


Extracellular Release of HMGB1 as an Early Potential Biomarker for the Therapeutic Response in a Xenograft Model of Boron Neutron Capture Therapy
 , , , , ,
, , , , , 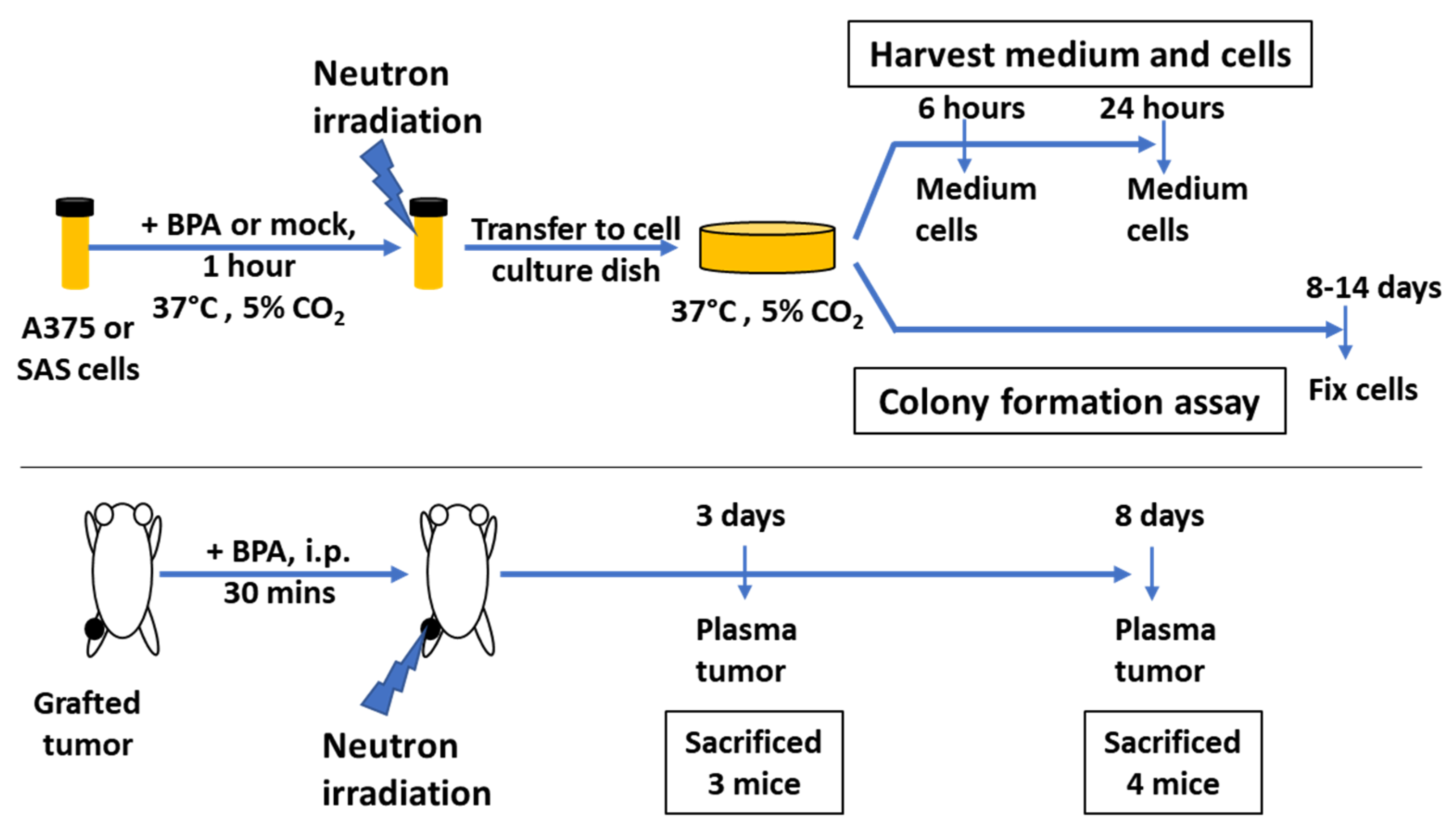{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Irradiation
2.3. Clonogenic Assay
2.4. ELISA
2.5. Immunohistochemistry (IHC)
2.6. Tumorigenesis Analysis and Irradiation
2.7. Statistical Analysis
3. Results
3.1. Effect of BNCT on the Survival of SAS Cells
3.2. Enhanced Extracellular HMGB1 Release from BPA-Pretreated SAS Cells after Neutron Irradiation
3.3. Extracellular HMGB1 Release from BPA-Pretreated A375 Cells after Neutron Irradiation
3.4. HMGB1 Distribution in Tumors after BPA Pretreatment and Neutron Irradiation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Suzuki, M. Boron neutron capture therapy (bnct): A unique role in radiotherapy with a view to entering the accelerator-based bnct era. Int. J. Clin. Oncol. 2020, 25, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Miyatake, S.I.; Wanibuchi, M.; Hu, N.; Ono, K. Boron neutron capture therapy for malignant brain tumors. J. Neurooncol. 2020, 149, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sauerwein, W.; Wittig, A.; Moss, R.; Nakagawa, Y. Neutron Capture Therapy Principles and Applications; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2012. [Google Scholar]
- Kinashi, Y.; Sakurai, Y.; Masunaga, S.; Suzuki, M.; Nagata, K.; Ono, K. Evaluation of micronucleus induction in lymphocytes of patients following boron-neutron-capture-therapy: A comparison with thyroid cancer patients treated with radioiodine. J. Radiat. Res. 2007, 48, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Masunaga, S.I.; Kinashi, Y.; Takagaki, M.; Akaboshi, M.; Kobayashi, T.; Akuta, K. Radiobiological evidence suggesting heterogeneous microdistribution of boron compounds in tumors: Its relation to quiescent cell population and tumor cure in neutron capture therapy. Int. J. Radiat. Oncol. Biol. Phys. 1996, 34, 1081–1086. [Google Scholar] [CrossRef]
- Kondo, N.; Michiue, H.; Sakurai, Y.; Tanaka, H.; Nakagawa, Y.; Watanabe, T.; Narabayashi, M.; Kinashi, Y.; Suzuki, M.; Masunaga, S.; et al. Detection of gammah2ax foci in mouse normal brain and brain tumor after boron neutron capture therapy. Rep. Pract. Oncol. Radiother. 2016, 21, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, C.; Carpano, M.; Curotto, P.; Thorp, S.; Casal, M.; Juvenal, G.; Pisarev, M.; Dagrosa, M.A. In vitro studies of DNA damage and repair mechanisms induced by bnct in a poorly differentiated thyroid carcinoma cell line. Radiat. Environ. Biophys. 2018, 57, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Masutani, M.; Baiseitov, D.; Itoh, T.; Hirai, T.; Berikkhanova, K.; Murakami, Y.; Zhumadilov, Z.; Imahori, Y.; Hoshi, M.; Itami, J. Histological and biochemical analysis of DNA damage after bnct in rat model. Appl. Radiat. Isot. 2014, 88, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Itoh, T.; Imamichi, S.; Kikuhara, S.; Fujimori, H.; Hirai, T.; Saito, S.; Sakurai, Y.; Tanaka, H.; Nakamura, H.; et al. Proteomic analysis of cellular response induced by boron neutron capture reaction in human squamous cell carcinoma sas cells. Appl. Radiat. Isot. 2015, 106, 213–219. [Google Scholar] [CrossRef]
- Chiba, S.; Baghdadi, M.; Akiba, H.; Yoshiyama, H.; Kinoshita, I.; Dosaka-Akita, H.; Fujioka, Y.; Ohba, Y.; Gorman, J.V.; Colgan, J.D.; et al. Tumor-infiltrating dcs suppress nucleic acid-mediated innate immune responses through interactions between the receptor tim-3 and the alarmin hmgb1. Nat. Immunol. 2012, 13, 832–842. [Google Scholar] [CrossRef]
- Andersson, U.; Tracey, K.J. Hmgb1 is a therapeutic target for sterile inflammation and infection. Annu. Rev. Immunol. 2011, 29, 139–162. [Google Scholar] [CrossRef]
- Scaffidi, P.; Misteli, T.; Bianchi, M.E. Release of chromatin protein hmgb1 by necrotic cells triggers inflammation. Nature 2002, 418, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Gardella, S.; Andrei, C.; Ferrera, D.; Lotti, L.V.; Torrisi, M.R.; Bianchi, M.E.; Rubartelli, A. The nuclear protein hmgb1 is secreted by monocytes via a non-classical, vesicle-mediated secretory pathway. EMBO Rep. 2002, 3, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Stott, K.; Tang, G.S.; Lee, K.B.; Thomas, J.O. Structure of a complex of tandem hmg boxes and DNA. J. Mol. Biol. 2006, 360, 90–104. [Google Scholar] [CrossRef] [PubMed]
- Kokkola, R.; Andersson, A.; Mullins, G.; Ostberg, T.; Treutiger, C.J.; Arnold, B.; Nawroth, P.; Andersson, U.; Harris, R.A.; Harris, H.E. Rage is the major receptor for the proinflammatory activity of hmgb1 in rodent macrophages. Scand. J. Immunol. 2005, 61, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Avalos, A.M.; Mao, S.Y.; Chen, B.; Senthil, K.; Wu, H.; Parroche, P.; Drabic, S.; Golenbock, D.; Sirois, C.; et al. Toll-like receptor 9-dependent activation by DNA-containing immune complexes is mediated by hmgb1 and rage. Nat. Immunol. 2007, 8, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Zurolo, E.; Iyer, A.; Maroso, M.; Carbonell, C.; Anink, J.J.; Ravizza, T.; Fluiter, K.; Spliet, W.G.; van Rijen, P.C.; Vezzani, A.; et al. Activation of toll-like receptor, rage and hmgb1 signalling in malformations of cortical development. Brain 2011, 134, 1015–1032. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Evans, J.E.; Rock, K.L. Molecular identification of a danger signal that alerts the immune system to dying cells. Nature 2003, 425, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Antoine, D.J.; Jenkins, R.E.; Dear, J.W.; Williams, D.P.; McGill, M.R.; Sharpe, M.R.; Craig, D.G.; Simpson, K.J.; Jaeschke, H.; Park, B.K. Molecular forms of hmgb1 and keratin-18 as mechanistic biomarkers for mode of cell death and prognosis during clinical acetaminophen hepatotoxicity. J. Hepatol. 2012, 56, 1070–1079. [Google Scholar] [CrossRef]
- Balkwill, F. Tumour necrosis factor and cancer. Nat. Rev. Cancer 2009, 9, 361–371. [Google Scholar] [CrossRef]
- Green, D.R. The end and after: How dying cells impact the living organism. Immunity 2011, 35, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Masunaga, S.; Ono, K.; Takahashi, A.; Sakurai, Y.; Ohnishi, K.; Kobayashi, T.; Kinashi, Y.; Takagaki, M.; Ohnishi, T. Impact of the p53 status of the tumor cells on the effect of reactor neutron beam irradiation, with emphasis on the response of intratumor quiescent cells. Jpn. J. Cancer Res. 2002, 93, 1366–1377. [Google Scholar] [CrossRef] [PubMed]
- Kinashi, Y.; Takahashi, S.; Kashino, G.; Okayasu, R.; Masunaga, S.; Suzuki, M.; Ono, K. DNA double-strand break induction in ku80-deficient cho cells following boron neutron capture reaction. Radiat. Oncol. 2011, 6, 106. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, K.; Suzuki, A.; Mori, Y.; Kakihana, H.; Honda, C.; Mishima, Y.; Kobayashi, T.; Kanda, K. Improvement of solubility of p-boronophenylalanine by complex formation with monosaccharides. Strahlenther. Onkol. Organ Der Dtsch. Rontgenges. 1989, 165, 127–129. [Google Scholar]
- Fujihara, H.; Nozaki, T.; Tsutsumi, M.; Isumi, M.; Shimoda, S.; Hamada, Y.; Masutani, M. Spontaneous development of dental dysplasia in aged parp-1 knockout mice. Cells 2019, 8, 1157. [Google Scholar] [CrossRef]
- Madaan, A.; Verma, R.; Singh, A.; Jain, S.; Jaggi, M. A stepwise procedure for isolation of murine bone marrow and generation of dendritic cells. J. Biol. Methods 2014, 1, 1–6. [Google Scholar] [CrossRef]
- Ugrinova, I.; Pasheva, E. Hmgb1 protein: A therapeutic target inside and outside the cell. Adv. Protein Chem. Struct. Biol. 2017, 107, 37–76. [Google Scholar]
- Zandarashvili, L.; Sahu, D.; Lee, K.; Lee, Y.S.; Singh, P.; Rajarathnam, K.; Iwahara, J. Real-time kinetics of high-mobility group box 1 (hmgb1) oxidation in extracellular fluids studied by in situ protein nmr spectroscopy. J. Biol. Chem. 2013, 288, 11621–11627. [Google Scholar] [CrossRef]
- Schlueter, C.; Weber, H.; Meyer, B.; Rogalla, P.; Röser, K.; Hauke, S.; Bullerdiek, J. Angiogenetic signaling through hypoxia: Hmgb1: An angiogenetic switch molecule. Am. J. Pathol. 2005, 166, 1259–1263. [Google Scholar] [CrossRef]
- Withers, H.R.; Taylor, J.M.; Maciejewski, B. The hazard of accelerated tumor clonogen repopulation during radiotherapy. Acta Oncol. 1988, 27, 131–146. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Cheng, J.; Sun, L.; Wang, Y.; Wang, C.; Liu, X.; Zhang, Z.; Zhao, M.; Luo, Y.; Tian, L.; et al. Hmgb1 released by irradiated tumor cells promotes living tumor cell proliferation via paracrine effect. Cell Death Dis. 2018, 9, 648. [Google Scholar] [CrossRef] [PubMed]
- Takata, T.; Kondo, N.; Sakurai, Y.; Tanaka, H.; Hasegawa, T.; Kume, K.; Suzuki, M. Localized dose delivering by ion beam irradiation for experimental trial of establishing brain necrosis model. Appl. Radiat. Isot. 2015, 105, 32–34. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.D.; Agostinis, P. Cell death and immunity in cancer: From danger signals to mimicry of pathogen defense responses. Immunol. Rev. 2017, 280, 126–148. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.E. Hmgb1 loves company. J. Leukoc. Biol. 2009, 86, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Minsart, C.; Liefferinckx, C.; Lemmers, A.; Dressen, C.; Quertinmont, E.; Leclercq, I.; Deviere, J.; Moreau, R.; Gustot, T. New insights in acetaminophen toxicity: Hmgb1 contributes by itself to amplify hepatocyte necrosis in vitro through the tlr4-trif-ripk3 axis. Sci. Rep. 2020, 10, 5557. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, M.; Zhou, L.; Feng, X.; Cheng, J.; Yu, Y.; Gong, Y.; Zhu, Y.; Li, C.; Tian, L.; et al. Increased hmgb1 and cleaved caspase-3 stimulate the proliferation of tumor cells and are correlated with the poor prognosis in colorectal cancer. J. Exp. Clin. Cancer Res. 2015, 34, 51. [Google Scholar] [CrossRef] [PubMed]
- Lv, D.J.; Song, X.L.; Huang, B.; Yu, Y.Z.; Shu, F.P.; Wang, C.; Chen, H.; Zhang, H.B.; Zhao, S.C. Hmgb1 promotes prostate cancer development and metastasis by interacting with brahma-related gene 1 and activating the akt signaling pathway. Theranostics 2019, 9, 5166–5182. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imamichi, S.; Chen, L.; Ito, T.; Tong, Y.; Onodera, T.; Sasaki, Y.; Nakamura, S.; Mauri, P.; Sanada, Y.; Igaki, H.; et al. Extracellular Release of HMGB1 as an Early Potential Biomarker for the Therapeutic Response in a Xenograft Model of Boron Neutron Capture Therapy. Biology 2022, 11, 420. https://doi.org/10.3390/biology11030420
Imamichi S, Chen L, Ito T, Tong Y, Onodera T, Sasaki Y, Nakamura S, Mauri P, Sanada Y, Igaki H, et al. Extracellular Release of HMGB1 as an Early Potential Biomarker for the Therapeutic Response in a Xenograft Model of Boron Neutron Capture Therapy. Biology. 2022; 11(3):420. https://doi.org/10.3390/biology11030420
Chicago/Turabian StyleImamichi, Shoji, Lichao Chen, Tasuku Ito, Ying Tong, Takae Onodera, Yuka Sasaki, Satoshi Nakamura, PierLuigi Mauri, Yu Sanada, Hiroshi Igaki, and et al. 2022. "Extracellular Release of HMGB1 as an Early Potential Biomarker for the Therapeutic Response in a Xenograft Model of Boron Neutron Capture Therapy" Biology 11, no. 3: 420. https://doi.org/10.3390/biology11030420