
Anti-Osteoporotic Effect of Lactobacillus brevis AR281 in an Ovariectomized Mouse Model Mediated by Inhibition of Osteoclast Differentiation


Abstract
:Simple Summary
Abstract
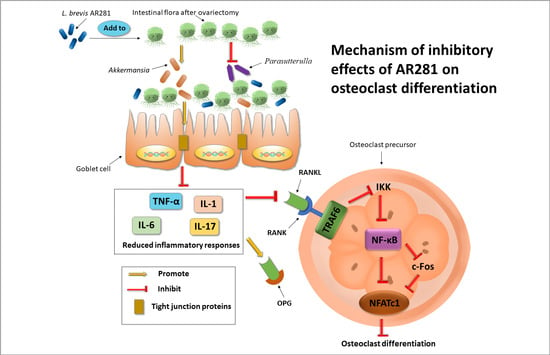
1. Introduction
2. Materials and Methods
2.1. Animal Study
2.2. Detection of Biochemical Parameters, Fluorescein Isothiocyanate-Dextran and Endotoxin
2.3. Micro-Computed Tomography and Biomechanical Detection
2.4. Cell Culture and Induction
2.5. TRAP Staining
2.6. Cell Counting Kit-8 Assay
2.7. RNA Extraction from Small Intestine, Tibia, Fecal Samples and RAW264.7 Cells
2.8. Real-Time Quantitative Polymerase Chain Reaction, Mus Primers, and High-Throughput 16SrDNA Sequencing
2.9. Statistics
3. Results
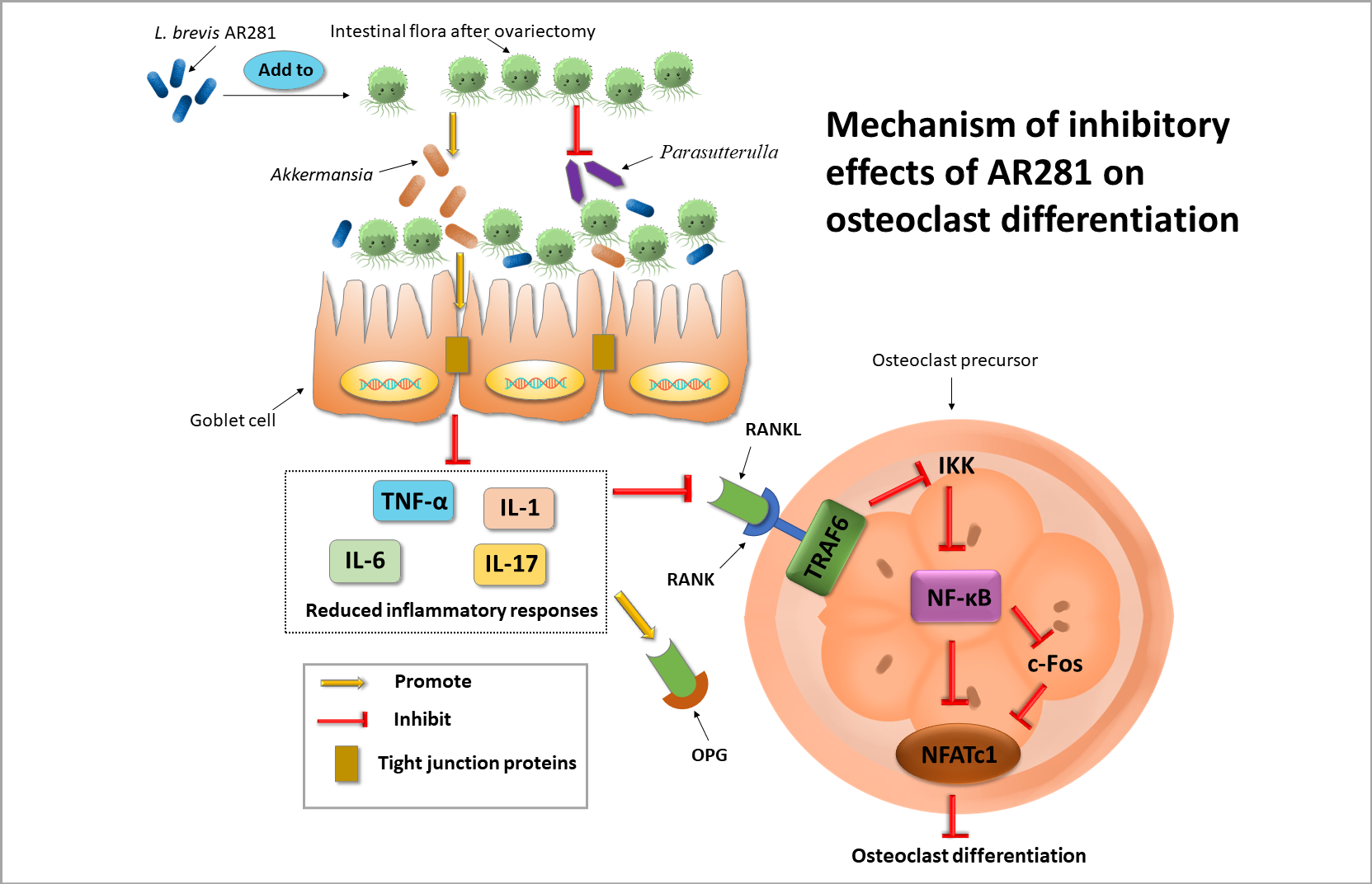
3.1. Effects of L. brevis AR281 on Skeletal Parameters and Biomechanics
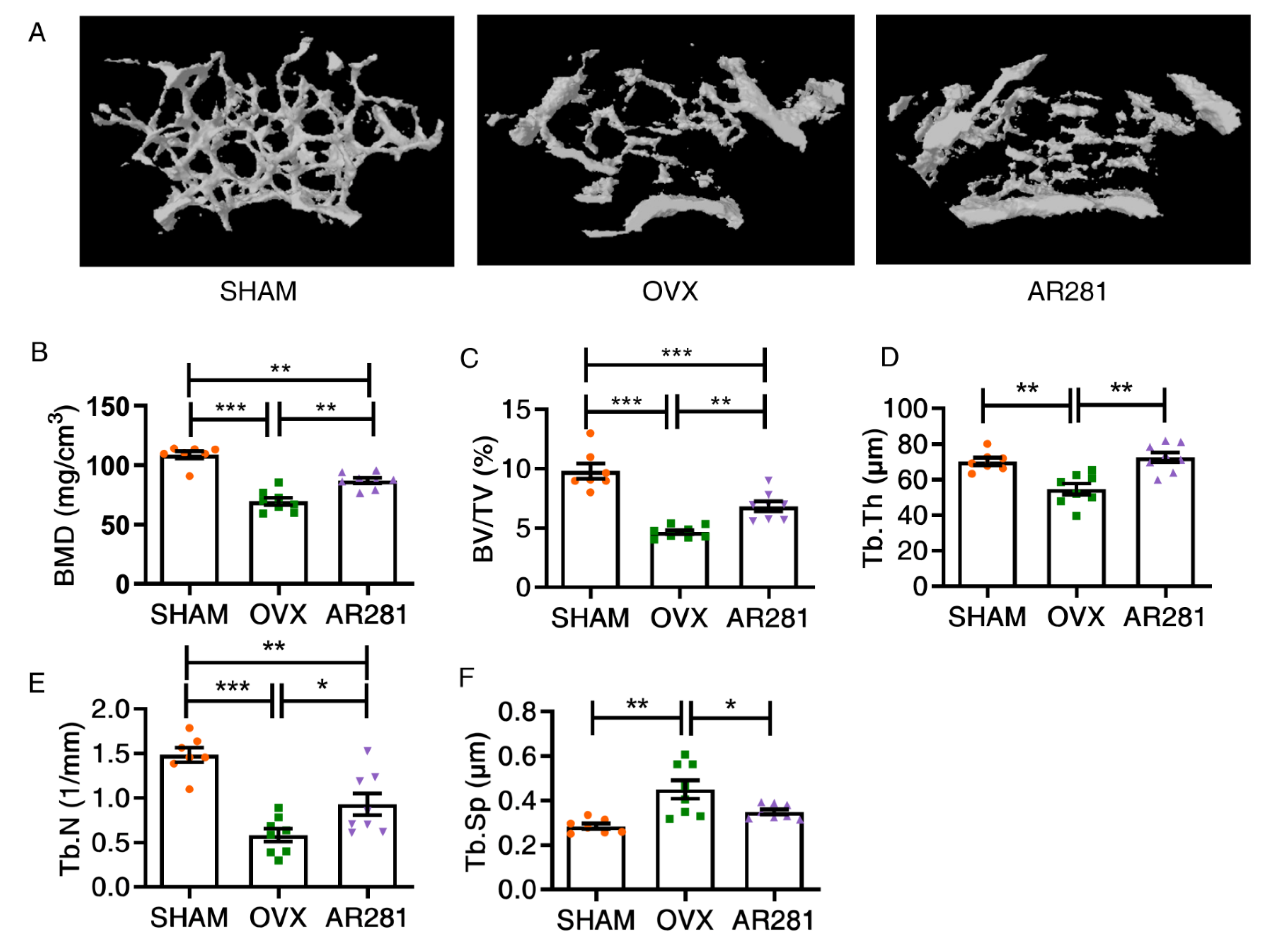
3.2. Effects of L. brevis AR281 on the Integrity of Gut Epithelial Barrier
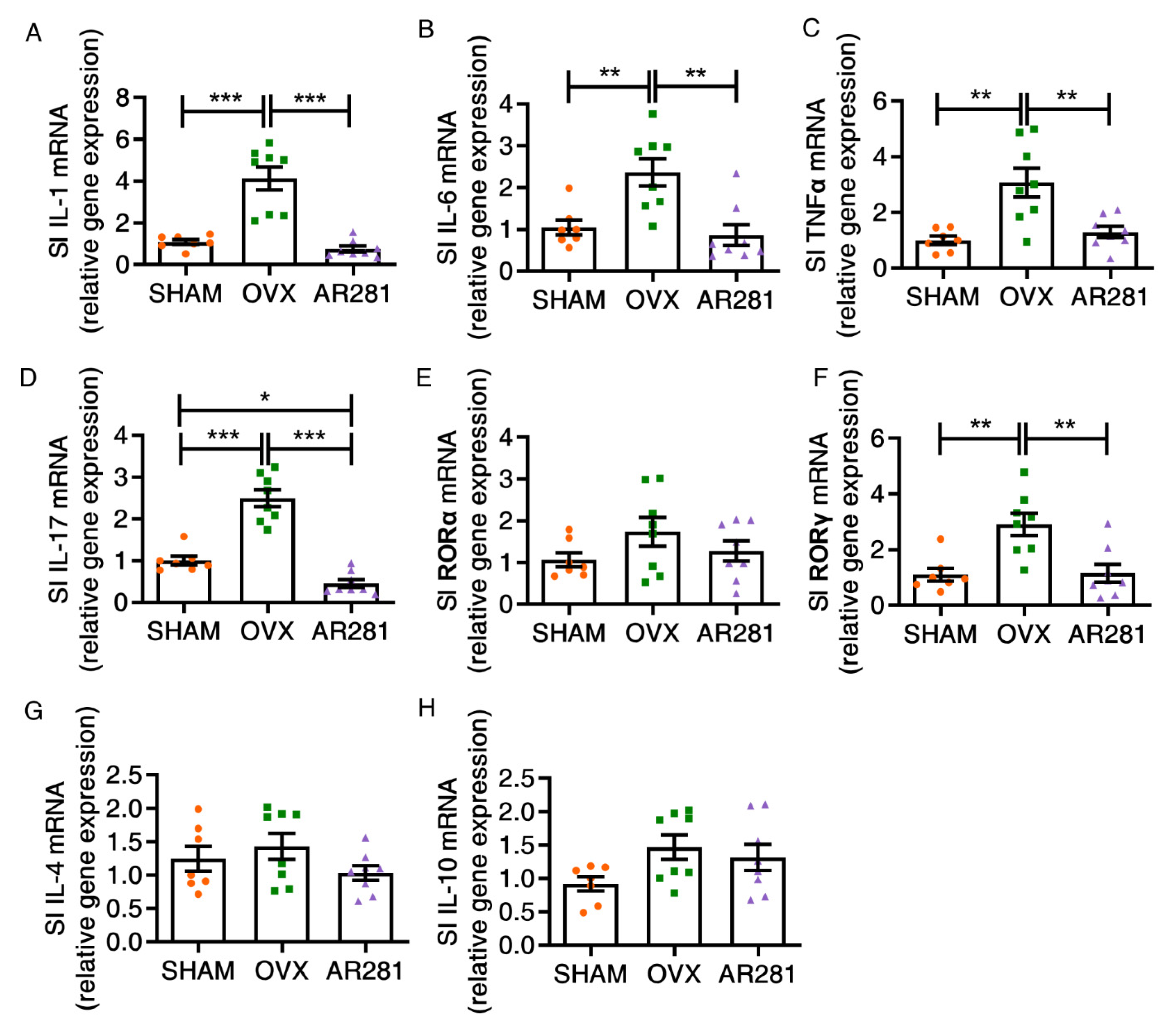
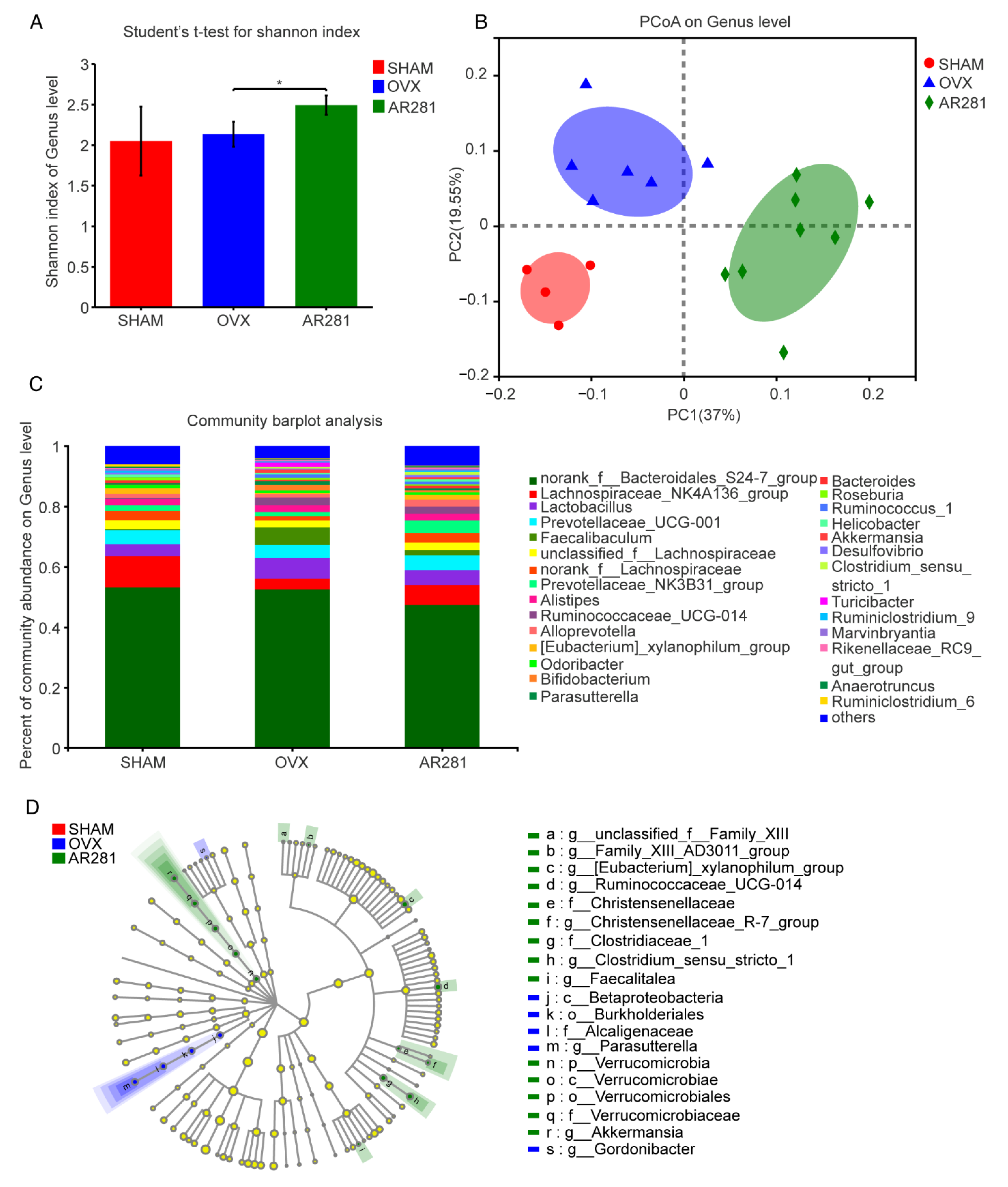
3.3. Effects of L. brevis AR281 on Intestinal Inflammatory Responses and Intestinal Flora
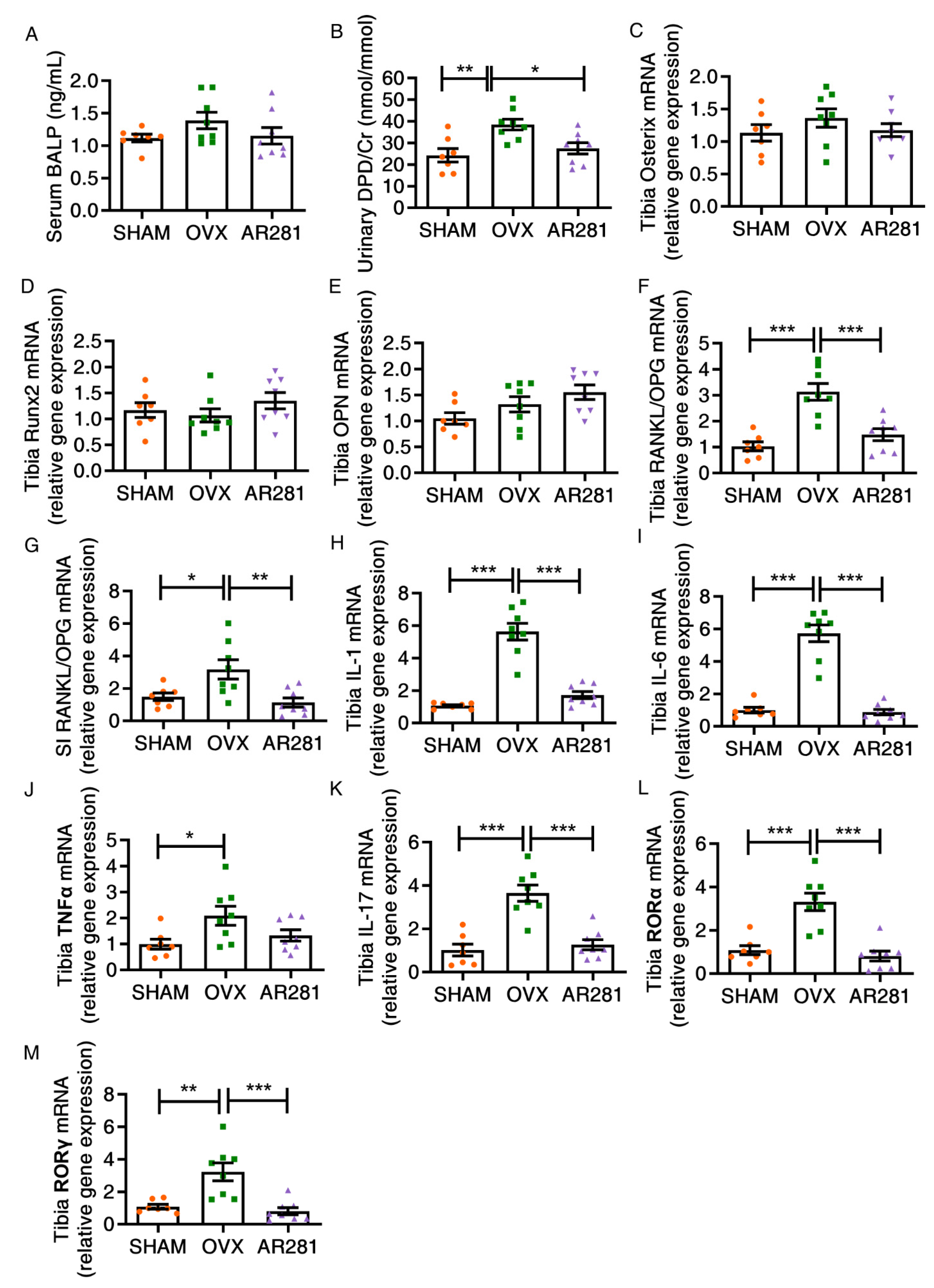
3.4. Effects of L. brevis AR281 on Bone Metabolism
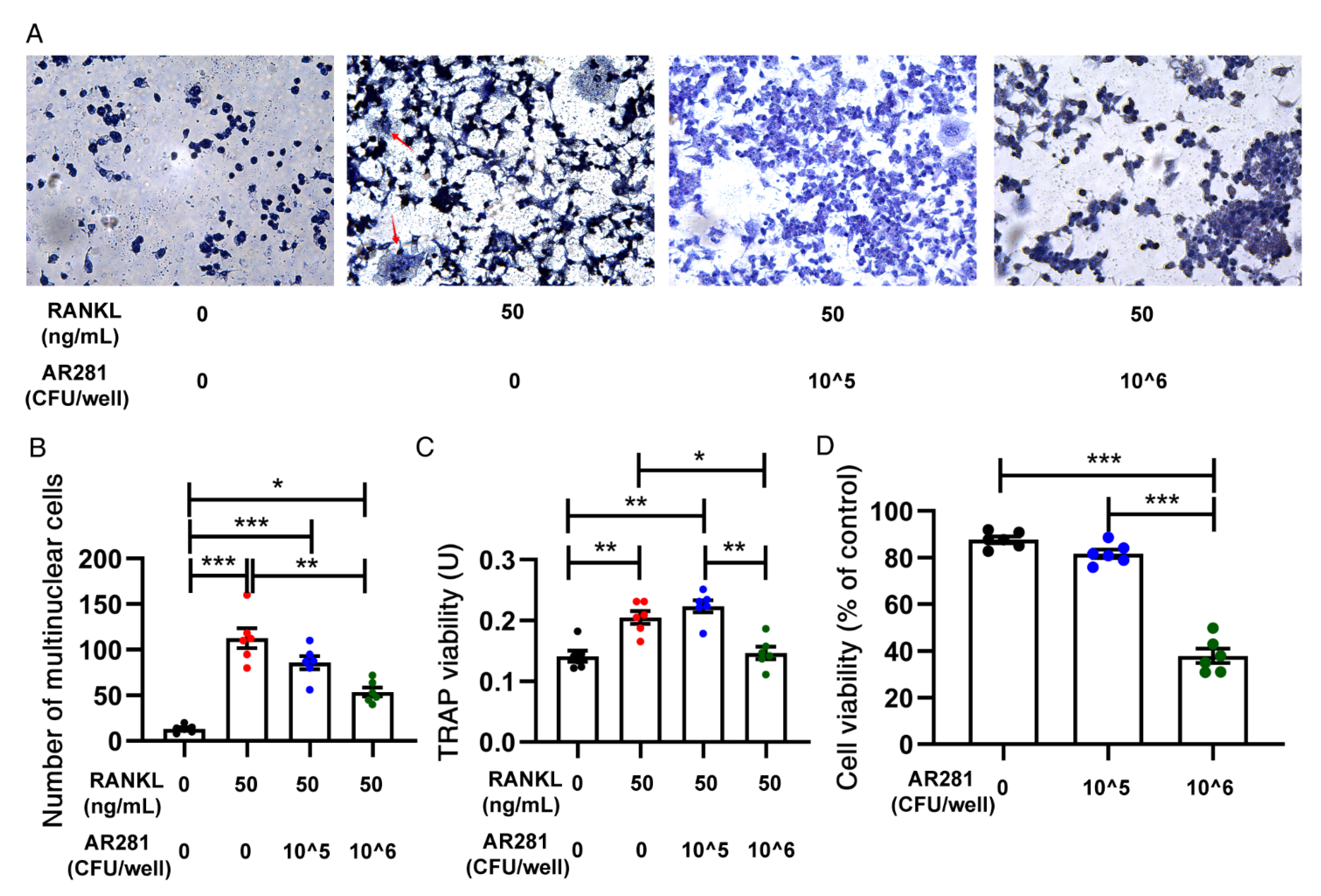
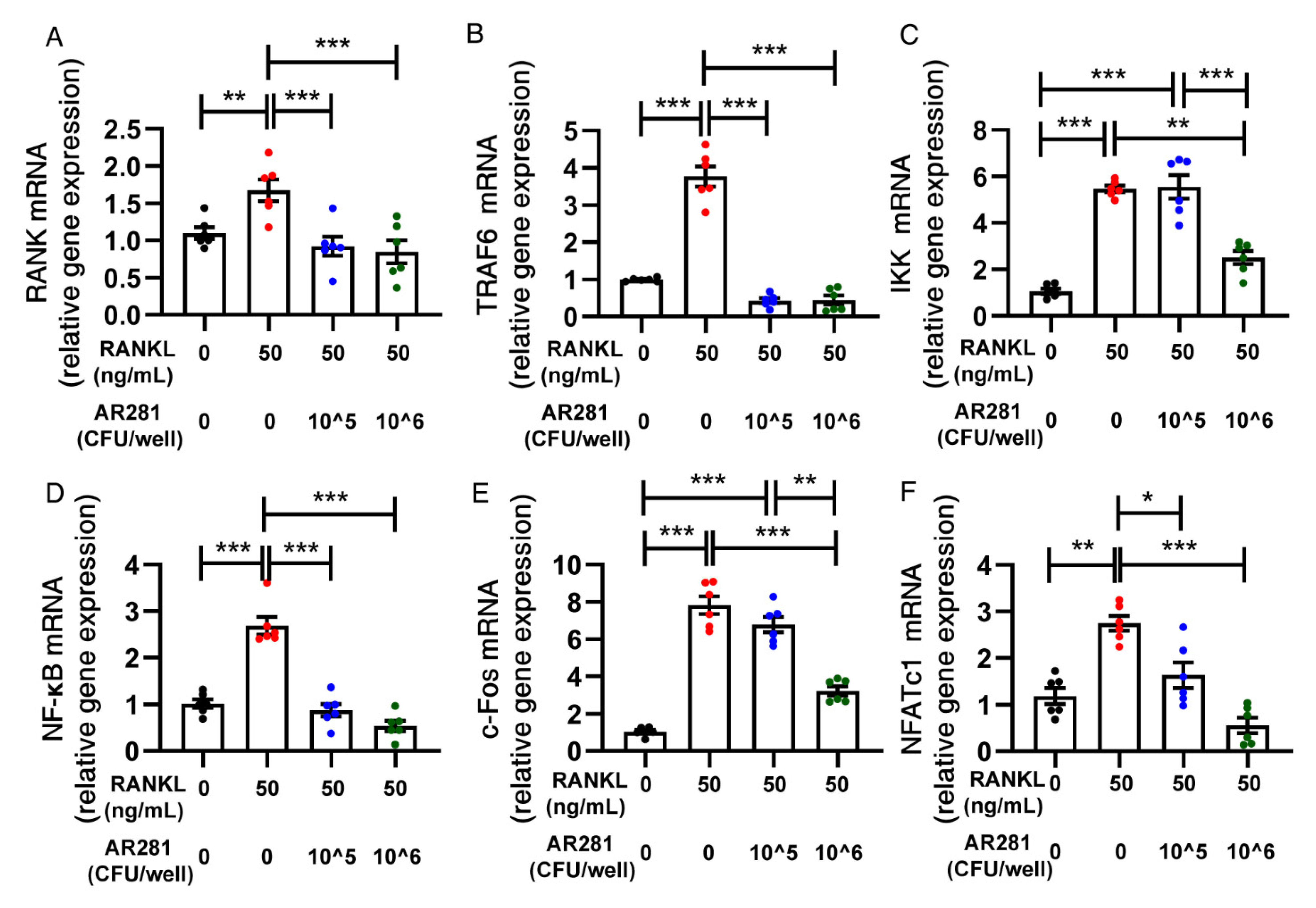
3.5. Effects of AR281 on RANKL-Induced Osteoclastogenesis in RAW264.7 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salt, R. Strontium Ranelate Treatment and Prevention of Osteoporosis, Bone Resorption Inhibitor, Bone Formation Stimulant. Drugs Future 2003, 28, 328–335. [Google Scholar]
- Rinonapoli, G.; Ruggiero, C.; Meccariello, L.; Bisaccia, M.; Caraffa, A. Osteoporosis in Men: A Review of an Underestimated Bone Condition. Int. J. Mol. Sci. 2021, 22, 2105. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.M.J.; Delaissé, J.M.; Olesen, J.B.; Madsen, J.S.; Canto, L.M.; Bechmann, T.; Rogatto, S.R.; Søe, K. Aging and Menopause Reprogram Osteoclast Precursors for Aggressive Bone Resorption. Bone Res. 2020, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, R.; Hart, D.M.; Forrest, C.; Baird, C. Prevention of Spinal Osteoporosis in Oophorectomised Women. Lancet 1980, 2, 1151–1154. [Google Scholar] [CrossRef]
- Rossouw, J.E.; Prentice, R.L.; Manson, J.E.; Wu, L.; Barad, D.; Barnabei, V.M.; Ko, M.; LaCroix, A.Z.; Margolis, K.L.; Stefanick, M.L. Postmenopausal Hormone Therapy and Risk of Cardiovascular Disease by Age and Years since Menopause. JAMA 2007, 297, 1465–1477. [Google Scholar] [CrossRef]
- Di Donato, V.; Palaia, I.; D’Aniello, D.; Musacchio, L.; Santangelo, G.; Pinto, A.D.; Musella, A.; Fischetti, M.; Tomao, F.; Perniola, G. Does Hormone Replacement Therapy Impact the Prognosis in Endometrial Cancer Survivors? A Systematic Review. Oncology 2020, 98, 195–201. [Google Scholar] [CrossRef]
- Vinogradova, Y.; Coupland, C.; Hippisley-Cox, J. Use of Hormone Replacement Therapy and Risk of Breast Cancer: Nested Case-Control Studies Using the Qresearch and Cprd Databases. BMJ 2020, 371, m3873. [Google Scholar] [CrossRef]
- Abrahamsen, B.; Einhorn, T.A. Beyond a Reasonable Doubt? Bisphosphonates and Atypical Femur Fractures. Bone 2012, 50, 1196–1200. [Google Scholar] [CrossRef]
- Rizzoli, R.; Reginster, J.Y.; Boonen, S.; Bréart, G.; Diez-Perez, A.; Felsenberg, D.; Kaufman, J.M.; Kanis, J.A.; Cooper, C. Adverse Reactions and Drug-Drug Interactions in the Management of Women with Postmenopausal Osteoporosis. Calcif. Tissue Int. 2011, 89, 91–104. [Google Scholar] [CrossRef] [Green Version]
- Gennari, L.; Merlotti, D.; Stolakis, K.; Nuti, R. Lasofoxifene, from the Preclinical Drug Discovery to the Treatment of Postmenopausal Osteoporosis. Expert Opin. Drug Discov. 2011, 6, 205–217. [Google Scholar] [CrossRef]
- McCabe, L.R.; Irwin, R.; Schaefer, L.; Britton, R.A. Probiotic Use Decreases Intestinal Inflammation and Increases Bone Density in Healthy Male but Not Female Mice. J. Cell Physiol. 2013, 228, 1793–1798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.S.; Zhao, X.; Yang, Y. Effects of Emodin on Inflammatory Bowel Disease-Related Osteoporosis. Biosci. Rep. 2020, 40, BSR20192317. [Google Scholar] [CrossRef] [PubMed]
- Chedid, V.G.; Kane, S.V. Bone Health in Patients with Inflammatory Bowel Diseases. J. Clin. Densitom. 2020, 23, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Masip, E.; Donat, E.; Miquel, B.P.; Ribes-Koninckx, C. Bone Mineral Density in Spanish Children at the Diagnosis of Inflammatory Bowel Disease. Arch. Osteoporos. 2021, 16, 96. [Google Scholar] [CrossRef] [PubMed]
- Greendale, G.A.; Jackson, N.J.; Han, W.; Huang, M.; Cauley, J.A.; Karvonen-Gutierrez, C.; Karlamangla, A.S. Increase in C-Reactive Protein Predicts Increase in Rate of Bone Mineral Density Loss: The Study of Women’s Health across the Nation. JBMR Plus 2021, 5, e10480. [Google Scholar] [CrossRef]
- Mun, H.; Liu, B.; Pham, T.H.A.; Wu, Q. C-Reactive Protein and Fracture Risk: An Updated Systematic Review and Meta-Analysis of Cohort Studies through the Use of Both Frequentist and Bayesian Approaches. Osteoporos. Int. 2021, 32, 425–435. [Google Scholar] [CrossRef]
- Brüünsgaard, H.; Pedersen, B.K. Age-Related Inflammatory Cytokines and Disease. Immunol. Allergy Clin. N. Am. 2003, 23, 15–39. [Google Scholar] [CrossRef]
- Bruunsgaard, H. Effects of Tumor Necrosis Factor-Alpha and Interleukin-6 in Elderly Populations. Eur. Cytokine Netw. 2002, 13, 389–391. [Google Scholar]
- Nanes, M.S. Tumor Necrosis Factor-Alpha: Molecular and Cellular Mechanisms in Skeletal Pathology. Gene 2003, 321, 1–15. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Kim, K.; Kim, H. The Potential Impact of Salivary Il-1 on the Diagnosis of Periodontal Disease: A Pilot Study. Healthcare 2021, 9, 729. [Google Scholar] [CrossRef]
- Ansari, M.A.; Nadeem, A.; Bakheet, S.A.; Attia, S.M.; Ahmad, S.F. Chemokine Receptor 5 Antagonism Causes Reduction in Joint Inflammation in a Collagen-Induced Arthritis Mouse Model. Molecules 2021, 26, 1839. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Jiang, S.; Dong, Q.; Dang, J.; Liu, Z.; Han, H.; Tao, Y.; Yue, H. Anti-Rheumatoid Arthritis Effects of Iridoid Glucosides from Lamiophlomis rotata (Benth.) Kudo on Adjuvant-Induced Arthritis in Rats by Opg/Rankl/Nf-Κb Signaling Pathways. J. Ethnopharmacol. 2021, 266, 113402. [Google Scholar] [CrossRef] [PubMed]
- Weitzmann, M.N.; Ofotokun, I. Physiological and Pathophysiological Bone Turnover—Role of the Immune System. Nat. Rev. Endocrinol. 2016, 12, 518–532. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-Y.; Chassaing, B.; Tyagi, A.M.; Vaccaro, C.; Luo, T.; Adams, J.; Darby, T.M.; Weitzmann, M.N.; Mulle, J.G.; Gewirtz, A.T.; et al. Sex Steroid Deficiency–Associated Bone Loss Is Microbiota Dependent and Prevented by Probiotics. J. Clin. Investig. 2016, 126, 2049–2063. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Zhan, S.; Tian, Z.; Liu, C.; Xie, Z.; Zhang, S.; Chen, M.; Zeng, Z.; Zhuang, X. Alterations in Bile Acid Metabolism Associated with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2021, 27, 1525–1540. [Google Scholar] [CrossRef]
- Zaiss, M.M.; Rapin, A.; Lebon, L.; Dubey, L.K.; Harris, N.L. The Intestinal Microbiota Contributes to the Ability of Helminths to Modulate Allergic Inflammation. Immunity 2015, 43, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Aron-Wisnewsky, J.; Warmbrunn, M.V.; Nieuwdorp, M.; Clément, K. Metabolism and Metabolic Disorders and the Microbiome: The Intestinal Microbiota Associated with Obesity, Lipid Metabolism, and Metabolic Health—Pathophysiology and Therapeutic Strategies. Gastroenterology 2021, 160, 573–599. [Google Scholar] [CrossRef]
- Alpízar-Rodríguez, D.; Finckh, A.; Gilbert, B. The Role of Nutritional Factors and Intestinal Microbiota in Rheumatoid Arthritis Development. Nutrients 2021, 13, 96. [Google Scholar] [CrossRef]
- Hang, Y.; Yu, J.; Lin, X.; Zhou, W.; Xia, Y.; Wang, G.; Zhang, H.; Xiong, Z.; Ai, L. Screening and Identification of Probiotics with Improvement Effects of Calcium and Phosphorous Absorption on Ovariectomized Mice. Ind. Microbiol. 2018, 48, 12–17. [Google Scholar]
- Feyereisen, M.; Mahony, J.; Lugli, G.A.; Ventura, M.; Neve, H.; Franz, C.; Noben, J.P.; O’Sullivan, T.; Sinderen, D.V. Isolation and Characterization of Lactobacillus brevis Phages. Viruses 2019, 11, 393. [Google Scholar] [CrossRef] [Green Version]
- Rönkä, E.; Malinen, E.; Saarela, M.; Rinta-Koski, M.; Aarnikunnas, J.; Palva, A. Probiotic and Milk Technological Properties of Lactobacillus brevis. Int. J. Food Microbiol. 2003, 83, 63–74. [Google Scholar] [CrossRef]
- Li, Y.; Sun, Y.; Xu, X.; Fang, C. Antimicrobial Activity Characteristic on Cell-Free Broth of Potential Lactobacillus Brevis P-319 Isolated from Sauerkraut. In Proceedings of the 2011 International Conference on Remote Sensing, Environment and Transportation Engineering, Nanjing, China, 24–26 June 2011; pp. 8065–8068. [Google Scholar]
- Yu, J.; Xia, Y.; Wang, G.; Xiong, Z.; Zhang, H.; Lai, P.F.-H.; Song, X.; Ai, L. Anti-Osteoporotic Potential of Lactobacillus Plantarum Ar237 and Ar495 in Ovariectomized Mice. J. Funct. Foods 2021, 87, 104762. [Google Scholar] [CrossRef]
- Jämsä, T.; Jalovaara, P.; Peng, Z.; Väänänen, H.K.; Tuukkanen, J. Comparison of Three-Point Bending Test and Peripheral Quantitative Computed Tomography Analysis in the Evaluation of the Strength of Mouse Femur and Tibia. Bone 1998, 23, 155–161. [Google Scholar] [CrossRef]
- Ueta, M.; Takaoka, K.; Yamamura, M.; Maeda, H.; Tamaoka, J.; Nakano, Y.; Noguchi, K.; Kishimoto, H. Effects of Tgf-Β1 on the Migration and Morphology of Raw264. 7 Cells in Vitro. Mol. Med. Rep. 2019, 20, 4331–4339. [Google Scholar] [PubMed]
- Goel, R.; Ormond, M.; Hullah, E.; Nayee, S.; Escudier, M.; Sanderson, J. Pth-067an Evaluation Study of Lactobacillus brevis Cd2 in Orofacial Granuolomatosis. Gut 2015, 64, A435–A436. [Google Scholar] [CrossRef]
- Takauji, S.; Konishi, H.; Fujiya, M.; Ueno, N.; Okumura, T. Polyphosphate, Derived from Lactobacillus brevis, Modulates the Intestinal Microbiome and Attenuates Acute Pancreatitis. Dig. Dis. Sci. 2021, 66, 3872–3884. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, T.; Hajishengallis, G. Topical Treatment with Probiotic Lactobacillus Brevis Cd2 Inhibits Experimental Periodontal Inflammation and Bone Loss. J. Periodontal Res. 2015, 49, 785–791. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, J. From the Gut to Bone: Connecting the Gut Microbiota with Th17 T Lymphocytes and Postmenopausal Osteoporosis. J. Clin. Investig. 2021, 131, e146619. [Google Scholar] [CrossRef]
- Chiodini, R.J.; Dowd, S.E.; Chamberlin, W.M.; Susan, G.; Brian, D.; Angela, G.; Hauke, S. Microbial Population Differentials between Mucosal and Submucosal Intestinal Tissues in Advanced Crohn’s Disease of the Ileum. PLoS ONE 2015, 10, e0134382. [Google Scholar] [CrossRef]
- Hu, J.; Oscar, F.; Pei, Z.; Steven, I.; Inga, P. P-237 Multiple Double-Barcoding 16s Sequencing on the Miseq Platform to Study the Gut Microbiome in Ashkenazi Jews with Crohn’s Disease. Inflamm. Bowel Dis. 2013, 19, S119–S121. [Google Scholar] [CrossRef]
- Becken, B.; Davey, L.; Middleton, D.R.; Mueller, K.D.; Sharma, A.; Holmes, Z.C.; Dallow, E.; Remick, B.; Barton, G.M.; David, L.A. Genotypic and Phenotypic Diversity among Human Isolates of Akkermansia muciniphila. mBio 2021, 12, e00478-21. [Google Scholar] [CrossRef] [PubMed]
- Grondin, J.; Wang, H.; Haq, S.; Kwon, E.Y.; Surette, M.; Khan, W.I. A219 Protective Effects of Akkermansia Muciniphila on Intestinal Barrier Function and Inflammation. J. Can. Assoc. Gastroenterol. 2020, 3, 93. [Google Scholar] [CrossRef] [Green Version]
- Xi, M.; Li, J.; Hao, G.; An, X.; Song, Y.; Wei, H.; Ge, W. Stachyose Increases Intestinal Barrier through Akkermansia Muciniphila and Reduces Gut Inflammation in Germ-Free Mice after Human Fecal Transplantation. Food Res. Int. 2020, 137, 109288. [Google Scholar] [CrossRef] [PubMed]
- Zhai, R.; Xue, X.; Zhang, L.; Yang, X.; Zhao, L.; Zhang, C. Strain-Specific Anti-Inflammatory Properties of Two Akkermansia Muciniphila Strains on Chronic Colitis in Mice. Front. Cell. Infect. Microbiol. 2019, 9, 239. [Google Scholar] [CrossRef]
- Yang, Y.; Zhong, Z.; Wang, B.; Xia, X.; Yao, W.; Huang, L.; Wang, Y.; Ding, W. Early-Life High-Fat Diet-Induced Obesity Programs Hippocampal Development and Cognitive Functions Via Regulation of Gut Commensal Akkermansia Muciniphila. Neuropsychopharmacology 2019, 44, 2054–2064. [Google Scholar] [CrossRef]
- Yasuda, H. Discovery of the Rankl/Rank/Opg System. J. Bone Miner. Metab. 2021, 39, 2–11. [Google Scholar] [CrossRef]
- Liu, H.; Gu, R.; Li, W.; Zhou, W.; Cong, Z.; Xue, J.; Liu, Y.; Wei, Q.; Zhou, Y. Lactobacillus Rhamnosus Gg Attenuates Tenofovir Disoproxil Fumarate-Induced Bone Loss in Male Mice Via Gut-Microbiota-Dependent Anti-Inflammation. Ther. Adv. Chronic Dis. 2019, 10, 2040622319860653. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.S.; Kim, J.-Y.; Kim, B.K.; Lee, I.O.; Park, N.H.; Kim, S.H. Lactobacillus-Fermented Milk Products Attenuate Bone Loss in an Experimental Rat Model of Ovariectomy-Induced Post-Menopausal Primary Osteoporosis. J. Appl. Microbiol. 2021, 130, 2041–2062. [Google Scholar] [CrossRef]
- Giehler, F.; Ostertag, M.; Sommermann, T.; Weidl, D.; Sterz, K.; Kutz, H.; Feller, S.; Geerlof, A.; Biesinger, B.; Popowicz, G. Epstein-Barr Virus-Driven B Cell Lymphoma Mediated by a Unique Lmp1-Traf6 Complex. Res. Sq. 2022, 1. [Google Scholar] [CrossRef]
- Fang, J.; Muto, T.; Kleppe, M.; Bolanos, L.C.; Hueneman, K.M.; Walker, C.S.; Sampson, L.; Wellendorf, A.M.; Chetal, K.; Choi, K.; et al. Traf6 Mediates Basal Activation of Nf-Κb Necessary for Hematopoietic Stem Cell Homeostasis. Cell Rep. 2018, 22, 1250–1262. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.; Kim, J.H.; Kim, K.; Seong, S.; Lee, K.; Kim, N. Irf2 Enhances Rankl-Induced Osteoclast Differentiation Via Regulating Nf-Κb/Nfatc1 Signaling. BMB Rep. 2021, 54, 482. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Lee, H.; Ha, I.J.; Yang, W.M. Zanthoxylum Piperitum Alleviates the Bone Loss in Osteoporosis Via Inhibition of Rankl-Induced C-Fos/Nfatc1/Nf-Κb Pathway. Phytomedicine 2021, 80, 153397. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Kim, I.S.; Lim, B.O. Black Rice (Oryza sativa L.) Fermented with Lactobacillus Casei Attenuates Osteoclastogenesis and Ovariectomy-Induced Osteoporosis. BioMed Res. Int. 2019, 2019, 5073085. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bending Stress (MPa) | Bending Load (N) | Young’s Bending Modulus (MPa) | |
|---|---|---|---|
| SHAM | 357.84 ± 22.65 | 14.05 ± 0.89 | 29,834.95 ± 1888.83 * |
| OVX | 338.75 ± 13.92 | 13.30 ± 0.55 | 25,263.29 ± 2572.43 |
| AR281 | 378.38 ± 23.64 * | 14.86 ± 0.93 | 31,743.22 ± 3073.48 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Hang, Y.; Sun, W.; Wang, G.; Xiong, Z.; Ai, L.; Xia, Y. Anti-Osteoporotic Effect of Lactobacillus brevis AR281 in an Ovariectomized Mouse Model Mediated by Inhibition of Osteoclast Differentiation. Biology 2022, 11, 359. https://doi.org/10.3390/biology11030359
Yu J, Hang Y, Sun W, Wang G, Xiong Z, Ai L, Xia Y. Anti-Osteoporotic Effect of Lactobacillus brevis AR281 in an Ovariectomized Mouse Model Mediated by Inhibition of Osteoclast Differentiation. Biology. 2022; 11(3):359. https://doi.org/10.3390/biology11030359
Chicago/Turabian StyleYu, Jing, Yiling Hang, Wenni Sun, Guangqiang Wang, Zhiqiang Xiong, Lianzhong Ai, and Yongjun Xia. 2022. "Anti-Osteoporotic Effect of Lactobacillus brevis AR281 in an Ovariectomized Mouse Model Mediated by Inhibition of Osteoclast Differentiation" Biology 11, no. 3: 359. https://doi.org/10.3390/biology11030359