
Microglia Are Necessary to Regulate Sleep after an Immune Challenge

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rigor
2.2. Animals
2.3. Study Design, Plexxikon Administration, and LPS Injections
2.4. Collection of Physiological Sleep–Wake Parameters
2.5. Tissue Collection and Immunohistochemistry
2.6. Imaging and Microglia Skeletal Analysis
2.7. Statistical Analyses
3. Results
3.1. Microglia Depletion Resulted in a Transient Sleep Increase during the Light Period
3.2. Mice with Depleted Microglia Slept More during the Dark Period Following LPS1 Administration
3.3. Sleep Changes during Microglia Repopulation Were Nominal
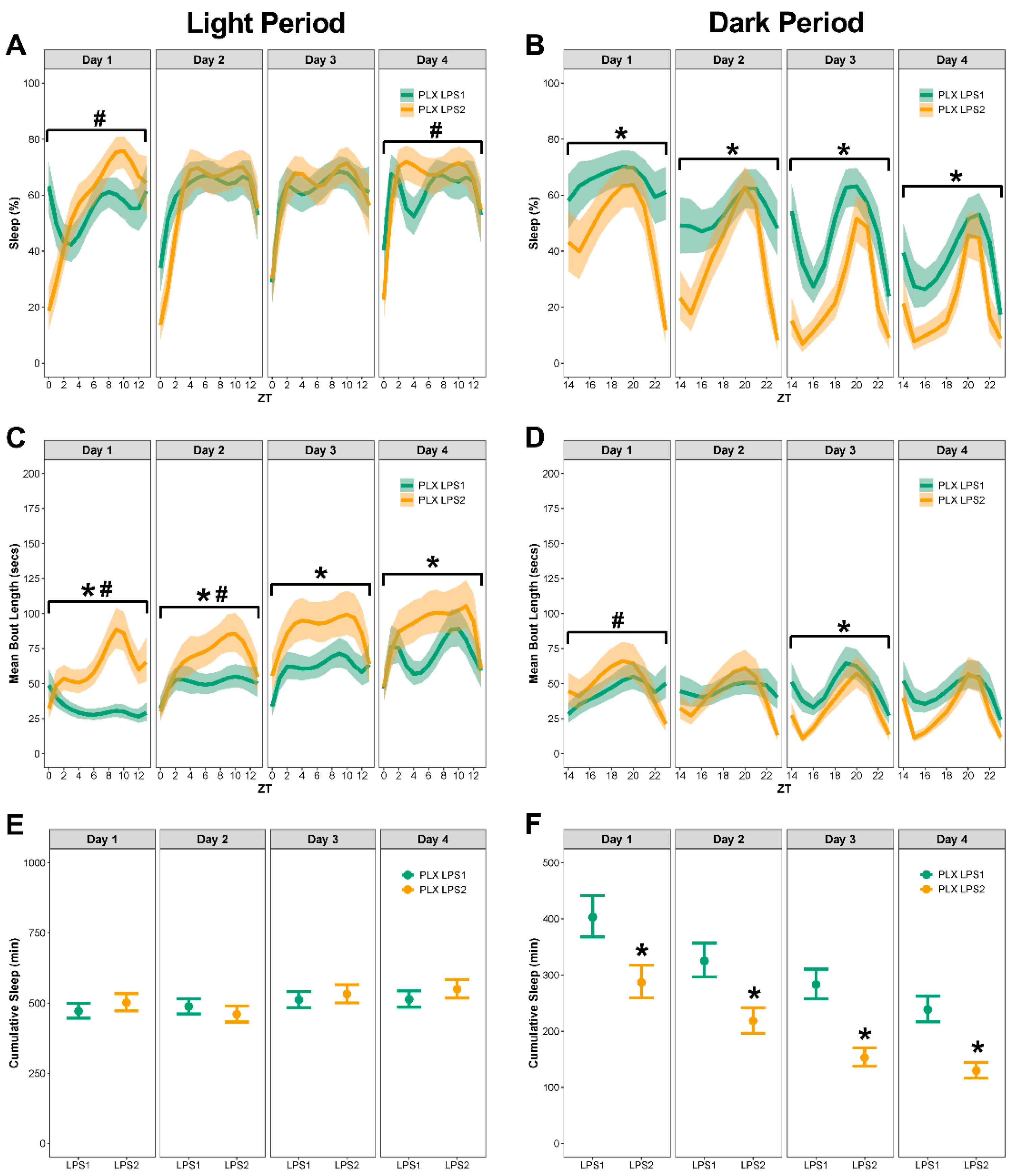
3.4. In Mice with Repopulated Microglia, LPS2 Resulted in Longer Bouts during the Light Period and No Differences in Sleep during the Dark Period Compared to Controls
3.5. In Control Mice, LPS2 Did Not Change the Total Time Spent Sleeping but Resulted in Temporal Pattern Changes in Percent Sleep and Bout Lengths
3.6. In Mice with Repopulated Microglia, LPS2 Resulted in Longer Bouts during the Light Period and Less Sleep during the Dark Period
3.7. Repopulated Microglia Had a Reactive Morphology with Fewer Endpoints per Microglial Cell Compared to Control Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frank, M.G. The Role of Glia in Sleep Regulation and Function. Handb. Exp. Pharmacol. 2019, 253, 83–96. [Google Scholar]
- Garofalo, S.; Picard, K.; Limatola, C.; Nadjar, A.; Pascual, O.; Tremblay, M.E. Role of Glia in the Regulation of Sleep in Health and Disease. Compr. Physiol. 2020, 10, 687–712. [Google Scholar]
- Krueger, J.M. The Role of Cytokines in Sleep Regulation. Curr. Pharm. Design 2008, 14, 3408–3416. [Google Scholar] [CrossRef] [Green Version]
- Krueger, J.M.; Majde, J.A. Cytokines and sleep. Int. Arch. Allergy Imm. 1995, 106, 97–100. [Google Scholar] [CrossRef]
- Krueger, J.M.; Obal, F.; Fang, J.D.; Kubota, T.; Taishi, P. The role of cytokines in physiological sleep regulation. Ann. N. Y. Acad. Sci. 2001, 933, 211–221. [Google Scholar] [CrossRef]
- Opp, M.R. Cytokines and sleep. Sleep Med. Rev. 2005, 9, 355–364. [Google Scholar] [CrossRef]
- Krueger, J.M.; Rector, D.M.; Churchill, L. Sleep and cytokines. Sleep Med. Clin. 2007, 2, 161–169. [Google Scholar] [CrossRef]
- Loane, D.J.; Faden, A.I. Neuroprotection for traumatic brain injury: Translational challenges and emerging therapeutic strategies. Trends Pharmacol. Sci. 2010, 31, 596–604. [Google Scholar] [CrossRef] [Green Version]
- Bilbo, S.; Stevens, B. Microglia: The Brain’s First Responders. Cerebrum 2017, 2017, 14–17. [Google Scholar]
- Catalin, B.; Cupido, A.; Iancau, M.; Albu, C.V.; Kirchhoff, F. Microglia: First responders in the central nervous system. Rom. J. Morphol. Embryol. 2013, 54, 467–472. [Google Scholar]
- Miwa, T.; Furukawa, S.; Nakajima, K.; Furukawa, Y.; Kohsaka, S. Lipopolysaccharide enhances synthesis of brain-derived neurotrophic factor in cultured rat microglia. J. Neurosci. Res. 1997, 50, 1023–1029. [Google Scholar] [CrossRef]
- Nakajima, K.; Kohsaka, S. Microglia: Neuroprotective and neurotrophic cells in the central nervous system. Curr. Drug Targets Cardiovasc. Haematol. Disord. 2004, 4, 65–84. [Google Scholar] [CrossRef] [PubMed]
- Myer, D.J.; Gurkoff, G.G.; Lee, S.M.; Hovda, D.A.; Sofroniew, M.V. Essential protective roles of reactive astrocytes in traumatic brain injury. Brain 2006, 129, 2761–2772. [Google Scholar] [CrossRef] [PubMed]
- Cardona, A.E.; Pioro, E.P.; Sasse, M.E.; Kostenko, V.; Cardona, S.M.; Dijkstra, I.M.; Huang, D.; Kidd, G.; Dombrowski, S.; Dutta, R.; et al. Control of microglial neurotoxicity by the fractalkine receptor. Nat. Neurosci. 2006, 9, 917–924. [Google Scholar] [CrossRef]
- Limatola, C.; Ransohoff, R.M. Modulating neurotoxicity through CX3CL1/CX3CR1 signaling. Front. Cell Neurosci. 2014, 8, 229. [Google Scholar] [CrossRef]
- Stoll, G.; Jander, S.; Schroeter, M. Detrimental and beneficial effects of injury-induced inflammation and cytokine expression in the nervous system. Adv. Exp. Med. Biol. 2002, 513, 87–113. [Google Scholar]
- Dickstein, J.B.; Moldofsky, H. Sleep, cytokines and immune function. Sleep Med. Rev. 1999, 3, 219–228. [Google Scholar] [CrossRef]
- Krueger, J.M.; Majde, J.A.; Rector, D.M. Cytokines in immune function and sleep regulation. Handb. Clin. Neurol. 2011, 98, 229–240. [Google Scholar]
- Imeri, L.; Opp, M.R. How (and why) the immune system makes us sleep. Nat. Rev. Neurosci. 2009, 10, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Krueger, J.M.; Rector, D.M.; Roy, S.; van Dongen, H.P.; Belenky, G.; Panksepp, J. Sleep as a fundamental property of neuronal assemblies. Nat. Rev. Neurosci. 2008, 9, 910–919. [Google Scholar] [CrossRef]
- Dinges, D.F.; Douglas, S.D.; Zaugg, L.; Campbell, D.E.; McMann, J.M.; Whitehouse, W.G.; Orne, E.C.; Kapoor, S.C.; Icaza, E.; Orne, M.T. Leukocytosis and natural killer cell function parallel neurobehavioral fatigue induced by 64 hours of sleep deprivation. J. Clin. Investig. 1994, 93, 1930–1939. [Google Scholar] [CrossRef]
- Frey, D.J.; Fleshner, M.; Wright, K.P., Jr. The effects of 40 hours of total sleep deprivation on inflammatory markers in healthy young adults. Brain Behav. Immun. 2007, 21, 1050–1057. [Google Scholar] [CrossRef]
- Haack, M.; Sanchez, E.; Mullington, J.M. Elevated inflammatory markers in response to prolonged sleep restriction are associated with increased pain experience in healthy volunteers. Sleep 2007, 30, 1145–1152. [Google Scholar] [CrossRef]
- Haack, M.; Pollmacher, T.; Mullington, J.M. Diurnal and sleep-wake dependent variations of soluble TNF- and IL-2 receptors in healthy volunteers. Brain Behav. Immun. 2004, 18, 361–367. [Google Scholar] [CrossRef]
- Irwin, M.R.; Carrillo, C.; Olmstead, R. Sleep loss activates cellular markers of inflammation: Sex differences. Brain Behav. Immun. 2010, 24, 54–57. [Google Scholar] [CrossRef] [Green Version]
- Irwin, M. Effects of sleep and sleep loss on immunity and cytokines. Brain Behav. Immun. 2002, 16, 503–512. [Google Scholar] [CrossRef]
- Irwin, M.R.; Wang, M.; Campomayor, C.O.; Collado-Hidalgo, A.; Cole, S. Sleep deprivation and activation of morning levels of cellular and genomic markers of inflammation. Arch. Intern. Med. 2006, 166, 1756–1762. [Google Scholar] [CrossRef] [Green Version]
- Lekander, M.; Andreasson, A.N.; Kecklund, G.; Ekman, R.; Ingre, M.; Akerstedt, T.; Axelsson, J. Subjective health perception in healthy young men changes in response to experimentally restricted sleep and subsequent recovery sleep. Brain Behav. Immun. 2013, 34, 43–46. [Google Scholar] [CrossRef]
- Meier-Ewert, H.K.; Ridker, P.M.; Rifai, N.; Regan, M.M.; Price, N.J.; Dinges, D.F.; Mullington, J.M. Effect of sleep loss on C-reactive protein, an inflammatory marker of cardiovascular risk. J. Am. Coll. Cardiol. 2004, 43, 678–683. [Google Scholar] [CrossRef] [Green Version]
- Mullington, J.M.; Simpson, N.S.; Meier-Ewert, H.K.; Haack, M. Sleep loss and inflammation. Best Pract. Res. Clin. Endocrinol. Metab. 2010, 24, 775–784. [Google Scholar] [CrossRef] [Green Version]
- Simpson, N.; Dinges, D.F. Sleep and inflammation. Nutr. Rev. 2007, 65, S244–S252. [Google Scholar] [CrossRef] [PubMed]
- Artiushin, G.; Sehgal, A. The Glial Perspective on Sleep and Circadian Rhythms. Annu. Rev. Neurosci. 2020, 43, 119–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, J.C.; Lee, Y.S.; Chang, C.N.; Chuang, H.L.; Ling, E.A.; Lan, C.T. Sleep deprivation inhibits expression of NADPH-d and NOS while activating microglia and astroglia in the rat hippocampus. Cells Tissues Organs 2003, 173, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Bellesi, M.; de Vivo, L.; Chini, M.; Gilli, F.; Tononi, G.; Cirelli, C. Sleep Loss Promotes Astrocytic Phagocytosis and Microglial Activation in Mouse Cerebral Cortex. J. Neurosci. 2017, 37, 5263–5273. [Google Scholar] [CrossRef]
- Hall, S.; Deurveilher, S.; Robertson, G.S.; Semba, K. Homeostatic state of microglia in a rat model of chronic sleep restriction. Sleep 2020, 43, zsaa108. [Google Scholar] [CrossRef]
- Deurveilher, S.; Golovin, T.; Hall, S.; Semba, K. Microglia dynamics in sleep/wake states and in response to sleep loss. Neurochem. Int. 2021, 143, 104944. [Google Scholar] [CrossRef]
- Green, T.R.F.; Ortiz, J.B.; Wonnacott, S.; Williams, R.J.; Rowe, R.K. The bidirectional relationship between sleep and inflammation links traumatic brain injury and alzheimer’s disease. Front. Neurosci. 2020, 14, 894. [Google Scholar] [CrossRef]
- Nadjar, A.; Wigren, H.M.; Tremblay, M.E. Roles of Microglial Phagocytosis and Inflammatory Mediators in the Pathophysiology of Sleep Disorders. Front. Cell Neurosci. 2017, 11, 250. [Google Scholar] [CrossRef] [Green Version]
- Wisor, J.P.; Schmidt, M.A.; Clegern, W.C. Evidence for neuroinflammatory and microglial changes in the cerebral response to sleep loss. Sleep 2011, 34, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Morrow, J.D.; Opp, M.R. Diurnal variation of lipopolysaccharide-induced alterations in sleep and body temperature of interleukin-6-deficient mice. Brain Behav. Immun. 2005, 19, 40–51. [Google Scholar] [CrossRef]
- Ingiosi, A.M.; Opp, M.R. Sleep and immunomodulatory responses to systemic lipopolysaccharide in mice selectively expressing interleukin-1 receptor 1 on neurons or astrocytes. Glia 2016, 64, 780–791. [Google Scholar] [CrossRef]
- Norden, D.M.; Trojanowski, P.J.; Villanueva, E.; Navarro, E.; Godbout, J.P. Sequential activation of microglia and astrocyte cytokine expression precedes increased Iba-1 or GFAP immunoreactivity following systemic immune challenge. Glia 2016, 64, 300–316. [Google Scholar] [CrossRef] [Green Version]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef]
- Rowe, R.K.; Harrison, J.L.; Morrison, H.; Subbian, V.; Murphy, S.M.; Lifshitz, J. Acute post-traumatic sleep may define vulnerability to a second traumatic brain injury in mice. J. Neurotrauma 2018, 36, 1318–1334. [Google Scholar] [CrossRef]
- Harrison, J.L.; Rowe, R.K.; Ellis, T.W.; Yee, N.S.; O’Hara, B.F.; Adelson, P.D.; Lifshitz, J. Resolvins AT-D1 and E1 differentially impact functional outcome, post-traumatic sleep, and microglial activation following diffuse brain injury in the mouse. Brain Behav. Immun. 2015, 47, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Saber, M.; Giordano, K.R.; Hur, Y.; Ortiz, J.B.; Morrison, H.; Godbout, J.P.; Murphy, S.M.; Lifshitz, J.; Rowe, R.K. Acute peripheral inflammation and post-traumatic sleep differ between sexes after experimental diffuse brain injury. Eur. J. Neurosci. 2019, 52, 2791–2814. [Google Scholar] [CrossRef]
- Mang, G.M.; Nicod, J.; Emmenegger, Y.; Donohue, K.D.; O’Hara, B.F.; Franken, P. Evaluation of a piezoelectric system as an alternative to electroencephalogram/ electromyogram recordings in mouse sleep studies. Sleep 2014, 37, 1383–1392. [Google Scholar] [CrossRef] [Green Version]
- Donohue, K.D.; Medonza, D.C.; Crane, E.R.; O’Hara, B.F. Assessment of a non-invasive high-throughput classifier for behaviours associated with sleep and wake in mice. Biomed. Eng. Online 2008, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Green, T.R.F.; Ortiz, J.B.; Harrison, J.L.; Lifshitz, J.; Rowe, R.K. Simultaneous Cryosectioning of Multiple Rodent Brains. J. Vis. Exp. 2018, 139, e58513. [Google Scholar] [CrossRef] [Green Version]
- Hoogland, I.C.; Houbolt, C.; van Westerloo, D.J.; van Gool, W.A.; van de Beek, D. Systemic inflammation and microglial activation: Systematic review of animal experiments. J. Neuroinflammation 2015, 12, 114. [Google Scholar] [CrossRef] [Green Version]
- Faraway, J.J. Extending the Linear Model with R: Generalized Linear, Mixed Effects and Nonparametric Regression Models, 2nd ed.; Chapman and Hall: New York, NY, USA, 2016. [Google Scholar]
- Stroup, W.W. Generalized Linear Mixed Models: Modern Concepts, Methods, and Applications; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.r-project.org/ (accessed on 3 June 2022).
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Mangnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Machler, M.; Bollker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, S.L.P.; Cribari-Neto, F. Beta regression for modelling rates and proportions. J. Appl. Stat. 2004, 31, 799–815. [Google Scholar] [CrossRef]
- Hilbe, J.M. Negative Binomial Regression. In Modeling Count Data; Cambridge University Press: New York, NY, 2014; pp. 126–161. [Google Scholar]
- Kedem, B.; Fokianos, K. Regression Models for Time Series Analysis; John Wiley & Sons: Hoboken, NJ, USA, 2002. [Google Scholar]
- Woolridge, J.M. Part 2, Regression Analysis with Time Series Data; South-Western Cengage Learning: Mason, OH, USA, 2016. [Google Scholar]
- Saber, M.; Murphy, S.M.; Cho, Y.; Lifshitz, J.; Rowe, R.K. Experimental diffuse brain injury and a model of Alzheimer’s disease exhibit disease-specific changes in sleep and incongruous peripheral inflammation. J. Neurosci. Res. 2021, 99, 1136–1160. [Google Scholar] [CrossRef]
- De Boor, C. Calculation of the smoothing spline with weighted roughness measure. Math. Models Methods Appl. Sci. 2001, 11, 33–41. [Google Scholar] [CrossRef]
- Perperoglou, A.; Sauerbrei, W.; Abrahamowicz, M.; Schmid, M. A review of spline function procedures in R. BMC Med. Res. Methodol. 2019, 19, 46. [Google Scholar] [CrossRef] [Green Version]
- Sawilowsky, S.S. New Effect Size Rule of Thumb. J. Mod. Appl. Stat. Methods 2009, 8, 597–599. [Google Scholar] [CrossRef]
- Dunn, O.J. Multiple comparisons among means. J. Am. Stat. Assoc. 1961, 56, 52–64. [Google Scholar] [CrossRef]
- Lenth, R.V.; Buerkner, P.; Herve, M.; Love, J.; Miiguez, F.; Riebl, H.; Singmann, H. Emmeans: Estimated Marginal Means, aka Least-Squares Means, R package version 1.7.3; 2021; Available online: https://www.cran.r-project.org/package=emmeans (accessed on 3 June 2022).
- Prather, A.A.; Vogelzangs, N.; Penninx, B.W. Sleep duration, insomnia, and markers of systemic inflammation: Results from the Netherlands Study of Depression and Anxiety (NESDA). J. Psychiatr. Res. 2015, 60, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Mendoza, J.; Baker, J.H.; Vgontzas, A.N.; Gaines, J.; Liao, D.; Bixler, E.O. Insomnia symptoms with objective short sleep duration are associated with systemic inflammation in adolescents. Brain Behav. Immun. 2017, 61, 110–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peled, N.; Kassirer, M.; Shitrit, D.; Kogan, Y.; Shlomi, D.; Berliner, A.S.; Kramer, M.R. The association of OSA with insulin resistance, inflammation and metabolic syndrome. Respir. Med. 2007, 101, 1696–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kent, B.D.; Ryan, S.; McNicholas, W.T. Obstructive sleep apnea and inflammation: Relationship to cardiovascular co-morbidity. Respir. Physiol. Neurobiol. 2011, 178, 475–481. [Google Scholar] [CrossRef]
- Gozal, D.; Kheirandish-Gozal, L. Cardiovascular morbidity in obstructive sleep apnea: Oxidative stress, inflammation, and much more. Am. J. Respir. Crit. Care Med. 2008, 177, 369–375. [Google Scholar] [CrossRef] [Green Version]
- Rajabally, Y.A.; Shah, R.S. Restless legs syndrome in chronic inflammatory demyelinating polyneuropathy. Muscle Nerve 2010, 42, 252–256. [Google Scholar] [CrossRef]
- Weinstock, L.B.; Walters, A.S.; Paueksakon, P. Restless legs syndrome—Theoretical roles of inflammatory and immune mechanisms. Sleep Med. Rev. 2012, 16, 341–354. [Google Scholar] [CrossRef]
- Irwin, M.R.; Olmstead, R.E.; Ganz, P.A.; Haque, R. Sleep disturbance, inflammation and depression risk in cancer survivors. Brain Behav. Immun. 2013, 30, S58–S67. [Google Scholar] [CrossRef] [Green Version]
- Kwekkeboom, K.L.; Tostrud, L.; Costanzo, E.; Coe, C.L.; Serlin, R.C.; Ward, S.E.; Zhang, Y. The Role of Inflammation in the Pain, Fatigue, and Sleep Disturbance Symptom Cluster in Advanced Cancer. J. Pain Symptom. Manag. 2018, 55, 1286–1295. [Google Scholar] [CrossRef] [Green Version]
- Bower, J.E.; Ganz, P.A.; Irwin, M.R.; Kwan, L.; Breen, E.C.; Cole, S.W. Inflammation and behavioral symptoms after breast cancer treatment: Do fatigue, depression, and sleep disturbance share a common underlying mechanism? J. Clin. Oncol. 2011, 29, 3517–3522. [Google Scholar] [CrossRef] [Green Version]
- Lacourt, T.E.; Kavelaars, A.; Galloway-Pena, J.R.; Sahasrabhojane, P.V.; Shah, N.D.; Futreal, A.; Kontoyiannis, D.P.; Shelburne, S.A.; Heijnen, C.J. Associations of inflammation with symptom burden in patients with acute myeloid leukemia. Psychoneuroendocrinology 2018, 89, 203–208. [Google Scholar] [CrossRef]
- Pearce, S.C.; Stolwyk, R.J.; New, P.W.; Anderson, C. Sleep disturbance and deficits of sustained attention following stroke. J. Clin. Exp. Neuropsychol. 2016, 38, 1–11. [Google Scholar] [CrossRef]
- Kim, J.; Kim, Y.; Yang, K.I.; Kim, D.E.; Kim, S.A. The Relationship Between Sleep Disturbance and Functional Status in Mild Stroke Patients. Ann. Rehabil. Med. 2015, 39, 545–552. [Google Scholar] [CrossRef] [Green Version]
- Baumann, C.R.; Werth, E.; Stocker, R.; Ludwig, S.; Bassetti, C.L. Sleep-wake disturbances 6 months after traumatic brain injury: A prospective study. Brain 2007, 130, 1873–1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaulieu-Bonneau, S.; Morin, C.M. Sleepiness and fatigue following traumatic brain injury. Sleep Med. 2012, 13, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Castriotta, R.J.; Murthy, J.N. Sleep disorders in patients with traumatic brain injury: A review. CNS Drugs 2011, 25, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Kempf, J.; Werth, E.; Kaiser, P.R.; Bassetti, C.L.; Baumann, C.R. Sleep-wake disturbances 3 years after traumatic brain injury. J. Neurol. Neurosurg. Psychiatry 2010, 81, 1402–1405. [Google Scholar] [CrossRef] [Green Version]
- Irwin, M.R.; Vitiello, M.V. Implications of sleep disturbance and inflammation for Alzheimer’s disease dementia. Lancet Neurol. 2019, 18, 296–306. [Google Scholar] [CrossRef]
- Urrestarazu, E.; Iriarte, J. Clinical management of sleep disturbances in Alzheimer’s disease: Current and emerging strategies. Nat. Sci. Sleep 2016, 8, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Vitiello, M.V.; Borson, S. Sleep disturbances in patients with Alzheimer’s disease: Epidemiology, pathophysiology and treatment. CNS Drugs 2001, 15, 777–796. [Google Scholar] [CrossRef]
- Winsky-Sommerer, R.; de Oliveira, P.; Loomis, S.; Wafford, K.; Dijk, D.J.; Gilmour, G. Disturbances of sleep quality, timing and structure and their relationship with other neuropsychiatric symptoms in Alzheimer’s disease and schizophrenia: Insights from studies in patient populations and animal models. Neurosci. Biobehav. Rev. 2019, 97, 112–137. [Google Scholar] [CrossRef]
- Vanderheyden, W.M.; Lim, M.M.; Musiek, E.S.; Gerstner, J.R. Alzheimer’s Disease and Sleep-Wake Disturbances: Amyloid, Astrocytes, and Animal Models. J. Neurosci. 2018, 38, 2901–2910. [Google Scholar] [CrossRef] [Green Version]
- Braley, T.J.; Boudreau, E.A. Sleep Disorders in Multiple Sclerosis. Curr. Neurol. Neurosci. Rep. 2016, 16, 50. [Google Scholar] [CrossRef]
- Veauthier, C.; Gaede, G.; Radbruch, H.; Wernecke, K.D.; Paul, F. Sleep Disorders Reduce Health-Related Quality of Life in Multiple Sclerosis (Nottingham Health Profile Data in Patients with Multiple Sclerosis). Int. J. Mol. Sci. 2015, 16, 16514–16528. [Google Scholar] [CrossRef] [Green Version]
- Ali, T.; Orr, W.C. Sleep disturbances and inflammatory bowel disease. Inflamm. Bowel Dis. 2014, 20, 1986–1995. [Google Scholar] [CrossRef]
- Abad, V.C.; Sarinas, P.S.; Guilleminault, C. Sleep and rheumatologic disorders. Sleep Med. Rev. 2008, 12, 211–228. [Google Scholar] [CrossRef]
- Yin, J.; Valin, K.L.; Dixon, M.L.; Leavenworth, J.W. The Role of Microglia and Macrophages in CNS Homeostasis, Autoimmunity, and Cancer. J. Immunol. Res. 2017, 2017, 5150678. [Google Scholar] [CrossRef] [Green Version]
- Acharya, M.M.; Green, K.N.; Allen, B.D.; Najafi, A.R.; Syage, A.; Minasyan, H.; Le, M.T.; Kawashita, T.; Giedzinski, E.; Parihar, V.K.; et al. Elimination of microglia improves cognitive function following cranial irradiation. Sci. Rep. 2016, 6, 31545. [Google Scholar] [CrossRef]
- Szalay, G.; Martinecz, B.; Lenart, N.; Kornyei, Z.; Orsolits, B.; Judak, L.; Csaszar, E.; Fekete, R.; West, B.L.; Katona, G.; et al. Microglia protect against brain injury and their selective elimination dysregulates neuronal network activity after stroke. Nat. Commun. 2016, 7, 11499. [Google Scholar] [CrossRef] [Green Version]
- Mandell, G.L. Cytokines, phagocytes, and pentoxifylline. J. Cardiovasc. Pharmacol. 1995, 25 (Suppl. S2), S20–S22. [Google Scholar] [CrossRef]
- Kumakura, S.; Yamaguchi, Y.; Murakawa, Y.; Kasukabe, T. Effect of Cytokines and Hyperthermia on Phagocytosis and Phosphatidylserine Externalization: Implication for the Pathophysiology of Hemophagocytic Syndrome. Ann. Clin. Lab. Sci. 2018, 48, 314–322. [Google Scholar]
- Barahona, R.A.; Morabito, S.; Swarup, V.; Green, K.N. Cortical diurnal rhythms remain intact with microglial depletion. Sci. Rep. 2022, 12, 114. [Google Scholar] [CrossRef]
- Corsi, G.; Picard, K.; di Castro, M.A.; Garofalo, S.; Tucci, F.; Chece, G.; Del Percio, C.; Golia, M.T.; Raspa, M.; Scavizzi, F.; et al. Microglia modulate hippocampal synaptic transmission and sleep duration along the light/dark cycle. Glia 2022, 70, 89–105. [Google Scholar] [CrossRef]
- Sominsky, L.; Dangel, T.; Malik, S.; de Luca, S.N.; Singewald, N.; Spencer, S.J. Microglial ablation in rats disrupts the circadian system. FASEB J. 2021, 35, e21195. [Google Scholar] [CrossRef]
- Bedolla, A.; Taranov, A.; Luo, F.; Wang, J.; Turcato, F.; Fugate, E.M.; Greig, N.H.; Lindquist, D.M.; Crone, S.A.; Goto, J.; et al. Diphtheria toxin induced but not CSF1R inhibitor mediated microglia ablation model leads to the loss of CSF/ventricular spaces in vivo that is independent of cytokine upregulation. J. Neuroinflammation 2022, 19, 3. [Google Scholar] [CrossRef] [PubMed]
- Ingiosi, A.M.; Opp, M.R.; Krueger, J.M. Sleep and immune function: Glial contributions and consequences of aging. Curr. Opin. Neurobiol. 2013, 23, 806–811. [Google Scholar] [CrossRef] [Green Version]
- Toth, L.A.; Opp, M.R. Cytokine- and microbially induced sleep responses of interleukin-10 deficient mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R1806–R1814. [Google Scholar] [CrossRef]
- Vichaya, E.G.; Malik, S.; Sominsky, L.; Ford, B.G.; Spencer, S.J.; Dantzer, R. Microglia depletion fails to abrogate inflammation-induced sickness in mice and rats. J. Neuroinflammation 2020, 17, 172. [Google Scholar] [CrossRef]
- Besedovsky, L.; Lange, T.; Born, J. Sleep and immune function. Pflugers Arch. 2012, 463, 121–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opp, M.R.; Krueger, J.M. Sleep and immunity: A growing field with clinical impact. Brain Behav. Immun. 2015, 47, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Biesmans, S.; Meert, T.F.; Bouwknecht, J.A.; Acton, P.D.; Davoodi, N.; de Haes, P.; Kuijlaars, J.; Langlois, X.; Matthews, L.J.; Ver Donck, L.; et al. Systemic immune activation leads to neuroinflammation and sickness behavior in mice. Mediat. Inflamm. 2013, 2013, 271359. [Google Scholar] [CrossRef] [PubMed]
- Granger, J.I.; Ratti, P.L.; Datta, S.C.; Raymond, R.M.; Opp, M.R. Sepsis-induced morbidity in mice: Effects on body temperature, body weight, cage activity, social behavior and cytokines in brain. Psychoneuroendocrinology 2013, 38, 1047–1057. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Henry, C.J.; Dantzer, R.; Johnson, R.W.; Godbout, J.P. Exaggerated sickness behavior and brain proinflammatory cytokine expression in aged mice in response to intracerebroventricular lipopolysaccharide. Neurobiol. Aging 2008, 29, 1744–1753. [Google Scholar] [CrossRef] [Green Version]
- Szentirmai, E.; Krueger, J.M. Sickness behaviour after lipopolysaccharide treatment in ghrelin deficient mice. Brain Behav. Immun. 2014, 36, 200–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neher, J.J.; Cunningham, C. Priming Microglia for Innate Immune Memory in the Brain. Trends Immunol. 2019, 40, 358–374. [Google Scholar] [CrossRef]
- Zhang, X.; Kracht, L.; Lerario, A.M.; Dubbelaar, M.L.; Brouwer, N.; Wesseling, E.M.; Boddeke, E.; Eggen, B.J.L.; Kooistra, S.M. Epigenetic regulation of innate immune memory in microglia. J. Neuroinflamm. 2022, 19, 111. [Google Scholar] [CrossRef]
- Schaafsma, W.; Zhang, X.; van Zomeren, K.C.; Jacobs, S.; Georgieva, P.B.; Wolf, S.A.; Kettenmann, H.; Janova, H.; Saiepour, N.; Hanisch, U.K.; et al. Long-lasting pro-inflammatory suppression of microglia by LPS-preconditioning is mediated by RelB-dependent epigenetic silencing. Brain Behav. Immun. 2015, 48, 205–221. [Google Scholar] [CrossRef]
- Green, T.R.F.; Murphy, S.M.; Ortiz, J.B.; Rowe, R.K. Age-At-Injury Influences the Glial Response to Traumatic Brain Injury in the Cortex of Male Juvenile Rats. Front. Neurol. 2021, 12, 804139. [Google Scholar] [CrossRef]
- Morrison, H.; Young, K.; Qureshi, M.; Rowe, R.K.; Lifshitz, J. Quantitative microglia analyses reveal diverse morphologic responses in the rat cortex after diffuse brain injury. Sci. Rep. 2017, 7, 13211. [Google Scholar] [CrossRef] [Green Version]
- Elmore, M.R.P.; Hohsfield, L.A.; Kramar, E.A.; Soreq, L.; Lee, R.J.; Pham, S.T.; Najafi, A.R.; Spangenberg, E.E.; Wood, M.A.; West, B.L.; et al. Replacement of microglia in the aged brain reverses cognitive, synaptic, and neuronal deficits in mice. Aging Cell 2018, 17, e12832. [Google Scholar] [CrossRef] [Green Version]
- Elmore, M.R.; Lee, R.J.; West, B.L.; Green, K.N. Characterizing newly repopulated microglia in the adult mouse: Impacts on animal behavior, cell morphology, and neuroinflammation. PLoS ONE 2015, 10, e0122912. [Google Scholar] [CrossRef]
- Corona, A.W.; Norden, D.M.; Skendelas, J.P.; Huang, Y.; O’Connor, J.C.; Lawson, M.; Dantzer, R.; Kelley, K.W.; Godbout, J.P. Indoleamine 2,3-dioxygenase inhibition attenuates lipopolysaccharide induced persistent microglial activation and depressive-like complications in fractalkine receptor (CX(3)CR1)-deficient mice. Brain Behav. Immun. 2013, 31, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Lei, F.; Cui, N.; Zhou, C.; Chodosh, J.; Vavvas, D.G.; Paschalis, E.I. CSF1R inhibition by a small-molecule inhibitor is not microglia specific; affecting hematopoiesis and the function of macrophages. Proc. Natl. Acad. Sci. USA 2020, 117, 23336–23338. [Google Scholar] [CrossRef]
- Van Zeller, M.; Sebastiao, A.M.; Valente, C.A. Microglia Depletion from Primary Glial Cultures Enables to Accurately Address the Immune Response of Astrocytes. Biomolecules 2022, 12, 666. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Spangenberg, E.E.; Tang, B.; Holmes, T.C.; Green, K.N.; Xu, X. Microglia Elimination Increases Neural Circuit Connectivity and Activity in Adult Mouse Cortex. J. Neurosci. 2021, 41, 1274–1287. [Google Scholar] [CrossRef] [PubMed]
- Dockman, R.L.; Carpenter, J.M.; Diaz, A.N.; Benbow, R.A.; Filipov, N.M. Sex differences in behavior, response to LPS, and glucose homeostasis in middle-aged mice. Behav. Brain Res. 2022, 418, 113628. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rowe, R.K.; Green, T.R.F.; Giordano, K.R.; Ortiz, J.B.; Murphy, S.M.; Opp, M.R. Microglia Are Necessary to Regulate Sleep after an Immune Challenge. Biology 2022, 11, 1241. https://doi.org/10.3390/biology11081241
Rowe RK, Green TRF, Giordano KR, Ortiz JB, Murphy SM, Opp MR. Microglia Are Necessary to Regulate Sleep after an Immune Challenge. Biology. 2022; 11(8):1241. https://doi.org/10.3390/biology11081241
Chicago/Turabian StyleRowe, Rachel K., Tabitha R. F. Green, Katherine R. Giordano, J. Bryce Ortiz, Sean M. Murphy, and Mark R. Opp. 2022. "Microglia Are Necessary to Regulate Sleep after an Immune Challenge" Biology 11, no. 8: 1241. https://doi.org/10.3390/biology11081241