

Assessment of Drought and Zinc Stress Tolerance of Novel Miscanthus Hybrids and Arundo donax Clones Using Physiological, Biochemical, and Morphological Traits

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material, Growing Conditions and Experimental Design
2.2. Growth and Biomass Characterization
2.3. Chlorophyll Fluorescence Measurements
2.4. Determination of Relative Water Content (RWC)%
2.5. Biochemical Assays
2.5.1. Analysis of Proline, Phenol and Malondialdehyde (MDA)
2.5.2. Determination of Total Soluble Protein and Antioxidant Enzymes Activity
2.6. Drought and Zn Tolerance Evaluation
2.7. Statistical Analysis and Clustering
3. Results
3.1. Growth Attributes and Biomass Accumulation
3.2. Physiological Responses to Drought and Zn Stress
3.2.1. Chlorophyll Fluorescence under Drought and Zn Stress
3.2.2. Effect of Drought and Zn Stress on Water Content (RWC)
3.3. Biochemical Responses to Drought and Zn Stress
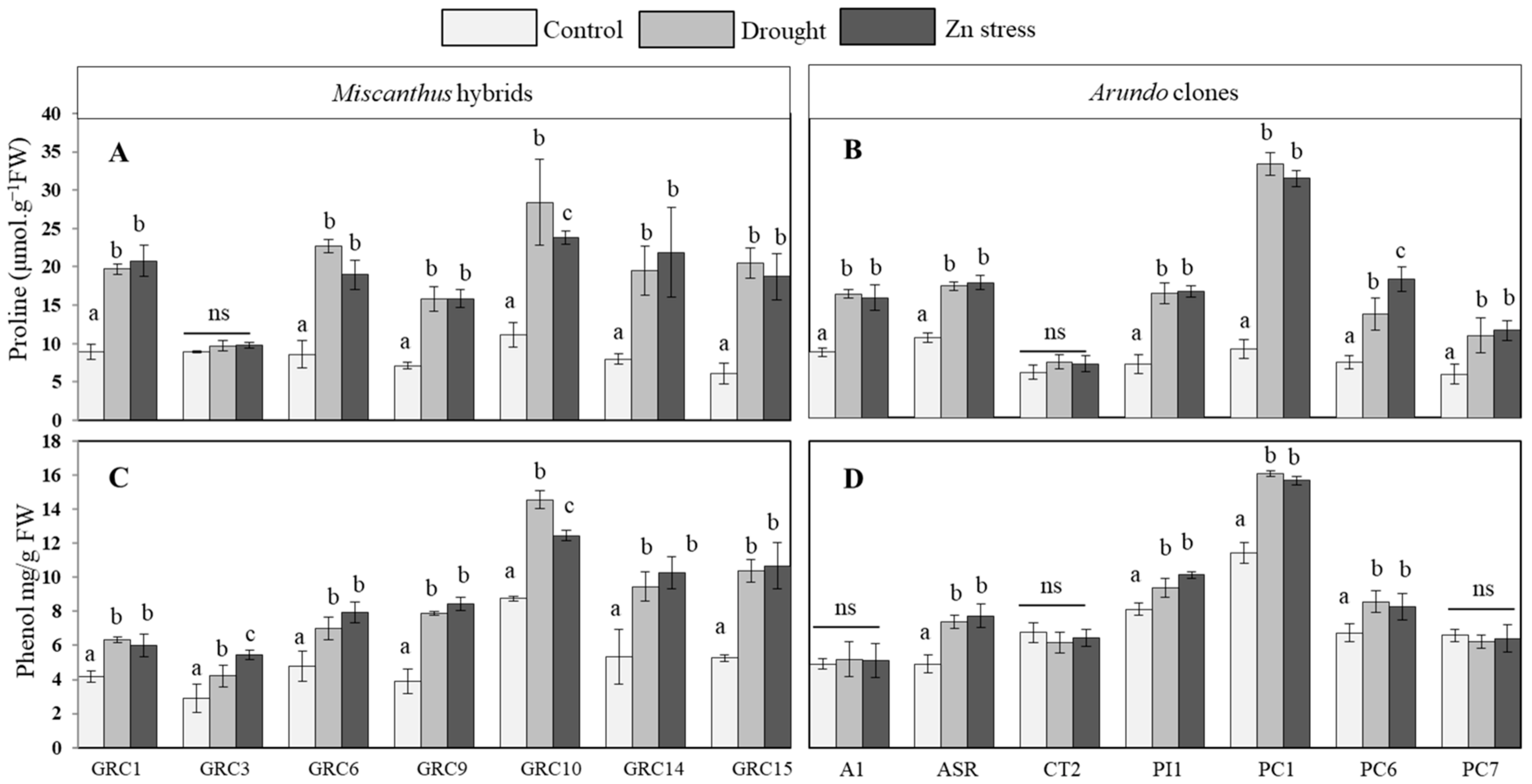
3.3.1. Effect of Drought and Zn Stress on Proline and Phenol Accumulation
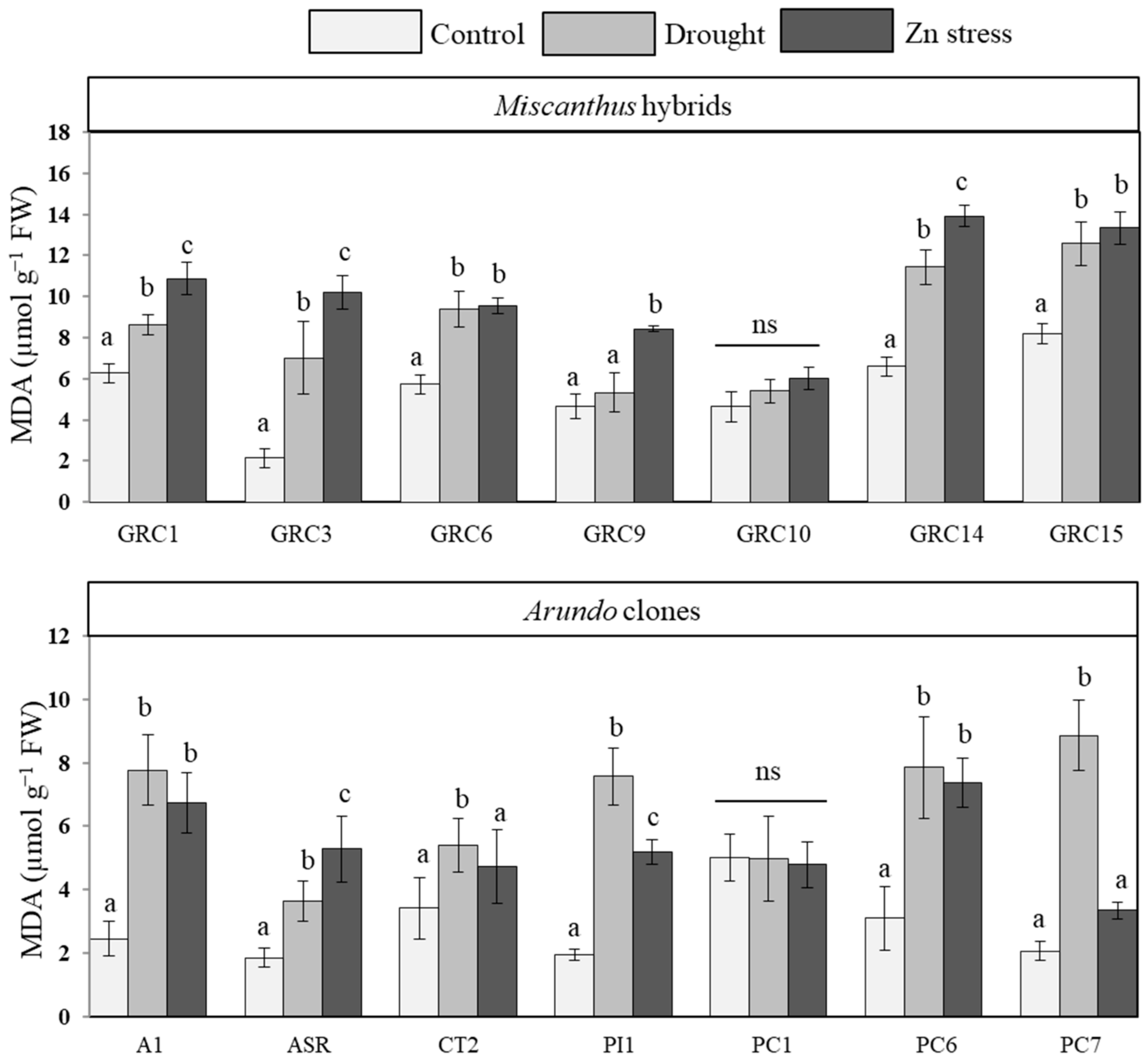
3.3.2. Lipid Peroxidation under Drought and Zn Stress
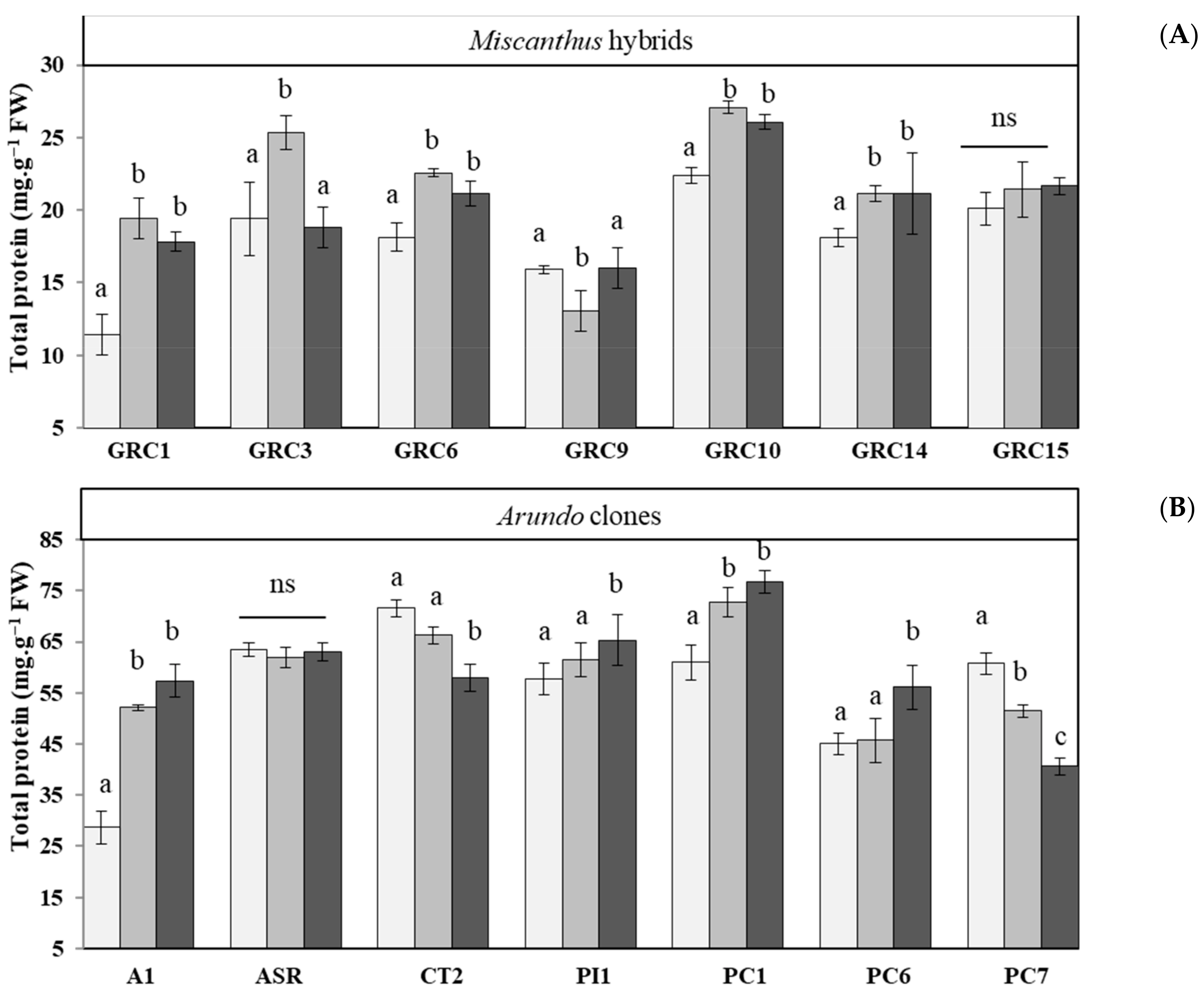
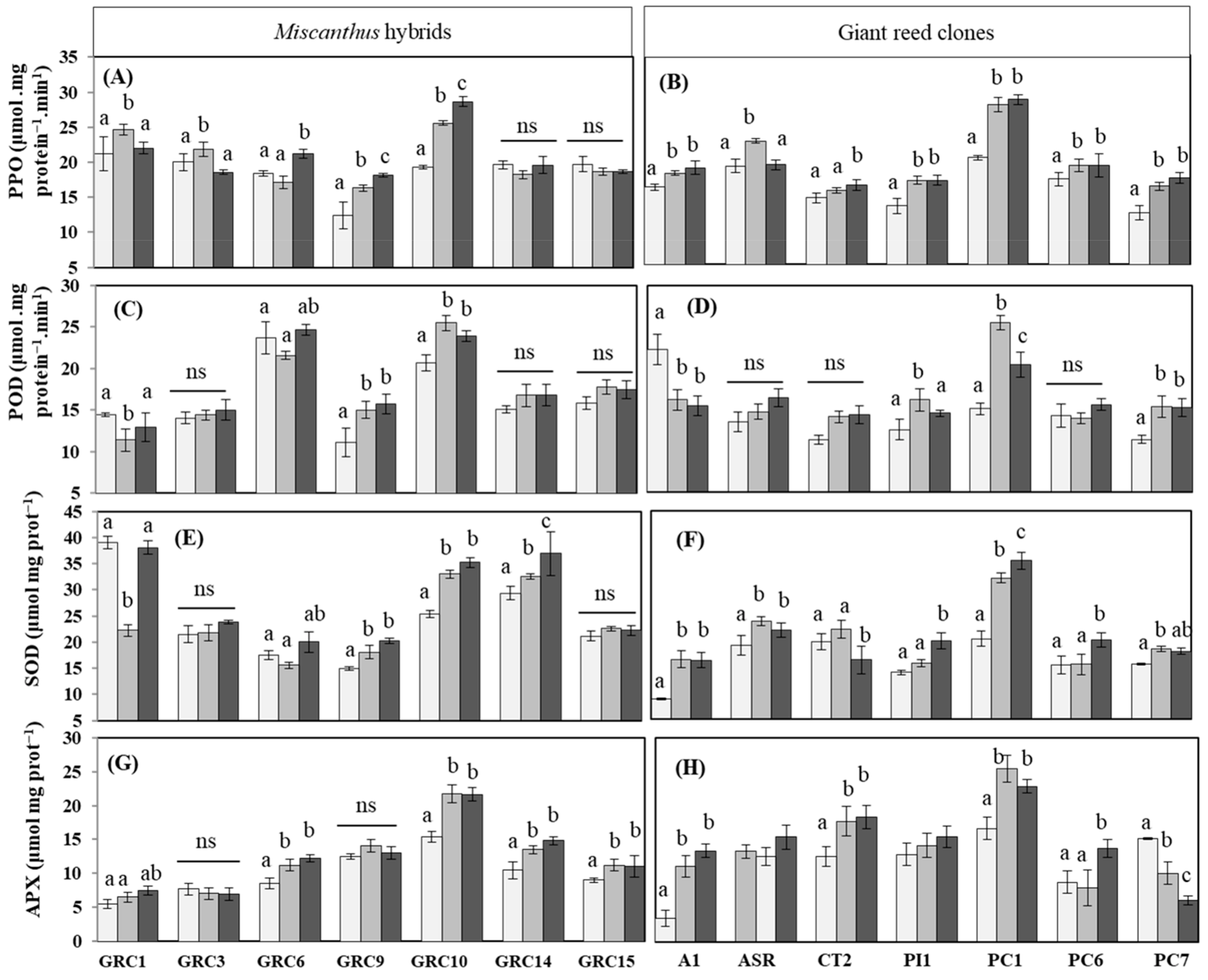
3.3.3. Effect of Drought and Zn Stress on Soluble Protein Accumulation and Activities of Antioxidant Enzymes
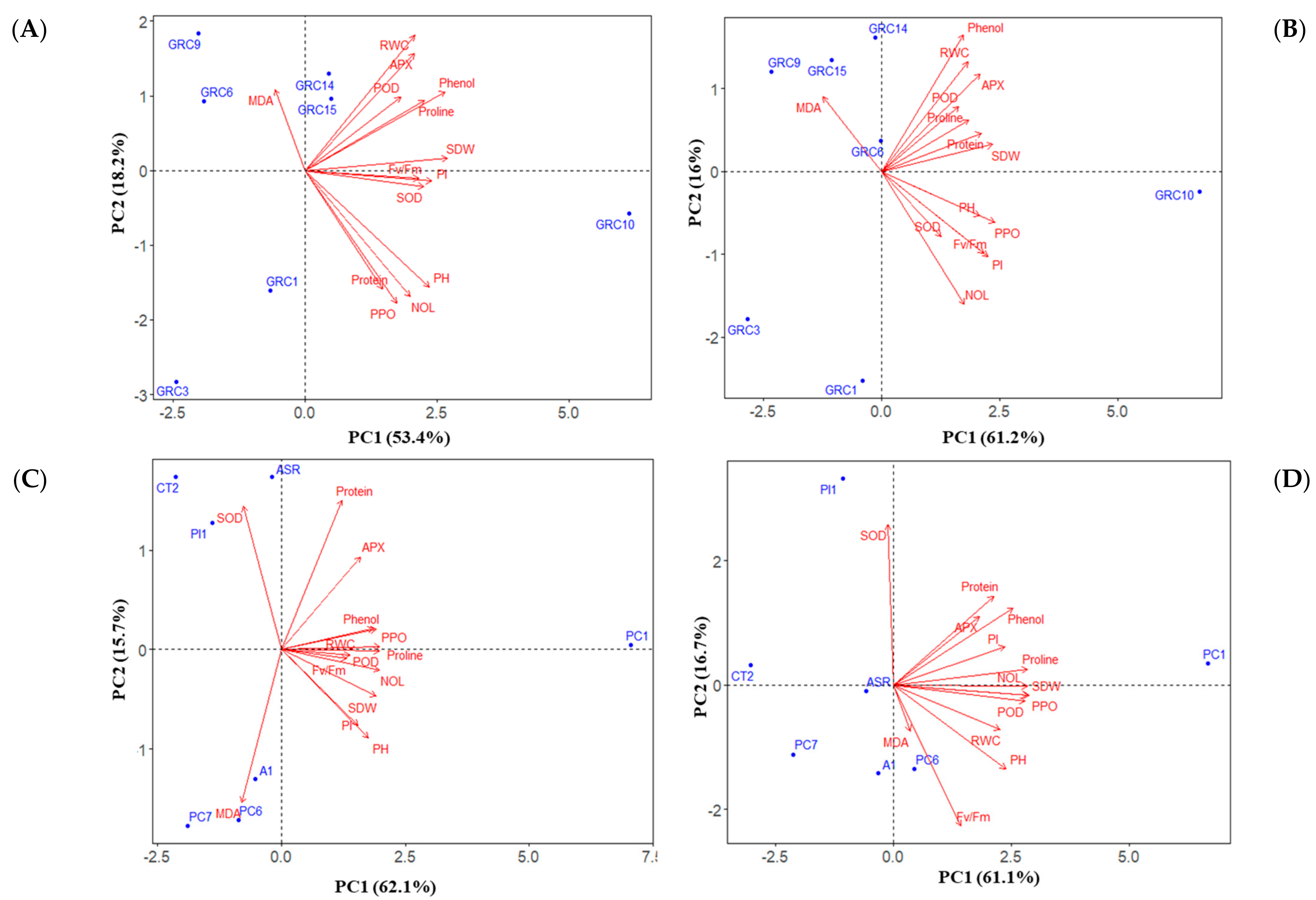
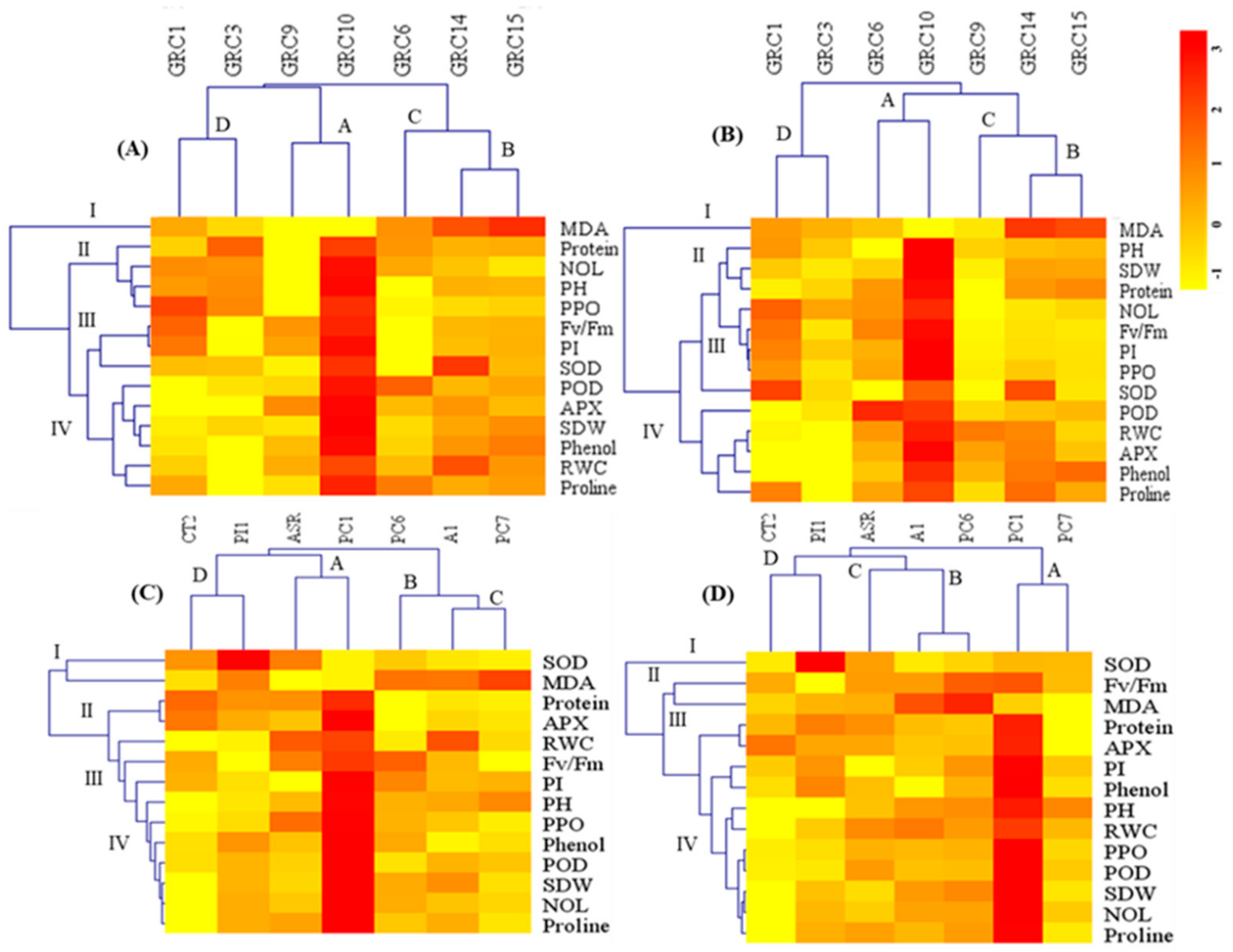
3.4. Ranking among the Hybrids and Clones to Drought and Zn Stress Tolerance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- COM. Annex to the Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee, the Committee of the Regions and the European Investment Bank; Third Report on the State of Energy Union; European Commission: Brussels, Belgium, 2017; p. 688. Available online: https://www.cde.unict.it/sites/cde.unict.it/files/1_2012_doc.pdf (accessed on 21 November 2023).
- Lewandowski, I.; Scurlock, J.M.O.; Lindvall, E.; Christou, M. The development and current status of perennial rhizomatous grasses as energy crops in the US and Europe. Biomass Bioenergy 2003, 25, 335–361. [Google Scholar] [CrossRef]
- Manzone, M.; Bergante, S.; Facciotto, G. Energy and economic evaluation of a poplar plantation for woodchips production in Italy. Biomass Bioenergy 2014, 60, 164–170. [Google Scholar] [CrossRef]
- Amaducci, S.; Perego, A. Field evaluation of Arundo donax clones for bioenergy production. Ind. Crops. Prod. 2015, 75, 122–128. [Google Scholar] [CrossRef]
- Amaducci, S.; Facciotto, G.; Bergante, S.; Perego, A.; Serra, P.; Ferrarini, A.; Chimento, C. Biomass production and energy balance of herbaceous and woody crops on marginal soils in the Po Valley. GCB Bioenergy 2017, 9, 31–45. [Google Scholar] [CrossRef]
- Larsen, S.U.; Jørgensen, U.; Lærke, P.E. Biomass yield, nutrient concentration and nutrient uptake by SRC willow cultivars grown on different sites in Denmark. Biomass Bioenergy 2018, 116, 161–170. [Google Scholar] [CrossRef]
- Jeguirim, M.; Trouvé, G. Pyrolysis characteristics and kinetics of Arundo donax using thermogravimetric analysis. Bioresour. Technol. 2009, 100, 4026–4031. [Google Scholar] [CrossRef] [PubMed]
- Pilu, R.; Badone, F.C.; Michela, L. Giant reed (Arundo donax L.): A weed plant or a promising energy crop? Afr. J. Biotechnol. 2012, 11, 9163–9174. [Google Scholar]
- Di Girolamo, G.; Grigatti, M.; Barbanti, L.; Angelidaki, I. Effects of hydrothermal pre-treatments on Giant reed (Arundo donax) methane yield. Bioresour. Technol. 2013, 147, 152–159. [Google Scholar] [CrossRef]
- Ragaglini, G.; Dragoni, F.; Simone, M.; Bonari, E. Suitability of giant reed (Arundo donax L.) for anaerobic digestion: Effect of harvest time and frequency on the biomethane yield potential. Bioresour. Technol. 2014, 152, 107–115. [Google Scholar] [CrossRef]
- Cattivelli, L.; Rizza, F.; Badeck, F.; Mazzucotelli, E.; Mastrangelo, A.M.; Francia, E.; Marè, C.; Tondelli, A.; Stanca, A.M. Drought tolerance improvement in crop plants: An integrated view from breeding to genomics. Field Crop. Res. 2008, 105, 1–14. [Google Scholar] [CrossRef]
- Saxena, P.; Misra, N. Remediation of heavy metal contaminated tropical land. In Soil Heavy Metals; Springer: Berlin, Heidelberg, 2010; pp. 431–477. [Google Scholar]
- Maughan, M.; Bollero, G.; Lee, D.K.; Darmody, R.; Bonos, S.; Cortese, L.; Murphy, J.; Gaussoin, R.; Sousek, M.; Williams, D.; et al. Miscanthus × giganteus productivity: The effects of management in different environments. GCB Bioenergy 2012, 4, 253–265. [Google Scholar] [CrossRef]
- Clifton-Brown, J.C.; Lewandowski, I. Overwintering problems of newly established Miscanthus plantations can be overcome by identifying genotypes with improved rhizome cold tolerance. New Phytol. 2000, 148, 287–294. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I.; Comas, P. Changed plant and animal life cycles from 1952 to 2000 in the Mediterranean region. Glob. Chang. Biol. 2002, 8, 531–544. [Google Scholar] [CrossRef]
- Sardans, J.; Penuelas, J. Trace element accumulation in the moss Hypnum cupressiforme Hedw. and the trees Quercus ilex L. and Pinus halepensis Mill. in Catalonia. Chemosphere 2005, 60, 1293–1307. [Google Scholar]
- Sensuła, B.; Fagel, N.; Michczyński, A. Radiocarbon, trace elements and pb isotope composition of pine needles from a highly industrialized region in southern Poland. Radiocarbon 2021, 63, 713–726. [Google Scholar] [CrossRef]
- Government Decree on the Assessment of Soil Contamination and Remediation Needs; Ministry of the Environment: Helsinki, Finland, 2021; p. 214. Available online: https://finlex.fi/en/laki/kaannokset/2007/en20070214.pdf (accessed on 10 July 2021).
- Tóth, G.; Hermann, T.; Da Silva, M.R.; Montanarella, L. Heavy metals in agricultural soils of the European Union with implications for food safety. Environ. Int. 2016, 88, 299–309. [Google Scholar] [CrossRef]
- Yap, C.K.; Al-Mutairi, K.A. Ecological-health risk assessments of heavy metals (Cu, Pb, and Zn) in aquatic sediments from the ASEAN-5 emerging developing countries: A review and synthesis. Biology 2021, 11, 7. [Google Scholar] [CrossRef]
- Gojon, A.; Cassan, O.; Bach, L.; Lejay, L.; Martin, A. The decline of plant mineral nutrition under rising CO2: Physiological and molecular aspects of a bad deal. Trends Plant Sci. 2023, 28, 185–198. [Google Scholar] [CrossRef]
- Sagardoy, R.; Morales, F.; López-Millán, A.F.; Abadía, A.; Abadía, J. Effects of zinc toxicity on sugar beet (Beta vulgaris L.) plants grown in hydroponics. Plant Biol. 2009, 11, 339–350. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; Taylor and Francis: Boca Raton, FL, USA, 2011; p. 505. [Google Scholar]
- Andrejić, G.; Gajić, G.; Prica, M.; Dželetović, Ž.; Rakić, T. Zinc accumulation, photosynthetic gas exchange, and chlorophyll a fluorescence in Zn-stressed Miscanthus × giganteus plants. Photosynthetica 2018, 56, 1249–1258. [Google Scholar] [CrossRef]
- Bonnet, M.; Camares, O.; Veisseire, P. Effects of zinc and influence of Acremonium lolii on growth parameters, chlorophyll a fluorescence and antioxidant enzyme activities of ryegrass (Lolium prenne L. cv. Apollo). J. Exp. Bot. 2000, 51, 945–953. [Google Scholar] [PubMed]
- Lei, Y.; Yin, C.; Li, C. Differences in some morphological, physiological, and biochemical responses to drought stress in two contrasting populations of Populus przewalskii. Physiol. Plant. 2006, 127, 182–191.28. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Yang, F.; Zhang, S.; Korpelainen, H.; Li, C. Physiological and proteomic responses of two contrasting Populus cathayana populations to drought stress. Physiol. Plant. 2009, 136, 150–168. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, D.; Zhao, L.; Chen, A.; Li, J.; Tang, H.; Pan, G.; Chang, L.; Deng, Y.; Huang, S. Comparative transcriptome combined with physiological analyses revealed key factors for differential cadmium tolerance in two contrasting hemp (Cannabis sativa L.) cultivars. Ind. Crops. Prod. 2019, 140, 111638. [Google Scholar] [CrossRef]
- Zgallaï, H.; Steppe, K.; Lemeur, R. Effects of different levels of water stress on leaf water potential, stomatal resistance, protein and chlorophyll content and certain antioxidative enzymes in tomato plants. J. Integr. Plant. Biol. 2006, 48, 679–685. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Bajji, M.; Lutts, S.; Kinet, J.M. Water deficit effects on solute contribution to osmotic adjustment as a function of leaf ageing in three durum wheat (Triticum durum Desf.) cultivars performing differently in arid conditions. Plant. Sci. 2001, 160, 669–681. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Y.; Guo, K.; Fan, D.; Li, G.; Zheng, Y.; Yu, L.; Yang, R. Effect of drought on pigments, osmotic adjustment and antioxidant enzymes in six woody plant species in karst habitats of southwestern China. Environ. Exp. Bot. 2011, 71, 174–183. [Google Scholar] [CrossRef]
- Papazoglou, E.G.; Karantounias, G.A.; Vemmos, S.N.; Bouranis, D.L. Photosynthesis and growth responses of giant reed (Arundo donax L.) to the heavy metals Cd and Ni. Environ. Int. 2005, 31, 243–249. [Google Scholar] [CrossRef]
- Ings, J.; Mur, L.A.; Robson, P.R.; Bosch, M. Physiological and growth responses to water deficit in the bioenergy crop Miscanthus × giganteus. Front. Plant Sci. 2013, 4, 468. [Google Scholar] [CrossRef]
- Cristaldi, A.; Conti, G.O.; Cosentino, S.L.; Mauromicale, G.; Copat, C.; Grasso, A.; Ferrante, M. Phytoremediation potential of Arundo donax (Giant Reed) in contaminated soil by heavy metals. Environ. Res. 2020, 185, 109427. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, B.; Boléo, S.; Sidella, S.; Costa, J.; Duarte, M.P.; Mendes, B.; Fernando, A.L. Phytoremediation of heavy metal-contaminated soils using the perennial energy crops Miscanthus spp. and Arundo donax L. Bioenergy Res. 2015, 8, 1500–1511. [Google Scholar] [CrossRef]
- Impollonia, G.; Croci, M.; Martani, E.; Ferrarini, A.; Kam, J.; Trindade, L.M.; Clifton-Brown, J.; Amaducci, S. Moisture content estimation and senescence phenotyping of novel Miscanthus hybrids combining UAV-based remote sensing and machine learning. GCB Bioenergy 2022, 14, 639–656. [Google Scholar] [CrossRef]
- Cavallaro, V.; Scordia, D.; Cosentino, S.L.; Copani, V. Up-scaling agamic propagation of giant reed (Arundo donax L.) by means of single-node stem cuttings. Ind. Crops. Prod. 2019, 128, 534–544. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. Probing Photosynth. Mech. Regul. Adapt. 2000, 25, 445–483. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant. Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Yu, L.; Fan, J.; Xu, C. Peroxisomal fatty acid β-oxidation negatively impacts plant survival under salt stress. Plant Signal. Behav. 2019, 14, 1561121. [Google Scholar] [CrossRef]
- Smith, P.E.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Kar, M.; Mishra, D. Catalase, peroxidase, and polyphenoloxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef]
- Goud, P.B.; Kachole, M.S. Antioxidant enzyme changes in neem, pigeonpea and mulberry leaves in two stages of maturity. Plant Signal. Behavior. 2012, 7, 1258–1262. [Google Scholar] [CrossRef]
- Gepstein, S.H.; Glick, B.R. Strategies to ameliorate abiotic stress-induced plant senescence. Plant Mol. Biol. 2013, 82, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Talebi, R.; Fayaz, F.; Naji, A.M. Effective selection criteria for assessing drought stress tolerance in durum wheat (Triticum durum Desf.). General Appl. Plant Phys. 2009, 35, 64–74. [Google Scholar]
- Zhu, X.; Chang, G.; He, D.; Zhao, H.; Ma, C. Evaluation of new onion varieties using cluster analysis and principal component analysis methods. Gansu Agr. Sci. 2014, 10, 25–28. [Google Scholar]
- Shaheen, S.; Ahmad, R.; Mahmood, Q.; Pervez, A.; Maroof Shah, M.; Hafeez, F. Gene expression and biochemical response of giant reed under Ni and Cu stress. Int. J. Phytoremediat. 2019, 21, 1474–1485.50. [Google Scholar] [CrossRef] [PubMed]
- Styles, D.; Thorne, F.; Jones, M.B. Energy crops in Ireland: An economic comparison of willow and Miscanthus production with conventional farming systems. Biomass Bioenergy 2008, 32, 407–421. [Google Scholar] [CrossRef]
- Clifton-Brown, J.C.; Lewandowski, I.; Bangerth, F.; Jones, M.B. Comparative responses to water stress in stay-green, rapid-and slow senescing genotypes of the biomass crop, Miscanthus. New Phytol. 2002, 154, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Stavridou, E.; Webster, R.J.; Robson, P.R. Novel Miscanthus genotypes selected for different drought tolerance phenotypes show enhanced tolerance across combinations of salinity and drought treatments. Ann. Bot. 2019, 124, 653–674. [Google Scholar] [CrossRef]
- Haworth, M.; Centritto, M.; Giovannelli, A.; Marino, G.; Proietti, N.; Capitani, D.; Loreto, F. Xylem morphology determines the drought response of two Arundo donax ecotypes from contrasting habitats. GCB Bioenergy 2017, 9, 119–131.57. [Google Scholar] [CrossRef]
- Zegada-Lizarazu, W.; Salvi, S.; Monti, A. Assessment of mutagenized giant reed clones for yield, drought resistance and biomass quality. Biomass Bioenergy 2020, 134, 105501. [Google Scholar] [CrossRef]
- Shao, H.; Chu, L.; Jaleel, C.A.; Zhao, C. Water-deficit stress-induced anatomical changes in higher plants. C. R. Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Dambreville, A.; Griolet, M.; Rolland, G.; Dauzat, M.; Bédiée, A.; Balsera, C.; Muller, B.; Vile, D.; Granier, C. Phenotyping oilseed rape growth-related traits and their responses to water deficit: The disturbing pot size effect. Funct. Plant Biol. 2017, 44, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Nogués, S.; Alegre, L.; Araus, J.; Perez-Aranda, L.; Lannoye, R. Modulatedchlorophyll fluorescence and photosynthetic gas exchange as rapid screeningmethods for drought tolerance in barley genotypes. Photosynthetica 1994, 30, 465–474. [Google Scholar]
- Misra, A.N.; Srivastava, A.; Strasser, R.J. Utilization of fast chlorophyll a fluorescence technique in assessing the salt/ion sensitivity of mung bean and Brassica seedlings. J. Plant Physiol. 2001, 158, 1173–1181. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rodríguez, E.; Rubio-Wilhelmi, M.; Cervilla, L.M.; Blasco, B.; Rios, J.J.; Rosales, M.A.; Ruiz, J.M. Genotypic differences in some physiological parameters symptomatic for oxidative stress under moderate drought in tomato plants. Plant Sci. 2010, 178, 30–40. [Google Scholar] [CrossRef]
- Siddique, M.R.B.; Hamid, A.I.M.S.; Islam, M.S. Drought stress effects on water relations of wheat. Bot. Bull. Acad. Sin. 2000, 41, 35–39. [Google Scholar]
- Islam, M.; Begum, M.C.; Kabir, A.H.; Alam, M.F. Molecular and biochemical mechanisms associated with differential responses to drought tolerance in wheat (Triticum aestivum L.). J. Plant Interact. 2015, 10, 195–201. [Google Scholar] [CrossRef]
- Aghaie, P.; Tafreshi, S.A.H.; Ebrahimi, M.A.; Haerinasab, M. Tolerance evaluation and clustering of fourteen tomato cultivars grown under mild and severe drought conditions. Sci. Hortic. 2018, 232, 1–12. [Google Scholar] [CrossRef]
- Fracasso, A.; Trindade, L.; Amaducci, S. Drought tolerance strategies highlighted by two Sorghum bicolor races in a dry-down experiment. J. Plant Physiol. 2016, 190, 1–14. [Google Scholar] [CrossRef]
- Janas, K.M.; Cvikrová, M.; Pałagiewicz, A.; Szafranska, K.; Posmyk, M.M. Constitutive elevated accumulation of phenylpropanoids in soybean roots at low temperature. Plant Sci. 2002, 163, 369–373. [Google Scholar] [CrossRef]
- Pandey, P.; Tripathi, A.; Dwivedi, S.; Lal, K.; Jhang, T. Deciphering the mechanisms, hormonal signaling, and potential applications of endophytic microbes to mediate stress tolerance in medicinal plants. Front. Plant Sci. 2023, 14, 1250020. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, V.; Alooparampil, S.; Pandya, R.V.; Tank, J.G. Physiological Function of Phenolic Compounds in Plant Defense System. In Phenolic Compounds-Chemistry, Synthesis, Diversity, Non-Conventional Industrial. Pharm. Thera App; IntechOpen: London, UK, 2021. [Google Scholar]
- De Vega, J.J.; Teshome, A.; Klaas, M.; Grant, J.; Finnan, J.; Barth, S. Physiological and transcriptional response to drought stress among bioenergy grass Miscanthus species. Biotechnol. Biofuels 2021, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Mihaličová Malčovská, S.; Dučaiová, Z.; Maslaňáková, I.; Bačkor, M. Effect of silicon on growth, photosynthesis, oxidative status and phenolic compounds of maize (Zea mays L.) grown in cadmium excess. Water Air Soil Pollut. 2014, 225, 2056. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids. 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Wang, X.; Liu, H.; Yu, F.; Hu, B.; Jia, Y.; Sha, H.; Zhao, H. Differential activity of the antioxidant defence system and alterations in the accumulation of osmolyte and reactive oxygen species under drought stress and recovery in rice (Oryza sativa L.) tillering. Sci. Rep. 2019, 9, 8543. [Google Scholar] [CrossRef]
- Guo, H.; Hong, C.; Chen, X.; Xu, Y.; Liu, Y.; Jiang, D.; Zheng, B. Different growth and physiological responses to cadmium of the three Miscanthus species. PLoS ONE 2016, 11, e0153475. [Google Scholar] [CrossRef]
- Mafakheri, A.; Siosemardeh, A.F.; Bahramnejad, B.; Struik, P.C.; Sohrabi, Y. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust. J. Crop Sci. 2010, 4, 580–585. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Plant response to salt stress and role of exogenous protectants to mitigate salt-induced damages. In Ecophysiology and Responses of Plants under Salt Stress; Springer: New York, NY, USA, 2013; pp. 25–87. [Google Scholar]
- Medeiros, D.B.; Silva, E.C.D.; Santos, H.R.B.; Pacheco, C.M.; Musser, R.D.S.; Nogueira, R.J.M.C. Physiological and biochemical responses to drought stress in Barbados cherry. Braz. J. Plant Physiol. 2012, 24, 181–192. [Google Scholar] [CrossRef]
- Akther, M.S.; Das, U.; Tahura, S.; Prity, S.A.; Islam, M.; Kabir, A.H. Regulation of Zn uptake and redox status confers Zn deficiency tolerance in tomato. Sci. Hortic. 2020, 273, 109–624.77. [Google Scholar]
- Ozkur, O.; Ozdemir, F.; Bor, M.; Turkan, I. Physiochemical and antioxidant responses of the perennial xerophyte Capparis ovata Desf. to drought. Environ. Exp. Bot. 2009, 66, 487–492. [Google Scholar] [CrossRef]
- Csiszár, J. Effect of osmotic stress on antioxidant enzyme activities in transgenic wheat calli bearing MsALR gene. Acta Biol. Szeged. 2005, 49, 49–50. [Google Scholar]
- Sarathambal, C.; Khankhane, P.J.; Gharde, Y.; Kumar, B.; Varun, M.; Arun, S. The effect of plant growth-promoting rhizobacteria on the growth, physiology, and Cd uptake of Arundo donax L. Int. J. Phytoremediat. 2017, 19, 360–370. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Miscanthus Hybrids | Arundo Clones | |||
|---|---|---|---|---|
| Code | Type | Hybrid | Clones Code | Origin |
| GRC 1 | Seed-based plugs | M. sinensis × M. sinensis | A1 | Italy |
| GRC 3 | Seed-based plugs | M. sinensis × M. sinensis | ASR | Rome |
| GRC 6 | Seed-based plugs | M. sinensis × M. sinensis | CT2 | Sicily |
| GRC 9 | Rhizomes | Miscanthus × giganteus | PI1 | Tuscany |
| GRC 10 | Seed-based plugs | M. sinensis × M. sacchariflorus | PC1 | Piacenza |
| GRC 14 | Seed-based plugs | M. sinensis × M. sacchariflorus | PC6 | Piacenza |
| GRC 15 | Rhizomes | M. sinensis × M. sacchariflorus | PC7 | Piacenza |
| PH | NOL | SDW | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Miscanthus Hybrids | Control | Drought | Zn | Control | Drought | Zn | Control | Drought | Zn |
| GRC1 | 79.0 ± 7.8 a | 65.6 ± 3.0 b | 68.2 ± 2.0 b | 9.0 ± 1.0 a | 6.3 ± 0.5 b | 7.0 ± 1.4 b | 13.1 ± 3.9 a | 6.0 ± 2.1 b | 8.3 ± 0.5 b |
| GRC3 | 77.2 ± 2.2 a | 67.5 ± 3.6 b | 63.2 ± 2.2 b | 8.5 ± 0.5 a | 6.2 ± 0.5 b | 6.0 ± 0.8 b | 13.0 ± 3.5 a | 7.5 ± 1.5 b | 6.3 ± 1.2 b |
| GRC6 | 67.6 ± 1.5 a | 51.3 ± 3.2 b | 56.2 ± 3.4 b | 8.3 ± 0.5 a | 6.0 ± 0.3 b | 6.2 ± 0.5 b | 15.4 ± 2.3 a | 7.1 ± 1.2 b | 8.0 ± 0.5 b |
| GRC9 | 76.0 ± 2.6 a | 51.7 ± 3.3 b | 62.3 ± 1.5 c | 6.0 ± 0.4 a | 4.5 ± 0.5 b | 4.0 ± 0.4 b | 14.4 ± 1.8 a | 6.6 ± 0.6 b | 5.9 ± 1.8 b |
| GRC10 | 88.7 ± 4.5 a | 83.5 ± 3.1 a | 83.7 ± 2.6 a | 8.2 ± 0.5 a | 7.7 ± 0.5 a | 7.7 ± 0.5 a | 22.7 ± 5.3 a | 20.7 ± 4.0 a | 21.6 ± 2.8 a |
| GRC14 | 70.3 ± 1.5 a | 63.0 ± 3.0 b | 64.0 ± 1.8 b | 7.3 ± 0.5 a | 5.6 ± 0.5 b | 5.0 ± 0.4 b | 14.7 ± 2.3 a | 10.3 ± 1.7 b | 10.9 ± 1.7 b |
| GRC15 | 81.6 ± 2.5 a | 62.2 ± 2.2 b | 64.7 ± 2.2 b | 6.6 ± 0.5 a | 5.2 ± 0.5 b | 5.2 ± 0.5 b | 16.3 ± 0.3 a | 11.7 ± 0.6 b | 10.6 ± 1.7 b |
| Arundo clones | |||||||||
| A1 | 78.3 ± 2.8 a | 57.6 ± 1.1 b | 62.7 ± 2.2 c | 9.3 ± 0.5 a | 5.3 ± 0.5 b | 6.2 ± 0.8 b | 29.3 ± 0.5 a | 16.1 ± 1.9 b | 15.3 ± 1.6 b |
| ASR | 70.6 ± 1.5 a | 52.5 ± 1.2 b | 52.5 ± 3.5 b | 8.3 ± 0.5 a | 5.2 ± 0.9 b | 5.2 ± 0.9 b | 13.6 ± 0.6 a | 3.5 ± 0.3 b | 3.4 ± 0.2 b |
| CT2 | 45.7 ± 4.1 a | 26.2 ± 1.2 b | 30.0 ± 2.1 c | 6.2 ± 0.5 a | 4.2 ± 0.5 b | 3.5 ± 0.7 b | 13.9 ± 1.1 a | 1.4 ± 0.5 b | 1.5 ± 0.4 b |
| PI1 | 53.2 ± 1.7 a | 41.5 ± 1.2 b | 36.7 ± 1.5 c | 7.0 ± 0.8 a | 5.7 ± 0.5 b | 5.7 ± 0.5 b | 19.8 ± 1.0 a | 5.9 ± 0.4 b | 5.5 ± 0.7 b |
| PC1 | 104.0 ± 3.6 a | 97.0 ± 0.8 b | 91.2 ± 0.9 c | 10.7 ± 0.9 a | 9.0 ± 0.3 b | 10.0 ± 0.8 ab | 50.9 ± 4.2 a | 41.2 ± 0.7 b | 42.4 ± 1.2 b |
| PC6 | 77.5 ± 1.0 a | 55.2 ± 1.2 b | 64.7 ± 1.5 c | 8.5 ± 1.2 a | 5.7 ± 0.5 b | 6.2 ± 0.5 b | 35.5 ± 2.7 a | 13.0 ± 0.5 b | 13.7 ± 0.9 b |
| PC7 | 85.3 ± 1.5 a | 65.0 ± 1.0 b | 66.6 ± 1.5 b | 6.0 ± 1.0 a | 5.0 ± 0.8 a | 5.3 ± 0.6 a | 20.0 ± 0.2 a | 6.5 ± 0.7 b | 5.7 ± 0.7 b |
| Fv/Fm | PI-ABS | RWC% | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Miscanthus Hybrids | Control | Drought | Zn | Control | Drought | Zn | Control | Drought | Zn |
| GRC1 | 0.6 ± 0.03 a | 0.51 ± 0.05 b | 0.45 ± 0.03 c | 4.25 ± 0.03 a | 2.83 ± 0.39 b | 2.51 ± 0.21 c | 77.4 ± 2.32 a | 69.0 ± 1.79 b | 60.70 ± 3.89 c |
| GRC3 | 0.5 ± 0.01 a | 0.26 ± 0.01 b | 0.28 ± 0.01 b | 3.69 ± 0.26 a | 1.34 ± 0.08 b | 1.74 ± 0.24 c | 71.0 ± 5.40 a | 52.8 ± 3.56 b | 54.3 ± 4.96 b |
| GRC6 | 0.58 ± 0.00 a | 0.21 ± 0.07 b | 0.43 ± 0.01 c | 3.52 ± 0.21 a | 1.12 ± 0.38 b | 2.03 ± 0.15 c | 83.8 ± 3.61 a | 71.4 ± 0.84 b | 73.5 ± 2.33 b |
| GRC9 | 0.59 ± 0.02 a | 0.43 ± 0.09 b | 0.25 ± 0.01 c | 3.51 ± 0.12 a | 2.36 ± 0.49 b | 1.31 ± 0.03 c | 84.0 ± 1.47 a | 73.7 ± 0.42 b | 77.1 ± 0.81 ab |
| GRC10 | 0.64 ± 0.01 a | 0.60 ± 0.02 a | 0.60 ± 0.04 ab | 4.34 ± 0.27 a | 3.91 ± 0.29 b | 3.90 ± 0.45 b | 97.2 ± 7.04 a | 87.2 ± 6.06 b | 88.6 ± 6.88 b |
| GRC14 | 0.51 ± 0.02 a | 0.38 ± 0.01 b | 0.28 ± 0.04 c | 3.68 ± 0.16 a | 2.05 ± 0.12 b | 1.52 ± 0.22 c | 94.6 ± 4.91 a | 85.9 ± 3.54 b | 75.5 ± 3.44 c |
| GRC15 | 0.53 ± 0.03 a | 0.38 ± 0.01 b | 0.27 ± 0.03 c | 3.74 ± 0.28 a | 2.19 ± 0.17 b | 1.46 ± 0.17 c | 83.9 ± 3.35 a | 76.8 ± 1.96 a | 65.1 ± 2.94 b |
| Arundo clones | |||||||||
| A1 | 0.81 ± 0.02 a | 0.65 ± 0.07 b | 0.70 ± 0.01 b | 5.26 ± 1.67 a | 2.94 ± 1.28 b | 2.44 ± 0.50 b | 90.7 ± 0.08 a | 75.9 ± 0.33 b | 79.0 ± 0.69 c |
| ASR | 0.78 ± 0.07 a | 0.73 ± 0.04 a | 0.70 ± 0.06 a | 2.62 ± 1.03 a | 0.74 ± 0.72 b | 0.57 ± 0.35 b | 92.9 ± 0.24 a | 75.4 ± 0.78 b | 77.3 ± 0.85 b |
| CT2 | 0.80 ± 0.03 a | 0.67 ± 0.06 b | 0.68 ± 0.05 b | 4.22 ± 2.25 a | 3.16 ± 1.49 a | 2.46 ± 1.20 ab | 88.7 ± 0.41 a | 64.1 ± 0.64 b | 58.8 ± 1.88 c |
| PI1 | 0.81 ± 0.03 a | 0.50 ± 0.10 b | 0.40 ± 0.25 b | 4.78 ± 1.57 a | 2.13 ± 0.72 b | 3.73 ± 2.04 ab | 93.1 ± 0.97 a | 65.5 ± 0.53 b | 71.2 ± 0.26 c |
| PC1 | 0.82 ± 0.01 a | 0.82 ± 0.02 a | 0.81 ± 0.01 a | 7.25 ± 0.91 a | 6.78 ± 0.32 a | 6.98 ± 2.31 a | 92.4 ± 1.93 a | 76.8 ± 1.61 b | 84.5 ± 1.81 c |
| PC6 | 0.81 ± 0.03 a | 0.78 ± 0.05 a | 0.80 ± 0.02 a | 4.93 ± 0.83 a | 4.10 ± 2.17 a | 3.74 ± 1.47 a | 90.2 ± 0.40 a | 65.9 ± 1.89 b | 75.8 ± 1.19 c |
| PC7 | 0.81 ± 0.01 a | 0.56 ± 0.08 b | 0.66 ± 0.08 b | 4.52 ± 1.24 a | 3.14 ± 0.86 a | 2.57 ± 1.41 a | 87.0 ± 1.70 a | 67.0 ± 1.55 b | 73.1 ± 1.83 c |
| Variable | PH | NOL | SDW | Fv/Fm | PI-ABS | RWC% | Protein | PPO | POD | SOD | APX | MDA | Phenol | Proline |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Miscanthus hybrids | ||||||||||||||
| Treatments | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| Hybrids | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| Treatments × hybrids | *** | * | NS | *** | *** | ** | *** | *** | *** | *** | *** | *** | *** | *** |
| Arundo clones | ||||||||||||||
| Treatments | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| Clones | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| Treatments × clones | *** | ** | *** | *** | NS | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| Miscanthus | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Drought | Zn | ||||||||||
| Hybrids | PC1 | PC2 | PC3 | Ranking | Numeric Rank | Hybrids | PC1 | PC2 | PC3 | Ranking | Numeric Rank |
| GRC10 | 15.54 | −0.85 | −0.28 | 8.11 | 1 | GRC10 | 18.24 | −0.33 | −0.62 | 11.34 | 1 |
| GRC15 | 1.25 | 1.42 | 1.56 | 2.14 | 2 | GRC14 | −0.33 | 2.24 | 2.26 | 1.32 | 2 |
| GRC14 | 1.13 | 1.91 | 0.44 | 1.96 | 3 | GRC6 | −0.02 | 0.51 | −1.92 | −0.16 | 3 |
| GRC1 | −1.69 | −2.38 | −1.92 | −1.58 | 4 | GRC1 | −1.05 | −3.50 | 1.90 | −0.95 | 4 |
| GRC6 | −4.87 | 1.37 | 2.09 | −2.09 | 5 | GRC15 | −2.86 | 1.87 | 0.66 | −1.45 | 5 |
| GRC9 | −5.15 | 2.71 | −2.70 | −2.60 | 6 | GRC9 | −6.30 | 1.67 | −1.09 | −3.84 | 6 |
| GRC3 | −6.21 | −4.18 | 0.82 | −3.97 | 7 | GRC3 | −7.69 | −2.47 | −1.19 | −5.32 | 7 |
| Arundo | |||||||||||
| Drought | Zn | ||||||||||
| Clones | PC1 | PC2 | PC3 | Ranking | Numeric Rank | Clones | PC1 | PC2 | PC3 | Ranking | Numeric Rank |
| PC1 | 19.30 | 0.06 | 0.51 | 12.22 | 1 | PC1 | 18.04 | 0.48 | −0.12 | 11.39 | 1 |
| ASR | 0.99 | 6.18 | 9.41 | 2.45 | 2 | PC6 | 1.22 | −1.92 | 0.79 | 1.56 | 2 |
| A1 | −1.53 | −1.86 | −0.89 | −1.33 | 3 | A1 | −0.79 | −2.04 | 0.25 | −0.79 | 3 |
| PC6 | −2.44 | −2.41 | −0.30 | −1.93 | 4 | ASR | −1.60 | −0.17 | 0.22 | −1.01 | 4 |
| PI1 | −3.80 | 1.64 | 2.18 | −1.94 | 5 | PI1 | −3.16 | 4.65 | −0.80 | −1.33 | 5 |
| PC7 | −5.19 | −2.34 | 0.64 | −3.57 | 6 | PC7 | −6.14 | −1.64 | −2.49 | −4.34 | 6 |
| CT2 | −5.84 | 2.37 | 0.20 | −3.31 | 7 | CT2 | −9.76 | 0.57 | 2.13 | −5.84 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.; Ferrarini, A.; Ali, A.; Kam, J.; Trindade, L.M.; Clifton-Brown, J.; Amaducci, S. Assessment of Drought and Zinc Stress Tolerance of Novel Miscanthus Hybrids and Arundo donax Clones Using Physiological, Biochemical, and Morphological Traits. Biology 2023, 12, 1525. https://doi.org/10.3390/biology12121525
Islam M, Ferrarini A, Ali A, Kam J, Trindade LM, Clifton-Brown J, Amaducci S. Assessment of Drought and Zinc Stress Tolerance of Novel Miscanthus Hybrids and Arundo donax Clones Using Physiological, Biochemical, and Morphological Traits. Biology. 2023; 12(12):1525. https://doi.org/10.3390/biology12121525
Chicago/Turabian StyleIslam, Monirul, Andrea Ferrarini, Amjad Ali, Jason Kam, Luisa M. Trindade, John Clifton-Brown, and Stefano Amaducci. 2023. "Assessment of Drought and Zinc Stress Tolerance of Novel Miscanthus Hybrids and Arundo donax Clones Using Physiological, Biochemical, and Morphological Traits" Biology 12, no. 12: 1525. https://doi.org/10.3390/biology12121525