
Comparison of Gut Bacterial Communities of Locusta migratoria manilensis (Meyen) Reared on Different Food Plants

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. DNA Extraction
2.3. Metagenomic Analysis
2.4. Statistical and Bioinformatic Analysis
3. Results
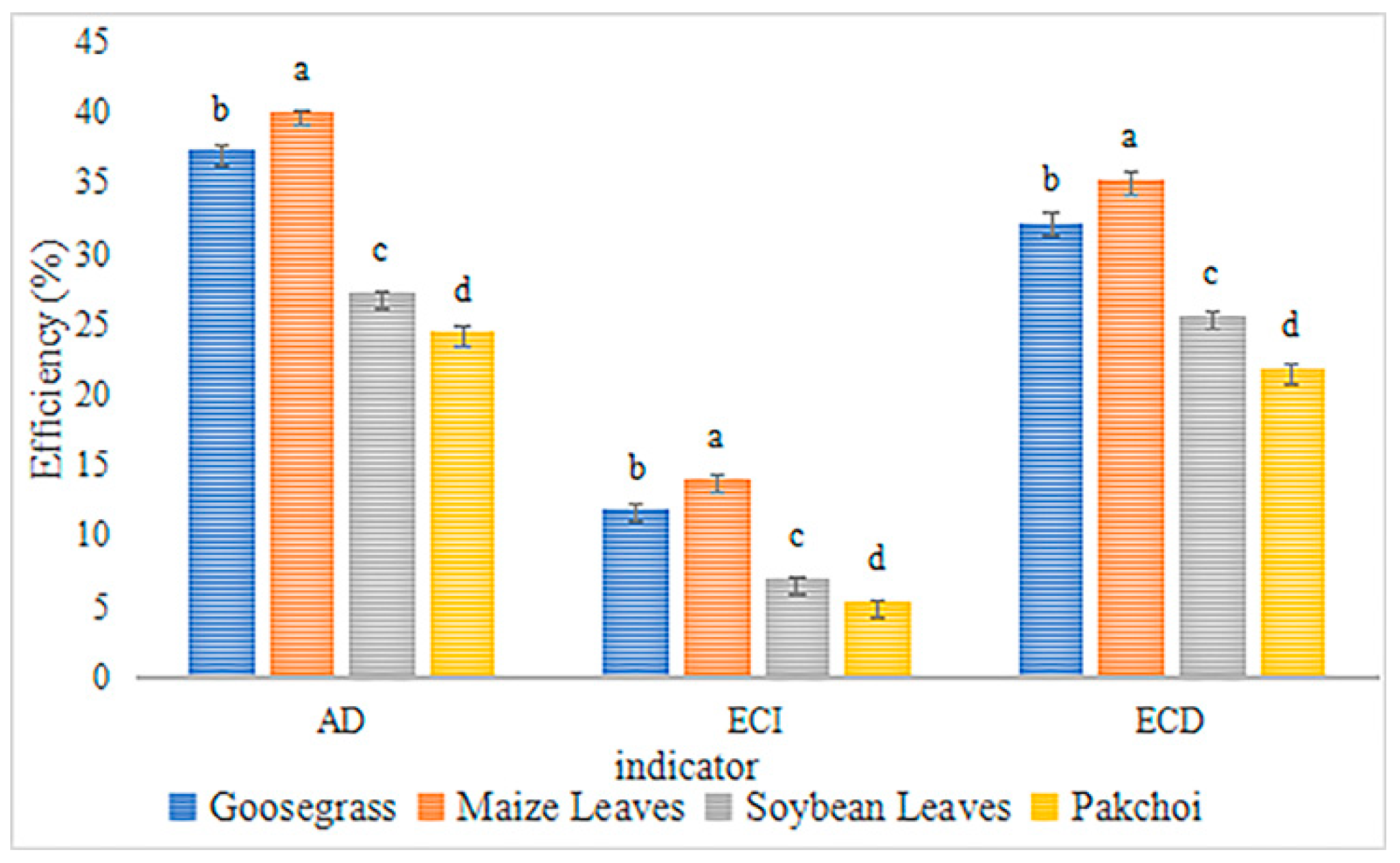
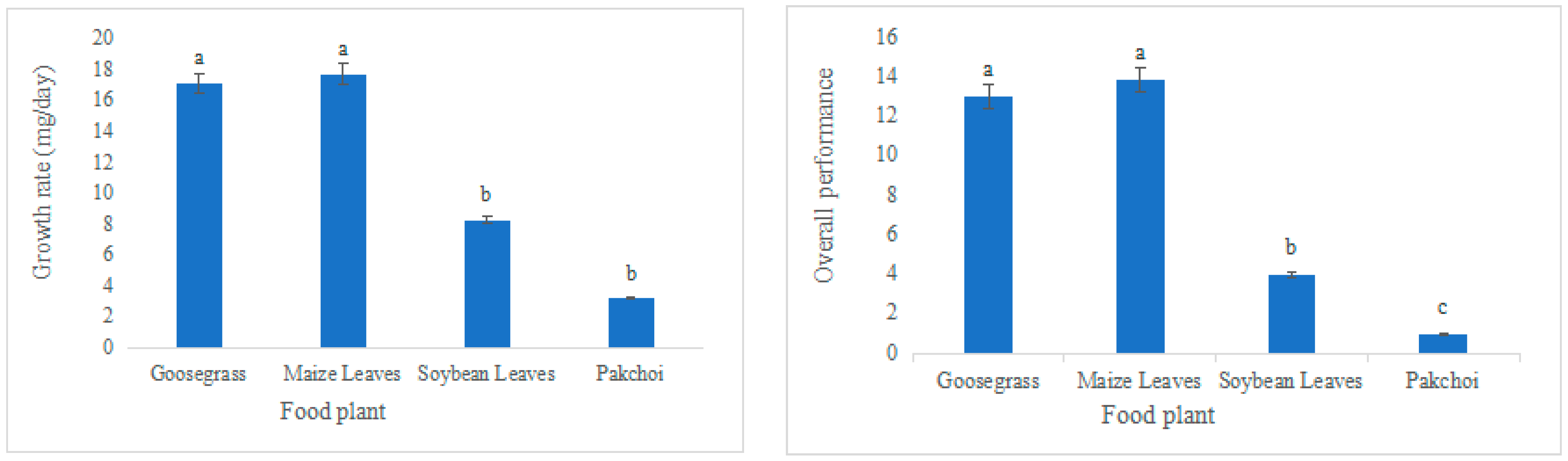
3.1. Development Rate of L. migratoria manilensis Reared on Different Food Plant
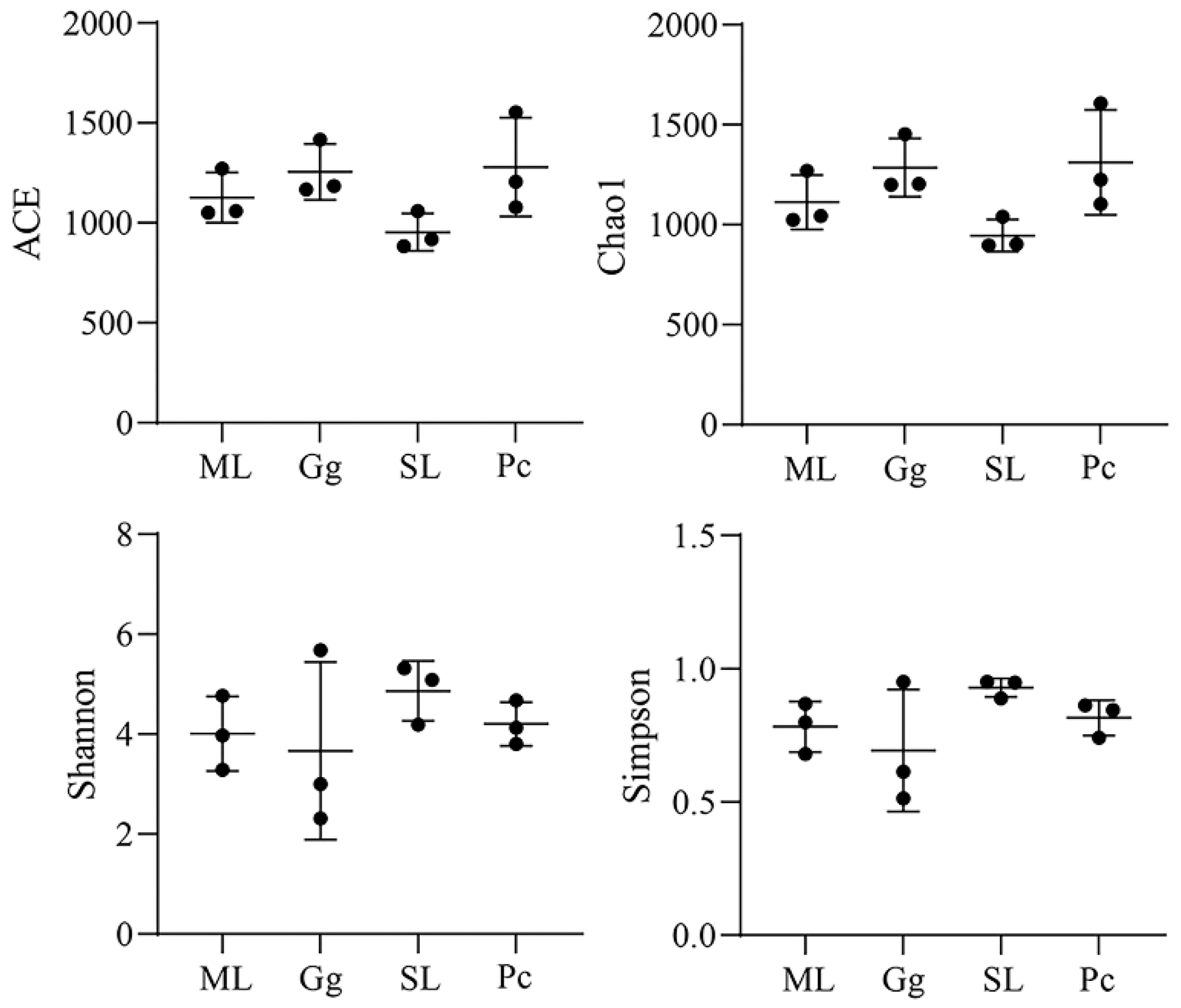
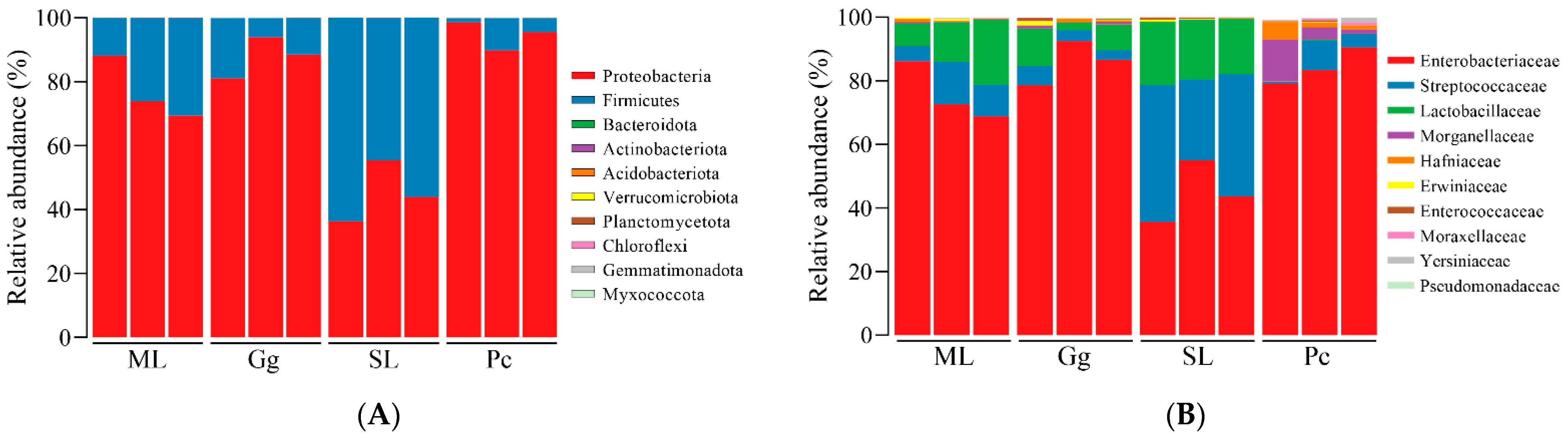
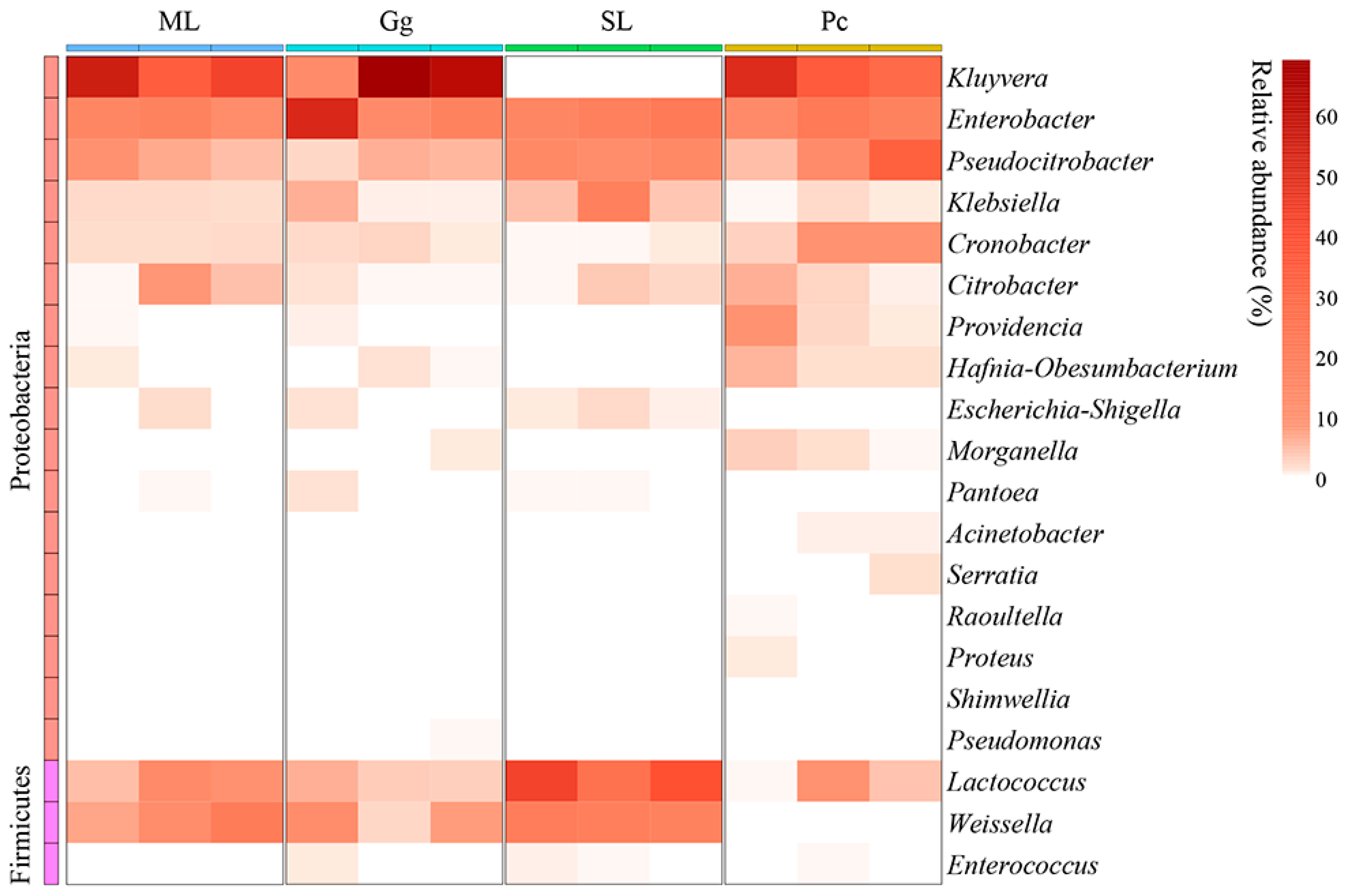
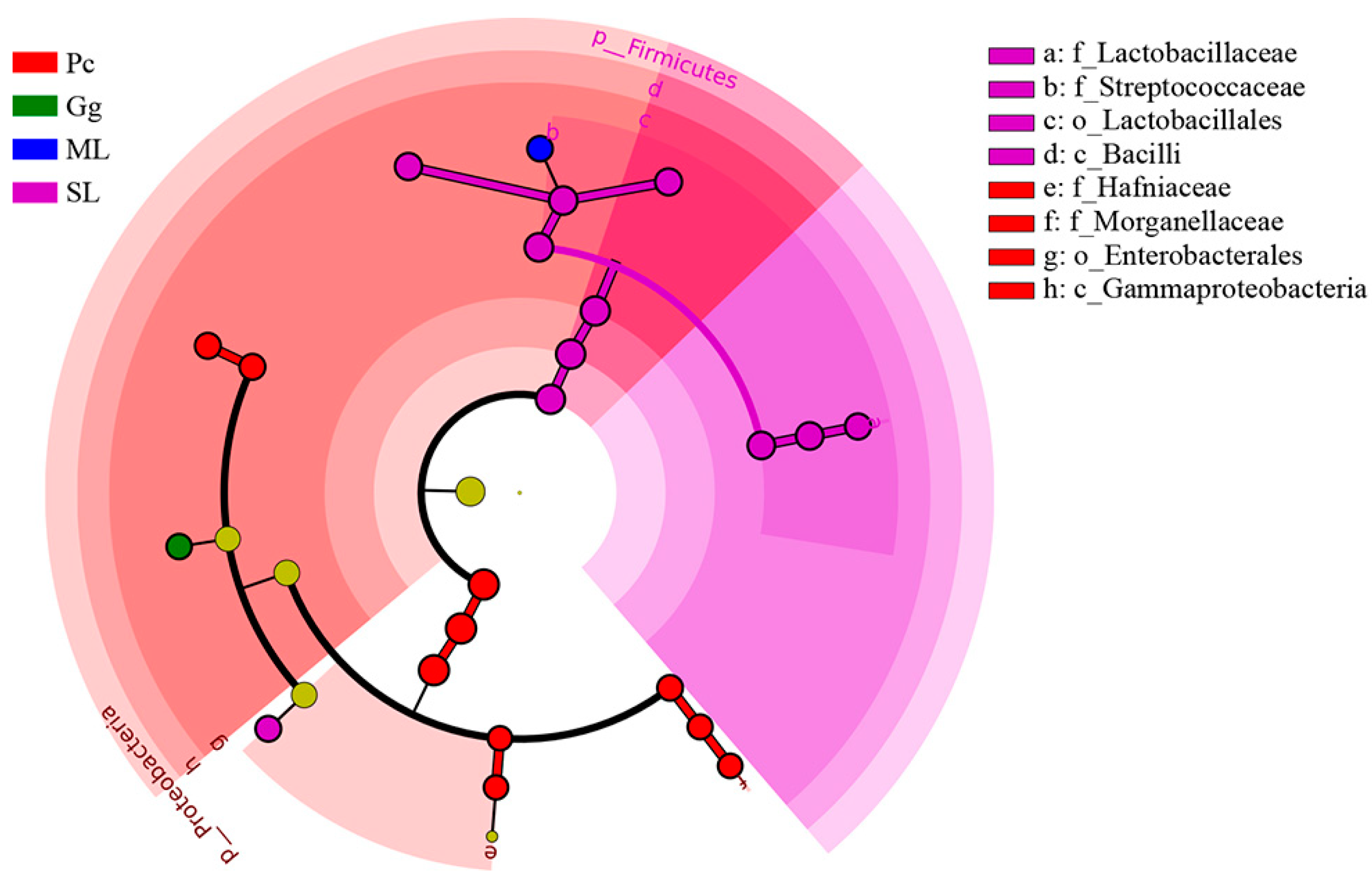
3.2. Metagenomic Analysis
3.3. Metagenomic Analysis
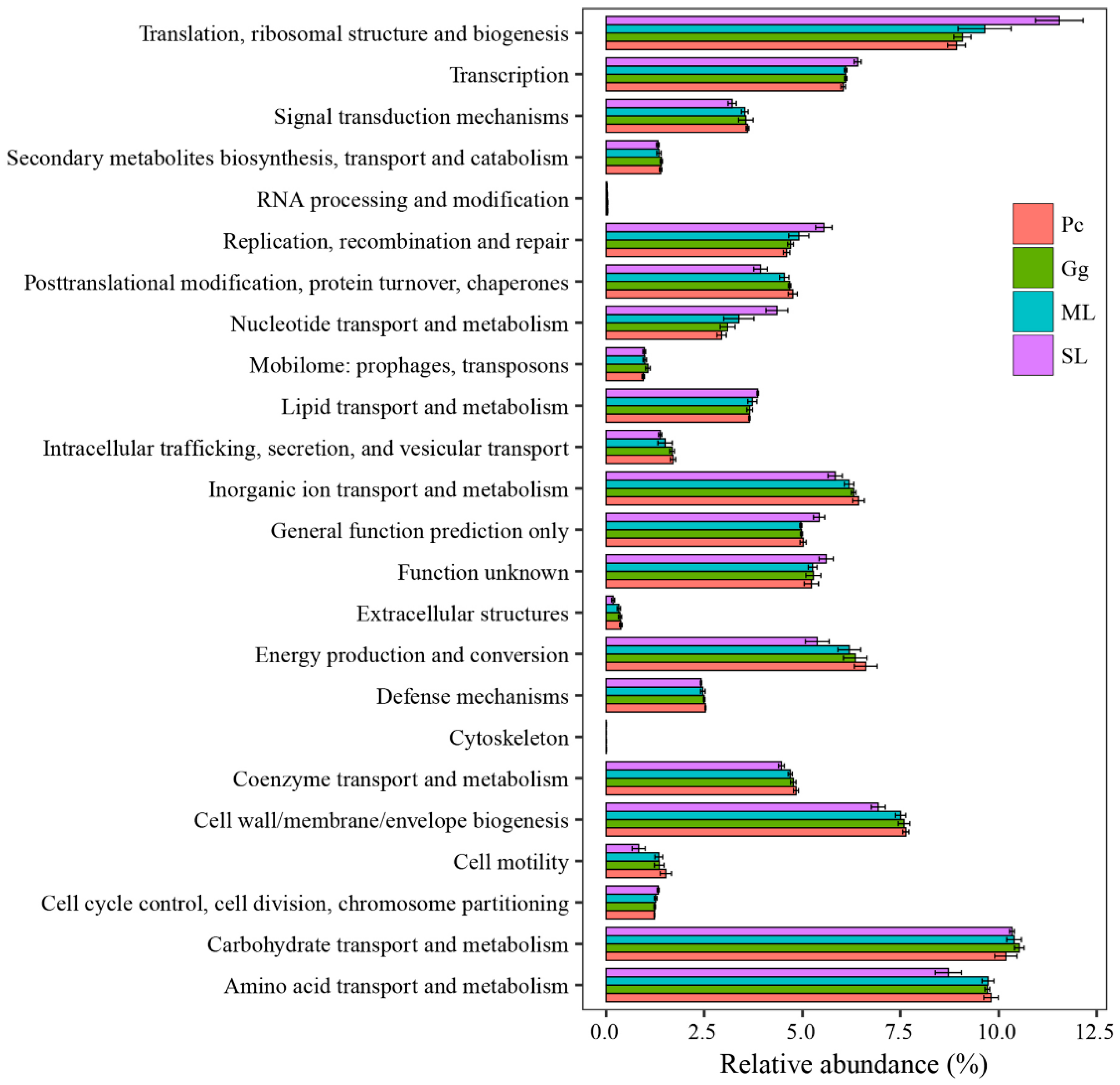
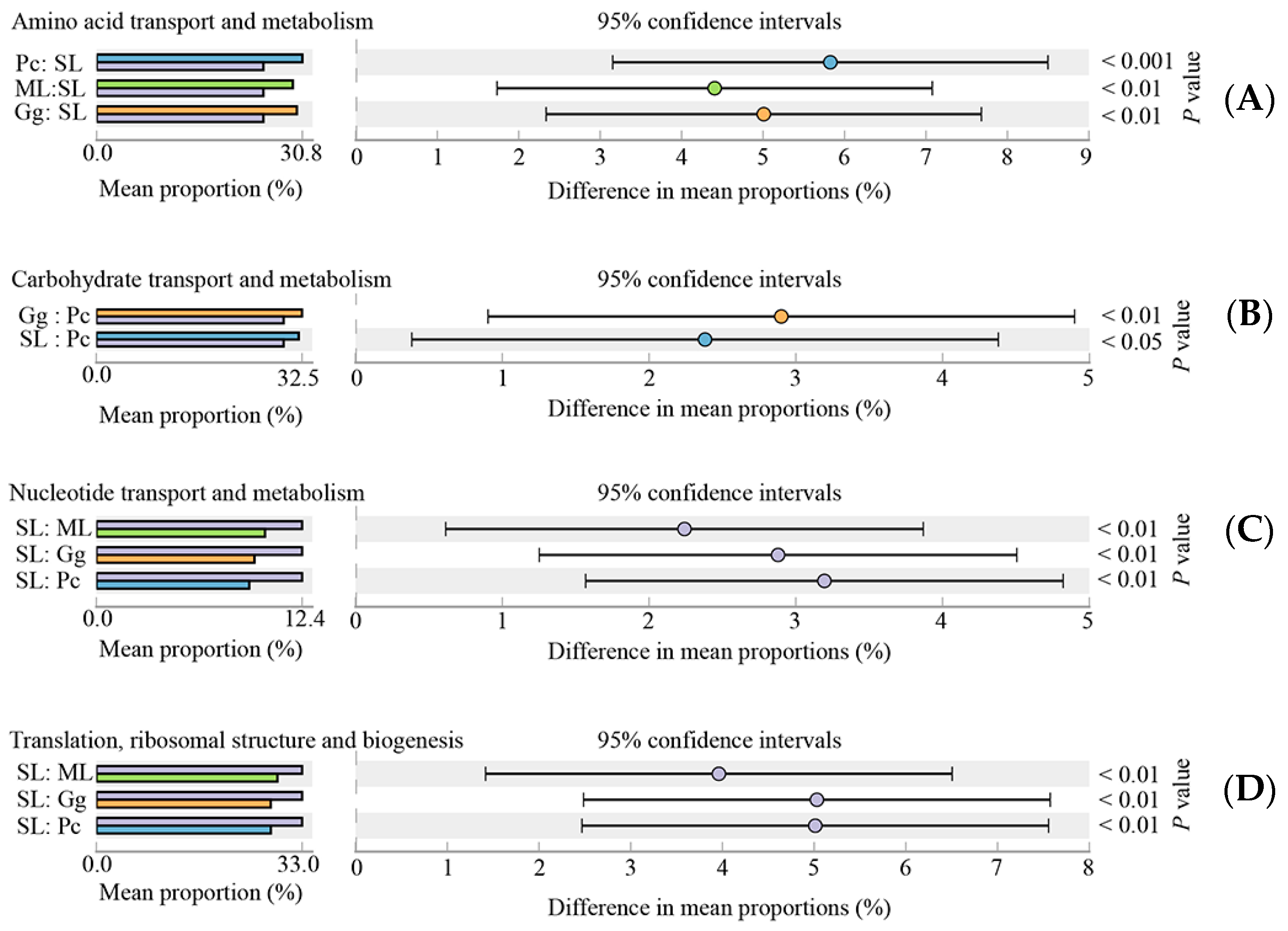
3.4. Functional Annotation of Gut Microbial Community in L. migratoria manilensis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, E.L. Prospects of integrated Locust management for twenty-first century in China. In Proceedings of the Symposium on Integrated Pest Management in China, Kunming, China, 2 December 1996; pp. 1031–1036. [Google Scholar]
- Cao, C.Q.; Huang, M.Y.; Chen, S.Z.; Liu, Y.S. Problems and Counter measures for the Resource Utilization of Locust. J. Anhui Agric. Sci. 2008, 36, 15491–15492, 15499. [Google Scholar] [CrossRef]
- Liu, S.P.; He, F.; Feng, W.J.; Li, Q.X. Research progress on edible insects of Orthoptera. Light Ind. Sci. Technol. 2016, 32, 9–11. [Google Scholar]
- Hou, S.K.; Hu, H.Y.; Liu, J.; Zhang, J. Development and Utilization of a New Type of Renewable Animal Protein Feed Resource-Insect Powder. Feed. Rev. 2017, 12, 35–38. [Google Scholar]
- Egonyu, J.P.; Subramanian, S.; Tanga, C.M.; Dubois, T.; Ekesi, S.; Kelemu, S. Global overview of locusts as food, feed and other uses. Glob. Food Secur. 2021, 31, 100574. [Google Scholar] [CrossRef]
- Qin, J.D.; Guo, F.; Zheng, Z.Y. Food specialization and food utilization of the Oriental migratory locust and the influence of different food plants on its growth and fecundity. Acta Entomol. Sin. 1957, 7, 143–166. [Google Scholar] [CrossRef]
- Chen, Y.L. Major achievements in the study of Locusta migratoria and their management in China. Chin. J. Appl. Entomol. 2000, 37, 50–59. [Google Scholar]
- Liu, Y.S. Captive breeding of Locusta migratoria manilensis (Meyen). Sci. Farming 2006, 02, 36–37. [Google Scholar] [CrossRef]
- Zhao, C. Study on the Feeding Ability on Maize Straw of Locusta migratoria manilensis (Meyen). Master’s Thesis, Shandong Agricultural University, Shandong, China, 2015. [Google Scholar]
- Salama, S.M. Nutrient Composition and Bioactive Components of the Migratory Locust (Locusta migratoria). In African Edible Insects as Alternative Source of Food, Oil, Protein and Bioactive Components; Adam Mariod, A., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 231–239. [Google Scholar]
- Steinhaus, E.A., II. The importance of environmental factors in the insect-microbe ecosystem. Microbiol. Mol. Biol. Rev. 1960, 24, 365–373. [Google Scholar] [CrossRef]
- Fang, J.; Liu, M.; Zhang, S.; Liu, F.; Zhang, Z.; Zhang, Q.; Kong, X. Chemical signal interactions of the bark beetle with fungal symbionts, and host/non-host trees. J. Exp. Bot. 2020, 71, 6084–6091. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, B. Advances in the study of insect gut microbes. Technol. Innov. Appl. 2017, 05, 50. [Google Scholar]
- Mei, C.; Fan, S.; Yang, H. The strategies of isolation of insect gut microorganisms. Acta Microbiol. Sin. 2018, 58, 985–994. [Google Scholar] [CrossRef]
- Yuan, X.Q.; Zhang, X.; Liu, X.Y.; Dong, Y.L.; Yan, Z.Z.; Lv, D.B.; Wang, P.; Li, Y.P. Comparison of Gut Bacterial Communities of Grapholita molesta (Lepidoptera: Tortricidae) Reared on Different Host Plants. Int. J. Mol. Sci. 2021, 22, 6843. [Google Scholar] [CrossRef] [PubMed]
- Cazemier, A.E.; Hackstein, J.H.P.; Op Den Camp, H.J.M.; Rosenberg, J.; van der Drift, C. Bacteria in the Intestinal Tract of Different Species of Arthropods. Microb. Ecol. 1997, 33, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Dillon, R.J.; Dillon, V.M. THE GUT BACTERIA OF INSECTS: Nonpathogenic Interactions. Annu. Rev. Entomol. 2004, 49, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Dillon, R.; Charnley, K. Mutualism between the desert locust Schistocerca gregaria and its gut microbiota. Res. Microbiol. 2002, 153, 503–509. [Google Scholar] [CrossRef]
- Bashir, Z.; Kondapalli, V.K.; Adlakha, N.; Sharma, A.; Bhatnagar, R.K.; Chandel, G.; Yazdani, S.S. Diversity and functional significance of cellulolytic microbes living in termite, pill-bug and stem-borer guts. Sci. Rep. 2013, 3, 2558. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N.A. The gut microbiota of insects-diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef]
- Lee, K.-A.; Kim, S.-H.; Kim, E.-K.; Ha, E.-M.; You, H.; Kim, B.; Kim, M.-J.; Kwon, Y.; Ryu, J.-H.; Lee, W.-J. Bacterial-Derived Uracil as a Modulator of Mucosal Immunity and Gut-Microbe Homeostasis in Drosophila. Cell 2013, 153, 797–811. [Google Scholar] [CrossRef]
- Calcagnile, M.; Tredici, S.M.; Talà, A.; Alifano, P. Bacterial Semiochemicals and Transkingdom Interactions with Insects and Plants. Insects 2019, 10, 441. [Google Scholar] [CrossRef]
- Suenami, S.; Nobu, M.K.; Miyazaki, R. Community analysis of gut microbiota in hornets, the largest eusocial wasps, Vespa mandarinia and V. simillima. Sci. Rep. 2019, 9, 9830. [Google Scholar] [CrossRef]
- Li, F.; Huang, S.S. Isolation and Identification of Lignin-degrading Bacteria from the Gut of Termite and Their Degradation Characteristics. Biotechnol. Bull. 2020, 36, 61–68. [Google Scholar]
- Wang, S.C.; Wang, L.Y.; Fan, X.; Yu, C.; Feng, L.; Yi, L. An Insight into Diversity and Functionalities of Gut Microbiota in Insects. Curr. Microbiol. 2020, 77, 1976–1986. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.Q.; Wang, X.Q.; Feng, W.; Zhou, W.; Xie, H.X.; Wan, Y.J. Comparative analysis of the composition of dominant intestinal microflora in silkworm reared with different forages. Acta Ecol. Sin. 2010, 30, 3875–3882. [Google Scholar]
- Pérez-Cobas, A.E.; Maiques, E.; Angelova, A.; Carrasco, P.; Moya, A.; Latorre, A. Diet shapes the gut microbiota of the omnivorous cockroach Blattella germanica. FEMS Microbiol. Ecol. 2015, 91, 22. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Choi, M.-Y.; Kim, J.-W.; Lee, S.A.; Ahn, J.-H.; Song, J.; Kim, S.-H.; Weon, H.-Y. Effects of diet type, developmental stage, and gut compartment in the gut bacterial communities of two Cerambycidae species (Coleoptera). J. Microbiol. 2017, 55, 21–30. [Google Scholar] [CrossRef]
- Mikaelyan, A.; Dietrich, C.; Köhler, T.; Poulsen, M.; Sillam-Dussès, D.; Brune, A. Diet is the primary determinant of bacterial community structure in the guts of higher termites. Mol. Ecol. 2015, 24, 5284–5295. [Google Scholar] [CrossRef]
- Priya, N.G.; Ojha, A.; Kajla, M.K.; Raj, A.; Rajagopal, R. Host plant induced variation in gut bacteria of Helicoverpa armigera. PLoS ONE 2017, 7, e30768. [Google Scholar] [CrossRef]
- Strano, C.P.; Malacrinò, A.; Campolo, O.; Palmeri, V. Influence of Host Plant on Thaumetopoea pityocampa Gut Bacterial Community. Microb. Ecol. 2018, 75, 487–494. [Google Scholar] [CrossRef]
- Liu, Y.J.; Shen, Z.J.; Yu, J.M.; Li, Z.; Liu, X.X.; Xu, H.L. Comparison of gut bacterial communities and their associations with host diets in four fruit borers. Pest Manag. Sci. 2020, 76, 1353–1362. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Ma, S.J. Occurrence dynamics of Locusta migratoria manilensis (Meyen) in China. Acta Entomol. Sin. 1958, 8, 1–40, 98–99. [Google Scholar] [CrossRef]
- Chen, Y.L. Characteristics, Causes and Ecological Management of Locust Disaster. Bull. Biol. 2000, 35, 1–5. [Google Scholar]
- Wu, W.W. Occurrence characteristics and control strategies of Oriental migratory locust. Hortic. Seed 2003, 23, 366. [Google Scholar]
- Colman, D.R.; Toolson, E.C.; Takacs-Vesbach, C.D. Do diet and taxonomy influence insect gut bacterial communities? Mol. Ecol. 2012, 21, 5124–5137. [Google Scholar] [CrossRef]
- Chandler, J.A.; Lang, J.M.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial communities of diverse Drosophila species: Ecological context of a host-microbe model system. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.L. Diets Affects Hemolymphatic Metabolomics Hemolymph and Gut Microbial Diversity in the Silkworm, Bombyx mori. Master’s Thesis, Soochow University, Suzhou, China, 2017. [Google Scholar]
- Leite-Mondin, M.; DiLegge, M.J.; Manter, D.K.; Weir, T.L.; Silva-Filho, M.C.; Vivanco, J.M. The gut microbiota composition of Trichoplusia ni is altered by diet and may influence its polyphagous behavior. Sci. Rep. 2021, 11, 5786. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.Z. Effect to Growth and Development of Locusta migratoria manilensis (Meyen) Reared on Different Kinds of Diets and Preliminary Identification of Intestinal Bacterium of Locusta migratoria manilensis (Meyen). Master’s Thesis, Shandong Agricultural University, Shandong, China, 2008. [Google Scholar]
- Lee, J.-H.; Lee, K.-A.; Lee, W.-J. Microbiota, Gut Physiology, and Insect Immunity; Academic Press: Cambridge, MA, USA, 2017; Volume 52, pp. 111–138. [Google Scholar] [CrossRef]
- Long, W.M. Molecular Ecological Study on the Intestinal Flora Structure of Grasshoppers. Master’s Thesis, Shanxi University, Shanxi, China, 2009. [Google Scholar]
- Shi, W.B.; Xie, S.X.; Chen, X.Y.; Sun, S.; Zhou, X.; Liu, L.T.; Gao, P.; Kyrpides, N.C.; No, E.-G.; Yuan, J.S. Comparative genomic analysis of the microbiome [corrected] of herbivorous insects reveals eco-environmental adaptations: Biotechnology applications. PLoS Genet. 2013, 9, e1003131. [Google Scholar] [CrossRef]
- Tinker, K.A.; Ottesen, E.A. The Core Gut Microbiome of the American Cockroach, Periplaneta americana, Is Stable and Resilient to Dietary Shifts. Appl. Environ. Microbiol. 2016, 82, 6603–6610. [Google Scholar] [CrossRef]
- Wu, X.L.; Xia, X.F.; Chen, J.H.; GURR, G.M.; You, M.S. Effects of different diets on the diversity of larval gut bacteria of the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Acta Entomol. Sin. 2019, 62, 1172–1185. [Google Scholar] [CrossRef]
- Lv, F.; Liu, Y.S. The identification of dominant intestinal bacteriae of Clanis bilineata tsingtauica Mell larva. Chin. J. Microecol. 2009, 21, 435–439. [Google Scholar] [CrossRef]
- Jin, L.; Wang, H.C.; Wang, H.X.; Zhang, J.G.; Gang, Y.; Jin, H.Y. Bacterial Community in Midguts of Ectropic oblique Larvae by PCR-DGGE and 16SrRNA Gene Library Analysis. Jiangxi Sci. 2013, 31, 759–763, 829. [Google Scholar] [CrossRef]
- Yun, J.-H.; Roh, S.W.; Whon, T.W.; Jung, M.-J.; Kim, M.-S.; Park, D.-S.; Yoon, C.; Nam, Y.-D.; Kim, Y.-J.; Choi, J.-H.; et al. Insect gut bacterial diversity determined by environmental habitat, diet, developmental stage, and phylogeny of host. Appl. Environ. Microbiol. 2014, 80, 5254–5264. [Google Scholar] [CrossRef]
- Mao, Y.X.; Tan, R.R.; Wang, Y.P.; Chen, X.; Wang, H.J.; Huang, D.J.; Gong, Z.M. Analysis of the bacterial diversity in adults of Empoasca (Matsumurasca) onukii based on 16SrDNA sequences. Plant Prot. 2018, 44, 17–23, 48. [Google Scholar] [CrossRef]
- Genta, F.A.; Dillon, R.J.; Terra, W.R.; Ferreira, C. Potential role for gut microbiota in cell wall digestion and glucoside detoxification in Tenebrio molitor larvae. J. Insect Physiol. 2006, 52, 593–601. [Google Scholar] [CrossRef]
- Warnecke, F.; Luginbühl, P.; Ivanova, N.; Ghassemian, M.; Richardson, T.H.; Stege, J.T.; Cayouette, M.; McHardy, A.C.; Djordjevic, G.; Aboushadi, N.; et al. Metagenomic and functional analysis of hindgut microbiota of a wood-feeding higher termite. Nature 2007, 450, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Sun, S.J.; Yang, X.L.; Cheng, J.; Wei, H.S.; Li, Z.; Michaud, J.P.; Liu, X.X. Variability of Gut Microbiota Across the Life Cycle of Grapholita molesta (Lepidoptera: Tortricidae). Front. Microbiol. 2020, 11, 1366. [Google Scholar] [CrossRef] [PubMed]
- Yong, H.-S.; Song, S.-L.; Chua, K.-O.; Lim, P.-E. High Diversity of Bacterial Communities in Developmental Stages of Bactrocera carambolae (Insecta: Tephritidae) Revealed by Illumina MiSeq Sequencing of 16S rRNA Gene. Curr. Microbiol. 2017, 74, 1076–1082. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.F.; Zhang, X.Y.; Chen, Z.S.; Wang, Z.; Lu, Y.Y.; Cheng, D.F. The Divergence in Bacterial Components Associated with Bactrocera dorsalis across Developmental Stages. Front. Microbiol. 2018, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Joynson, R.; Swamy, A.; Bou, P.A.; Chapuis, A.; Ferry, N. Characterization of cellulolytic activity in the gut of the terrestrial land slug Arion ater: Biochemical identification of targets for intensive study. Comp. Biochem. Physiol. Part. B 2014, 177, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X. Biological Nitrogen Fixation and Mechanism of Bite-Degrading Polystyrene Insects. Master’s Thesis, Wuhan University of Technology, Wuhan, China, 2019. [Google Scholar]
- Osimani, A.; Milanović, V.; Cardinali, F.; Garofalo, C.; Clementi, F.; Pasquini, M.; Riolo, P.; Ruschioni, S.; Isidoro, N.; Loreto, N.; et al. The bacterial biota of laboratory-reared edible mealworms (Tenebrio molitor L.): From feed to frass. Int. J. Food Microbiol. 2018, 272, 49–60. [Google Scholar] [CrossRef]
- Wei, D.F.; Wang, X.J.; Yang, J.; Geng, Y.X.; Chen, M. Analysis of the diversity and difference of intestinal bacteria in larvae Hyphantria cunea Drury (Lepidoptera: Arctiidae) on different diets. J. Environ. Entomol. 2017, 39, 515–524. [Google Scholar] [CrossRef]
- He, X.H.; Liu, Y.S.; Zheng, J.F. Effects of different feeding plants on the intestinal bacteria of Locusta migratoria manilensis. Chin. J. Microecol. 2010, 22, 492–497. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Evaluated Parameters | Goosegrass | Maize Leaves | Soybean Leaves | Pakchoi |
|---|---|---|---|---|
| Food intake (g) | 2.715 ± 0.028 a | 2.752 ± 0.029 a | 1.963 ± 0.022 b | 1.448 ± 0.013 c |
| Dung production (g) | 1.625 ± 0.015 a | 1.643 ± 0.015 a | 1.214 ± 0.014 b | 0.973 ± 0.044 c |
| Weight gain (g) | 0.385 ± 0.008 a | 0.392 ± 0.005 a | 0.244 ± 0.004 b | 0.110 ± 0.001 c |
| Survival rate (%) | 76.40 ± 0.51 a | 78.20 ± 0.58 a | 47.60 ± 0.93 b | 27.60 ± 1.36 c |
| Egg production (eggs) | 251.12 ± 12.72 a | 273.35 ± 13.47 a | 85.52 ± 6.45 b | 84.97 ± 11.04 b |
| Developmental Stage | Goosegrass | Maize Leaves | Soybean Leaves | Pakchoi | |
|---|---|---|---|---|---|
| Pre-adult stage (d) | 43.68 ± 0.32 c | 43 ± 0.29 c | 51.63 ± 0.4 b | 55.43 ± 0.41 a | |
| Adult pre-oviposition period (d) | 14.72 ± 0.33 b | 14.26 ± 0.27 b | 16.43 ± 0.34 a | 16.13 ± 0.4 a | |
| Adult stage (d) | Female insects | 43.18 ± 1.7 a | 44.35 ± 1.49 a | 22.29 ± 0.77 b | 21.38 ± 0.75 b |
| Male insects | 41.5 ± 3.14 a | 42.13 ± 2.91 a | 19.4 ± 1.21 b | 19.67 ± 1.2 b | |
| Average generation period (d) | 66.67 ± 0.43 c | 65.53 ± 0.51 d | 70.31 ± 0.45 b | 74.24 ± 0.41 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Liu, Y.; Yin, X. Comparison of Gut Bacterial Communities of Locusta migratoria manilensis (Meyen) Reared on Different Food Plants. Biology 2022, 11, 1347. https://doi.org/10.3390/biology11091347
Wang Q, Liu Y, Yin X. Comparison of Gut Bacterial Communities of Locusta migratoria manilensis (Meyen) Reared on Different Food Plants. Biology. 2022; 11(9):1347. https://doi.org/10.3390/biology11091347
Chicago/Turabian StyleWang, Qian, Yusheng Liu, and Xiangchu Yin. 2022. "Comparison of Gut Bacterial Communities of Locusta migratoria manilensis (Meyen) Reared on Different Food Plants" Biology 11, no. 9: 1347. https://doi.org/10.3390/biology11091347