
Dental Paleobiology in a Juvenile Neanderthal (Combe-Grenal, Southwestern France)
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Brief Reminder of Tartar (Dental Calculus) Etiology
3. Materials and Methods
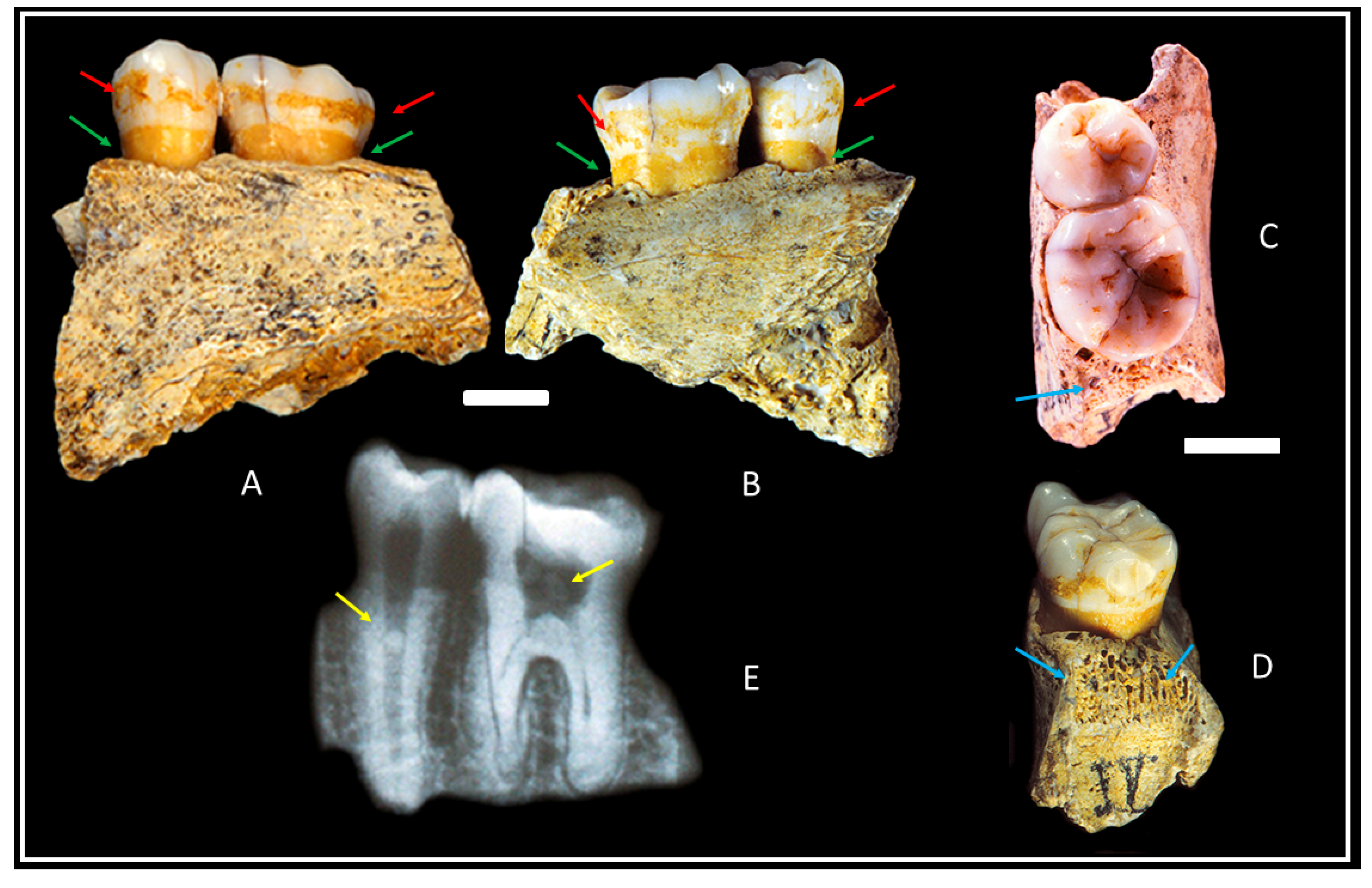
3.1. Materials
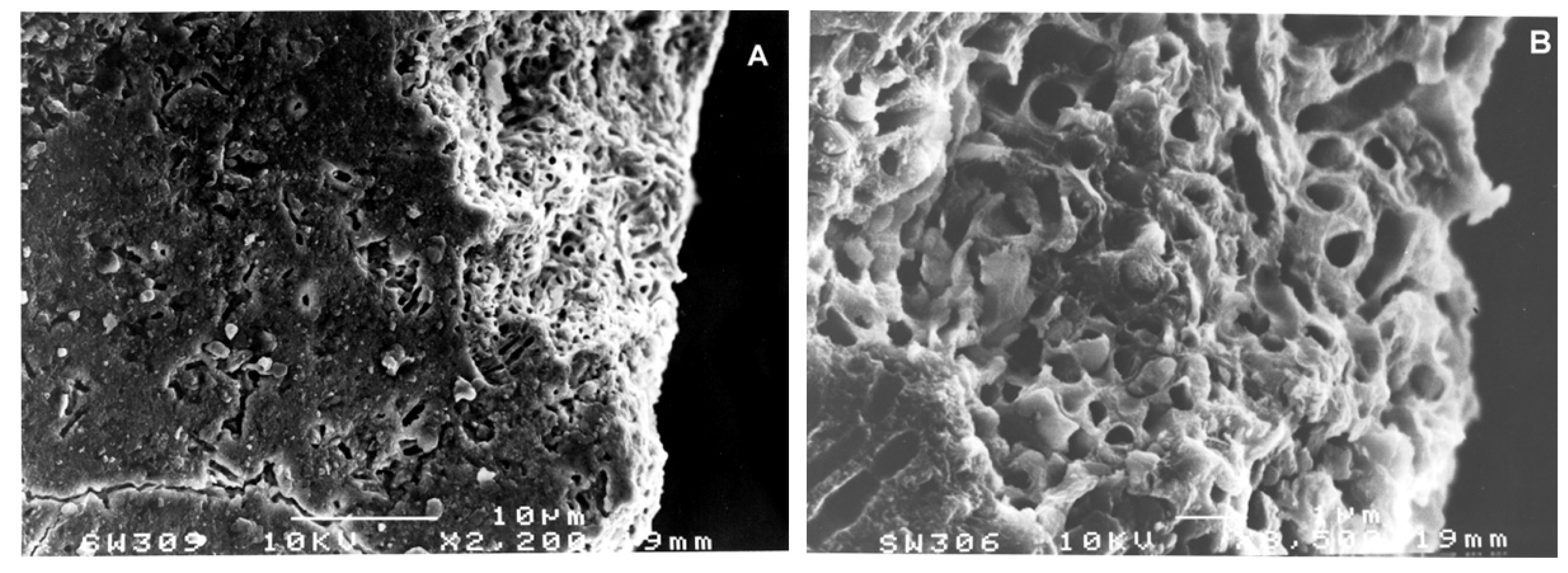
3.2. Methods
4. Results
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bordes, F. La stratigraphie de la grotte de Combe-Grenal (Dordogne). Note préliminaire. Bull. Soc. Préhistorique Française 1955, 52, 426–429. [Google Scholar] [CrossRef]
- Bordes, F. A Tale of Two Caves; Harper & Row: New York, NY, USA, 1972. [Google Scholar]
- Bordes, F.; Prat, F. Observations sur les faunes du Riss et du Würm I en Dordogne. l’Anthropologie 1965, 69, 31–46. [Google Scholar]
- Bordes, F.; Laville, H.; Paquerau, M.M. Observations sur le Pléistocéne supérieur du gisement de Combe-Grenal (Dordogne). Actes Soc. Linnéenne Bordx. 1966, 103, 3–19. [Google Scholar]
- Guadelli, J.L.; Laville, H. L’environnement climatique de la fin du Moustérien à Combe-Grenal et à Camiac. Confrontation des Données Naturalistes et Implications. In Paléolithique Moyen Récent et Paléolithique Superieur Ancien en Europe; Farizy, C., Ed.; Mém. du Musée de Préhistoire d’Ile de France: Nemours, France, 1990; pp. 43–48. [Google Scholar]
- Garralda, M.D.; Vandermeersch, B. Les Néandertaliens de la Grotte de Combe-Grenal (Domme, France). Paléo Rev. D’archéologie Préhistorique 2000, 12, 213–259. [Google Scholar]
- Garralda, M.D.; Giacobini, G.; Vandermeersch, B. Neanderthal cutmarks: Combe-Grenal and Marillac (France). A SEM analysis. Anthropologie 2005, 43, 189–197. [Google Scholar]
- Maureille, B.; Garralda, M.D.; Madelaine, S.; Turq, A.; Vandermeersch, B. Le plus ancien enfant d’Aquitaine, Combe-Grenal 31 (Domme, France). PALEO. Rev. D’archéologie Préhistorique 2011, 21, 189–202. [Google Scholar] [CrossRef]
- Gómez-Olivencia, A.; Garralda, M.D.; Vandermeersch, B.; Madelaine, S.; Arsuaga, J.-L.; Maureille, B. Two newly identified mousterian human rib fragments from Combe-Grenal. PALEO. Rev. D’archéologie Préhistorique 2013, 24, 229–235. [Google Scholar] [CrossRef]
- Lang, N.P.; Mombelli, A.; Attström, R. Dental Plaque and Calculus. In Clinical Periodontology and Implant Dentistry, 3rd ed.; Lindle, J., Karring, T., Lang, N.P., Eds.; Munsksgaard: Copenhagen, Denmark, 1998; pp. 102–137. [Google Scholar]
- Ottoni, C.; Guelli, M.; Ozga, A.T.; Stone, A.C.; Kersten, O.; Bramanti, B.; Porcier, B.; Van Neer, W. Metagenomic analysis of dental calculus in ancient Egyptian baboons. Sci. Rep. 2019, 9, 19637. [Google Scholar] [CrossRef]
- Gleize, Y.; Castex, D.; Duday, H.; Chapoulie, R. Analyse préliminaire et discussion sur la nature d’un dépôt dentaire très particulier. Bull. Mémoires Société d’Anthropologie Paris. BMSAP 2005, 17, 5–12. [Google Scholar] [CrossRef]
- Verger-Pratoucy, J.-C. Commentaires sur l’article: Analyse préliminaire et discussion sur la nature d’un dépôt dentaire très particulier. Bull. Mémoires Société d’Anthropologie Paris. BMSAP 2007, 19, 95–102. [Google Scholar]
- Smith, B.H. Patterns of Molar Wear in Hunter-Gatherers and Agriculturalists. Amer. J. Phys. Anthropol. 1984, 63, 39–56. [Google Scholar] [CrossRef]
- Ubelaker, D. Human Skeletal Remains, 2nd ed.; Taraxacum Press: Washington, DC, USA, 1989. [Google Scholar]
- AlQahtani, S.J.; Hector, M.P.; Liversidge, H.M. The London Atlas of Human Tooth Development and Eruption. Am. J. Phys. Anthropol. 2010, 142, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Lavigne, S.E.; Molto, J.E. System of measurement of the severity of periodontal disease in Past Populations. Int. J. Osteoarch. 1995, 5, 265–273. [Google Scholar] [CrossRef]
- Brothwell, D. Digging Up Bones, 3rd ed.; Cornell University Press: Ithaca, NY, USA, 1981. [Google Scholar]
- Dietrich, P.; Ower, P.; Tank, M.; West, N.X.; Walter, C.; Needleman, I.; Hughes, F.J.; Wadia, R.; Milward, M.R.; Hodge, P.J.; et al. Periodontal diagnosis in the context of the 2017 classification system of periodontal diseases and conditions—Implementation in clinical practice. Br. Dent. J. 2019, 226, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Vandermeersch, B.; Arensburg, B.; Tillier, A.-M.; Rak, Y.; Weiner, S.; Spiers, M.; Aspillaga, E. Middle Paleolithic dental Bacteria from Kebara, Israel. Comptes Rendus L Acad. Sci. Ser. Ii 1994, 319, 727–731. [Google Scholar]
- Littner, M.; Tillier, A.-M.; Arensburg, B.; Kaffe, I. A Middle Paleolithic mandible from Kebara Israel (60.000 BP) in View of Oral Health of Modern Humans. J. Isr. Dent. Assoc. 1996, 13, 1. [Google Scholar]
- Lindhe, J.; Karring, T.; Lang, N.P. Clinical Periodontology and Implant Dentistry, 3rd ed.; Munksgaard: Copenhagen, Denmark, 1998. [Google Scholar]
- Pap, I.; Tillier, A.-M.; Arensburg, B.; Weiner, S.; Chech, M. First scanning electron microscope analysis of dental calculus from European Neanderthals: Subalyuk (Middle Paleolithic, Hungary). Bull. Mémoires Société d'Anthropologie Paris 1995, 7, 69–72. [Google Scholar] [CrossRef]
- Hillson, S. Dental Anthropology; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Ripamonti, U. Paleopathology in Australopithecus africanus: A suggested case of a 3 million-year-old prepubertal periodontitis. Amer. J. Phys. Anthropol. 1988, 76, 197–210. [Google Scholar] [CrossRef]
- Bartucz, L.; Dancza, J.; Hollendonner, F.; Kadic, O.; Mottl, M.; Pataki, V.; Palosi, E.; Szabo, J.; Vendl, A. Die Mussolini-Höhle (Subalyuk) bei Cserépfalu; Editio Instituti Regni Hungarici Geologici: Budapest, Hungary, 1939; Volume 14. [Google Scholar]
- Bar-Yosef, O. Middle Paleolithic Adaptations in the Mediterranean Levant. In The Evolution and Dispersal of Modern Humans in Asia; Akazawa, T., Aoki, K., Kimura, T., Eds.; Hokusen-Sha: Tokyo, Japan, 1992; pp. 189–216. [Google Scholar]
- Bocherens, H.; Billiou, D.; Patou-Mathis, M.; Otte, M.; Bonjean, D.; Toussaint, M.; Mariotti, A. Palaeoenvironmental and palaeoditary implications of isotopic biogeochemistry of late interglacial Neandertal and mammal bones in Scladina Cave (Belgium). J. Archeol. Sci. 1999, 26, 599–607. [Google Scholar] [CrossRef]
- Bocherens, H. Isotopic biogeochemistry as a marker of Neandertal diet. Anthrop. Anz. 1997, 55, 101–120. [Google Scholar] [CrossRef]
- Richards, M.P.; Pettitt, P.B.; Trinkaus, E.; Smith, F.H.; Paunovic, M.; Karavanic, I. Neanderthal diet at Vindija and Neanderthal predation: The evidence from stable isotopes. Proc. Natl. Acad. Sci. USA 2000, 97, 7663–7666. [Google Scholar] [CrossRef] [PubMed]
- Balter, V.; Simon, L. Diet and behavior of the Saint-Césaire Neanderthal inferred from biogeochemical data inversion. J. Hum. Evol. 2006, 51, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Bocherens, H.; Drucker, D.; Billiou, D.; Pathou-Mathis, M.; Vandermeersch, B. Isotopic evidence for diet and subsistence pattern of the Saint-Césaire 1 Neanderthal review and use of a multi-source mixing model. J. Hum. Evol. 2005, 49, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Richards, M.P.; Taylor, G.; Steele, T.; McPherron, S.; Soressi, M.; Jaubert, J.; Orschiedt, J.; Mallye, J.-B.; Rendu, W.; Hublin, J.-J. Isotopic analysis of a Neanderthal and associated fauna from the site of Jonzac (Charente-Maritime, France). J. Hum. Evol. 2008, 55, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Wibing, C.; Rougier, H.; Baumann, C.; Comeyne, A.; Crevecoeur, I.; Drucker, D.; Gaudzinski-Windheuser, S.; Germonpré, M.; Gomez-Olivencia, A.; Krause, J.; et al. Stable isotopes reveal patterns of diet and mobility in the last Neanderthls and first modern humans in Europe. Sci. Rep. 2019, 9, 4433. [Google Scholar] [CrossRef]
- Dodat, P.-J.; Tacail, T.; Albalat, E.; Gómez-Olivencia, A.; Couture-Veschambre, C.; Holliday, T.; Madelaine, S.; Martin, J.E.; Rmoutilova, R.; Maureille, B.; et al. The dietary reconstruction of the Regourdou 1 Neandertal (MIS 5, France) using bone calcium isotopes. J. Hum. Evol. 2021, 10, 21230. [Google Scholar]
- Bar-Yosef, O. Eat what is there: Hunting and Gathering in the World of Neanderthals and their Neighbours. Int. J. Osteoarch. 2004, 14, 333–342. [Google Scholar] [CrossRef]
- Lev, E.; Kislev, M.E.; Bar-Yosef, O. Mousterian vegetal food in Kebara Cave, Mt. Carmel. J. Archaeol. Sci. 2005, 32, 475–484. [Google Scholar] [CrossRef]
- Hardy, K.; Buckley, S.; Collins, M.J.; Estalrrich, A.; Brothwel, D.; Copeland, L.; García-Tabernero, A.; García-Vargas, S.; de la Rasilla, M.; Lalueza-Fox, C.; et al. Neanderthal medics? Evidence for food, cooking, and medicinal plants entrapped in dental calculus. Naturwissenschaften 2012, 99, 617–626. [Google Scholar] [CrossRef]
- Naito, Y.I.; Chikaraishi, Y.; Drucker, D.G.; Ohkouchi, N.; Semal, P.; Wißing, C.; Bocherens, H. Ecological niche of Neanderthals from Spy Cave revealed by nitrogen isotopes of individual amino acids in collagen. J. Hum. Evol. 2016, 93, 82–90. [Google Scholar] [CrossRef]
- Duday, H.; Arensburg, B. La Pathologie. In Le Squelette Moustérien de Kébara 2; Bar-Yosef, O., Vandermeersch, B., Eds.; Cahiers de Paléoanthropologie, CNRS: Paris, France, 1991; pp. 179–193. [Google Scholar]
- Lozano, M.; Subirà, M.E.; Aparicio, J.; Lorenzo, C.; Gómez-Merino, G. Toothpicking and periodontal disease in a Neanderthal specimen from Cova Foradà site (Valencia, Spain). PLoS ONE 2013, 8, e76852. [Google Scholar] [CrossRef] [PubMed]
- van der Merwe, N.J.; Williamson, R.F.; Pfeiffer, S.; Thomas, S.C.; Allegretto, K.O. The Moatfield ossuary: Isotopic dietary analysis of an Iroquoian community, using dental tissue. J. Anthropol. Archaeol. 2003, 22, 245–261. [Google Scholar] [CrossRef]
- Schoeninger, M.J.; Reeser, H.; Hallin, K. Paleoenvironment of Australopithecus anamensis at Allia Bay, East Turkana, Kenya: Evidence from mammalian herbivore enamel stable isotopes. J. Anthropol. Archaeol. 2003, 22, 200–207. [Google Scholar] [CrossRef]
- Lee-Thorp, J.; Sponheimer, M. Three case studies used to reassess the reliability of fossil bone and enamel isotope signals for paleodietary studies. J. Anthropol. Archaeol. 2003, 22, 208–216. [Google Scholar] [CrossRef]
- Richards, M.P.; Schmitz, R.W. Isotope evidence for the diet of the Neanderthal type specimen. Antiquity 2008, 82, 553–559. [Google Scholar] [CrossRef]
- Bocherens, H. Diet and Ecology: Implications from C and N isotopes. In Neanderthal Lifeways, Subsistence and Technology; Conard, N., Richter, N.H., Eds.; Vertebrate Paleobiology and Paleoanthropology Series; Springer: Dordrecht, The Netherlands, 2011. [Google Scholar] [CrossRef]
- Bocherens, H.; Díaz-Zorita, M.; Daujeard, C.; Fernandes, P.; Raynal, J.-P.; Moncel, M.-H. Direct Isotopic evidence for subsistence variability in Middle Pleistocene Neanderthals (Payre, southeastern France). Quat. Sci. Rev. 2016, 154, 226–236. [Google Scholar] [CrossRef]
- Harrison, R.G.; Katzenberg, M.A. Paleodiet studies using stable carbon isotopes from bone apatite and collagen: Examples from Southern Ontario and San Nicolas Island, California. J. Anthropol. Archaeol. 2003, 22, 227–244. [Google Scholar] [CrossRef]
- Weyrich, L.S.; Duchene, S.; Soubrier, J.; Arriola, L.; Llamas, B.; Breen, J.; Morris, A.G.; Alt, K.W.; Caramelli, D.; Dresely, V.; et al. Neanderthal behaviour, diet, and disease inferred from ancient DNA in dental calculus. Nature 2017, 554, 357. [Google Scholar] [CrossRef]
- Charlier, P.; Gaultier, F.; Héry-Arnaud, G. Interbreeding between Neanderthals and modern humans: Remarks and methodological dangers of a dental calculus microbioma analysis. J. Hum.Evol. 2019, 126, 124–126. [Google Scholar] [CrossRef]
- Mann, A.E.; Fellows Yates, J.A.; Fagernäs, Z.; Austin, R.M.; Nelson, E.A.; Hofman, C.A. Do I have something in my teeth? The trouble with genetic analyses of diet from archaeological dental calculus. Quat. Int. 2022; in press. [Google Scholar] [CrossRef]
- Costamagno, S.; Meignen, L.; Cédric, B.; Vandermeersch, B.; Maureille, B. Les Pradelles (Marillac-le-Franc, France): A Mousterian reindeer hunting camp? J. Anthrop. Archaeol. 2006, 26, 466–484. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurements | N | Mean | Std. Dev. | Std. Error | Minimum | Maximum | Variance | |
|---|---|---|---|---|---|---|---|---|
| C. Grenal 304 (10.000 M) | Diameter | 16 | 0.493 | 0.109 | 0.027 | 0.360 | 0.700 | 0.012 |
| Length | 7 | 0.791 | 0.262 | 0.099 | 0.590 | 1.340 | 0.069 | |
| C.-Grenal 306 (3.500 M) | Diameter | 31 | 0.601 | 0.138 | 0.025 | 0.310 | 0.850 | 0.019 |
| Length | 10 | 1.608 | 0.472 | 0.149 | 1.050 | 2.550 | 0.222 | |
| C.-Grenal 308 (20.000 M) | Diameter | 15 | 0.477 | 0.165 | 0.043 | 0.330 | 0.840 | 0.027 |
| Length | 7 | 1.499 | 0.376 | 0.142 | 1.160 | 2.200 | 0.141 | |
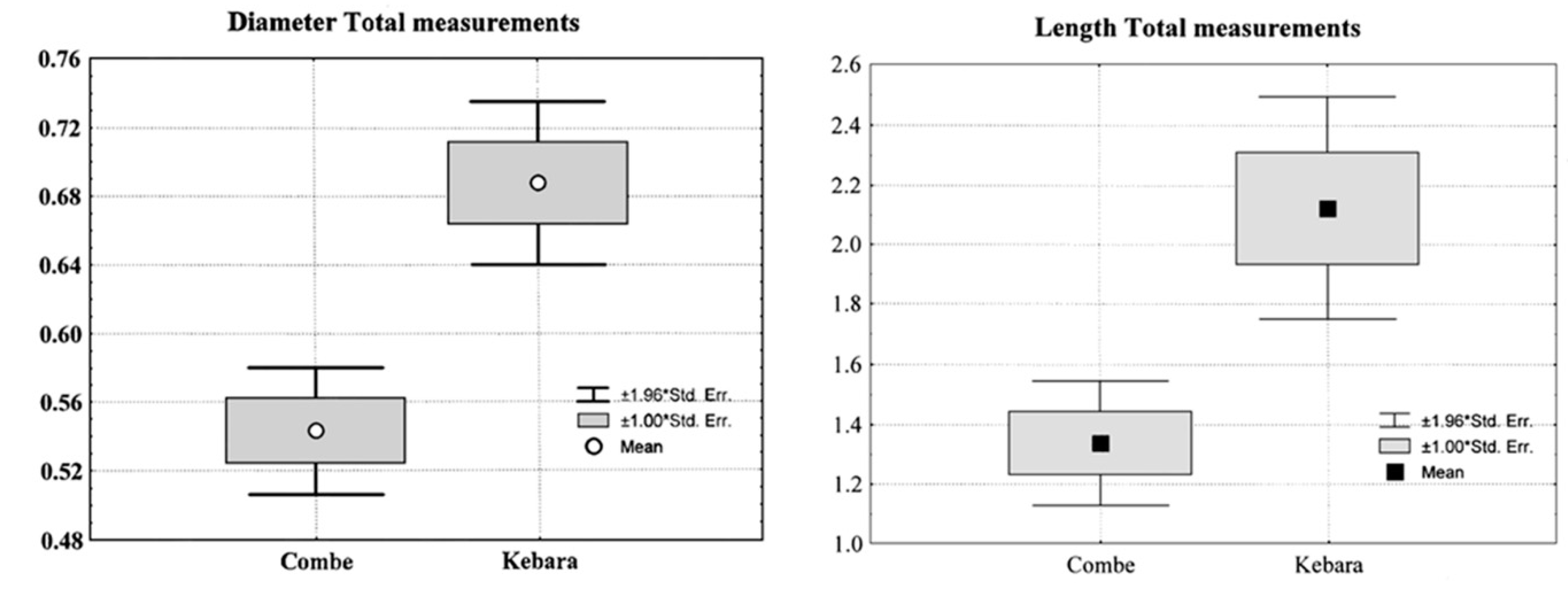
| Total C.-Grenal IV | Diameter | 62 | 0.543 | 0.149 | 0.019 | 0.330 | 0.850 | 0.022 |
| Length | 24 | 1.338 | 0.522 | 0.107 | 0.590 | 2.550 | 0.272 |
| Pictures | Measurements | N | Mean | Std. Dev. | Std. Error | Minimum | Maximum | Variance |
|---|---|---|---|---|---|---|---|---|
| Kebara 601 (10.000 M) | Diameter | 20 | 0.718 | 0.161 | 0.036 | 0.350 | 0.940 | 0.026 |
| Length | 18 | 1.241 | 0.422 | 0.100 | 0.830 | 2.360 | 0.178 | |
| Kebara 603 (5.000 M) | Diameter | 17 | 0.825 | 0.154 | 0.037 | 0.570 | 1.140 | 0.024 |
| Length | 16 | 3.116 | 0.719 | 0.180 | 1.940 | 4.310 | 0.517 | |
| Kebara 606 (5.000 M) | Diameter | 22 | 0.555 | 0.142 | 0.030 | 0.400 | 0.810 | 0.020 |
| Length | 0 | |||||||
| Total Kebara | Diameter | 59 | 0.687 | 0.187 | 0.024 | 0.400 | 1.140 | 0.035 |
| Length | 34 | 2.123 | 1.109 | 0.190 | 0.830 | 4.310 | 1.229 |
| Individuals | Traits | N | Mean | Std. Error | P |
|---|---|---|---|---|---|
| Total Kebara 2 | Diameter | 59 | 0.687 | 0.187 | 0.0001 |
| Total Combe-Grenal IV | 62 | 0.543 | 0.149 | ||
| Total Kebara 2 | Length | 34 | 2.123 | 1.109 | 0.02 |
| Total Combe-Grenal IV | 24 | 1.338 | 0.522 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garralda, M.D.; Weiner, S.; Arensburg, B.; Maureille, B.; Vandermeersch, B. Dental Paleobiology in a Juvenile Neanderthal (Combe-Grenal, Southwestern France). Biology 2022, 11, 1352. https://doi.org/10.3390/biology11091352
Garralda MD, Weiner S, Arensburg B, Maureille B, Vandermeersch B. Dental Paleobiology in a Juvenile Neanderthal (Combe-Grenal, Southwestern France). Biology. 2022; 11(9):1352. https://doi.org/10.3390/biology11091352
Chicago/Turabian StyleGarralda, María Dolores, Steve Weiner, Baruch Arensburg, Bruno Maureille, and Bernard Vandermeersch. 2022. "Dental Paleobiology in a Juvenile Neanderthal (Combe-Grenal, Southwestern France)" Biology 11, no. 9: 1352. https://doi.org/10.3390/biology11091352