org/10.3390/biology12020216
Yu J, Zhang J, Hong H. Characterization and Expression Analysis of Four Cadmium-Tolerance-Associated Genes of Avicennia marina (Forsk.). Biology. 2023; 12(2):216. https://doi.org/10.3390/biology12020216
Chicago/Turabian StyleYu, Jinfeng, Jicheng Zhang, and Hualong Hong. 2023. "Characterization and Expression Analysis of Four Cadmium-Tolerance-Associated Genes of Avicennia marina (Forsk.)" Biology 12, no. 2: 216. https://doi.org/10.3390/biology12020216