
Multidrug Resistance in Enterococci Isolated from Cheese and Capable of Producing Benzalkonium Chloride-Resistant Biofilms

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Enterococcus Isolates from Cheeses
2.2. Antibiotic Resistance Profile, Virulence, and Resistance Genes
2.3. Evaluation of the Susceptibility to Benzalkonium Chloride (BAC)
2.4. Evaluation of Biofilm-Forming Ability
2.5. Minimum Biofilm Eradication Concentration (MBEC) of BAC
2.6. Data Analysis and Interpretation
3. Results
3.1. Antibiotic Resistance Profile, Antibiotic Resistance Genes, and Virulence Genes
3.2. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) of Benzalkonium Chloride (BAC)
3.3. Biofilm-Forming Ability According to Incubation Time and Temperature
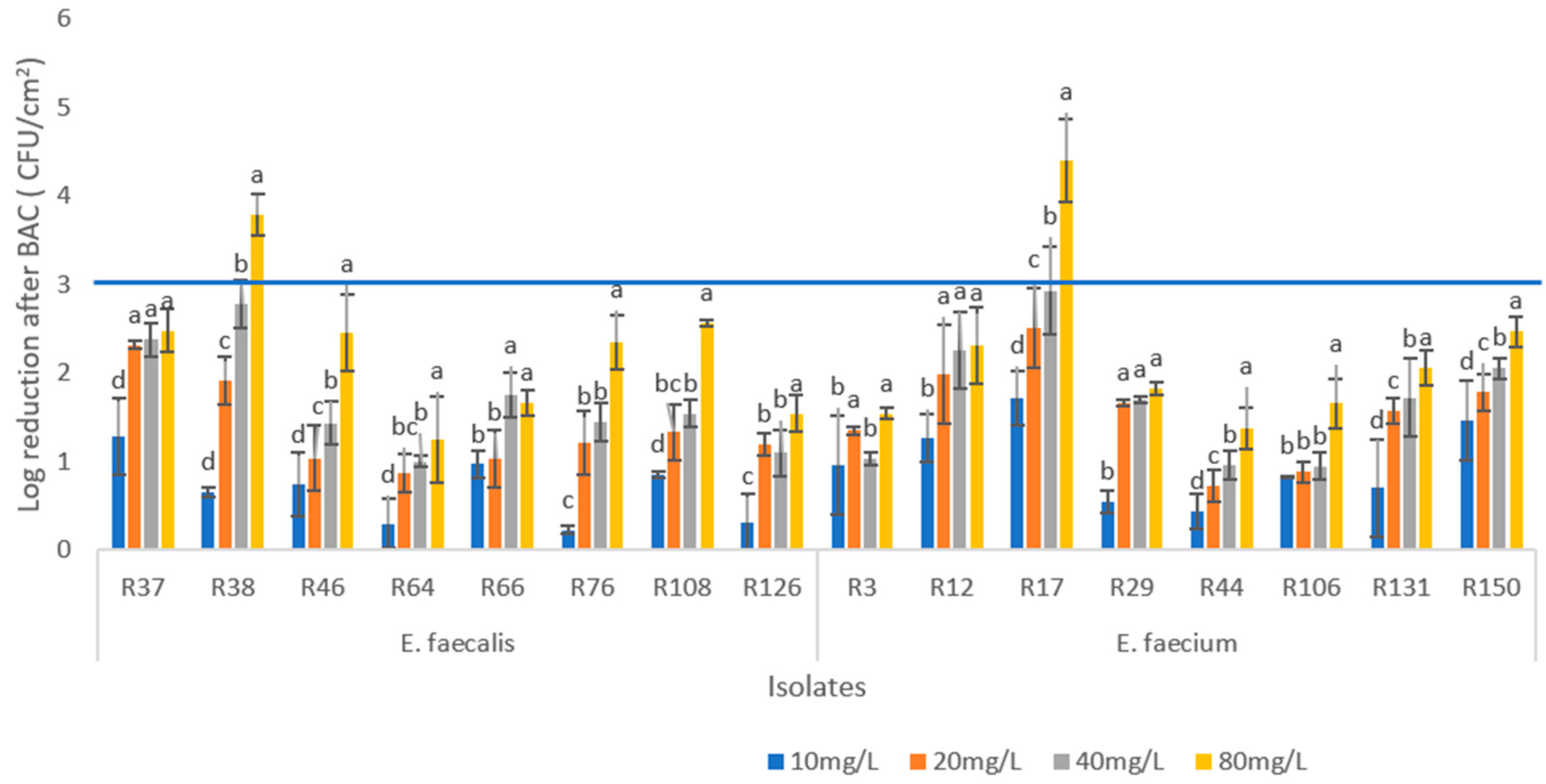
3.4. Assessment of the Minimum Biofilm Eradication Concentration by BAC (MBEC)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubin, K.; Pamer, E.G. Enterococci and Their Interactions with the Intestinal Microbiome. Microbiol. Spectr. 2017, 5, 5–6. [Google Scholar] [CrossRef]
- Coelho, M.C.; Malcata, F.X.; Silva, C.C.G. Lactic Acid Bacteria in Raw-Milk Cheeses: From Starter Cultures to Probiotic Functions. Foods 2022, 11, 2276. [Google Scholar] [CrossRef]
- Ben Braïek, O.; Smaoui, S. Enterococci: Between Emerging Pathogens and Potential Probiotics. Biomed. Res. Int. 2019, 2019, 5938210. [Google Scholar] [CrossRef] [PubMed]
- Berreta, A.; Baumgardner, R.M.; Kopper, J.J. Evaluation of Commercial Veterinary Probiotics Containing Enterococci for Transferrable Vancomycin Resistance Genes. BMC Res. Notes 2020, 13, 275. [Google Scholar] [CrossRef] [PubMed]
- Nami, Y.; Haghshenas, B.; Vaseghi Bakhshayesh, R.; Mohammadzadeh Jalaly, H.; Lotfi, H.; Eslami, S.; Hejazi, M.A. Novel Autochthonous Lactobacilli with Probiotic Aptitudes as a Main Starter Culture for Probiotic Fermented Milk. LWT 2018, 98, 85–93. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Forssten, S.; Hibberd, A.A.; Lyra, A.; Stahl, B. Probiotic Approach to Prevent Antibiotic Resistance. Ann. Med. 2016, 48, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Terzić-Vidojević, A.; Veljović, K.; Popović, N.; Tolinački, M.; Golić, N. Enterococci from Raw-Milk Cheeses: Current Knowledge on Safety, Technological, and Probiotic Concerns. Foods 2021, 10, 2753. [Google Scholar] [CrossRef]
- Shao, Y.; Zhen, W.; Guo, F.; Hu, Z.; Zhang, K.; Kong, L.; Guo, Y.; Wang, Z. Pretreatment with Probiotics Enterococcus faecium NCIMB 11181 Attenuated Salmonella Typhimurium-Induced Gut Injury through Modulating Intestinal Microbiome and Immune Responses with Barrier Function in Broiler Chickens. J. Anim. Sci. Biotechnol. 2022, 13, 130. [Google Scholar] [CrossRef]
- Wu, Y.; Zhen, W.; Geng, Y.; Wang, Z.; Guo, Y. Pretreatment with Probiotic Enterococcus faecium NCIMB 11181 Ameliorates Necrotic Enteritis-Induced Intestinal Barrier Injury in Broiler Chickens. Sci. Rep. 2019, 9, 10256. [Google Scholar] [CrossRef]
- Zhang, F.; Qiu, L.; Xu, X.; Liu, Z.; Zhan, H.; Tao, X.; Shah, N.P.; Wei, H. Beneficial Effects of Probiotic Cholesterol-Lowering Strain of Enterococcus faecium WEFA23 from Infants on Diet-Induced Metabolic Syndrome in Rats. J. Dairy. Sci. 2017, 100, 1618–1628. [Google Scholar] [CrossRef]
- Xu, W.; Zou, K.; Zhan, Y.; Cai, Y.; Zhang, Z.; Tao, X.; Qiu, L.; Wei, H. Enterococcus faecium GEFA01 Alleviates Hypercholesterolemia by Promoting Reverse Cholesterol Transportation via Modulating the Gut Microbiota-SCFA Axis. Front. Nutr. 2022, 9, 1020734. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Xie, X.; Li, Y.; Wu, L.; Fan, C.; Liang, T.; Xi, Y.; Yang, S.; Li, H.; Zhang, J.; et al. Evaluation of the Cholesterol-Lowering Mechanism of Enterococcus faecium Strain 132 and Lactobacillus paracasei Strain 201 in Hypercholesterolemia Rats. Nutrients 2021, 13, 1982. [Google Scholar] [CrossRef]
- Scheb-Wetzel, M.; Rohde, M.; Bravo, A.; Goldmann, O. New Insights into the Antimicrobial Effect of Mast Cells against Enterococcus faecalis. Infect. Immun. 2014, 82, 4496–4507. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, Y.; Xue, M.; Xiao, Z.; Fan, Y.; Zeng, L.; Zhou, Y. Effects of Dietary Enterococcus faecalis YFI-G720 on the Growth, Immunity, Serum Biochemical, Intestinal Morphology, Intestinal Microbiota, and Disease Resistance of Crucian Carp (Carassius auratus). Fishes 2022, 7, 18. [Google Scholar] [CrossRef]
- Hu, Y.; Dun, Y.; Li, S.; Zhang, D.; Peng, N.; Zhao, S.; Liang, Y. Dietary Enterococcus faecalis LAB31 Improves Growth Performance, Reduces Diarrhea, and Increases Fecal Lactobacillus Number of Weaned Piglets. PLoS ONE 2015, 10, e0116635. [Google Scholar] [CrossRef]
- Pinkes, M.E.; White, C.; Wong, C.S. Native-Valve Enterococcus hirae Endocarditis: A Case Report and Review of the Literature. BMC Infect. Dis. 2019, 19, 891. [Google Scholar] [CrossRef]
- Shankar, N.; Lockatell, C.V.; Baghdayan, A.S.; Drachenberg, C.; Gilmore, M.S.; Johnson, D.E. Role of Enterococcus faecalis Surface Protein Esp in the Pathogenesis of Ascending Urinary Tract Infection. Infect. Immun. 2001, 69, 4366–4372. [Google Scholar] [CrossRef] [PubMed]
- Barnes, A.M.T.; Frank, K.L.; Dunny, G.M. Enterococcal Endocarditis: Hiding in Plain Sight. Front. Cell Infect. Microbiol. 2021, 11, 722482. [Google Scholar] [CrossRef]
- Fiore, E.; Van Tyne, D.; Gilmore, M.S. Pathogenicity of Enterococci. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Bonilla, N.; Santiago, T.; Marcos, P.; Urdaneta, M.; Domingo, J.S.; Toranzos, G.A. Enterophages, a Group of Phages Infecting Enterococcus faecalis, and Their Potential as Alternate Indicators of Human Faecal Contamination. Water Sci. Technol. 2010, 61, 293–300. [Google Scholar] [CrossRef]
- Tran, N.H.; Gin, K.Y.-H.; Ngo, H.H. Fecal Pollution Source Tracking Toolbox for Identification, Evaluation and Characterization of Fecal Contamination in Receiving Urban Surface Waters and Groundwater. Sci. Total Environ. 2015, 538, 38–57. [Google Scholar] [CrossRef]
- Santiago-Rodriguez, T.M.; Marcos, P.; Monteiro, S.; Urdaneta, M.; Santos, R.; Toranzos, G.A. Evaluation of Enterococcus-Infecting Phages as Indices of Fecal Pollution. J. Water Health 2013, 11, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Jamet, E.; Akary, E.; Poisson, M.A.; Chamba, J.F.; Bertrand, X.; Serror, P. Prevalence and Characterization of Antibiotic Resistant Enterococcus faecalis in French Cheeses. Food Microbiol. 2012, 31, 191–198. [Google Scholar] [CrossRef]
- Amidi-Fazli, N.; Hanifian, S. Biodiversity, Antibiotic Resistance and Virulence Traits of Enterococcus Species in Artisanal Dairy Products. Int. Dairy. J. 2022, 129, 105287. [Google Scholar] [CrossRef]
- Elsayed, S.; Hamilton, N.; Boyd, D.; Mulvey, M.; Kariyama, R.; Mitsuhata, R.; Kumon, H. Improved Primer Design for Multiplex PCR Analysis of Vancomycin-Resistant Enterococcus spp. J. Clin. Microbiol. 2001, 39, 2367–2368. [Google Scholar] [CrossRef] [PubMed]
- Semedo-Lemsaddek, T.; Nóbrega, C.S.; Ribeiro, T.; Pedroso, N.M.; Sales-Luís, T.; Lemsaddek, A.; Tenreiro, R.; Tavares, L.; Vilela, C.L.; Oliveira, M. Virulence Traits and Antibiotic Resistance among Enterococci Isolated from Eurasian Otter (Lutra lutra). Vet. Microbiol. 2013, 163, 378–382. [Google Scholar] [CrossRef]
- Marinho, A.R.; Martins, P.D.; Ditmer, E.M.; d’Azevedo, P.A.; Frazzon, J.; Der Sand, S.T.V.; Frazzon, A.P.G. Biofilm Formation on Polystyrene under Different Temperatures by Antibiotic Resistant Enterococcus faecalis and Enterococcus faecium Isolated from Food. Braz. J. Microbiol. 2013, 44, 423–426. [Google Scholar] [CrossRef]
- Hashem, Y.A.; Amin, H.M.; Essam, T.M.; Yassin, A.S.; Aziz, R.K. Biofilm Formation in Enterococci: Genotype-Phenotype Correlations and Inhibition by Vancomycin. Sci. Rep. 2017, 7, 5733. [Google Scholar] [CrossRef]
- Wolfe, B.E.; Button, J.E.; Santarelli, M.; Dutton, R.J. Cheese Rind Communities Provide Tractable Systems for In Situ and In Vitro Studies of Microbial Diversity. Cell 2014, 158, 422–433. [Google Scholar] [CrossRef]
- Eaton, T.J.; Gasson, M.J. Molecular Screening of Enterococcus Virulence Determinants and Potential for Genetic Exchange between Food and Medical Isolates. Appl. Env. Microbiol. 2001, 67, 1628–1635. [Google Scholar] [CrossRef]
- Semedo, T.; Santos, M.A.; Fátima, M.; Lopes, S.; Marques, J.J.F.; Crespo, M.T.B.; Tenreiro, R. Virulence Factors in Food, Clinical and Reference Enterococci: A Common Trait in the Genus? Syst. Appl. Microbiol. 2003, 26, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Salamandane, A.; Silva, A.C.; Brito, L.; Malfeito-Ferreira, M. Microbiological Assessment of Street Foods at the Point of Sale in Maputo (Mozambique). Food Qual. Saf. 2021, 5, fyaa030. [Google Scholar] [CrossRef]
- ISO 6887-1; Microbiology of the Food Chain—Preparation of Test Samples, Initial Suspension and Decimal Dilutions for Microbiological Examination—Part 1: General Rules for the Preparation of the Initial Suspension and Decimal Dilutions. International Standard: Geneva, Switzerland, 2017.
- Rocha, P.A.B.; Marques, J.M.M.; Barreto, A.S.; Semedo-Lemsaddek, T. Enterococcus spp. from Azeitão and Nisa PDO-Cheeses: Surveillance for Antimicrobial Drug Resistance. LWT 2022, 154, 112622. [Google Scholar] [CrossRef]
- Salamandane, A.; Malfeito-Ferreira, M.; Brito, L. A High Level of Antibiotic Resistance in Klebsiella and Aeromonas Isolates from Street Water Sold in Mozambique, Associated with the Prevalence of Extended-Spectrum and AmpC ß-Lactamases. J. Environ. Sci. Health Part. B 2022, 57, 561–567. [Google Scholar] [CrossRef]
- CLSI. An Informational Supplement for Global Application Developed through the Clinical and Laboratory Standards Institute Consensus Process. In M100 Performance Standards for Antimicrobial Susceptibility Testing; CLSI: Wayne, PA, USA, 2021; Volume 27. [Google Scholar]
- Semedo-Lemsaddek, T.; Bettencourt Cota, J.; Ribeiro, T.; Pimentel, A.; Tavares, L.; Bernando, F.; Oliveira, M. Resistance and Virulence Distribution in Enterococci Isolated from Broilers Reared in Two Farming Systems. Ir. Vet. J. 2021, 74, 22. [Google Scholar] [CrossRef]
- Salamandane, A.; Oliveira, J.; Coelho, M.; Ramos, B.; Cunha, M.V.; Malfeito-Ferreira, M.; Brito, L. Enterotoxin- and Antibiotic-Resistance-Encoding Genes Are Present in Both Coagulase-Positive and Coagulase-Negative Foodborne Staphylococcus Strains. Appl. Microbiol. 2022, 2, 367–380. [Google Scholar] [CrossRef]
- Morrissey, I.; Oggioni, M.R.; Knight, D.; Curiao, T.; Coque, T.; Kalkanci, A.; Martinez, J.L.; Baldassarri, L.; Orefici, G.; Yetiş, Ü.; et al. Evaluation of Epidemiological Cut-off Values Indicates That Biocide Resistant Subpopulations Are Uncommon in Natural Isolates of Clinically-Relevant Microorganisms. PLoS ONE 2014, 9, e86669. [Google Scholar] [CrossRef]
- Costa, A.; Bertolotti, L.; Brito, L.; Civera, T. Biofilm Formation and Disinfectant Susceptibility of Persistent and Non-Persistent Listeria Monocytogenes Isolates from Gorgonzola Cheese Processing Plants. Foodborne Pathog. Dis. 2016, 13, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Salamandane, A.; Correia, J.; Muetanene, B.A.; dos Santos, M.; Malfeito-Ferreira, M.; Brito, L. Methicillin Resistance of Food-Borne Biofilm-Forming Staphylococci. Appl. Sci. 2023, 13, 7725. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Salamandane, A.; Alves, S.; Chambel, L.; Malfeito-Ferreira, M.; Brito, L. Characterization of Escherichia Coli from Water and Food Sold on the Streets of Maputo: Molecular Typing, Virulence Genes, and Antibiotic Resistance. Appl. Microbiol. 2022, 2, 133–147. [Google Scholar] [CrossRef]
- Palmer, K.L.; Kos, V.N.; Gilmore, M.S. Horizontal Gene Transfer and the Genomics of Enterococcal Antibiotic Resistance. Curr. Opin. Microbiol. 2010, 13, 632–639. [Google Scholar] [CrossRef]
- Výrostková, J.; Regecová, I.; Dudriková, E.; Marcinčák, S.; Vargová, M.; Kováčová, M.; Mal’ová, J. Antimicrobial Resistance of Enterococcus sp. Isolated from Sheep and Goat Cheeses. Foods 2021, 10, 1844. [Google Scholar] [CrossRef]
- Cetinkaya, Y.; Falk, P.; Mayhall, C.G. Vancomycin-Resistant Enterococci. Clin. Microbiol. Rev. 2000, 13, 686–707. [Google Scholar] [CrossRef]
- Werner, G. Current Trends of Emergence and Spread of Vancomycin-Resistant Enterococci. In Antibiotic Resistant Bacteria—A Continuous Challenge in the New Millennium; IntechOpen: London, UK, 2012. [Google Scholar]
- Tian, Y.; Yu, H.; Wang, Z. Distribution of Acquired Antibiotic Resistance Genes among Enterococcus spp. Isolated from a Hospital in Baotou, China. BMC Res. Notes 2019, 12, 27. [Google Scholar] [CrossRef]
- Saribas, Z.; Tunckanat, F.; Pinar, A. Prevalence of Erm Genes Encoding Macrolide-Lincosamide-Streptogramin (MLS) Resistance among Clinical Isolates of Staphylococcus Aureus in a Turkish University Hospital. Clin. Microbiol. Infect. 2006, 12, 797–799. [Google Scholar] [CrossRef] [PubMed]
- Kiruthiga, A.; Padmavathy, K.; Shabana, P.; Naveenkumar, V.; Gnanadesikan, S.; Malaiyan, J. Improved Detection of esp, hyl, asa1, gelE, cylA Virulence Genes among Clinical Isolates of Enterococci. BMC Res. Notes 2020, 13, 170. [Google Scholar] [CrossRef] [PubMed]
- Hashem, Y.A.; Abdelrahman, K.A.; Aziz, R.K. Phenotype–Genotype Correlations and Distribution of Key Virulence Factors in Enterococcus faecalis Isolated from Patients with Urinary Tract Infections. Infect. Drug Resist. 2021, 14, 1713–1723. [Google Scholar] [CrossRef] [PubMed]
- Gajewska, J.; Chajęcka-Wierzchowska, W.; Byczkowska-Rostkowska, Z.; Saki, M. Biofilm Formation Capacity and Presence of Virulence Determinants among Enterococcus Species from Milk and Raw Milk Cheeses. Life 2023, 13, 495. [Google Scholar] [CrossRef]
- Grudlewska-Buda, K.; Skowron, K.; Bauza-Kaszewska, J.; Budzyńska, A.; Wiktorczyk-Kapischke, N.; Wilk, M.; Wujak, M.; Paluszak, Z. Assessment of Antibiotic Resistance and Biofilm Formation of Enterococcus Species Isolated from Different Pig Farm Environments in Poland. BMC Microbiol. 2023, 23, 89. [Google Scholar] [CrossRef]
- Bonneville, L.; Ortiz, S.; Maia, V.; Brito, L.; Martínez-Suárez, J.V. Strain and Growth Conditions May Regulate Resistance of Listeria Monocytogenes Biofilms to Benzalkonium Chloride. Appl. Sci. 2020, 10, 988. [Google Scholar] [CrossRef]
- Mogana, R.; Adhikari, A.; Tzar, M.N.; Ramliza, R.; Wiart, C. Antibacterial Activities of the Extracts, Fractions and Isolated Compounds from Canarium Patentinervium Miq. against Bacterial Clinical Isolates. BMC Complement. Med. Ther. 2020, 20, 55. [Google Scholar] [CrossRef] [PubMed]
- Barroso, I.; Maia, V.; Cabrita, P.; Martínez-Suárez, J.V.; Brito, L. The Benzalkonium Chloride Resistant or Sensitive Phenotype of Listeria Monocytogenes Planktonic Cells Did Not Dictate the Susceptibility of Its Biofilm Counterparts. Food Res. Int. 2019, 123, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Van Tyne, D.; Martin, M.; Gilmore, M. Structure, Function, and Biology of the Enterococcus faecalis Cytolysin. Toxins 2013, 5, 895–911. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Isolate Code | Species | Antibiotic Resistance | Virulence Genes | |
|---|---|---|---|---|
| Phenotype | Genotype | |||
| R37 | E. faecalis | RD, ERY | ermC, vanA | FsrB, gelE, cylA |
| R38 | E. faecalis | TET, ERY, VAN | ermC, tetM, vanA | fsrB, gelE, esp, agg |
| R46 | E. faecalis | VAN | vanA, vanB | FsrB, gelE |
| R64 | E. faecalis | TET, RD, ERY, VAN | ermC, tetM, vanA, pbp5 | fsrB, gelE, cylA, esp |
| R66 | E. faecalis | P, TET, RD, ERY, VAN | ermC, tetM, vanA, blaZ | FsrB, gelE, cylA |
| R76 | E. faecalis | P, TET, ERY, RD, VAN | ermC, vanA, blaZ | fsrB, gelE, esp, agg |
| R108 | E. faecalis | TET, RD, ERY, VAN | tetM, ermC, vanA | fsrB, gelE, cylA, esp, agg |
| R126 | E. faecalis | TET, ERY, VAN | ermC, tetM, vanA | fsrB, gelE, cylA |
| R3 | E. faecium | TET, ERY, VAN | ermA, ermC, tetM, vanA | fsrB, gelE, cylA, esp |
| R12 | E. faecium | RD | pbp5 | fsrB, gelE, cylA |
| R17 | E. faecium | GEN, ERY, VAN | ermA, ermC, tetM, vanA | fsrB, gelE, esp |
| R29 | E. faecium | P, VAN | vanA | fsrB, gelE |
| R44 | E. faecium | ERY, VAN | vanA, pbp5 | fsrB, gelE, cylA, esp |
| R106 | E. faecium | P, RD, ERY, VAN | vanA, blaZ, pbp5 | fsrB, gelE, esp |
| R131 | E. faecium | P, GEN, RD, VAN | ermC, vanA, pbp5 | fsrB, gelE |
| R150 | E. faecium | P, TET, ERY, VAN | ermC, vanA, tetM, pbp5 | fsrB, gelE, cylA, esp |
| Isolate Code | Species | MIC (mg/L) | MBC (mg/L) | MBC/MIC |
|---|---|---|---|---|
| R37 | E. faecalis | 10 | 20 | 2 |
| R38 | E. faecalis | 20 | 20 | 1 |
| R46 | E. faecalis | 20 | 20 | 1 |
| R64 | E. faecalis | 10 | 10 | 1 |
| R66 | E. faecalis | 5 | 10 | 2 |
| R76 | E. faecalis | 5 | 10 | 2 |
| R108 | E. faecalis | 5 | 10 | 2 |
| R126 | E. faecalis | 5 | 10 | 2 |
| R3 | E. faecium | 5 | 10 | 2 |
| R12 | E. faecium | 20 | 40 | 2 |
| R17 | E. faecium | 20 | 40 | 2 |
| R29 | E. faecium | 20 | 40 | 2 |
| R44 | E. faecium | 20 | 40 | 2 |
| R106 | E. faecium | 10 | 20 | 2 |
| R131 | E. faecium | 5 | 10 | 2 |
| R150 | E. faecium | 5 | 10 | 2 |
| Isolate Code | Species | Biofilm-Forming Ability (Log CFU/cm2) | p-Value | |||
|---|---|---|---|---|---|---|
| 15 °C, 72 h | 37 °C, 24 h | 37 °C, 48 h | 37 °C, 72 h | |||
| R37 | E. faecalis | 7.66 ± 0.31 Bb | 6.40 ± 0.09 Dd | 7.07 ± 0.06 Cb | 8.82 ± 0.09 Aa | 3.7 × 10−6 |
| R38 | E. faecalis | 7.13 ± 0.05 Cd | 7.40 ± 0.04 Ba | 7.29 ± 0.02 Bb | 8.82 ± 0.10 Aa | 9.99 × 10−9 |
| R46 | E. faecalis | 7.25 ± 0.01 Cc | 7.13 ± 0.14 Cb | 7.60 ± 0.16 Ba | 8.89 ± 0.07 Aa | 1.1 × 10−6 |
| R64 | E. faecalis | 6.92 ± 0.16 Cd | 7.10 ± 0.05 Cb | 7.92 ± 0.07 Ba | 8.89 ± 0.07 Aa | 1.3 × 10−7 |
| R66 | E. faecalis | 7.87 ± 0.08 Ba | 6.76 ± 0.28 Cc | 7.87 ± 0.09 Ba | 8.78 ± 0.14 Aa | 1.8 × 10−5 |
| R76 | E. faecalis | 7.06 ± 0.14 Cd | 6.50 ± 0.01 Dc | 7.75 ± 0.10 Ba | 8.91 ± 0.06 Aa | 2.57 × 10−8 |
| R108 | E. faecalis | 7.52 ± 0.05 Cb | 7.28 ± 0.21 Ca | 7.81 ± 0.04 Ba | 8.89 ± 0.08 Aa | 3.1 × 10−6 |
| R126 | E. faecalis | 6.97 ± 0.07 Cd | 6.72 ± 0.11 Cc | 7.70 ± 0.16 Ba | 9.04 ± 0.18 Aa | 7.8 × 10−7 |
| R3 | E. faecium | 7.28 ± 0.13 Bc | 6.21 ± 0.08 Cd | 7.90 ± 0.06 Aa | 8.15 ± 0.46 Ab | 0.00019 |
| R12 | E. faecium | 6.69 ± 0.09 Be | 6.65 ± 0.11 Bc | 7.41 ± 0.38 Ab | 8.01 ± 0.77 Ab | 0.0418 |
| R17 | E. faecium | 7.31 ± 0.15 Bc | 7.48 ± 0.34 ABa | 7.49 ± 0.45 ABb | 7.96 ± 0.61 Ab | 0.0486 |
| R29 | E. faecium | 7.51 ± 0.02 Ab | 6.38 ± 0.11 Bd | 7.18 ± 0.01 Bb | 7.70 ± 0.27 Ab | 9.3 × 10−5 |
| R44 | E. faecium | 7.21 ± 0.11 Bc | 6.93 ± 0.05 Bb | 7.84 ± 0.21 Aa | 8.18 ± 0.45 Ab | 0.00415 |
| R106 | E. faecium | 6.82 ± 0.04 Be | 6.98 ± 0.04 Bb | 7.78 ± 0.02 Aa | 7.90 ± 0.05 Ab | 0.0418 |
| R131 | E. faecium | 6.67 ± 0.03 Ce | 6.15 ± 0.05 Dd | 7.24 ± 0.06 Bb | 7.93 ± 0.08 Ab | 2.3 × 10−9 |
| R150 | E. faecium | 7.28 ± 0.08 Bc | 7.08 ± 0.08 Bb | 7.22± 0.15 Bb | 7.79 ± 0.25 Ab | 0.0091 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salamandane, A.; Cahango, G.; Muetanene, B.A.; Malfeito-Ferreira, M.; Brito, L. Multidrug Resistance in Enterococci Isolated from Cheese and Capable of Producing Benzalkonium Chloride-Resistant Biofilms. Biology 2023, 12, 1353. https://doi.org/10.3390/biology12101353
Salamandane A, Cahango G, Muetanene BA, Malfeito-Ferreira M, Brito L. Multidrug Resistance in Enterococci Isolated from Cheese and Capable of Producing Benzalkonium Chloride-Resistant Biofilms. Biology. 2023; 12(10):1353. https://doi.org/10.3390/biology12101353
Chicago/Turabian StyleSalamandane, Acácio, Gomes Cahango, Belo Afonso Muetanene, Manuel Malfeito-Ferreira, and Luísa Brito. 2023. "Multidrug Resistance in Enterococci Isolated from Cheese and Capable of Producing Benzalkonium Chloride-Resistant Biofilms" Biology 12, no. 10: 1353. https://doi.org/10.3390/biology12101353