
Confirmation of the Applicability of Skeletochronology and Estimating the Age Structure of Kaloula borealis (Microhylidae: Anura) at Lake Sihwa, South Korea



Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
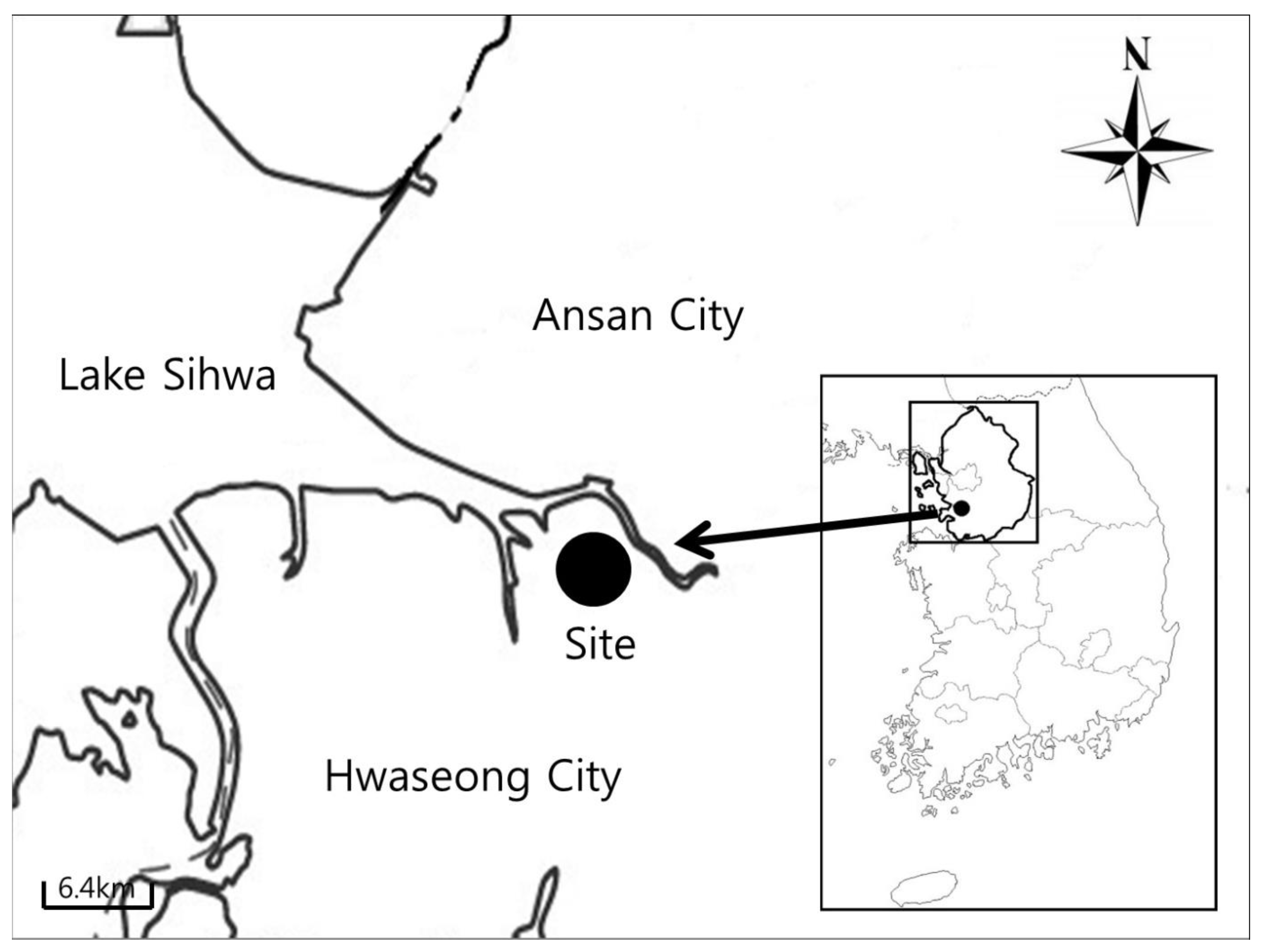
2.1. Study Site
2.2. Sample Collection
2.3. Rearing of Juveniles and the Determination of Reference ML and LAG1 Diameter
2.4. Skeletochronology
2.5. Estimation of the Growth Curve
2.6. Estimation of Sexual Dimorphism
2.7. Statistical Analysis
3. Results
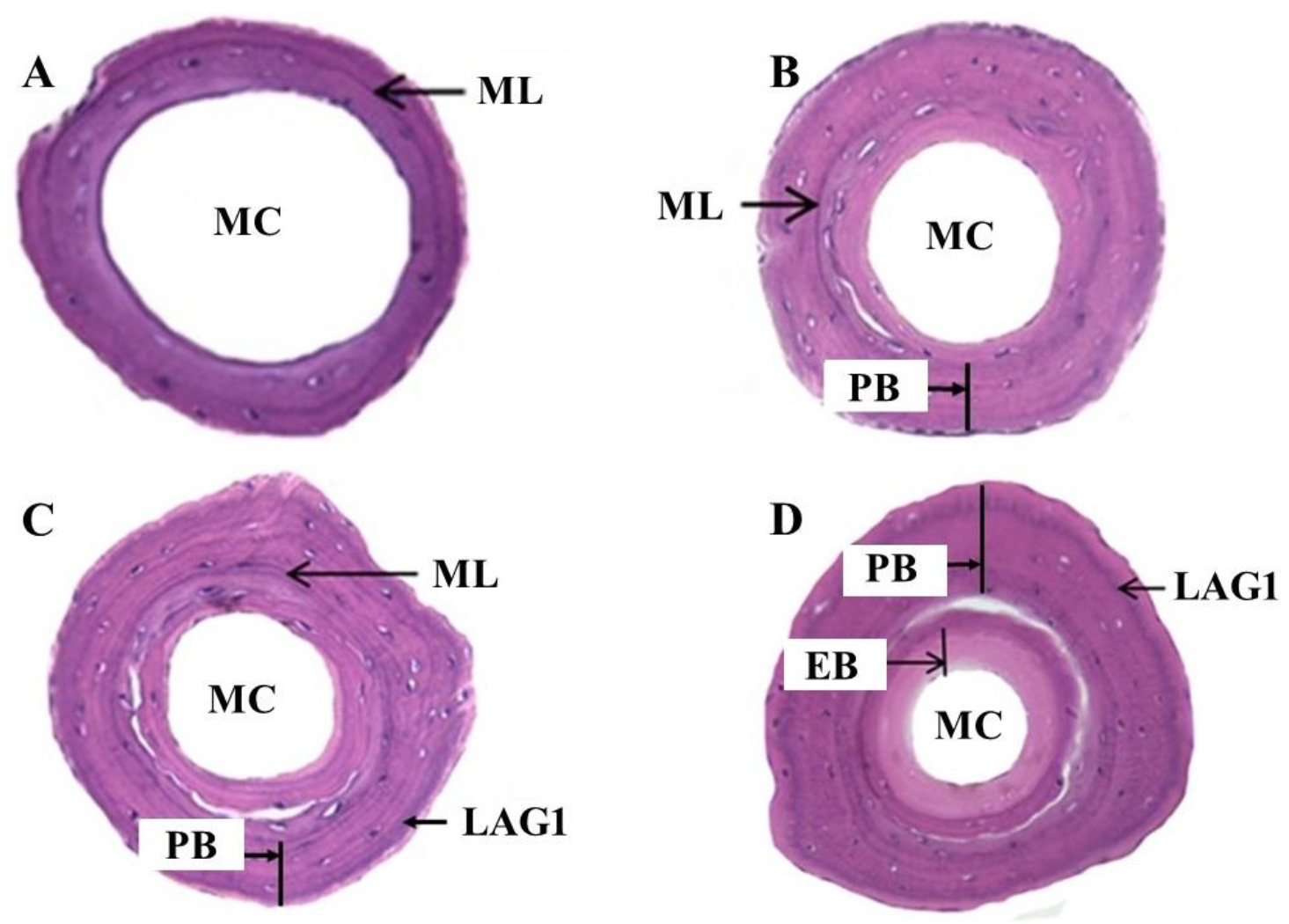
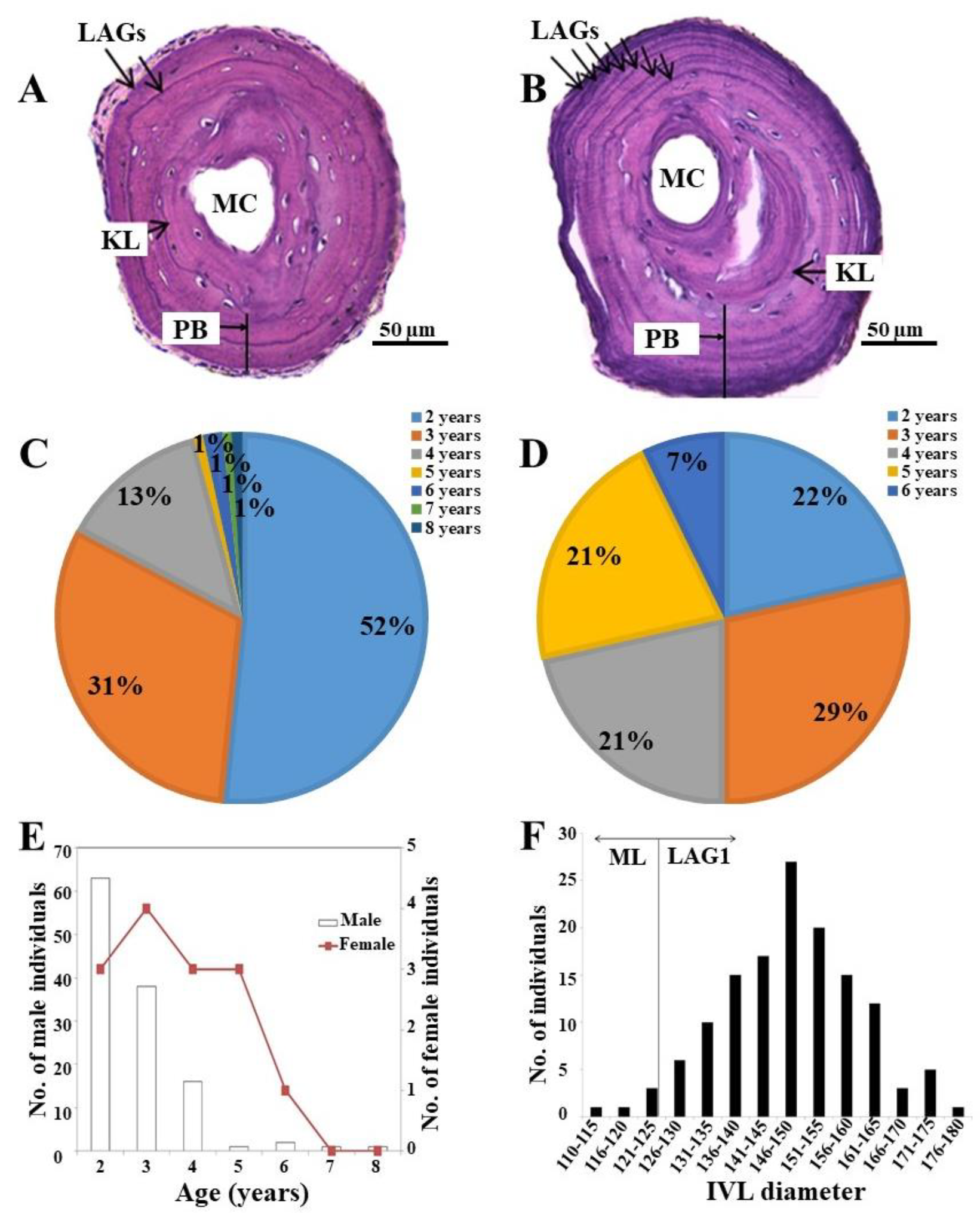
3.1. Bone Formation and the Appearance of LAG
3.2. Threshold Diameter for ML and LAG1
3.3. The Age Structure of the K. borealis Population in Lake Sihwa
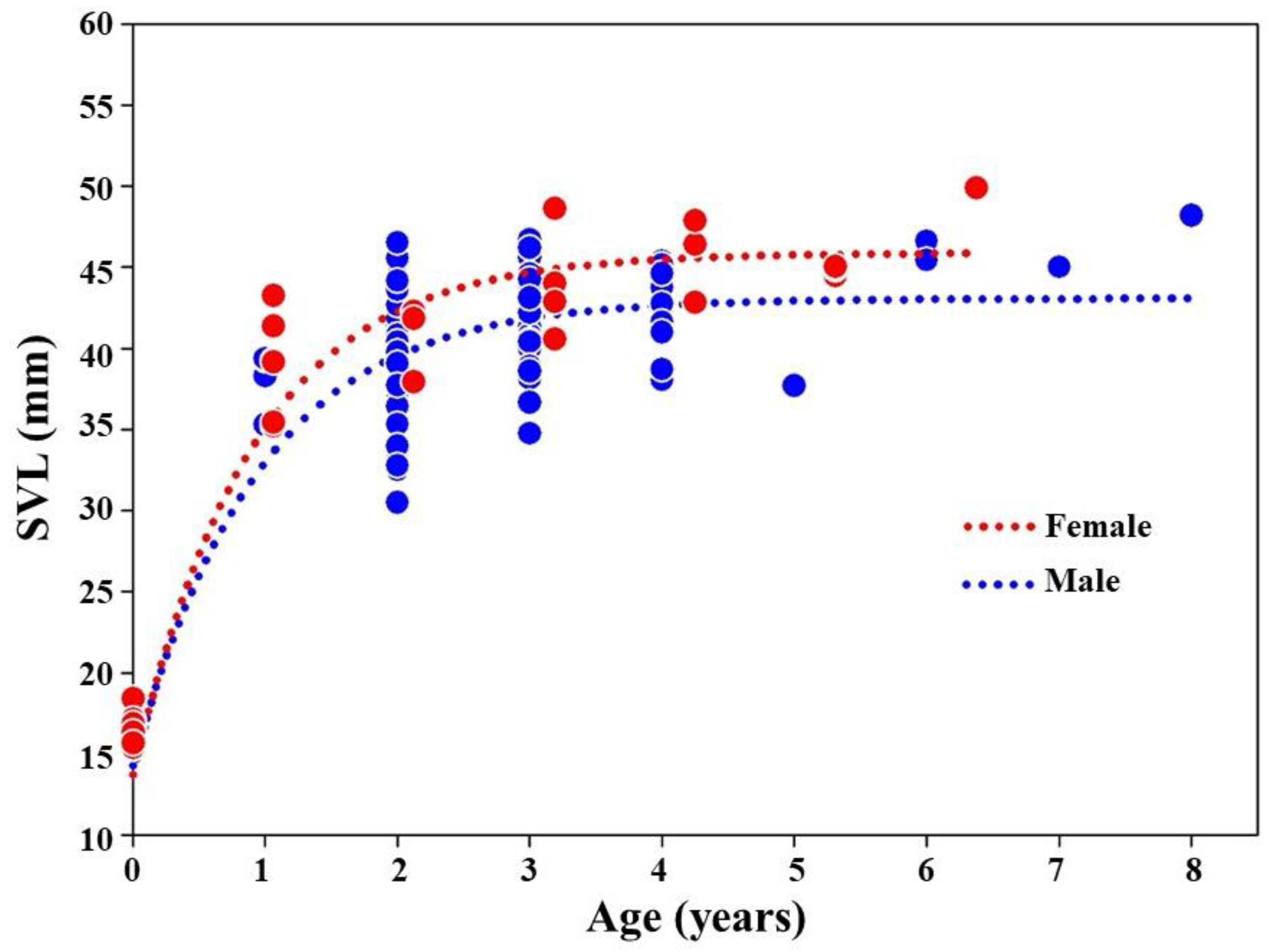
3.4. The Growth Curves
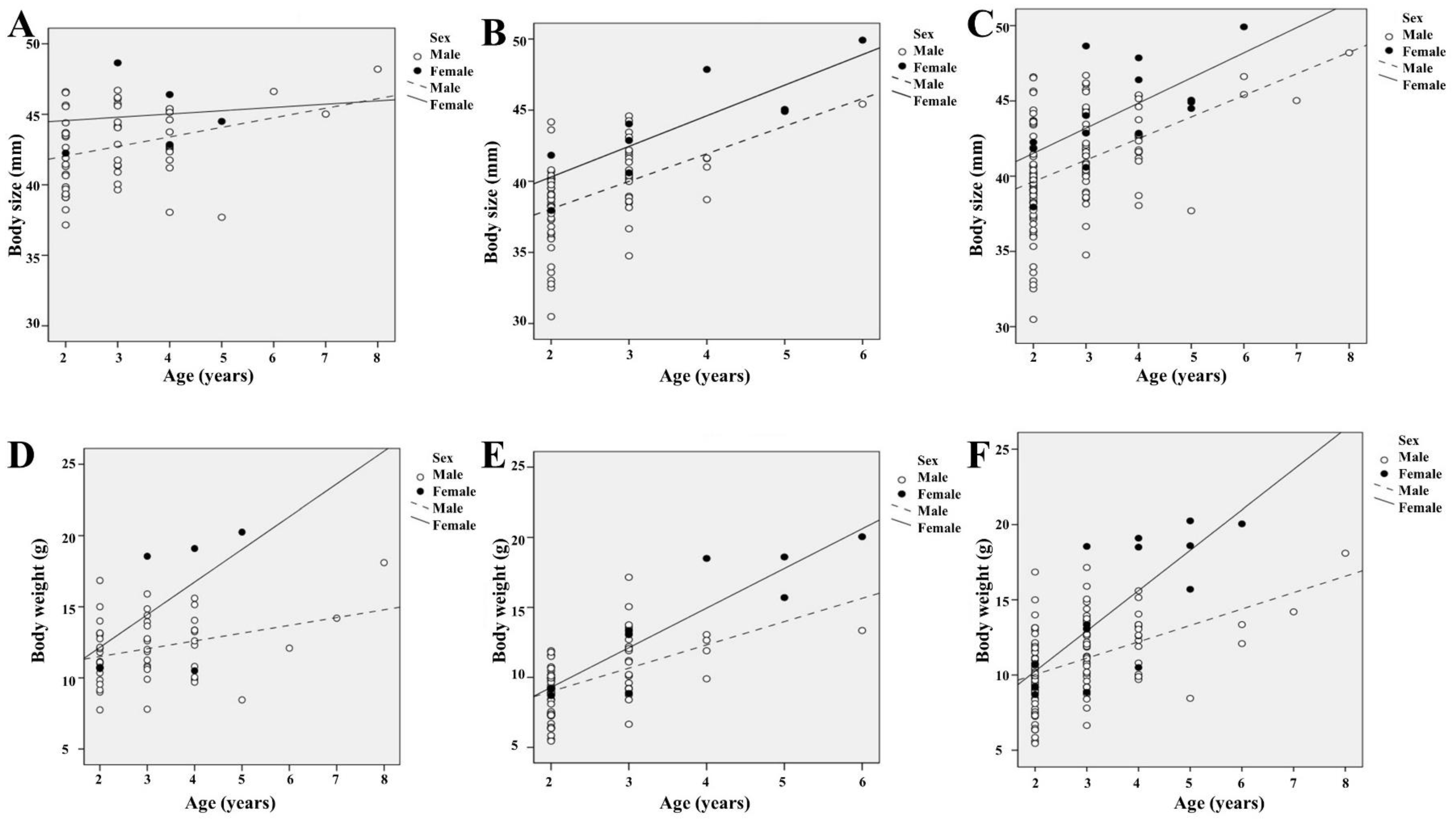
3.5. Relationship of Age with Body Size and Weight
3.6. Sexual Size Dimorphism
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biek, R.; Funk, W.C.; Maxell, B.A.; Mills, L.S. What is missing in amphibian decline research: Insights from ecological sensitivity analysis. Conserv. Biol. 2002, 16, 728–734. [Google Scholar] [CrossRef]
- Green, D.M. The ecology of extinction: Population fluctuation and decline in amphibians. Biol. Conserv. 2003, 111, 331–343. [Google Scholar] [CrossRef]
- Middleton, J.; Green, D.M. Adult age-structure variability in an amphibian in relation to population decline. Herpetologica 2015, 71, 190–195. [Google Scholar] [CrossRef]
- Zhong, M.J.; Yu, X.; Liao, W.B. A Review for life-history traits variation in frogs especially for anurans in China. Asian Herpetol. Res. 2018, 9, 165–174. [Google Scholar]
- Castanet, J. Age estimation and longevity in reptiles. Gerontology 1994, 40, 174–192. [Google Scholar] [CrossRef]
- Kusano, T.; Ueda, T.; Nakagawa, H. Body Size and Age Structure of Breeding Populations of the Salamander, Hynobius tokyoensis (Caudata: Hynobiidae). Curr. Herpetol. 2006, 25, 71–78. [Google Scholar]
- Meunier, F.J. Skeletochronological studies of cyclical growth of freshwater fishes in French Guiana: A review. Cybium 2012, 36, 55–62. [Google Scholar]
- Sinsch, U. Review: Skeletochronological assessment of demographic life-history traits in amphibians. Herpetol. J. 2015, 25, 5–13. [Google Scholar]
- Rahman, M.M. Impact of temperature fluctuations on gut histology of the cricket frog, Fejervarya limnocharis (Anura: Dicroglossidae). J. Bio-Sci. 2016, 22, 15–20. [Google Scholar] [CrossRef]
- Rozenblut, B.; Ogielska, M. Development and growth of long bones in european water frogs (Amphibia: Anura: Ranidae), with remarks on age determination. J. Morphol. 2005, 265, 304–317. [Google Scholar] [CrossRef]
- Smirina, E.M. Annual layers in bones of rana temporaria. Zool. Zhurnal 1972, 51, 1529–1534. [Google Scholar]
- Gibbons, M.M.; McCarthy, T.K. Age determination of frogs and toads (Amphibia, Anura) from North-Western Europe. Zool. Scr. 1983, 12, 145–151. [Google Scholar] [CrossRef]
- Hemelaar, A. An improved method to estimate the number of year rings resorbed in phalanges of Bufo bufo (L.) and its application to populations from different latitudes and altitudes. Amphib.-Reptil. 1985, 6, 323–341. [Google Scholar] [CrossRef]
- Castanet, J.; Smirina, E. Introduction to the skeletochronology method in amphibians and reptiles. Ann. Des Sci. Nat. Zool. 1990, 11, 191–196. [Google Scholar]
- Sagor, E.S.; Ouellet, M.; Barten, E.; Green, D.M. Skeletochronology and geographic variation in age structure in the wood frog, Rana sylvatica. J. Herpetol. 1998, 32, 469. [Google Scholar] [CrossRef]
- Tsiora, A.; Kyriakopoulou-Sklavounou, P. A skeletochronological study of age and growth in relation to adult size in the water frog Rana epeirotica. Zoology 2002, 105, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Zhang, L.; Lu, X. Global gaps in age data based on skeletochronology for amphibians. Integr. Zool. 2021, 00, 1–12. [Google Scholar] [CrossRef]
- Lee, J.-H.; Park, D.-S. Effects of physical parameters and age on the order of entrance of Hynobius leechii to a breeding pond. J. Ecol. Environ. 2008, 31, 183–191. [Google Scholar] [CrossRef]
- Lee, J.-H.; Ra, N.-Y.; Eom, J.-H.; Park, D.-S. Population dynamics of the long-tailed clawed salamander larva, Onychodactylus fischeri, and its age structure in Korea. J. Ecol. Environ. 2008, 31, 31–36. [Google Scholar] [CrossRef]
- Lee, J.-H.; Min, M.-S.; Kim, T.-H.; Baek, H.-J.; Lee, H.; Park, D. Age structure and growth rates of two korean salamander species (Hynobius yangi and Hynobius quelpaertensis) from field populations. Anim. Cells Syst. 2010, 14, 315–322. [Google Scholar] [CrossRef]
- Cheong, S.-W.; Park, D.-S.; Sung, H.-C.; Lee, J.-H.; Park, S.-R. Skeletochronological age determination and comparative demographic analysis of two populations of the gold-spotted pond frog (Rana chosenica). J. Ecol. Environ. 2007, 30, 57–62. [Google Scholar] [CrossRef]
- Su, M.J. Physical Characteristics of Breeding Huanren Brown Frogs (Rana huanrenensis), and the Age Structure of Their Population in the Research Forest of Kangwon National University. Master’s Thesis, Kangwon National University, Chuncheon, Korea, 2011; pp. 1–26. [Google Scholar]
- Ko, S.B.; Lee, J.H.; Oh, H.S. Age structure analysis of Kaloula borealis. Korean J. Environ. Ecol. 2011, 25, 861–866. [Google Scholar]
- Cheong, S.; Yoo, J.-H.; Park, S.-R.; Sung, H.-C. Age estimation by skeletochronology and advertisement call variation in the black-spotted pond frog (Rana nigromaculata). Anim. Cells Syst. 2013, 17, 141–146. [Google Scholar] [CrossRef]
- Ham, C.H. Morphology, Age Structure and Mating Call Characteristics of Japanese Tree Frog (Hyla japonica) and Suweon Tree Frog (Hyla suweonensis). Master’s Thesis, Chonnam National University, Gwangju, Korea, 2014; pp. 1–32. [Google Scholar]
- Kim, S.-B.; Suh, M.-H.; Lee, B.-Y.; Kim, S.T.; Park, C.-H.; Oh, H.-K.; Kim, H.-Y.; Lee, J.-H.; Lee, S.Y. (Eds.) Korean Red List of Threatened Species, 2nd ed.; National Institute of Biological Resources: Incheon, Korea, 2014; p. 71. [Google Scholar]
- IUCN. 2021. Available online: https://www.iucnredlist.org/species/57849/63887081#assessment-information (accessed on 2 June 2022).
- Yoon, J.; Kwon, K.; Yoo, J.; Yoo, N. Current status and future prospects of endangered species restoration projects for freshwater fishes, amphibians, and reptiles in South Korea. Proc. Natl. Inst. Ecol. Repub. Korea 2021, 2, 247–258. [Google Scholar]
- Gosner, L.K. A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica 1960, 16, 183–190. [Google Scholar]
- Von Bertalanffy, L. A quantitative theory of organic growth. Hum. Biol. 1938, 10, 181–213. [Google Scholar]
- Lovich, J.E.; Gibbons, J.W. A review of techniques for quantifying sexual size dimorphism. Growth Dev. Aging 1992, 56, 269–281. [Google Scholar]
- Bourdon, E.; Castanet, J.; de Ricqlès, A.; Scofield, P.; Tennyson, A.; Lamrous, H.; Cubo, J. Bone growth marks reveal protracted growth in New Zealand Kiwi (Aves, Apterygidae). Biol. Lett. 2009, 5, 639–642. [Google Scholar] [CrossRef]
- Marín-Moratalla, N.; Jordana, X.; Köhler, M. Bone histology as an approach to providing data on certain key life history traits in mammals: Implications for conservation biology. Mamm. Biol. 2013, 78, 422–429. [Google Scholar] [CrossRef]
- Bülbül, U.; Koç-Gür, H.; Albayrak, M.; Bayrak, M.O.; Özkan, H.; Kutrup, B. Body size and age structure of theLebanon lizard Phoenicolacerta laevis (Gray, 1838) (Reptilia: Lacertidae) in a lowland Turkish population. Acta Zool. Bulg. 2021, 73, 225–231. [Google Scholar]
- Pellegrini, R.A.; Callahan, W.R.; Hastings, A.K.; Parris, D.C.; McCauley, J.D. Skeletochronology and paleohistology of Hyposaurus rogersii (Crocodyliformes, Dyrosauridae) from the early Paleogene of New Jersey, USA. Animals 2021, 11, 3067. [Google Scholar] [CrossRef] [PubMed]
- van Gelder, J.J.; Hemelaar, A.S.M. Annual growth rings in phalanges of Bufo bufo (Anura, Amphibia) from the Netherlands and their use for age determination. Neth. J. Zool. 1979, 30, 129–135. [Google Scholar] [CrossRef]
- Ento, K.; Matsui, M. Estimation of age structure by skeletochronology of a population of Hynobius nebulosus in a breeding season (Amphibia, Urodela). Zool. Sci. 2002, 19, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Shirose, L.J.; Brooks, R.J.; Barta, J.R.; Desser, S.S. Intersexual differences in growth, mortality, and size at maturity in bullfrogs in central ontario. Can. J. Zool. 1993, 71, 2363–2369. [Google Scholar] [CrossRef]
- Kyriakopoulou-Sklavounou, P.; Stylianou, P.; Tsiora, A. A Skeletochronological study of age, growth and longevity in a population of the frog Rana ridibunda from Southern Europe. Zoology 2008, 111, 30–36. [Google Scholar] [CrossRef]
- Felton, A.; Alford, R.A.; Felton, A.M.; Schwarzkopf, L. Multiple mate choice criteria and the importance of age for male mating success in the microhylid frog, Cophixalus ornatus. Behav. Ecol. Sociobiol. 2006, 59, 786–795. [Google Scholar] [CrossRef]
- Tessa, G.; Guarino, F.M.; Randrianirina, J.E.; Andreone, F. Age structure in the false tomato frog Dyscophus guinetifrom eastern madagascar compared to the closely related D. antongilii (Anura, Microhylidae). Afr. J. Herpetol. 2011, 60, 84–88. [Google Scholar] [CrossRef]
- Sinsch, U.; Leskovar, C.; Drobig, A.; König, A.; Grosse, W.-R. Life-history traits in green toad (Bufo viridis) populations: Indicators of habitat quality. Can. J. Zool. 2007, 85, 665–673. [Google Scholar] [CrossRef]
- Kutrup, B.; Cakir, E.; Colak, Z.; Bulbul, U.; Karaoglu, H. Age and growth of the green toad, Bufo Viridis (Laurenti, 1768) from an island and a mainland population in Giresun, Turkey. J. Anim. Vet. Adv. 2011, 10, 1469–1472. [Google Scholar] [CrossRef]
- Leclair, R.; Castanet, J. A skeletochronological assessment of age and growth in the frog Rana pipiens Schreber (Amphibia, Anura) from Southwestern Quebec. Copeia 1987, 1987, 361–369. [Google Scholar] [CrossRef]
- Cherry, M.I.; Francillon-Vieillot, H. Body size, age and reproduction in the leopard toad, Bufo pardalis. J. Zool. 1992, 228, 41–50. [Google Scholar] [CrossRef]
- Yuan, L.; Zhong, M.; Liao, W. Age structure of two species of odorous frogs (Odorrana margaretae and Odorrana grahami). Asian Herpetol. Res. 2021, 12, 308–314. [Google Scholar] [CrossRef]
- Liao, W.B.; Lu, X. Variation in body size, age and growth in the omei treefrog (Rhacophorus omeimontis) along an altitudinal gradient in Western China. Ethol. Ecol. Evol. 2011, 23, 248–261. [Google Scholar] [CrossRef]
- Shine, R. Ecological causes for the evolution of sexual dimorphism: A review of the evidence. Q. Rev. Biol. 1989, 64, 419–461. [Google Scholar] [CrossRef] [PubMed]
- Hemelaar, A.S.M. Age of Bufo bufo in amplexus over the spawning period. Oikos 1983, 40, 1–5. [Google Scholar] [CrossRef]
- Guarino, F.M.; Lunardi, S.; Carlomagno, M.; Mazzotti, S. A skeletochronological study of growth, longevity, and age at sexual maturity in a population of Rana latastei (Amphibia, Anura). J. Biosci. 2003, 28, 775–782. [Google Scholar] [CrossRef]
- Fairbairn, D.J. Allometry for sexual size dimorphism: Pattern and process in the coevolution of body size in males and females. Annu. Rev. Ecol. Syst. 1997, 28, 659–687. [Google Scholar] [CrossRef]
- Trivers, R.L. Parent–offspring conflict. Am. Zool. 1974, 14, 249–264. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Life Stage | Number of Individuals | Lines | Diameter (μm) | |||
|---|---|---|---|---|---|---|
| Mean ± SD | Threshold * | Min | Max | |||
| Juvenile | 10 | ML | 109.75 ± 8.64 | 127 | 96.80 | 123.38 |
| LAG1 | 162.90 ± 8.63 | 180 | 146.35 | 174.69 | ||
| Life Stage | Sex | Number of Individuals | Age (Years) | ||
|---|---|---|---|---|---|
| Mean ± SD | Range | Z | |||
| Adult | Male | 122 | 2.75 ± 1.05 | 2–8 | −2.845 |
| Female | 14 | 3.64 ± 1.28 | 2–6 | ||
| Sex | Number of Individuals | Maximum Growth Size (mm; Mean ± SD) | Growth Coefficient (K; Mean ± SD) |
|---|---|---|---|
| Male | 122 | 43.08 ± 0.63 | 1.05 ± 0.10 |
| Female | 14 | 45.90 ± 1.06 | 1.16 ± 0.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.M.; Lee, Y.-Y.; Park, S.-M.; Ham, C.-H.; Sung, H.-C. Confirmation of the Applicability of Skeletochronology and Estimating the Age Structure of Kaloula borealis (Microhylidae: Anura) at Lake Sihwa, South Korea. Biology 2022, 11, 898. https://doi.org/10.3390/biology11060898
Rahman MM, Lee Y-Y, Park S-M, Ham C-H, Sung H-C. Confirmation of the Applicability of Skeletochronology and Estimating the Age Structure of Kaloula borealis (Microhylidae: Anura) at Lake Sihwa, South Korea. Biology. 2022; 11(6):898. https://doi.org/10.3390/biology11060898
Chicago/Turabian StyleRahman, Md Mizanur, Yu-Young Lee, Seung-Min Park, Choong-Ho Ham, and Ha-Cheol Sung. 2022. "Confirmation of the Applicability of Skeletochronology and Estimating the Age Structure of Kaloula borealis (Microhylidae: Anura) at Lake Sihwa, South Korea" Biology 11, no. 6: 898. https://doi.org/10.3390/biology11060898