
Molecular Identification and Characterization of Two Groups of Phytoplasma and Candidatus Liberibacter Asiaticus in Single or Mixed Infection of Citrus maxima on Hainan Island of China


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
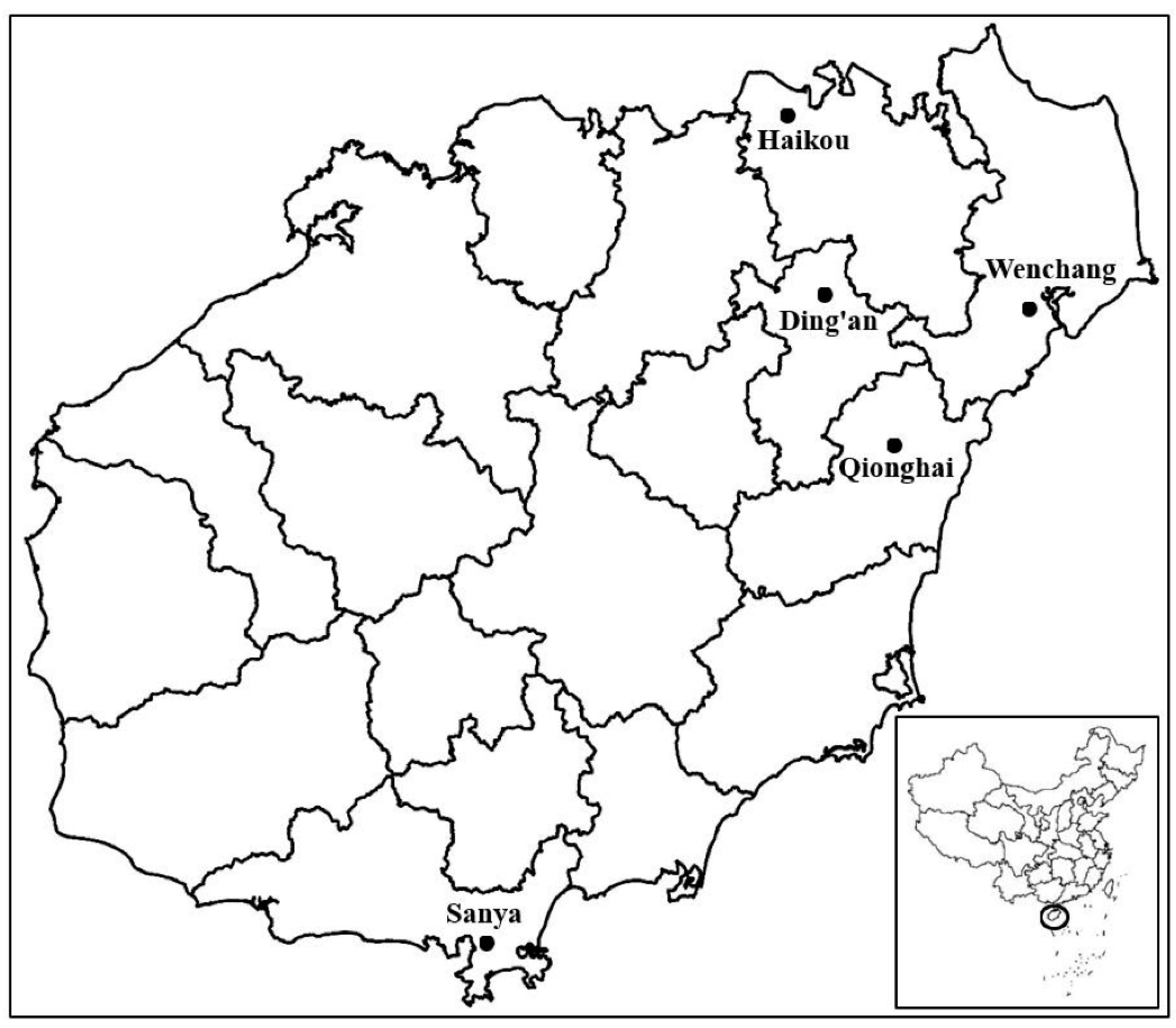
2.1. Plant Samples and DNA Preparation
2.2. PCR Amplification and DNA Sequencing
2.3. Sequence Analysis
2.4. Nucleotide Sequence Accession Numbers
3. Results
3.1. Molecular Detection and Identification of the Pathogens Associated with the Disease Symptoms in Citrus Maxima
3.2. Virtual RFLP Analysis of the Citrus Maxima Phytoplasma Strains
3.3. Genetic Variation Characteristics of the Phytoplasma and the Candidatus Liberibacter Asiaticus Infecting Citrus Maxima
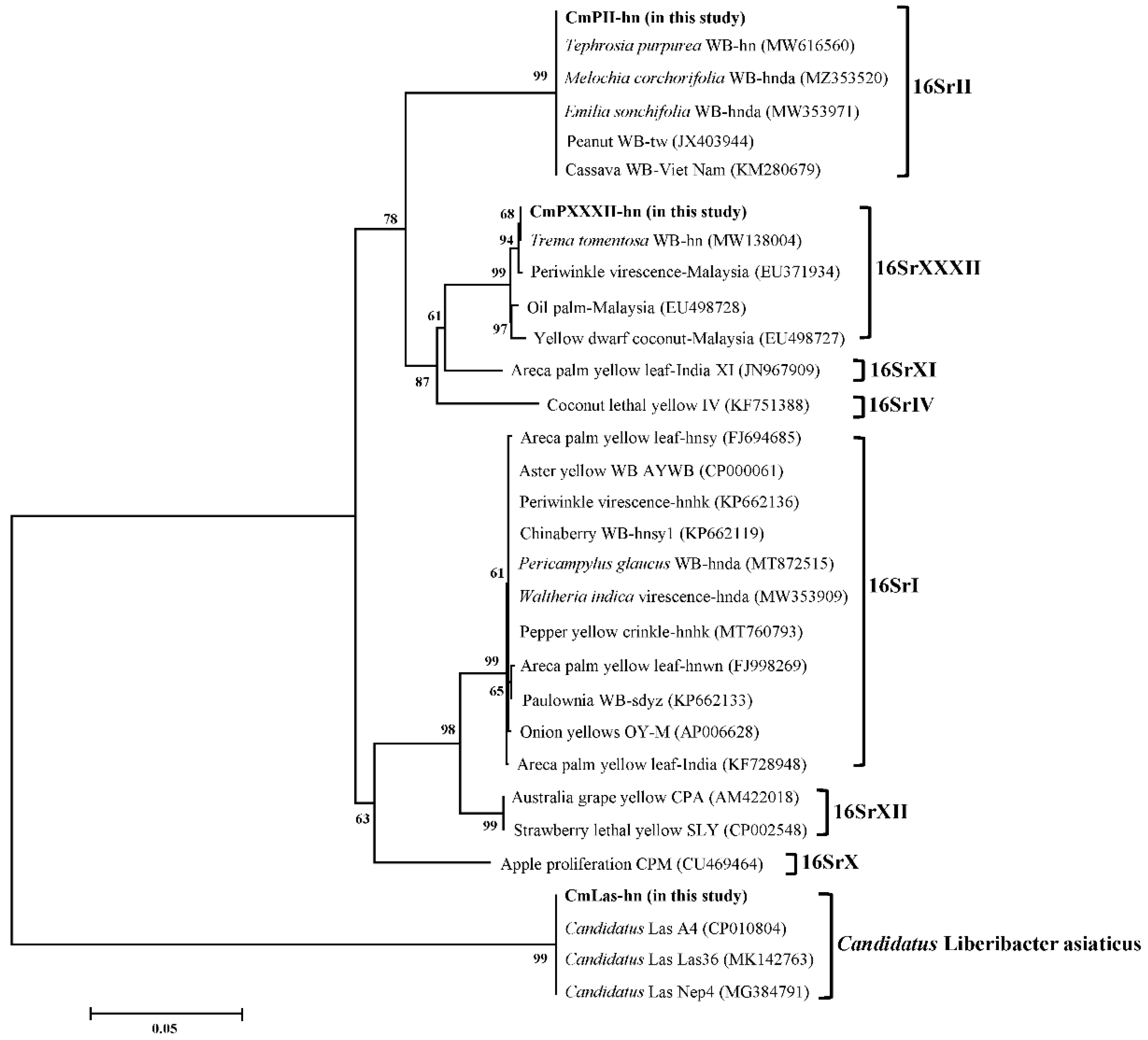
3.4. Phylogenetic Analysis of the Plant Pathogens Identified in the Study
4. Discussion
4.1. Citrus Maxima Single or Mixed Infection by the Phytoplasma and Candidatus Liberibacter Asiaticus
4.2. The Plant Host Diversity of the Phytoplasmas and the Epidemiology of the Related Phytoplasma Diseases
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Doi, Y.; Teranaka, M.; Yora, K.; Asuyama, H. Mycoplasma-or PLT group-like microorganisms found in the phloem elements of plants infected with mulberry dwarf, potato witches’ broom, aster yellows, or paulownia witches’-broom. Jpn. J. Phytopathol. 1967, 33, 259–266. [Google Scholar] [CrossRef]
- Wei, W.; Trivellone, V.; Dietrich, C.H.; Zhao, Y.; Bottner-Parker, K.D.; Ivanauskas, A. Identification of Phytoplasmas Representing Multiple New Genetic Lineages from Phloem-Feeding Leafhoppers Highlights the Diversity of Phytoplasmas and Their Potential Vectors. Pathogens 2021, 10, 352. [Google Scholar] [CrossRef]
- Hemmati, C.; Nikooei, M.; Al-Subhi, A.M.; Al-Sadi, A.M. History and Current Status of Phytoplasma Diseases in the Middle East. Biology 2021, 10, 226. [Google Scholar] [CrossRef]
- IRPCM. ‘Candidatus Phytoplasma’, a taxon for the wall-less, non-helical prokaryotes that colonize plant phloem and insects. Int. J. Syst. Evol. Microbiol. 2004, 54, 1243–1255. [Google Scholar] [CrossRef] [Green Version]
- Bertaccini, A.; Duduk, B.; Paltrinieri, S.; Contaldo, N. Phytoplasmas and phytoplasma diseases: A severe threat to agriculture. Am. J. Plant Sci. 2014, 5, 1763–1788. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, M.; Hodgetts, J. Phytoplasma: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2013. [Google Scholar]
- Sugio, A.; Maclean, A.M.; Kingdom, H.N.; Grieve, V.M.; Manimekalai, R.; Hogenhout, S.A. Diverse targets of phytoplasma effectors: From plant development to defense against insects. Annu. Rev. Phytopathol. 2011, 49, 175–195. [Google Scholar] [CrossRef]
- Ramos, A.; Esteves, M.B.; Cortes, M.T.B.; Lopes, J.R.S. Maize bushy stunt phytoplasma favors its spread by changing host preference of the insect vector. Insects 2020, 11, 600. [Google Scholar] [CrossRef]
- Mori, N.; Cargnus, E.; Martini, M.; Pavan, F. Relationships between Hyalesthes obsoletus, its herbaceous hosts and Bois Noir epidemiology in northern Italian Vineyards. Insects 2020, 11, 606. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, E.A.; Farrag, A.A.; Kheder, A.A.; Shaaban, A. Effect of Phytoplasma Associated with Sesame Phyllody on Ultrastructural Modification, Physio-Biochemical Traits, Productivity and Oil Quality. Plants 2022, 11, 477. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Inaba, J.; Zhao, Y.; Mowery, J.D.; Hammond, R. Phytoplasma Infection Blocks Starch Breakdown and Triggers Chloroplast Degradation, Leading to Premature Leaf Senescence, Sucrose Reallocation, and Spatiotemporal Redistribution of Phytohormones. Int. J. Mol. Sci. 2022, 23, 1810. [Google Scholar] [CrossRef]
- Lee, I.M.; Hammond, R.W.; Davis, R.E.; Gundersen, D.E. Universal amplification and analysis of pathogen 16S rDNA for classification and identification of mycoplasma like organisms. Phytopathology 1993, 83, 834–842. [Google Scholar] [CrossRef]
- Wei, W.; Davis, R.E.; Lee, I.M.; Zhao, Y. Computer-simulated RFLP analysis of 16S rRNA genes: Identification of ten new phytoplasma groups. Int. J. Syst. Evol. Microbiol. 2007, 57, 1855–1867. [Google Scholar] [CrossRef]
- Yu, S.S.; Li, Y.; Ren, Z.G.; Song, C.S.; Lin, C.L.; Piao, C.G.; Tian, G.Z. Multilocus sequence analysis for revealing finer genetic variation and phylogenetic interrelatedness of phytoplasma strains in 16SrI group in China. Sci. Silva Sin. 2017, 53, 105–118. [Google Scholar]
- Curcic, Ž.; Kosovac, A.; Stepanovic, J.; Rekanovic, E.; Kube, M.; Duduk, B. Multilocus Genotyping of ‘Candidatus Phytoplasma solani’ Associated with Rubbery Taproot Disease of Sugar Beet in the Pannonian Plain. Microorganisms 2021, 9, 1950. [Google Scholar] [CrossRef]
- Seemüller, E.; Marcone, C.; Lauer, U.; Ragozzino, A.; Goschl, M. Current status of molecular classification of the phytoplasmas. J. Plant Pathol. 1998, 80, 3–26. [Google Scholar]
- Bao, M.; Zheng, Z.; Chen, J.; Deng, X. Investigation of citrus HLB symptom variations associated with “Candidatus Liberibacter asiaticus” strains harboring different phages in Southern China. Agronomy 2021, 11, 2262. [Google Scholar] [CrossRef]
- Nan, J.; Zhang, S.; Zhan, P.; Jiang, L. Discovery of novel GMPS inhibitors of Candidatus Liberibacter asiaticus by structure based design and enzyme kinetic. Biology 2021, 10, 594. [Google Scholar] [CrossRef]
- Li, B.; Wang, S.; Zhang, Y.; Qiu, D. Acid soil improvement enhances disease tolerance in citrus infected by Candidatus Liberibacter asiaticus. Int. J. Mol. Sci. 2020, 21, 3614. [Google Scholar] [CrossRef]
- Dorado, C.; Bowman, K.D.; Cameron, R.G.; Manthey, J.A.; Bai, J.; Ferguson, K.L. Steam explosion (STEX) of Citrus × Poncirus hybrids with exceptional tolerance to Candidatus Liberibacter asiaticus (CLas) as useful sources of volatiles and other commercial products. Biology 2021, 10, 1285. [Google Scholar] [CrossRef] [PubMed]
- Arce-Leal, A.P.; Bautista, R.; Rodriguez-Negrete, E.A.; Manzanilla-Ramirez, M.A.; Velazquez-Monreal, J.J.; Santos-Cervantes, M.E.; Mendez-Lozano, J.; Beuzon, C.R.; Bejarano, E.R.; Castillo, A.G.; et al. Gene expression profile of Mexican Lime (Citrus aurantifolia) trees in response to Huanglongbing disease caused by Candidatus Liberibacter asiaticus. Microorganisms 2020, 8, 528. [Google Scholar] [CrossRef] [Green Version]
- Quiroga, N.; Gamboa, C.; Medina, G.; Contaldo, N.; Torres, F.; Bertaccini, A.; Zamorano, A.; Fiore, N. Survey for ‘Candidatus Liberibacter’ and ‘Candidatus Phytoplasma’ in Citrus in Chile. Pathogens 2022, 11, 48. [Google Scholar] [CrossRef]
- Yang, Y.; Li, D.; Duan, Y.; Jiang, L. Investigation and distribution of the Citrus Huanglongbing and identification of Huanglongbing liberibacter species in Hainan. Plant Quar. 2020, 34, 43–47. [Google Scholar]
- Chen, J.; Zheng, Z.; Deng, X. Research on the movement and propagation of “Candidatus Liberibacter asiaticus” and “Candidatus Phytoplasma asteri” in periwinkle and their associated symptoms. Acta Phytopathol. Sin. 2019, 49, 160–166. [Google Scholar]
- Teixeira, D.D.C.; Wulff, N.A.; Martins, E.; Elliot, K.; Bassanezi, R.B.; Ayres, A.J.; Eveillard, S.; Saillard, C.; Bové, J. A phytoplasma closely related to the pigeon pea witches’-broom phytoplasma (16SrIX) is associated with Citrus Huanglongbing symptoms in the State of São Paulo State, Brazil. Phytopathology 2008, 98, 977–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Pu, X.; Deng, X.; Liu, S.; Li, H.; Civerolo, E. A phytoplasma related to ‘Candidatus Phytoplasma asteri’ detected in citrus showing Huanglongbing (yellow shoot disease) symptoms in Guangdong, P.R. China. Phytopathology 2009, 99, 236–242. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.S.; Wu, Y.; Song, W.W. Occurrence of a 16SrII-V subgroup phytoplasma associated with witches’-broom disease in Melochia corchorifolia in China. Plant Dis. 2022, 106, 754. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.S.; Zhao, R.L.; Lin, M.X.; Wu, Y.; Chen, S.G.; Yu, F.Y.; Song, W.W.; Zhu, H. Occurrence of phytoplasma belongs to 16SrII group associated with witches’-broom symptoms in Emilia sonchifolia in Hainan Island of China. Plant Dis. 2021, 105, 4151. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.S.; Tang, Q.H.; Wu, Y.; Lin, M.X.; Zhao, R.L.; Song, W.W.; Qin, W.Q. First report of 16SrXXXII group related phytoplasma associated with Trema tomentosa witches’-broom disease in China. Plant Dis. 2021, 105, 1191. [Google Scholar] [CrossRef]
- Yu, S.S.; Zhao, R.L.; Lin, M.X.; Wu, Y.; Chen, S.G.; Yu, F.Y.; Song, W.W.; Zhu, H. Waltheria indica is a new host of phytoplasma belongs to 16SrI-B subgroup associated with virescence symptoms in China. Plant Dis. 2021, 105, 2012. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Gundersen, D.E.; Lee, I.M. Ultrasensitive detection of phytoplasmas by nested-PCR assays using two universal primer pairs. Phytopathol. Mediterr. 1996, 35, 144–151. [Google Scholar]
- Jagoueix, S.; Bove, J.M.; Garnier, M. PCR detection of two Candidatus Liberobacter species associated with greening disease of citrus. Mol. Cell. Probes 1996, 10, 43–50. [Google Scholar] [CrossRef]
- Hocquellet, A.; Toorawa, P.; Bové, J.M.; Garnier, M. Detection and identification of the two Candidatus Liberobacter species associated with citrus huanglongbing by PCR amplification of ribosomal protein genes of the β-operon. Mol. Cell. Probes 1999, 13, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wei, W.; Lee, I.M.; Shao, J.; Suo, X.; Davis, R.E. Construction of an interactive online phytoplasma classification tool, iPhyClassifier, and its application in analysis of the peach X-disease phytoplasma group (16SrIII). Int. J. Syst. Evol. Microbiol. 2009, 59, 2582–2593. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Che, H.; Wu, C.; Fu, R.; Wen, Y.; Ye, S.; Luo, D. Molecular identification of pathogens from arecanut yellow leaf disease in Hainan. Chin. J. Trop. Crops 2010, 31, 83–87. [Google Scholar]
- Yu, S.S.; Song, W.W.; Qin, W.Q. Molecular detection and phylogenetic relationship of areca palm yellow leaf phytoplasma in Hainan, China. Chin. J. Trop. Crops 2021, 42, 3066–3072. [Google Scholar]
- Sun, X.C.; Mou, H.Q.; Li, T.T.; Tian, Q.; Zhao, W.J. Mixed infection of two groups (16SrI and V) of phytoplasmas in a single jujube tree in China. J. Phytopathol. 2013, 161, 661–665. [Google Scholar] [CrossRef]
- Arratia-Castro, A.A.; Santos-Cervantes, M.E.; Fernández-Herrera, E.; Chávez-Medina, J.A.; Flores-Zamora, G.L.; Camacho-Beltrán, E.; Méndez-Lozano, J.; Leyva-López, N.E. Occurrence of ‘Candidatus Phytoplasma asteris’ in citrus showing Huanglongbing symptoms in Mexico. Crop Prot. 2014, 62, 144–151. [Google Scholar] [CrossRef]
- Yu, S.S.; Zhao, R.L.; Lin, M.X.; Wu, Y.; Song, W.W.; Yan, W. Tephrosia purpurea represents a new host of 16SrII-V subgroup phytoplasma associated with witches’-broom disease in China. Plant Dis. 2021, 105, 2235. [Google Scholar] [CrossRef]
- Chen, W.; Li, Y.; Liu, F.; Zeng, L.Y.; Ren, L.; Xu, L.; Chen, R.K.; Fang, X.P. Melochia corchorifolia, a new host of 16SrI-B phytoplasma in China. J. Plant Pathol. 2017, 99, 291. [Google Scholar]
- Gurr, G.M.; Johnson, A.C.; Ash, G.J.; Wilson, B.A.L.; Ero, M.M.; Pilotti, C.A.; Dewhurst, C.F.; You, M.S. Coconut lethal yellowing diseases: A phytoplasma threat to palms of global economic and social significance. Front. Plant Sci. 2016, 7, 1521. [Google Scholar] [CrossRef] [PubMed]
- Babu, M.; Thangeswari, S.; Josephrajkumar, A.; Krishnakumar, V.; Karthikeyan, A.; Selvamani, V.; Daliya, M.; Hegde, V.; Maheswarappa, H.P.; Karun, A. First report on the association of ‘Candidatus Phytoplasma asteris’ with lethal wilt disease of coconut (Cocos nucifera L.) in India. J. Gen. Plant Pathol. 2021, 87, 16–23. [Google Scholar] [CrossRef]
- Ramaswamy, M.; Nair, S.; Soumya, V.P.; Thomas, G.V. Phylogenetic analysis identifies, a ‘Candidatus Phytoplasma oryzae’-related strain associated with yellow leaf disease of areca palm (Areca catechu L.) in India. Int. J. Syst. Evol. Microbiol. 2013, 63, 1376–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, C.K.D.; Dahanayake, D.; Silva, R.D.; Dickinson, M.; Silva, K.M.N.D.; Udagama, P.V. Molecular and scanning electron microscopic proof of phytoplasma associated with areca palm yellow leaf disease in Sri Lanka. Plant Dis. 2015, 99, 1641. [Google Scholar] [CrossRef]
- Kazeem, S.A.; Inaba, J.; Zhao, Y.; Zwolinska, A.; Ogunfunmilayo, A.O.; Arogundade, O.; Wei, W. Molecular identification and characterization of ‘Candidatus Phytoplasma convolvuli’-related strains (representing a new 16SrXII-O subgroup) associated with papaya bunchy top disease in Nigeria. Crop Prot. 2021, 148, 105731. [Google Scholar] [CrossRef]
- Wang, J.; Tian, G.Z.; Xu, Q.C.; Liu, Y.G.; Gao, R.; Li, X.D.; Zhu, X.P. Molecular detection of phytoplasma strains from several plants around diseased paulownia infected with paulownia witches’-broom phytoplasma. Sci. Agric. Sin. 2010, 43, 304–312. [Google Scholar]
- Lin, J.; Yang, C.; Liu, J.; Yu, S.; Xing, J.; Huang, P.; Chen, W.; Bao, Y.; Hu, Q.; Chen, C.; et al. Identification and characterization of the phytoplasma associated with lettuce chlorotic leaf rot disease together with its natural reservoirs and leafhopper vectors in China. Crop Prot. 2020, 138, 105318. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytoplasma Strains | Subgroups | Polymorphic Nucleotide Sites | |||||||||||||||
| 1 | 1 | 1 | |||||||||||||||
| 1 | 1 | 3 | 6 | 8 | 8 | 8 | 9 | 1 | 2 | 2 | |||||||
| 3 | 3 | 3 | 3 | 4 | 1 | 3 | 4 | 8 | 1 | 2 | 2 | 6 | 8 | 6 | 7 | ||
| 6 | 7 | 8 | 9 | 2 | 5 | 0 | 6 | 6 | 8 | 1 | 9 | 7 | 3 | 3 | 2 | ||
| Trema tomentosa WB-hn (MW138004) | 16SrXXXII-D | C | A | T | T | C | A | T | T | A | G | T | T | C | A | T | C |
| CmPXXXII-hn (in this study) | 16SrXXXII-D | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • | • |
| Periwinkle virescence-Malaysia (EU371934) | 16SrXXXII-A | • | • | • | • | • | G | • | • | G | • | • | • | - | G | • | • |
| Yellow dwarf coconut-Malaysia (EU498727) | 16SrXXXII-B | T | G | A | C | T | G | • | • | • | • | A | C | • | • | - | G |
| Oil palm-Malaysia (EU498728) | 16SrXXXII-C | • | G | A | C | G | G | C | C | • | A | • | • | • | • | • | G |
| Phytoplasma Strains | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 CmPII-hn (in this study) | 100 | |||||||||||||||||||||
| 2 CmPXXXII-hn (in this study) | 90.6 | 100 | ||||||||||||||||||||
| 3 Tephrosia purpurea WB-hn (MW616560) | 100 | 90.6 | 100 | |||||||||||||||||||
| 4 Melochia corchorifolia WB-hnda (MZ353520) | 100 | 90.6 | 100 | 100 | ||||||||||||||||||
| 5 Pepper yellow crinkle-hnhk (MT760793) | 90.0 | 90.7 | 90.0 | 90.0 | 100 | |||||||||||||||||
| 6 Waltheria indica virescence-hnda (MW353909) | 90.0 | 90.7 | 90.0 | 90.0 | 100 | 100 | ||||||||||||||||
| 7 Emilia sonchifolia WB-hnda (MW353971) | 100 | 90.6 | 100 | 100 | 90.0 | 90.0 | 100 | |||||||||||||||
| 8 Pericampylus glaucus WB-hnda (MT872515) | 90.0 | 90.7 | 90.0 | 90.0 | 100 | 100 | 90.0 | 100 | ||||||||||||||
| 9 Areca palm yellow leaf-hnwn (FJ998269) | 90.0 | 90.6 | 90.0 | 90.0 | 99.8 | 99.8 | 90.0 | 99.8 | 100 | |||||||||||||
| 10 Coconut lethal yellow IV (KF751388) | 91.4 | 93.6 | 91.4 | 91.4 | 90.2 | 90.2 | 91.4 | 90.2 | 90.1 | 100 | ||||||||||||
| 11 Peanut WB-tw (JX403944) | 100 | 90.6 | 100 | 100 | 90.0 | 90.0 | 100 | 90.0 | 90.0 | 91.4 | 100 | |||||||||||
| 12 Chinaberry WB-hnsy1 (KP662119) | 90.0 | 90.7 | 90.0 | 90.0 | 100 | 100 | 90.0 | 100 | 99.8 | 90.2 | 90.0 | 100 | ||||||||||
| 13 Periwinkle virescence-hnhk (KP662136) | 90.0 | 90.7 | 90.0 | 90.0 | 100 | 100 | 90.0 | 100 | 99.8 | 90.2 | 90.0 | 100 | 100 | |||||||||
| 14 Trema tomentosa WB-hn (MW138004) | 90.6 | 100 | 90.6 | 90.6 | 90.7 | 90.7 | 90.6 | 90.7 | 90.6 | 93.6 | 90.6 | 90.7 | 90.7 | 100 | ||||||||
| 15 Periwinkle virescence-Malaysia (EU371934) | 90.5 | 99.8 | 90.5 | 90.5 | 90.7 | 90.7 | 90.5 | 90.7 | 90.5 | 93.5 | 90.5 | 90.7 | 90.7 | 99.8 | 100 | |||||||
| 16 Yellow dwarf coconut-Malaysia (EU498727) | 90.1 | 99.3 | 90.1 | 90.1 | 90.7 | 90.7 | 90.1 | 90.7 | 90.5 | 93.1 | 90.1 | 90.7 | 90.7 | 99.3 | 99.2 | 100 | ||||||
| 17 Oil palm-Malaysia (EU498728) | 90.2 | 99.4 | 90.2 | 90.2 | 90.7 | 90.7 | 90.2 | 90.7 | 90.6 | 93.2 | 90.2 | 90.7 | 90.7 | 99.4 | 99.3 | 99.4 | 100 | |||||
| 18 Onion yellows OY-M (AP006628) | 89.9 | 90.8 | 89.9 | 89.9 | 99.9 | 99.9 | 89.9 | 99.9 | 99.8 | 90.1 | 89.9 | 99.9 | 99.9 | 90.8 | 90.7 | 90.7 | 90.8 | 100 | ||||
| 19 Aster yellow WB AYWB (CP000061) | 90.0 | 90.7 | 90.0 | 90.0 | 100 | 100 | 90.0 | 100 | 99.8 | 90.2 | 90.0 | 100 | 100 | 90.7 | 90.6 | 90.6 | 90.7 | 99.9 | 100 | |||
| 20 Australia grape yellow CPA (AM422018) | 89.6 | 90.6 | 89.6 | 89.6 | 96.6 | 96.6 | 89.6 | 96.6 | 96.5 | 89.8 | 89.6 | 96.6 | 96.6 | 90.6 | 90.4 | 90.4 | 90.5 | 96.5 | 96.6 | 100 | ||
| 21 Apple proliferation CPM (CU469464) | 89.9 | 91.4 | 89.9 | 89.9 | 92.9 | 92.9 | 89.9 | 92.9 | 92.7 | 91.5 | 89.9 | 92.9 | 92.9 | 91.4 | 91.2 | 91.2 | 91.3 | 92.8 | 92.9 | 92.9 | 100 | |
| 22 Strawberry lethal yellow SLY (CP002548) | 89.6 | 90.6 | 89.6 | 89.6 | 96.6 | 96.6 | 89.6 | 96.6 | 96.5 | 89.8 | 89.6 | 96.6 | 96.6 | 90.6 | 90.4 | 90.4 | 90.5 | 96.5 | 96.6 | 100 | 92.9 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.-S.; Zhu, A.-N.; Song, W.-W.; Yan, W. Molecular Identification and Characterization of Two Groups of Phytoplasma and Candidatus Liberibacter Asiaticus in Single or Mixed Infection of Citrus maxima on Hainan Island of China. Biology 2022, 11, 869. https://doi.org/10.3390/biology11060869
Yu S-S, Zhu A-N, Song W-W, Yan W. Molecular Identification and Characterization of Two Groups of Phytoplasma and Candidatus Liberibacter Asiaticus in Single or Mixed Infection of Citrus maxima on Hainan Island of China. Biology. 2022; 11(6):869. https://doi.org/10.3390/biology11060869
Chicago/Turabian StyleYu, Shao-Shuai, An-Na Zhu, Wei-Wei Song, and Wei Yan. 2022. "Molecular Identification and Characterization of Two Groups of Phytoplasma and Candidatus Liberibacter Asiaticus in Single or Mixed Infection of Citrus maxima on Hainan Island of China" Biology 11, no. 6: 869. https://doi.org/10.3390/biology11060869