
The First Comprehensive Biodiversity Study of Culturable Fungal Communities Inhabiting Cryoconite Holes in the Werenskiold Glacier on Spitsbergen (Svalbard Archipelago, Arctic)

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
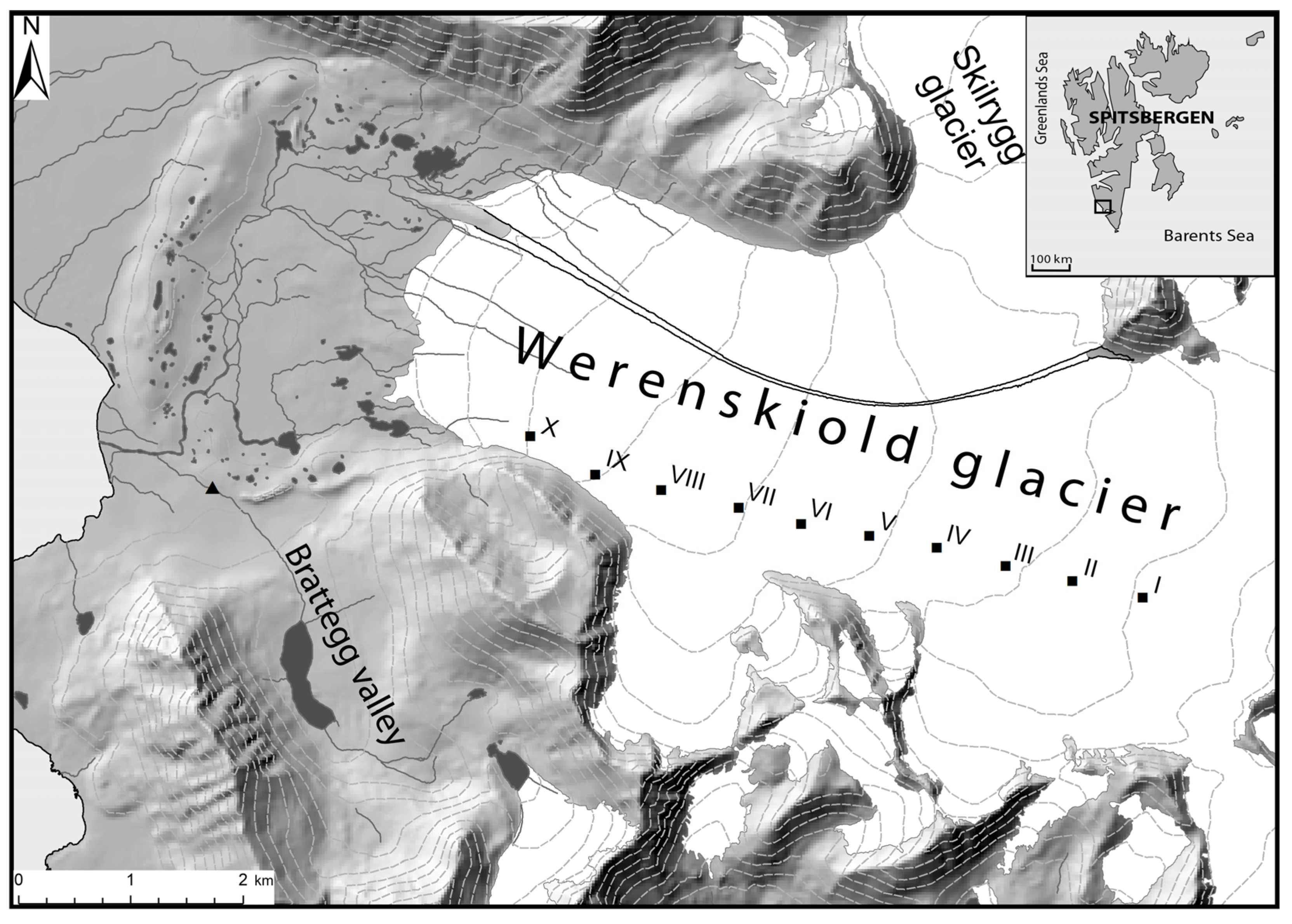
2.1. Study Area
2.2. Mycological Analysis of Samples
2.3. Fungal Identification
2.4. Data Analyses
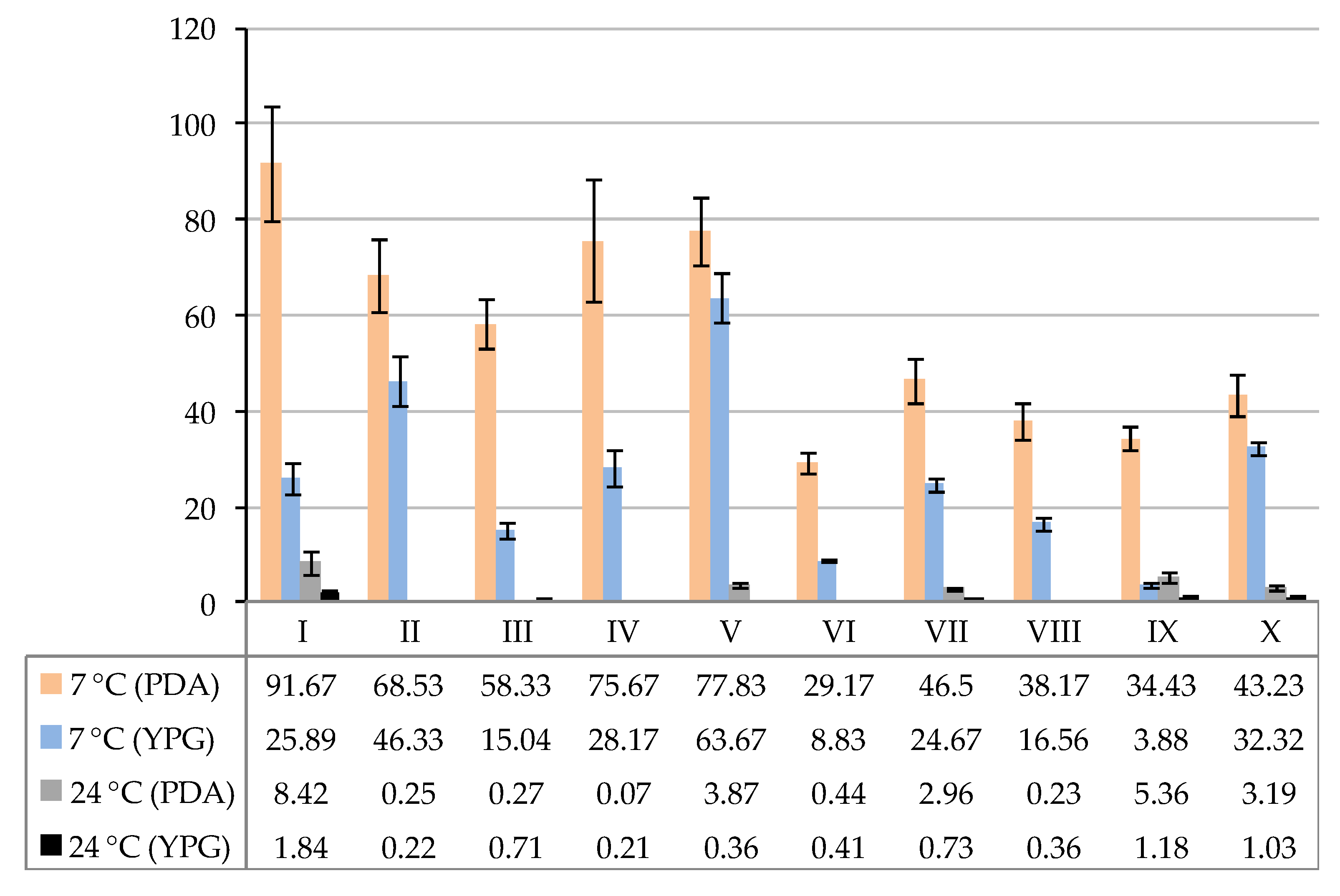
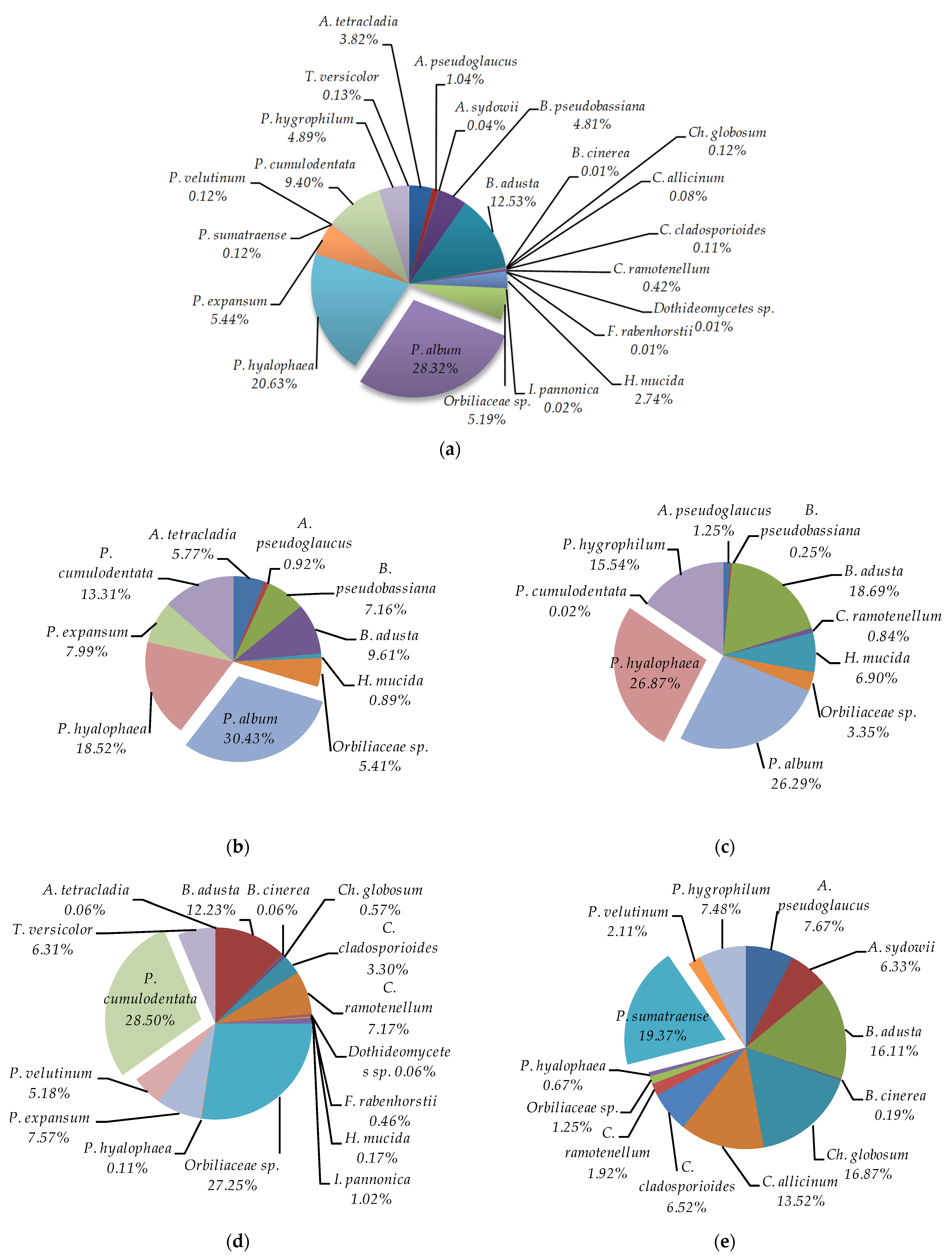
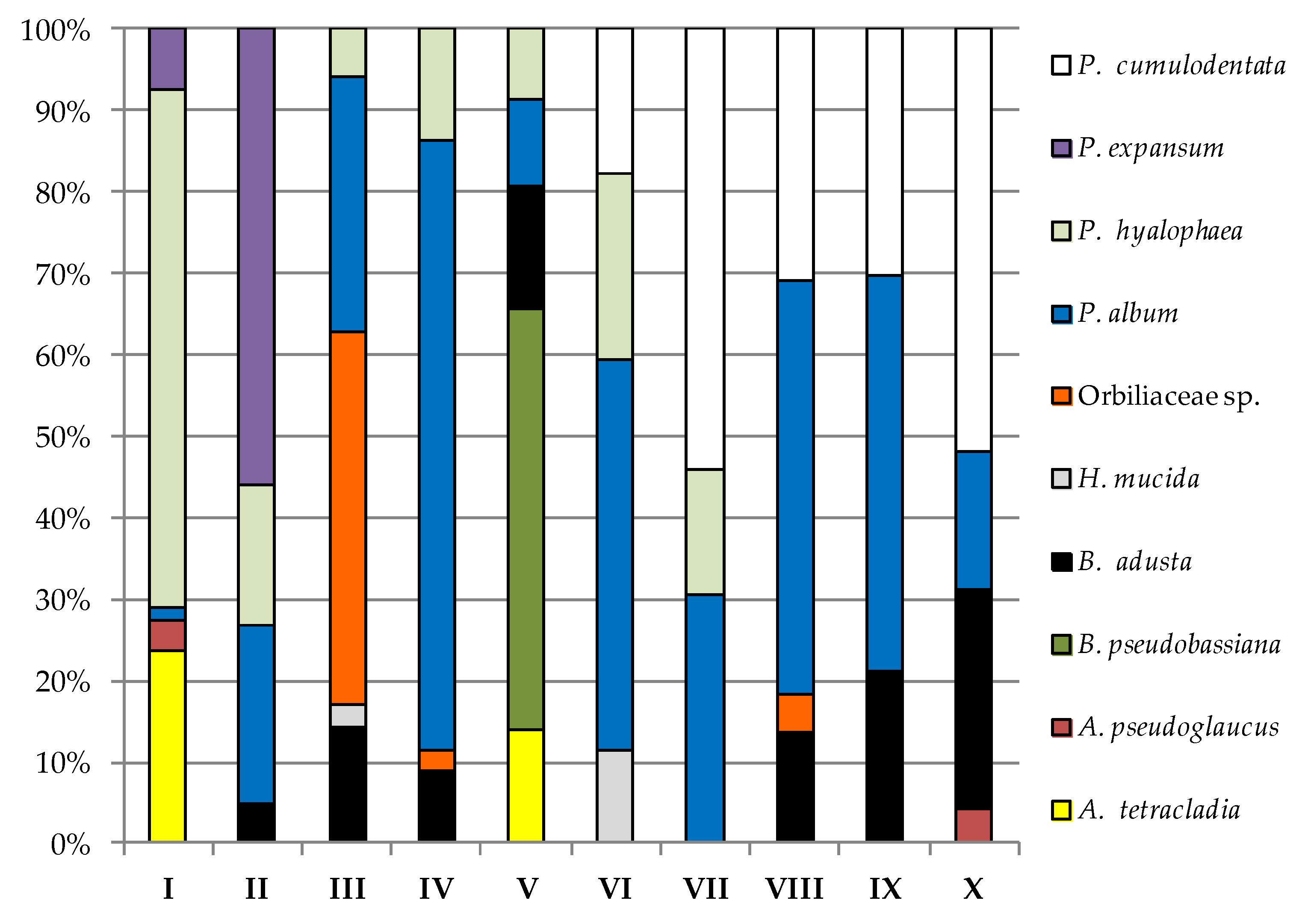
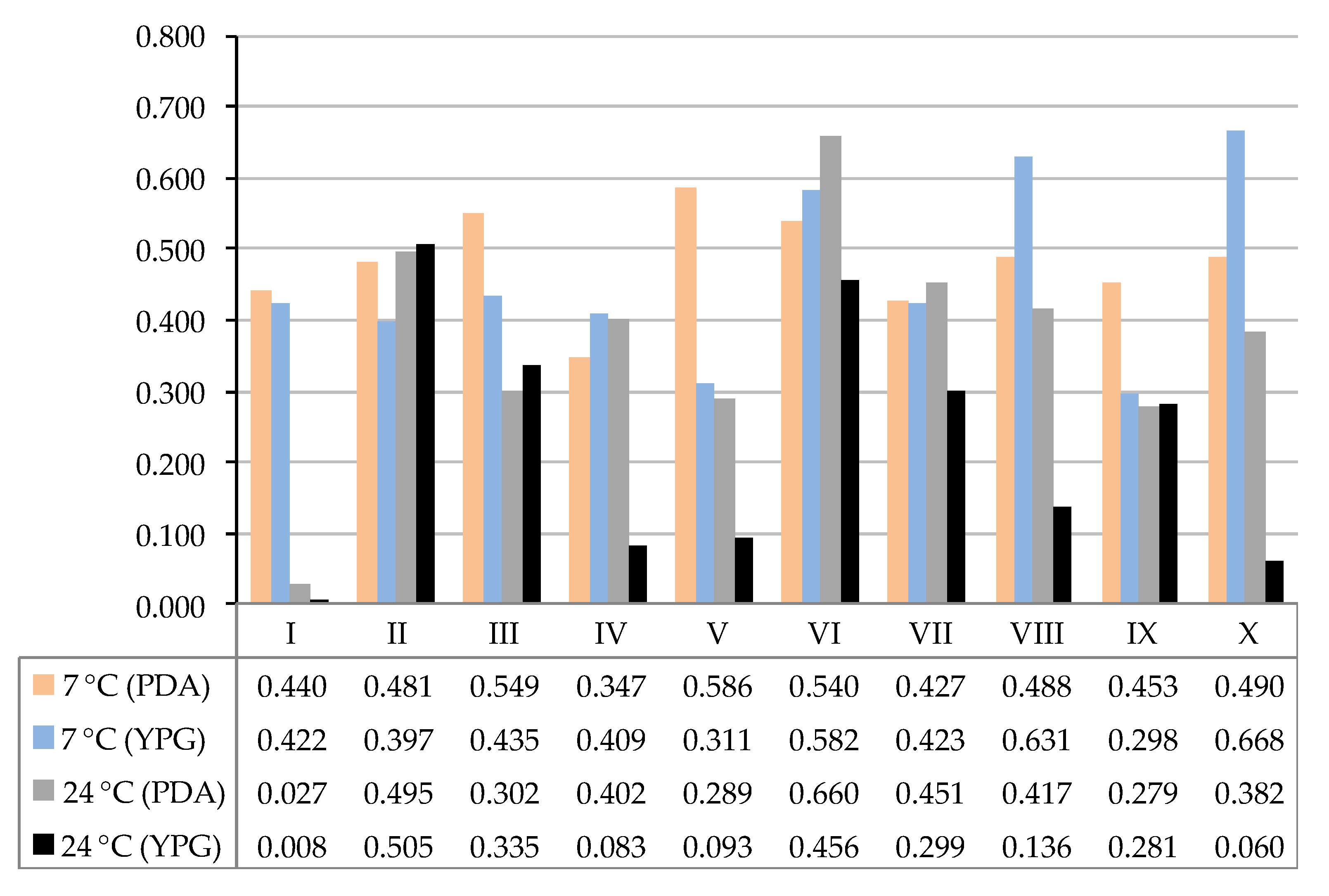
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
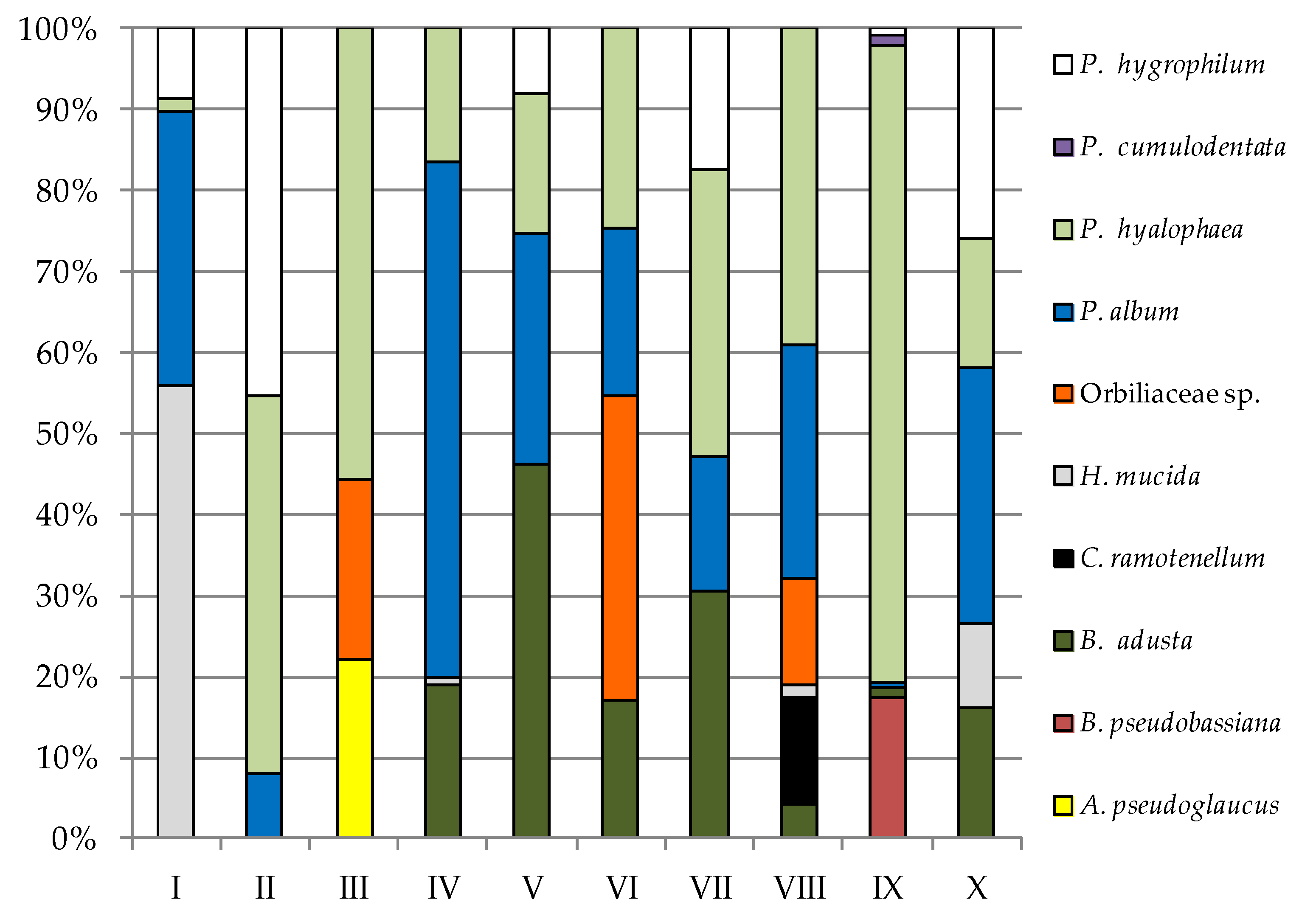
| Fungi | Study Sites | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I 1 | II | III | IV | V | VI | VII | VIII | IX | X | |||||||||||
| PDA | YPG | PDA | YPG | PDA | YPG | PDA | YPG | PDA | YPG | PDA | YPG | PDA | YPG | PDA | YPG | PDA | YPG | PDA | YPG | |
| A. tetracladia | 21.66 | — | — | — | — | — | — | — | 10.83 | — | — | — | — | — | — | — | — | — | — | — |
| A. pseudoglaucus | 3.33 | — | — | — | — | 3.33 | — | — | — | — | — | — | — | — | — | — | — | — | 1.83 | — |
| B. pseudobassiana | — 2 | — | — | — | — | — | — | — | 40.33 | — | — | — | — | — | — | — | — | 0.67 | — | — |
| B. adusta | — | — | 3.33 | — | 8.33 | — | 6.67 | 5.33 | 11.67 | 29.33 | — | 1.50 | — | 7.50 | 5.17 | 0.67 | 7.33 | 0.05 | 11.67 | 5.20 |
| C. ramotenellum | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | 2.22 | — | — | — | — |
| H. mucida | — | 14.44 | — | — | 1.67 | — | — | 0.25 | — | — | 3.33 | — | — | — | — | 0.22 | — | — | — | 3.40 |
| Orbiliaceae sp. | — | — | — | — | 26.67 | 3.34 | 2.00 | — | — | — | — | 3.33 | — | — | 1.83 | 2.22 | — | — | — | — |
| P. album | 1.67 | 8.78 | 15.00 | 3.67 | 18.33 | 0.02 | 56.67 | 17.92 | 8.33 | 18.33 | 14.00 | 1.83 | 14.17 | 4.17 | 19.33 | 4.78 | 16.67 | 0.03 | 7.33 | 10.22 |
| P. hyalophaea | 58.33 | 0.44 | 11.87 | 21.66 | 3.33 | 8.36 | 10.33 | 4.67 | 6.67 | 11.00 | 6.67 | 2.17 | 7.17 | 8.67 | — | 6.44 | — | 3.05 | — | 5.17 |
| P. expansum | 6.67 | — | 38.33 | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| P. cumulodentata | — | — | — | — | — | — | — | — | — | — | 5.17 | — | 25.17 | — | 11.83 | — | 10.43 | 0.05 | 22.40 | — |
| P. hygrophilum | — | 2.22 | — | 21.00 | — | — | — | — | — | 5.00 | — | — | — | 4.33 | — | — | — | 0.03 | — | 8.33 |
| Total species | 5 | 4 | 4 | 3 | 5 | 4 | 4 | 4 | 5 | 4 | 4 | 4 | 3 | 4 | 4 | 6 | 3 | 6 | 4 | 5 |
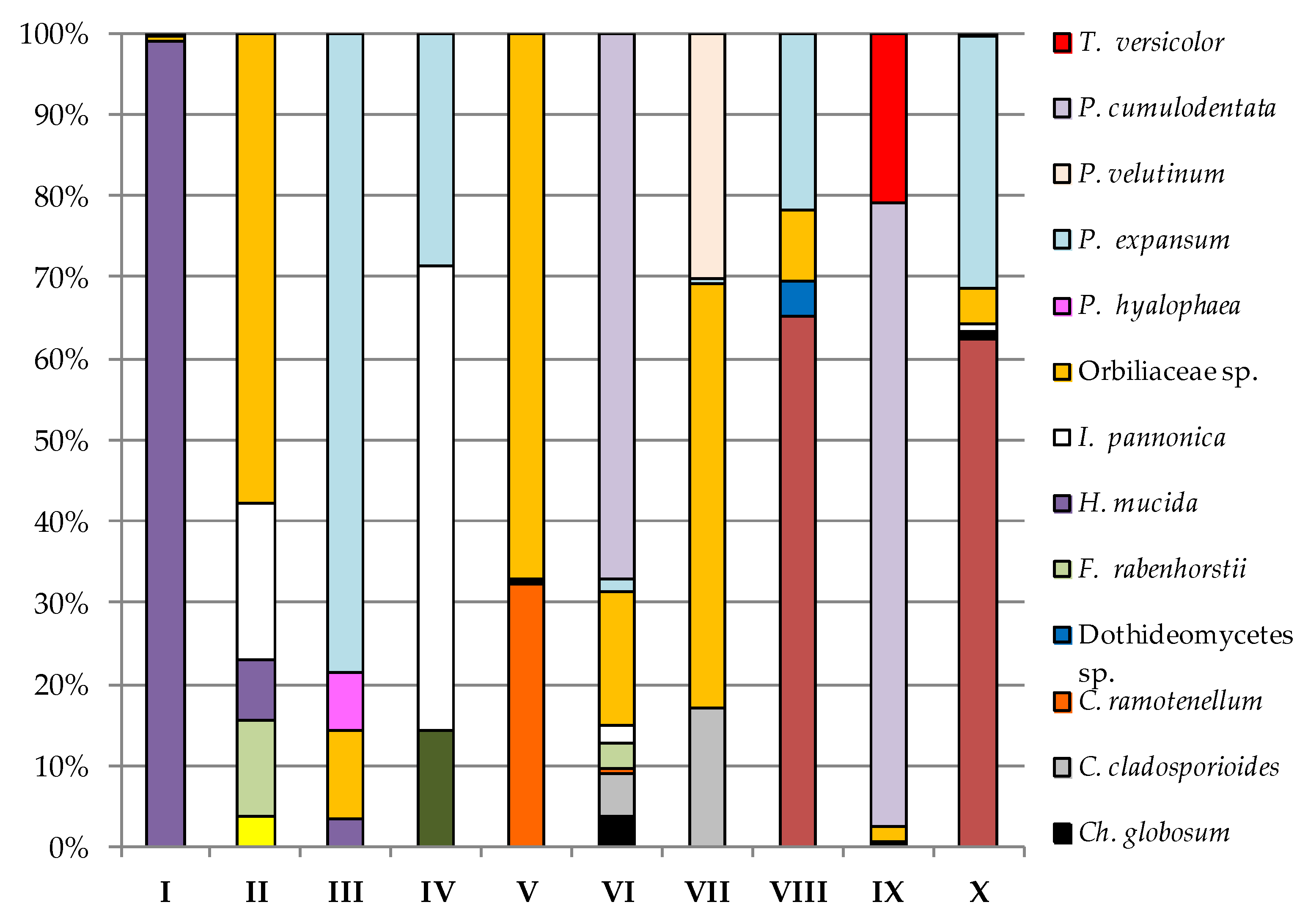
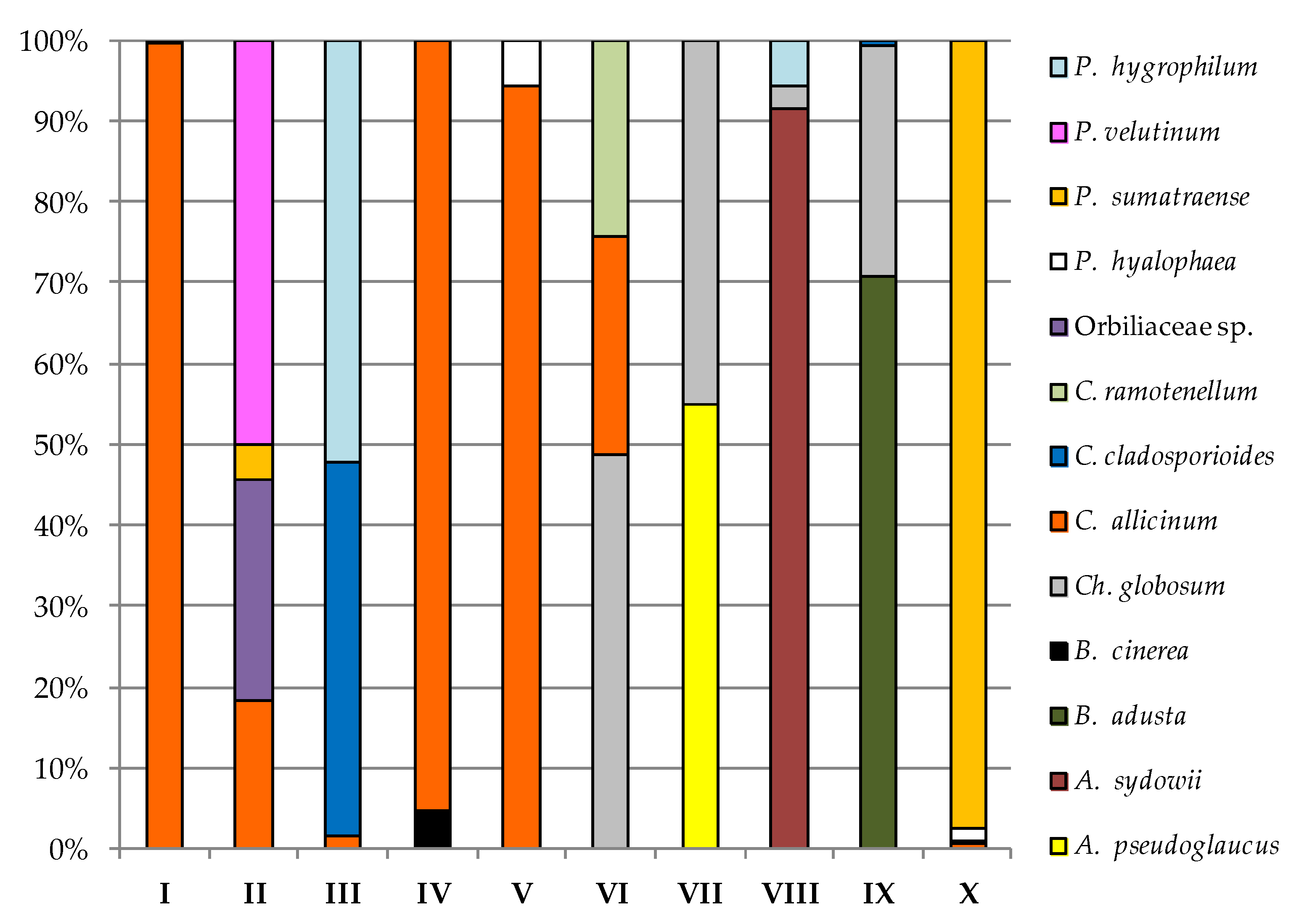
| Fungi | Study Sites | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I 1 | II | III | IV | V | VI | VII | VIII | IX | X | |||||||||||
| PDA | YPG | PDA | YPG | PDA | YPG | PDA | YPG | PDA | YPG | PDA | YPG | PDA | YPG | PDA | YPG | PDA | YPG | PDA | YPG | |
| A. tetracladia | — 2 | — | 0.01 | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| A. pseudoglaucus | — | — | — | — | — | — | — | — | — | — | — | — | — | 0.40 | — | — | — | — | — | — |
| A. sydowii | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | 0.33 | — | — | — | — |
| B. adusta | — | — | — | — | — | — | — | — | — | — | — | — | — | — | 0.15 | — | — | 0.84 | 2.00 | — |
| B. cinerea | — | — | — | — | — | — | 0.01 | 0.01 | — | — | — | — | — | — | — | — | — | — | — | — |
| Ch. globosum | — | — | — | — | — | — | — | — | — | — | 0.05 | 0.20 | — | 0.33 | — | 0.01 | 0.02 | 0.34 | 0.03 | — |
| C. allicinum | — | 1.83 | — | 0.04 | — | 0.01 | — | 0.2 | — | 0.34 | — | 0.11 | — | — | — | — | — | — | 0.005 | |
| C. cladosporioides | — | — | — | — | — | 0.33 | — | — | — | — | 0.07 | — | 0.51 | — | — | — | — | 0.01 | — | — |
| C. ramotenellum | — | — | — | — | — | — | — | — | 1.25 | — | 0.01 | 0.10 | — | — | — | — | — | — | — | — |
| Dothideomycetes sp. | — | — | — | — | — | — | — | — | — | — | — | — | — | — | 0.01 | — | — | — | — | — |
| F. rabenhorstii | — | — | 0.03 | — | — | — | — | — | 0.01 | — | 0.04 | — | — | — | — | — | — | — | — | — |
| H. mucida | 8.33 | — | 0.02 | — | 0.01 | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| I. pannonica | — | — | 0.05 | — | — | — | 0.04 | — | 0.01 | — | 0.03 | — | — | — | — | — | 0.02 | — | 0.03 | — |
| Orbiliaceae sp. | 0.05 | — | 0.15 | 0.06 | 0.03 | — | — | — | 2.60 | — | 0.22 | — | 1.54 | — | 0.02 | — | 0.09 | — | 0.14 | 0.005 |
| P. hyalophaea | — | 0.01 | — | — | 0.02 | — | — | — | — | 0.02 | — | — | — | — | — | — | — | — | — | 0.015 |
| P. expansum | 0.03 | — | — | — | 0.22 | — | 0.02 | — | — | — | 0.02 | — | 0.02 | — | 0.05 | — | — | — | 1.00 | — |
| P. sumatraense | — | — | — | 0.01 | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | 1.00 |
| P. velutinum | — | — | — | 0.11 | — | — | — | — | — | — | — | — | 0.90 | — | — | — | — | — | 0.01 | — |
| P. cumulodentata | — | — | — | — | — | — | — | — | — | — | 0.90 | — | — | — | — | — | 4.11 | — | — | — |
| P. hygrophilum | — | — | — | — | — | 0.37 | — | — | — | — | — | — | — | — | — | 0.02 | — | — | — | — |
| T. versicolor | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | 1.11 | — | — | — |
| Total species | 3 | 2 | 4 | 4 | 4 | 3 | 3 | 2 | 4 | 2 | 7 | 3 | 4 | 2 | 4 | 3 | 5 | 3 | 6 | 4 |
References
- Baccolo, G.; Di Mauro, B.; Massabò, D.; Clemenza, M.; Nastasi, M.; Delmonte, B.; Prata, M.; Prati, P.; Previtali, E.; Maggi, V. Cryoconite as a temporary sink for anthropogenic species stored in glaciers. Sci. Rep. 2017, 7, 9623. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.; Edwards, A.; Takeuchi, N.; Irvine-Fynn, T. Cryoconite: The dark biological secret of the cryosphere. Prog. Phys. Geogr. 2016, 40, 66–111. [Google Scholar] [CrossRef]
- Edwards, A.; Douglas, B.; Anesio, A.M.; Rassner, S.M.; Irvine-Fynn, T.D.L.; Sattler, B.; Griffith, G.W. A distinctive fungal community inhabiting cryoconite holes on glaciers in Svalbard. Fungal Ecol. 2013, 6, 168–176. [Google Scholar] [CrossRef]
- Sommers, P.; Darcy, J.L.; Porazinska, D.L.; Gendron, E.M.S.; Fountain, A.G.; Zamora, F.; Vincent, K.; Cawley, K.M.; Solon, A.J.; Vimercati, L.; et al. Comparison of Microbial Communities in the Sediments and Water Columns of Frozen Cryoconite Holes in the McMurdo Dry Valleys, Antarctica. Front. Microbiol. 2019, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Christner, B.C.; Kvitko, B.H.; Reeve, J.N. Molecular identification of Bacteria and Eukarya inhabiting an Antarctic cryoconite hole. Extremophiles 2003, 7, 177–183. [Google Scholar] [CrossRef]
- Singh, P.; Roy, U.; Tsuji, M. Characterisation of yeast and filamentous fungi from Brøggerbreen glaciers, Svalbard. Polar Record. 2016, 52, 442–449. [Google Scholar] [CrossRef]
- Zawierucha, K.; Trzebny, A.; Buda, J.; Bagshaw, E.; Franzetti, A.; Dabert, M.; Ambrosini, R. Trophic and symbiotic links between obligate-glacier water bears (Tardigrada) and cryoconite microorganisms. PLoS ONE 2022, 17, e0262039. [Google Scholar] [CrossRef]
- Millar, J.; Bagshaw, E.; Edwards, A.; Poniecka, E.; Jungblut, A. Polar Cryoconite Associated Microbiota Is Dominated by Hemispheric Specialist Genera. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Anesio, A.M.; Mindl, B.; Laybourn-Parry, J.; Hodson, A.J.; Sattler, B. Viral dynamics in cryoconite holes on a high Arctic glacier (Svalbard). J. Geophys. Res. 2007, 112, G04S31. [Google Scholar] [CrossRef]
- Säwström, C.; Mumford, P.; Marshall, W.; Hodson, A.; Laybourn-Parry, J. The microbial communities and primary productivity of cryoconite holes in an Arctic glacier (Svalbard 79 °N). Polar Biol. 2002, 25, 591–596. [Google Scholar] [CrossRef]
- Poniecka, A.E.; Bagshaw, E.A.; Sass, H.; Segar, A.; Webster, G.; Williamson, C.; Anesio, A.M.; Tranter, M. Physiological Capabilities of Cryoconite Hole Microorganisms. Front. Microbiol. 2020, 11, 1783. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, Ł.; Jakubowska, N.; Celewicz-Gołdyn, S.; Zawierucha, K. The microorganisms of cryoconite holes (algae, Archaea, bacteria, cyanobacteria, fungi, and Protista): A review. Polar Record. 2016, 52, 176–203. [Google Scholar] [CrossRef]
- Kutty, S.N.; Philip, R. Marine yeasts—A review. Yeast 2008, 25, 465–483. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Singh, S.M. Characterization of Yeast and Filamentous Fungi Isolated from Cryoconite Holes of Svalbard, Arctic. Polar Biol. 2012, 35, 575–583. [Google Scholar] [CrossRef]
- Ludley, K.E.; Robinson, C.H. Decomposer Basidiomycota in Arctic and Antarctic ecosystems. Soil Biol. Biochem. 2008, 40, 11–29. [Google Scholar] [CrossRef]
- Newsham, K.K.; Upson, R.; Read, D.J. Mycorrhizas and dark septate root endophytes in polar regions. Fungal Ecol. 2009, 2, 10–20. [Google Scholar] [CrossRef]
- Wang, M.; Tian, J.; Xiang, M.; Liu, X. Living strategy of cold-adapted fungi with the reference to several representative species. Mycology 2017, 30, 178–188. [Google Scholar] [CrossRef]
- Olsson, P.-A.; Eriksen, B.; Dahlberg, A. Colonisation by arbuscular mycorrhizal and fine endophytic fungi in herbaceous vegetation in Canadian High Arctic. Can. J. Botany 2004, 82, 1547–1556. [Google Scholar] [CrossRef]
- Failor, K.C.; Schmale, D.G.; Vinatzer, B.A.; Monteil, C.L. Ice nucleation active bacteria in precipitation are genetically diverse and nucleate ice by employing different mechanisms. ISME J. 2017, 11, 2740–2753. [Google Scholar] [CrossRef]
- D’Amico, S.; Collins, T.; Marx, J.C.; Feller, G.; Gerday, C. Psychrophilic microorganisms: Challenges for life. EMBO Rep. 2006, 7, 385–389. [Google Scholar] [CrossRef]
- Aislabie, J.M.; Balks, M.R.; Foght, J.M.; Waterhouse, E.J. Hydrocarbonspills on Antarctic soils: Effects and Management. Environ. Sci. Technol. 2004, l38, 1265–1274. [Google Scholar]
- Wynn-Williams, D.W. Ecological aspects of Antarctic microbiology. Adv. Microbial. Ecol 1990, 11, 71–146. [Google Scholar]
- Hamdan, A. Psychrophiles: Ecological significance and potential industrial application. S. Afr. J. Sci. 2018, 114, 1–6. [Google Scholar] [CrossRef]
- Ślubowska, M.A.; Koç, N.; Rasmussen, T.L.; Klitgaard-Kristensen, D. Changes in the flow of Atlantic water into the Arctic Ocean since the last deglaciation: Evidence from the northern Svalbard continental margin, 80 °N. Paleoceanography 2005, 20, PA4014. [Google Scholar] [CrossRef]
- Elven, R.; Murray, D.F.; Razzhivin, V.; Yurtsev, B.A. Checklist of the Panarctic Flora (PAF); Natural History Museum; University of Oslo: Oslo, Norway, 2011. [Google Scholar]
- Mehlum, F.; Gjertz, I. The Birds and Mammals of Svalbard; Norsk Polarinstitutt: Oslo, Norway, 1990; Volume 5. [Google Scholar]
- Lloyd, C.G. Mycological Notes 65. In Mycological Writings; Harvard University: Cambridge, MA, USA, 1921; Volume 6, pp. 1029–1101. [Google Scholar]
- Saccardo, P.A. Nova ascomycetum genera. Grevillea 1875, 4, 21–22. [Google Scholar]
- Chilvers, M.I.; du Toit, L.J. Detection and identification of Botrytis species associated with neck rot, scape blight, and umbel blight of onion: Online. Plant Health Prog. 2006, 7. [Google Scholar] [CrossRef]
- Bensch, K.; Braun, U.; Groenewald, J.Z.; Crous, P.W. The genus Cladosporium. Stud. Mycol. 2012, 72, 1–401. [Google Scholar] [CrossRef]
- Korniłłowicz-Kowalska, T.; Rybczyńska, K. Decolorization of Remazol Brilliant Blue (RBBR) and Poly R-478 dyes by Bjerkandera adusta CCBAS 930. Open Life Sci. 2012, 7, 948–956. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef]
- Kruys, Å.; Huhndorf, S.M.; Miller, A.N. Coprophilous contributions to the phylogeny of Lasiosphaeriaceae and allied taxa within Sordariales (Ascomycota, Fungi). Fungal Divers. 2015, 70, 101–113. [Google Scholar] [CrossRef]
- Liu, X.-Z.; Wang, Q.-M.; Göker, M.; Groenewald, M.; Kachalkin, A.V.; Lumbsch, H.T.; Millanes, A.M.; Wedin, M.; Yurkov, A.M.; Boekhout, T.; et al. Towards an integrated phylogenetic classification of the Tremellomycetes. Stud. Mycol. 2015, 81, 85–147. [Google Scholar] [CrossRef] [PubMed]
- Sogonov, M.V.; Schroers, H.-J.; Gams, W.; Dijksterhuis, J.; Summerbell, R.C. The hyphomycete Teberdinia hygrophila gen. nov., sp. nov. and related anamorphs of Pseudeurotium species. Mycologia 2005, 97, 695–709. [Google Scholar] [CrossRef] [PubMed]
- Volobuev, S.; Okun, M.; Ordynets, A.; Spirin, V. The Phanerochaete sordida group (Polyporales, Basidiomycota) in temperate Eurasia, with a note on Phanerochaete pallida. Mycol. Prog. 2015, 14, 80. [Google Scholar] [CrossRef]
- Soler-Hurtado, M.M.; Sandoval-Sierra, J.V.; Machordom, A.; Diéguez-Uribeondo, J. Aspergillus sydowii and Other Potential Fungal Pathogens in Gorgonian Octocorals of the Ecuadorian Pacific. PLoS ONE 2016, 11, e0165992. [Google Scholar] [CrossRef]
- Wang, X.W.; Lombard, L.; Groenewald, J.Z.; Li, J.; Videira, S.I.R.; Samson, R.A.; Liu, X.Z.; Crous, P.W. Phylogenetic reassessment of the Chaetomium globosum species complex. Persoonia 2016, 36, 83–133. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.J.; Hubka, V.; Frisvad, J.C.; Visagie, C.M.; Houbraken, J.; Meijer, M.; Varga, J.; Demirel, R.; Jurjević, Ž.; Kubátová, A.; et al. Polyphasic taxonomy of Aspergillus section Aspergillus (formerly Eurotium), and its occurrence in indoor environments and food. Stud. Mycol. 2017, 88, 37–135. [Google Scholar] [CrossRef]
- Fiuza, P.O.; Pérez, T.; Gulis, V.; Gusmão, L. Ingoldian fungi of Brazil: Some new records and a review including a checklist and a key. Phytotaxa 2017, 306, 171–200. [Google Scholar] [CrossRef]
- Schoch, C.L.; Crous, P.W.; Groenewald, J.Z.; Boehm, E.W.A.; Burgess, T.I.; de Gruyter, J.; de Hoog, G.S.; Dixon, L.J.; Grube, M.; Gueidan, C.; et al. A class-wide phylogenetic assessment of Dothideomycetes. Stud. Mycol. 2009, 64, 1–15. [Google Scholar] [CrossRef]
- Dyląg, M.; Sawicki, A.; Ogórek, R. Diversity of Species and Susceptibility Phenotypes toward Commercially Available Fungicides of Cultivable Fungi Colonizing Bones of Ursus spelaeus on Display in Niedźwiedzia Cave (Kletno, Poland). Diversity 2019, 11, 224. [Google Scholar] [CrossRef]
- Kanegae, H.; Tomino, N.; Nakamura, Y.; Minakawa, T.; Yaguchi, T.; Izawa, T.; Sano, A.; Nagakawa Itano, E.; Ueda, K. Parengyodontium album Isolated from Cutaneous Lesions of a Pacific White-Sided Dolphin (Lagenorhynchus obliquidens) During Treatment for Paracoccidioidomycosis Ceti. Mycopathologia 2020, 185, 1021–1031. [Google Scholar] [CrossRef]
- Kovač, M.; Gorczak, M.; Wrzosek, M.; Tkaczuk, C.; Pernek, M. Identification of Entomopathogenic Fungi as Naturally Occurring Enemies of the Invasive Oak Lace Bug, Corythucha arcuata (Say) (Hemiptera: Tingidae). Insects 2020, 11, 679. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics in PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Ogórek, R.; Dyląg, M.; Kozak, B. Dark stains on rock surfaces in Driny Cave (Little Carpathian Mountains, Slovakia). Extremophiles 2016, 20, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C.E.; Wiener, W. The Mathematical Theory of Communication; University Illinois Press: Champaign, IL, USA, 1963; Volume 360. [Google Scholar]
- Spellerberg, I.F.; Fedor, P.J. A tribute to Claude-Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the Shannon-Wiener Index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef]
- Grzesiak, J.; Górniak, D.; Świątecki, A.; Aleksandrzak-Piekarczyk, T.; Szatraj, K.; Zdanowski, M.K. Microbial community development on the surface of Hans and Werenskiold Glaciers (Svalbard, Arctic): A comparison. Extremophiles 2015, 19, 885–897. [Google Scholar] [CrossRef] [PubMed]
- Leplat, J.; François, A.; Bousta, F. Parengyodontium Album, a Frequently Reported Fungal Species in the Cultural Heritage Environment. Fungal Biol. Rev. 2020, 34, 126–135. [Google Scholar] [CrossRef]
- Xia, Y.-L.; Sun, J.-H.; Ai, S.-M.; Li, Y.; Du, X.; Sang, P.; Yang, L.-Q.; Fu, Y.-X.; Liu, S.-Q. Insights into the role of electrostatics in temperature adaptation: A comparative study of psychro- philic, mesophilic, and thermophilic subtilisin-like serine proteases. RSC Adv. 2018, 8, 29698–29713. [Google Scholar] [CrossRef]
- Bergero, R.; Girlanda, M.; Varese, G.; Intili, D.; Luppi, A. Psychrooligotrophic fungi from arctic soils of Franz Joseph land. Polar Biol. 1999, 21, 361–368. [Google Scholar] [CrossRef]
- Zucconi, L.; Selbmann, L.; Buzzini, P.; Turchetti, B.; Guglielmin, M.; Frisvad, J.; Onofri, S. Searching for eukaryotic life pre- served in Antarctic permafrost. Polar Biol. 2012, 35, 749–757. [Google Scholar] [CrossRef]
- Wainwright, M.; Wickramasinghe, N.C.; Narlikar, J.V.; Rajaratnam, P. Microorganisms cultured from stratospheric air samples obtained at 41 km. FEMS Microbiol. Lett. 2003, 218, 161–165. [Google Scholar] [CrossRef]
- Liñán, C.; Del Rosal, Y.; Carrasco, F.; Vadillo, I.; Benavente, J.; Ojeda, L. Highlighting the importance of transitional ventilation regimes in the management of Mediterranean show caves (Nerja-Pintada system, southern Spain). Sci. Total Environ. 2018, 631, 1268–1278. [Google Scholar] [CrossRef]
- Kirichuk, N.; Pivkin, M.; Polokhin, O. Fungal assemblages of submarine soils of the eastern Sakhalin shelf. Russ. J. Mar. Biol. 2012, 38, 375–380. [Google Scholar] [CrossRef]
- Yao, Q.; Wang, J.; Zhang, X.; Nong, X.; Xu, X.; Qi, S. Cytotoxic polyketides from the deep-sea-derived fungus Engyodontium album DFFSCS021. Mar. Drugs 2014, 12, 5902–5915. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-Y.; Zhang, Y.; Xu, X.-Y.; Qi, S.-H. Diverse deep-sea fungi from the South China Sea and their antimicrobial activity. Curr. Microbiol. 2013, 67, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Wiese, J.; Wenzel-Storjohann, A.; Malien, S.; Schmaljohann, R.; Imhoff, J.F. Engyodontochones, anti- biotic polyketides from the marine fungus Engyodontium album strain LF069. Chem. Eur. J. 2016, 22, 7452–7462. [Google Scholar] [CrossRef] [PubMed]
- Pindi, P.K. Diversity of fungi at various depths of marine water. Res. Biotechnol. 2012, 3. Available online: https://updatepublishing.com/journal/index.php/rib/article/view/2414 (accessed on 10 May 2021).
- Baral, H.O.; Carter, A. Patinella hyalophaea Sacc–Rediscovered in New Brunswick, Canada. Ascomycete 2013, 5, 91–96. [Google Scholar]
- Matočec, N.; Jukić, N.; Omerović, N.; Kušan, I. Dinaric karst poljes and their importance for mycobiota. In Dinaric Karst Poljes-Nature Conservation and Rural Development; Sackl, P., Ferger, S., Sarajlić, N., Kotrošan, D., Topić, G., Eds.; Ornitološko društvo “Naše ptice”: Sarajevo, Bosnia and Herzegovina, 2019; pp. 27–49. [Google Scholar]
- Held, B.W.; Blanchette, R.A. Deception Island, Antarctica, harbors a diverse assemblage of wood decay fungi. Fungal Biol. 2017, 121, 145–157. [Google Scholar] [CrossRef]
- Pedersen, N.B.; Matthiesen, H.; Blanchette, R.A.; Alfredsen, G.; Held, B.W.; Westergaard-Nielsen, A.; Hollesen, J. Fungal attack on archaeological wooden artefacts in the Arctic—Implications in a changing climate. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Ogaki, M.B.; Vieira, R.; Muniz, M.C.; Zani, C.L.; Alves, T.M.; Junior, P.A.; Murta, S.M.; Barbosa, E.C.; Oliveira, J.G.; Ceravolo, I.P.; et al. Diversity, ecology, and bioprospecting of culturable fungi in lakes impacted by anthropogenic activities in Maritime Antarctica. Extremophiles 2020, 24, 637–655. [Google Scholar] [CrossRef]
- Ogórek, R.; Borzęcka, J.; Kłosińska, K.; Piecuch, A.; Przymencki, M.; Litwiniak, K.; Suchodolski, J. A Culture-Based Study of Micromycetes Isolated from the Urban Nests of Grey Heron (Ardea cinerea) in SW Poland. Animals 2022, 12, 676. [Google Scholar] [CrossRef]
- Gilbertson, R.L.; Lombard, F.F. Wood-rotting Basidiomycetes--Itasca State Park Annotated List. Plant Biol. Commons 1967, 42, 25–31. [Google Scholar]
- Bolshakov, S.Y.; Kalinina, L.B.; Volobuev, S.V.; Rebriev, Y.A.; Shiryaev, A.G.; Khimich, Y.R.; Vlasenko, V.A.; Leostrin, A.V.; Shakhova, N.V.; Vlasenko, A.V.; et al. New Species for Regional Mycobiotas of Russia. 5. Report 2020. Mikol. I Fitopatol. 2020, 54, 404–413. [Google Scholar] [CrossRef]
- Smiri, M.; Kheireddine, A.; Hammami, R.; Rouissi, M.; Espeso, E.A.; Sadfi-Zouaoui, N. An Assessment of the Air Quality in Apple Warehouses: New Records of Aspergillus europaeus, Aspergillus pulverulentus, Penicillium allii and Penicillium sumatraense as Decay Agents. Arch. Microbiol. 2021, 203, 5975–5992. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.S.; Schmidt, O.; Liese, W. Susceptibility of Bamboo to Fungi. In Proceedings of the IXth World Bamboo Congress, Antwerp, Belgium, 10–13 April 2012; pp. 235–245. [Google Scholar]
- Giovannoni, M.; Larini, I.; Scafati, V.; Scortica, A.; Compri, M.; Pontiggia, D.; Zapparoli, G.; Vitulo, N.; Benedetti, M.; Mattei, B. A Novel Penicillium sumatraense Isolate Reveals an Arsenal of Degrading Enzymes Exploitable in Algal Bio-Refinery Processes. Biotechnol. Biofuels 2021, 14, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Cong, B.; Wang, N.; Liu, S.; Liu, F.; Yin, X.; Shen, J. Isolation, Characterization and Transcriptome Analysis of a Novel Antarctic Aspergillus Sydowii Strain MS-19 as a Potential Lignocellulosic Enzyme Source. BMC Microbiol. 2017, 17, 1–14. [Google Scholar] [CrossRef]
- Sonjak, S.; Frisvad, J.C.; Gunde-Cimerman, N. Penicillium Mycobiota in Arctic Subglacial Ice. Microb. Ecol. 2006, 52, 207–216. [Google Scholar] [CrossRef]
- Takenaka, S.; Nakabayashi, R.; Ogawa, C.; Kimura, Y.; Yokota, S.; Doi, M. Characterization of Surface Aspergillus Community Involved in Traditional Fermentation and Ripening of Katsuobushi. Int. J. Food Microbiol. 2020, 327, 108654. [Google Scholar] [CrossRef]
- Wen, R.; Li, X.; Han, G.; Chen, Q.; Kong, B. Fungal Community Succession and Volatile Compound Dynamics in Harbin Dry Sausage during Fermentation. Food Microbiol. 2021, 99, 103764. [Google Scholar] [CrossRef]
- Mouhamadou, B.; Sage, L.; Périgon, S.; Séguin, V.; Bouchart, V.; Legendre, P.; Caillat, M.; Yamouni, H.; Garon, D. Molecular Screening of Xerophilic Aspergillus Strains Producing Mycophenolic Acid. Fungal Biol. 2017, 121, 103–111. [Google Scholar] [CrossRef]
- Zhang, C.; Tao, Y.; Li, S.; Ke, T.; Wang, P.; Wei, S.; Chen, L. Bioremediation of Cadmium-Trichlorfon Co-Contaminated Soil by Indian Mustard (Brassica Juncea) Associated with the Trichlorfon-Degrading Microbe Aspergillus sydowii: Related Physiological Responses and Soil Enzyme Activities. Ecotoxicol. Environ. Saf. 2020, 188, 109756. [Google Scholar] [CrossRef]
- Tian, J.; Dong, Q.; Yu, C.; Zhao, R.; Wang, J.; Chen, L. Biodegradation of the Organophosphate Trichlorfon and Its Major Degradation Products by a Novel Aspergillus sydowii PA F-2. J. Agric. Food Chem. 2016, 64, 4280–4287. [Google Scholar] [CrossRef] [PubMed]
- Willian, G.A. Biodegradation of Chlorpyrifos by Whole Cells of Marine-Derived Fungi Aspergillus sydowii and Trichoderma sp. J. Microb. Biochem. Technol. 2015, 7. [Google Scholar] [CrossRef]
- Niu, S.; Yang, L.; Chen, T.; Hong, B.; Pei, S.; Shao, Z.; Zhang, G. New Monoterpenoids and Polyketides from the deep-sea sediment-derived fungus Aspergillus sydowii MCCC 3A00324. Mar. Drugs 2020, 324, 561. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, N.; Wei, Y.J.; Li, X.; Ge, H.M.; Jiao, R.H.; Tan, R.X. Citrofulvicin, an Antiosteoporotic Polyketide from Penicillium velutinum. Org. Lett. 2018, 20, 3741–3744. [Google Scholar] [CrossRef]
- Chen, Y.; Wei, Y.J.; Jiang, N.; Ge, H.M.; Jiao, R.H.; Cheng, X.; Tan, R.X. Spirocitromycetin, a Fungal Polyketide with an Antiosteoporotic Pharmacophore. J. Nat. Prod. 2022, 85, 1442–1447. [Google Scholar] [CrossRef]
- Ghosh, M.; Nanda, G. Purification and Some Properties of a Xylanase from Aspergillus Sydowii MG49. Appl. Environ. Microbiol. 1994, 60, 4620–4623. [Google Scholar] [CrossRef]
- Prata, M.B.; Mussatto, S.I.; Rodrigues, L.R.; Teixeira, J.A. Fructooligosaccharide Production by Penicillium expansum. Biotechnol. Lett. 2010, 32, 837–840. [Google Scholar] [CrossRef]
- Ciegler, A.; Vesonder, R.F.; Jackson, L.K. Production and Biological Activity of Patulin and Citrinin from Penicillium Expansum. Appl. Environ. Microbiol. 1977, 33, 1004–1006. [Google Scholar] [CrossRef]
- Andersen, B.; Smedsgaard, J.; Frisvad, J.C. Penicillium Expansum: Consistent Production of Patulin, Chaetoglobosins, and Other Secondary Metabolites in Culture and Their Natural Occurrence in Fruit Products. J. Agric. Food Chem. 2004, 52, 2421–2428. [Google Scholar] [CrossRef]
- Motai, H.; Ichishima, E.; Yoshida, F. Purification and Properties of Lipase from Torulopsis. Nature 1966, 210, 308–309. [Google Scholar] [CrossRef]
- Fawzi, E.M. Purification and Characterization of the Pectin Lyase and Protease Produced by Penicillium Velutinum Grown on Eichhornia Crassipes under Solid State Fermentation. Ann. Microbiol. 2009, 59, 755–761. [Google Scholar] [CrossRef]
- Janek, T.; Łukaszewicz, M.; Rezanka, T.; Krasowska, A. Isolation and characterization of two new lipopeptide biosurfactants produced by Pseudomonas fluorescens BD5 isolated from water from the Arctic Archipelago of Svalbard. Bioresour. Technol. 2010, 10, 6118–6123. [Google Scholar] [CrossRef] [PubMed]
- Janek, T.; Łukaszewicz, M.; Krasowska, A. Antiadhesive activity of the biosurfactant pseudofactin II secreted by the Arctic bacterium Pseudomonas fluorescens BD5. BMC Microbiol. 2012, 12, 24. [Google Scholar] [CrossRef] [PubMed]
- Janek, T.; Krasowska, A.; Radwańska, A.; Łukaszewicz, M. Lipopeptide Biosurfactant Pseudofactin II Induced Apoptosis of Melanoma A 375 Cells by Specific Interaction with the Plasma Membrane. PLoS ONE 2013, 8, e57991. [Google Scholar] [CrossRef]
- Biniarz, P.; Baranowska, G.; Feder-Kubis, J.; Krasowska, A. The lipopeptides pseudofactin II and surfactin effectively decrease Candida albicans adhesion and hydrophobicity. Antonie Van Leeuwenhoek 2015, 108, 343–353. [Google Scholar] [CrossRef]
- Biniarz, P.; Coutte, F.; Gancel, F.; Łukaszewicz, M. High-throughput optimization of medium components and culture conditions for the efficient production of a lipopeptide pseudofactin by Pseudomonas fluorescens BD5. Microb. Cell Fact. 2018, 17, 121. [Google Scholar] [CrossRef]
- Janek, T.; Czyżnikowska, Ż.; Łukaszewicz, M.; Gałęzowska, J. The effect of Pseudomonas fluorescens biosurfactant pseudofactin II on the conformational changes of bovine serum albumin: Pharmaceutical and biomedical applications. J. Mol. Liq. 2019, 288, 111001. [Google Scholar] [CrossRef]
- Kantarcıoğlu, A.S.; Yücel, A.; Hoog, G.S. Case report. Isolation of Cladosporium cladosporioides from cerebrospinal fluid. Mycoses 2002, 45, 500–503. [Google Scholar]
- Ogórek, R.; Lejman, A.; Pusz, W.; Miłuch, A.; Miodyńska, P. Characteristics and taxonomy of Cladosporium fungi. Mik. Lek. 2012, 19, 80–85. [Google Scholar]
- Tsuji, M.; Uetake, J.; Tanabe, Y. Changes in the fungal community of Austre Brøggerbreen deglaciation area, Ny-Ålesund, Svalbard, High Arctic. Mycoscience 2016, 57, 448–451. [Google Scholar] [CrossRef]
- Segers, F.J.J.; Meijer, M.; Houbraken, J.; Samson, R.A.; Wösten, H.A.B.; Dijksterhuis, J. Xerotolerant Cladosporium sphaerospermum Are Predominant on Indoor Surfaces Compared to Other Cladosporium Species. PLoS ONE 2015, 10, e0145415. [Google Scholar] [CrossRef] [PubMed]
- Andersen, B.; Frisvad, J.C.; Dunn, R.R.; Thrane, U. A Pilot Study on Baseline Fungi and Moisture Indicator Fungi in Danish Homes. J. Fungi 2021, 7, 71. [Google Scholar] [CrossRef]
- Pinzari, F.; Cornish, L.; Jungblut, A.D. Skeleton Bones in Museum Indoor Environments Offer Niches for Fungi and Are Affected by Weathering and Deposition of Secondary Minerals. Environ. Microbiol. 2020, 22, 59–75. [Google Scholar] [CrossRef] [PubMed]
- Cantabella, D.; Teixidó, N.; Segarra, G.; Torres, R.; Casanovas, M.; Dolcet-Sanjuan, R. Rhizosphere Microorganisms Enhance in Vitro Root and Plantlet Development of Pyrus and Prunus Rootstocks. Planta 2021, 253, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cantabella, D.; Dolcet-Sanjuan, R.; Casanovas, M.; Solsona, C.; Torres, R.; Teixidó, N. Inoculation of in Vitro Cultures with Rhizosphere Microorganisms Improve Plant Development and Acclimatization during Immature Embryo Rescue in Nectarine and Pear Breeding Programs. Sci. Hortic. 2020, 273. [Google Scholar] [CrossRef]
- Ding, S.; Li, N.; Cao, M.; Huang, Q.; Chen, G.; Xie, S.; Zhang, J.; Cheng, G.; Li, W. Diversity of Epiphytic Fungi on the Surface of Kyoho Grape Berries during Ripening Process in Summer and Winter at Nanning Region, Guangxi, China. Fungal Biol. 2019, 123, 283–289. [Google Scholar] [CrossRef]
- Celia, M. Pathogen Identification and Control of Sooty Spot Caused by Cladosporium Ramotenellum, Appearing on Fresh Easy Peeler Mandarins from Perú. J. Plant Sci. Phytopathol. 2021, 5, 44–52. [Google Scholar] [CrossRef]
- Ogórek, R.; Suchodolski, J.; Dudek, B. Droppings of Svalbard reindeer (Rangifer tarandus platyrhynchus) as a reservoir of cultivable micromycetes on Spitsbergen (Svalbard Archipelago, Arctic). Pol. Polar Res. 2022; in press. [Google Scholar]
- Arenz, B.E.; Held, B.W.; Jurgens, J.A.; Blanchette, R.A. Fungal colonization of exotic substrates in Antarctica. Fungal Diver. 2011, 49, 13–22. [Google Scholar] [CrossRef]
- Rodríguez-Couto, S. Industrial and environmental applications of white-rot fungi. Mycosphere 2017, 8, 456–466. [Google Scholar] [CrossRef]
- Heinfling, A.; Bergbauer, M.; Szewzyk, U. Biodegradation of Azo and Phthalocyanine Dyes by Trametes versicolor and Bjerkandera adusta. Appl. Microbiol. Biotechnol. 1997, 48, 261–266. [Google Scholar] [CrossRef]
- Gao, T.; Qin, D.; Zuo, S.; Peng, Y.; Xu, J.; Yu, B.; Song, H.; Dong, J. Decolorization and Detoxification of Triphenylmethane Dyes by Isolated Endophytic Fungus, Bjerkandera Adusta SWUSI4 under Non-Nutritive Conditions. Bioresour. Bioprocess. 2020, 7, 53. [Google Scholar] [CrossRef]
- Robinson, T.; Nigam, P.S. Remediation of Textile Dye Waste Water Using a White-Rot Fungus Bjerkandera Adusta through Solid-State Fermentation (SSF). Appl. Biochem. Biotechnol. 2008, 151, 618–628. [Google Scholar] [CrossRef] [PubMed]
- Bouacem, K.; Rekik, H.; Jaouadi, N.Z.; Zenati, B.; Kourdali, S.; El Hattab, M.; Badis, A.; Annane, R.; Bejar, S.; Hacene, H.; et al. Purification and Characterization of Two Novel Peroxidases from the Dye-Decolorizing Fungus Bjerkandera adusta Strain CX-9. Int. J. Biol. Macromol. 2018, 106, 636–646. [Google Scholar] [CrossRef]
- Gomi, N.; Yoshida, S.; Matsumoto, K.; Okudomi, M.; Konno, H.; Hisabori, T.; Sugano, Y. Degradation of the Synthetic Dye Amaranth by the Fungus Bjerkandera adusta Dec 1: Inference of the Degradation Pathway from an Analysis of Decolorized Products. Biodegradation 2011, 22, 1239–1245. [Google Scholar] [CrossRef]
- Heinfling, A.; Martínez, M.J.; Martínez, A.T.; Bergbauer, M.; Szewzyk, U. Transformation of Industrial Dyes by Manganese Peroxidases from Bjerkandera adusta and Pleurotus eryngii in a Manganese-Independent Reaction. Appl. Environ. Microbiol. 1998, 64, 2788–2793. [Google Scholar] [CrossRef]
- Anastasi, A.; Spina, F.; Prigione, V.; Tigini, V.; Giansanti, P.; Varese, G.C. Scale-up of a Bioprocess for Textile Wastewater Treatment Using Bjerkandera adusta. Bioresour. Technol. 2010, 101, 3067–3075. [Google Scholar] [CrossRef]
- Ramsay, J.A.; Nguyen, T. Decoloration of Textile Dyes by Trametes Versicolor and Its Effect on Dye Toxicity. Biotechnol. Lett. 2002, 24, 1757–1761. [Google Scholar] [CrossRef]
- Blánquez, P.; Casas, N.; Font, X.; Gabarrell, X.; Sarrà, M.; Caminal, G.; Vicent, T. Mechanism of Textile Metal Dye Biotransformation by Trametes versicolor. Water Res. 2004, 38, 2166–2172. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, J. Adsorption and Degradation of Synthetic Dyes on the Mycelium of Trametes versicolor. Water Sci. Technol. 1998, 38, 233–238. [Google Scholar] [CrossRef]
- Quintero, J.C.; Lú-Chau, T.A.; Moreira, M.T.; Feijoo, G.; Lema, J.M. Bioremediation of HCH Present in Soil by the White-Rot Fungus Bjerkandera adusta in a Slurry Batch Bioreactor. Int. Biodeterior. Biodegrad. 2007, 60, 319–326. [Google Scholar] [CrossRef]
- Davila-Vazquez, G.; Tinoco, R.; Pickard, M.A.; Vazquez-Duhalt, R. Transformation of Halogenated Pesticides by Versatile Peroxidase from Bjerkandera adusta. Enzyme Microb. Technol. 2005, 36, 223–231. [Google Scholar] [CrossRef]
- Rubilar, O.; Feijoo, G.; Diez, C.; Lu-Chau, T.A.; Moreira, M.T.; Lema, J.M. Biodegradation of Pentachlorophenol in Soil Slurry Cultures by Bjerkandera Adusta and Anthracophyllum discolor. Ind. Eng. Chem. Res. 2007, 46, 6744–6751. [Google Scholar] [CrossRef]
- Marco-Urrea, E.; Pérez-Trujillo, M.; Vicent, T.; Caminal, G. Ability of White-Rot Fungi to Remove Selected Pharmaceuticals and Identification of Degradation Products of Ibuprofen by Trametes versicolor. Chemosphere 2009, 74, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Majcherczyk, A.; Johannes, C.; Hüttermann, A. Oxidation of Polycyclic Aromatic Hydrocarbons (PAH) by Laccase of Trametes versicolor. Enzyme Microb. Technol. 1998, 22, 335–341. [Google Scholar] [CrossRef]
- Han, M.J.; Choi, H.T.; Song, H.G. Degradation of Phenanthrene by Trametes Versicolor and Its Laccase. J. Microbiol. 2004, 42, 94–98. [Google Scholar]
- Bayramoǧlu, G.; Bektaş, S.; Arica, M.Y. Biosorption of Heavy Metal Ions on Immobilized White-Rot Fungus Trametes versicolor. J. Hazard. Mater. 2003, 101, 285–300. [Google Scholar] [CrossRef]
- Mułenko, W.; Majewski, T.; Ruszkiewicz-Michalska, M. A Preliminary Checklist of Micromycetes in Poland; Wstępna lista grzybów mikroskopijnych Polski; W. Szafer Institute of Botany Polish Academy of Sciences: Kraków, Poland, 2008; ISBN 978-83-89648-75-4. (In Polish) [Google Scholar]
- Chmiel, M.A. Checklist of Polish Larger Ascomycetes; Krytyczna lista wielkoowocnikowych grzybów workowych Polski; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2006; ISBN 978-83-89648-46-4. (In Polish) [Google Scholar]
- Yu, Z.; Mo, M.; Zhang, Y.; Zhang, K.Q. Taxonomy of Nematode-Trapping Fungi from Orbiliaceae, Ascomycota. In Nematode-Trapping Fungi Fungal Diversity Research Series; Zhang, K.Q., Hyde, K., Eds.; Springer: Dordrecht, The Netherlands, 2014; p. 23. [Google Scholar]
- Zhang, Y.; Yu, Z.; Baral, H.; Mo, M.; Zhang, K. New Species and Records of Orbilia (Orbiliaceae, Ascomycota) from China. Fungal Divers. 2009, 36, 141–153. [Google Scholar]
- Yang, Y.; Yang, E.; An, Z.; Liu, X. Evolution of Nematode-Trapping Cells of Predatory Fungi of the Orbiliaceae Based on Evidence from RRNA-Encoding DNA and Multiprotein Sequences. Proc. Natl. Acad. Sci. USA 2007, 104, 8379–8384. [Google Scholar] [CrossRef]
- Hujslová, M.; Kubátová, A.; Bukovská, P.; Chudíčková, M.; Kolařík, M. Extremely Acidic Soils are Dominated by Species-Poor and Highly Specific Fungal Communities. Microb. Ecol. 2017, 73, 321–337. [Google Scholar] [CrossRef]
- Pem, D.; Jeewon, R.; Chethana, K.W.T.; Hongsanan, S.; Doilom, M.; Suwannarach, N.; Hyde, K.D. Species Concepts of Dothideomycetes: Classification, Phylogenetic Inconsistencies and Taxonomic Standardization; Springer: Dordrecht, The Netherlands, 2021; p. 109. ISBN 0123456789. [Google Scholar]
- Wijayawardene, N.N.; Crous, P.W.; Kirk, P.M.; Hawksworth, D.L.; Boonmee, S.; Braun, U.; Dai, D.Q.; D’souza, M.J.; Diederich, P.; Dissanayake, A.; et al. Naming and Outline of Dothideomycetes–2014 Including Proposals for the Protection or Suppression of Generic Names. Fungal Divers. 2014, 69, 1–55. [Google Scholar] [CrossRef]
- Dong, W.; Wang, B.; Hyde, K.D.; McKenzie, E.H.C.; Raja, H.A.; Tanaka, K.; Abdel-Wahab, M.A.; Abdel-Aziz, F.A.; Doilom, M.; Phookamsak, R.; et al. Freshwater Dothideomycetes; Springer: Dordrecht, The Netherlands, 2020; p. 105. ISBN 1322502000463. [Google Scholar]
- Shearer, C.A.; Raja, H.A.; Miller, A.N.; Nelson, P.; Tanaka, K.; Hirayama, K.; Marvanová, L.; Hyde, K.D.; Zhang, Y. The Molecular Phylogeny of Freshwater Dothideomycetes. Stud. Mycol. 2009, 64, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Meyling, N.V.; Schmidt, N.M.; Eilenberg, J. Occurrence and diversity of fungal entomopathogens in soils of low and high Arctic Greenland. Polar Biol. 2012, 35, 1439–1445. [Google Scholar] [CrossRef]
- Ozerskaya, S.M.; Kochkina, G.A.; Ivanushkina, N.E.; Knyazeva, E.V.; Gilichinskii, D.A. The structure of micromycete complexes in permafrost and cryopegs of the arctic. Microbiology 2008, 77, 482–489. [Google Scholar] [CrossRef]
- Leung, G. Genetic analysis and substrate utilization of fungal isolates from the standing dead material of the moss Schistidium apocarpum from a High Arctic site. Master’s Thesis, University of Manchester, Manchester, UK, 2011. [Google Scholar]
- Bjorbækmo, M.F.M.; Carlsen, T.; Brysting, A.; Vrålstad, T.; Høiland, K.; Ugland, K.I.; Geml, J.; Schumacher, T.; Kauserud, H. High diversity of root associated fungi in both alpine and arctic Dryas octopetala. BMC Plant Biol. 2010, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- de Garcia, V.; Brizzio, S.; van Broock, M.R. Yeasts from Glacial Ice of Patagonian Andes, Argentina. FEMS Microbiol. Ecol. 2012, 82, 540–555. [Google Scholar] [CrossRef]
- Rosa, L.H.; Ogaki, M.B.; Lirio, J.M.; Vieira, R.; Coria, S.H.; Pinto, O.H.B.; Carvalho-Silva, M.; Convey, P.; Rosa, C.A.; Câmara, P.E.A.S. Fungal Diversity in a Sediment Core from Climate Change Impacted Boeckella Lake, Hope Bay, North-Eastern Antarctic Peninsula Assessed Using Metabarcoding. Extremophiles 2022, 26, 1–10. [Google Scholar] [CrossRef]








| Fungi Isolated from Glacier Samples | Identity with Sequence from GenBank | |||||
|---|---|---|---|---|---|---|
| Isolate Number | Identified Fungi | Phylum | GenBank Accession No. | The Sequence Length (bp) | Identity, % | Accession |
| UWR_219 | Articulospora tetracladia | Ascomycota | MZ045861 | 409 | 99.27 | LC131004.1 |
| UWR_220 | Aspergillus pseudoglaucus | Ascomycota | MZ045862 | 488 | 100.00 | MH630012.1 |
| UWR_221 | Aspergillus sydowii | Ascomycota | MZ045863 | 513 | 100.00 | MN809362.1 |
| UWR_222 | Beauveria pseudobassiana | Ascomycota | MZ045864 | 502 | 100.00 | MT241786.1 |
| UWR_223 | Bjerkandera adusta | Basidiomycota | MZ045865 | 481 | 100.00 | MT133795.1 |
| UWR_224 | Botrytis cinerea | Ascomycota | MZ045866 | 439 | 100.00 | KP900730.1 |
| UWR_225 | Chaetomium globosum | Ascomycota | MZ045867 | 443 | 100.00 | MN453401.1 |
| UWR_226 | Cladosporium allicinum | Ascomycota | MZ045868 | 506 | 100.00 | MK460808.1 |
| UWR_227 | Cladosporium cladosporioides | Ascomycota | MZ045869 | 498 | 100.00 | MK761055.1 |
| UWR_228 | Cladosporium ramotenellum | Ascomycota | MZ045870 | 452 | 100.00 | N636231.1 |
| UWR_229 | Dothideomycetes sp. | Ascomycota | MZ045871 | 377 | 97.62 | KJ508303.1 |
| UWR_230 | Fimetariella rabenhorstii | Ascomycota | MZ045872 | 406 | 100.00 | MN984305.1 |
| UWR_231 | Holwaya mucida | Ascomycota | MZ045873 | 388 | 98.46 | MN749367.1 |
| UWR_232 | Itersonilia pannonica | Basidiomycota | MZ045874 | 525 | 100.00 | KX067837.1 |
| UWR_233 | Orbiliaceae sp. | Ascomycota | MZ045875 | 490 | 100.00 | LN901113.1 |
| UWR_234 | Parengyodontium album | Ascomycota | MZ045876 | 492 | 100.00 | MT279507.1 |
| UWR_235 | Patinella hyalophaea | Ascomycota | MZ045877 | 400 | 99.75 | MN833368.1 |
| UWR_236 | Penicillium expansum | Ascomycota | MZ045878 | 489 | 100.00 | MT218335.1 |
| UWR_237 | Penicillium sumatraense | Ascomycota | MZ045879 | 465 | 99.79 | MH971259.1 |
| UWR_238 | Penicillium velutinum | Ascomycota | MZ045880 | 515 | 100.00 | AF033448.1 |
| UWR_239 | Phanerochaete cumulodentata | Basidiomycota | MZ045881 | 432 | 99.77 | MH971273.1 |
| UWR_240 | Pseudeurotium hygrophilum | Ascomycota | MZ045882 | 448 | 100.00 | MF375774.1 |
| UWR_241 | Trametes versicolor | Basidiomycota | MZ045883 | 363 | 100.00 | MT000476.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borzęcka, J.; Suchodolski, J.; Dudek, B.; Matyaszczyk, L.; Spychała, K.; Ogórek, R. The First Comprehensive Biodiversity Study of Culturable Fungal Communities Inhabiting Cryoconite Holes in the Werenskiold Glacier on Spitsbergen (Svalbard Archipelago, Arctic). Biology 2022, 11, 1224. https://doi.org/10.3390/biology11081224
Borzęcka J, Suchodolski J, Dudek B, Matyaszczyk L, Spychała K, Ogórek R. The First Comprehensive Biodiversity Study of Culturable Fungal Communities Inhabiting Cryoconite Holes in the Werenskiold Glacier on Spitsbergen (Svalbard Archipelago, Arctic). Biology. 2022; 11(8):1224. https://doi.org/10.3390/biology11081224
Chicago/Turabian StyleBorzęcka, Justyna, Jakub Suchodolski, Bartłomiej Dudek, Lena Matyaszczyk, Klaudyna Spychała, and Rafał Ogórek. 2022. "The First Comprehensive Biodiversity Study of Culturable Fungal Communities Inhabiting Cryoconite Holes in the Werenskiold Glacier on Spitsbergen (Svalbard Archipelago, Arctic)" Biology 11, no. 8: 1224. https://doi.org/10.3390/biology11081224