
Role of Descending Serotonergic Fibers in the Development of Pathophysiology after Spinal Cord Injury (SCI): Contribution to Chronic Pain, Spasticity, and Autonomic Dysreflexia



Abstract
:Simple Summary
Abstract
1. Introduction
2. Serotonin Function in the Uninjured Nervous System
2.1. Overview of Descending Pathways and Their Function
2.1.1. Regulation of Sensory Processes and Pain
2.1.2. Regulation of Motor Behavior
2.1.3. Regulation of Autonomic Function
2.2. Overview of How 5-HT Affects Neural Function within the Spinal Cord
3. Impact of SCI on 5-HT Function
3.1. Impact of Injury on 5-HT Levels
3.2. Acute Effects of Impaired Serotonergic Activity
3.2.1. Descending Serotonergic Fibers Can Quell Nociceptive Sensitization
3.2.2. Only Uncontrollable Stimulation Induces Nociceptive Sensitization
3.2.3. Uncontrollable Stimulation Increases Tissue Loss and Impairs Recovery after a Contusion Injury
3.2.4. Descending 5-HT Fibers Help Preserve the Capacity to Learn
3.2.5. Behavioral Control and Brain-Derived Neurotrophic Factor (BDNF) Counter the Adverse Effects of Noxious Stimulation
3.3. Long-Term Effects of Impaired Serotonergic Activity
3.3.1. Damage to Serotonergic Pathways Promotes the Development of Neuropathic Pain
3.3.2. Damage to Serotonergic Pathways Fosters Spasticity
3.3.3. Damage to Serotonergic Pathways Fosters Autonomic Function
3.3.4. SCI Facilitates Pulvinar Reorganization and Dysfunction
4. Descending Serotonergic Fibers Regulate the Inhibitory Effect of GABA
4.1. Pretreatment with a GABA-A Antagonist Blocks the Development of Nociceptive Sensitization after SCI
4.2. Alterations in Intracellular Cl− Impact How GABA Affects Neural Activity
4.3. Exercise and Training Re-Establish GABAergic Inhibition after Injury
4.4. The Shift in GABA Function Is Tied to the Loss of Descending 5-HT Fibers
5. Role of Other Monoamines
5.1. Noradrenergic Fiber Pathways
5.2. Dopaminergic Fiber Pathways
6. Conclusions
6.1. Summary
6.2. Limitations and Issues for Future Research
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ben-Ari, Y.; Khalilov, I.; Kahle, K.T.; Cherubini, E. The GABA Excitatory/Inhibitory Shift in Brain Maturation and Neurological Disorders. Neuroscientist 2012, 18, 467–486. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Pearse, D.D. The role of the serotonergic system in locomotor recovery after spinal cord injury. Front. Neural Circuits 2014, 8, 151. [Google Scholar] [CrossRef] [PubMed]
- Hachoumi, L.; Sillar, K.T. Developmental stage-dependent switching in the neuromodulation of vertebrate locomotor central pattern generator networks. Dev. Neurobiol. 2020, 80, 42–57. [Google Scholar] [CrossRef] [PubMed]
- Perrin, F.E.; Noristani, H.N. Serotonergic mechanisms in spinal cord injury. Exp. Neurol. 2019, 318, 174–191. [Google Scholar] [CrossRef]
- Perrier, J.F.; Cotel, F. Serotonergic modulation of spinal motor control. Curr. Opin. Neurobiol. 2015, 33, 1–7. [Google Scholar] [CrossRef]
- Slawinska, U.; Miazga, K.; Jordan, L.M. The role of serotonin in the control of locomotor movements and strategies for restoring locomotion after spinal cord injury. Acta Neurobiol. Exp. 2014, 74, 172–187. [Google Scholar]
- Jordan, L.M.; Slawinska, U. Chapter 12—Modulation of rhythmic movement: Control of coordination. Prog. Brain Res. 2011, 188, 181–195. [Google Scholar] [CrossRef]
- Cummings, K.J.; Hodges, M.R. The serotonergic system and the control of breathing during development. Respir. Physiol. Neurobiol. 2019, 270, 103255. [Google Scholar] [CrossRef]
- Fields, D.P.; Mitchell, G.S. Spinal metaplasticity in respiratory motor control. Front. Neural Circuits 2015, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Bannister, K.; Dickenson, A.H. What do monoamines do in pain modulation? Curr. Opin. Supportive Palliat. Care 2017, 10, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Bardoni, R. Serotonergic Modulation of Nociceptive Circuits in Spinal Cord Dorsal Horn. Curr. Neuropharmacol. 2019, 17, 1133–1145. [Google Scholar] [CrossRef]
- Liu, Q.Q.; Yao, X.X.; Gao, S.H.; Li, R.; Li, B.J.; Yang, W.; Cui, R.J. Role of 5-HT receptors in neuropathic pain: Potential therapeutic implications. Pharmacol. Res. 2020, 159, 104949. [Google Scholar] [CrossRef]
- Dahlstroem, A.; Fuxe, K. Evidence for the existence of monoamine-containing neurons in the central nervous system. I. Demonstration of monoamines in the cell bodies of brain stem neurons. Acta Physiol. Scand. 1964, 62 (Suppl. S232), 231–255. [Google Scholar]
- Törk, I. Anatomy of the serotonergic system. Ann. N. Y. Acad. Sci. 1990, 600, 9–34. [Google Scholar] [CrossRef]
- Bowker, R.M.; Westlund, K.N.; Sullivan, M.C.; Coulter, J.D. Organization of descending serotonergic projections to the spinal cord. Prog. Brain Res. 1982, 57, 239–265. [Google Scholar] [CrossRef]
- Benarroch, E.E. Descending monoaminergic pain modulation: Bidirectional control and clinical relevance. Neurology 2008, 71, 217–221. [Google Scholar] [CrossRef]
- Millan, M.J. Descending control of pain. Prog. Neurobiol. 2002, 66, 355–474. [Google Scholar] [CrossRef]
- Eldahan, K.C.; Rabchevsky, A.G. Autonomic dysreflexia after spinal cord injury: Systemic pathophysiology and methods of management. Auton. Neurosci. 2018, 209, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Bowker, R.M.; Westlund, K.N.; Coulter, J.D. Origins of serotonergic projections to the spinal cord in rat: An immunocytochemical-retrograde transport study. Brain Res. 1981, 226, 187–199. [Google Scholar] [CrossRef]
- Bowker, R.M.; Westlund, K.N.; Sullivan, M.C.; Wilber, J.F.; Coulter, J.D. Descending serotonergic, peptidergic and cholinergic pathways from the raphe nuclei: A multiple transmitter complex. Brain Res. 1983, 288, 33–48. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Fields, H.L. Endogenous pain control systems: Brainstem spinal pathways and endorphin circuitry. Annu. Rev. Neurosci. 1984, 7, 309–338. [Google Scholar] [CrossRef]
- Stewart, W.; Maxwell, D.J. Morphological evidence for selective modulation by serotonin of a subpopulation of dorsal horn cells which possess the neurokinin-1 receptor. Eur. J. Neurosci. 2000, 12, 4583–4588. [Google Scholar]
- Millan, M.J. Endorphins and nociception: An overview. Methods Find. Exp. Clin. Pharmacol. 1982, 4, 445–462. [Google Scholar]
- Fields, H.L.; Basbaum, A.I. Brainstem control of spinal pain-transmission neurons. Annu. Rev. Physiol. 1978, 40, 217–248. [Google Scholar] [CrossRef]
- Zhuo, M.; Gebhart, G.F. Characterization of descending facilitation and inhibition of spinal nociceptive transmission from the nuclei reticularis gigantocellularis and gigantocellularis pars alpha in the rat. J. Neurophysiol. 1992, 67, 1599–1614. [Google Scholar] [CrossRef]
- Zhuo, M.; Gebhart, G.F. Spinal serotonin receptors mediate descending facilitation of a nociceptive reflex from the nuclei reticularis gigantocellularis and gigantocellularis pars alpha in the rat. Brain Res. 1991, 550, 35–48. [Google Scholar] [CrossRef]
- Zhuo, M.; Gebhart, G.F. Characterization of descending inhibition and facilitation from the nuclei reticularis gigantocellularis and gigantocellularis pars alpha in the rat. Pain 1990, 42, 337–350. [Google Scholar] [CrossRef]
- Zhuo, M.; Gebhart, G.F. Biphasic modulation of spinal nociceptive transmission from the medullary raphe nuclei in the rat. J. Neurophysiol. 1997, 78, 746–758. [Google Scholar] [CrossRef] [PubMed]
- Gebhart, G.F. Descending modulation of pain. Neurosci. Biobehav. Rev. 2004, 27, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Randich, A.; Gebhart, G.F. Vagal afferent modulation of a nociceptive reflex in rats: Involvement of spinal opioid and monoamine receptors. Brain Res. 1988, 446, 285–294. [Google Scholar] [CrossRef]
- Ren, K.; Randich, A.; Gebhart, G.F. Spinal serotonergic and kappa opioid receptors mediate facilitation of the tail flick reflex produced by vagal afferent stimulation. Pain 1991, 45, 321–329. [Google Scholar] [CrossRef]
- Mason, P.; Gao, K.; Genzen, J.R. Serotonergic raphe magnus cell discharge reflects ongoing autonomic and respiratory activities. J. Neurophysiol. 2007, 98, 1919–1927. [Google Scholar] [CrossRef] [Green Version]
- Heinricher, M.M.; Morgan, M.M.; Tortorici, V.; Fields, H.L. Disinhibition of off-cells and antinociception produced by an opioid action within the rostral ventromedial medulla. Neuroscience 1994, 63, 279–288. [Google Scholar] [CrossRef]
- Ossipov, M.H.; Morimura, K.; Porreca, F. Descending pain modulation and chronification of pain. Curr. Opin. Support. Palliat. Care 2014, 8, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Marlier, L.; Teilhac, J.R.; Cerruti, C.; Privat, A. Autoradiographic mapping of 5-HT1, 5-HT1A, 5-HT1B and 5-HT2 receptors in the rat spinal cord. Brain Res. 1991, 550, 15–23. [Google Scholar] [CrossRef]
- Otoshi, C.K.; Walwyn, W.M.; Tillakaratne, N.J.; Zhong, H.; Roy, R.R.; Edgerton, V.R. Distribution and localization of 5-HT(1A) receptors in the rat lumbar spinal cord after transection and deafferentation. J. Neurotrauma 2009, 26, 575–584. [Google Scholar] [CrossRef]
- Kia, H.K.; Miquel, M.C.; Brisorgueil, M.J.; Daval, G.; Riad, M.; El Mestikawy, S.; Hamon, M.; Vergé, D. Immunocytochemical localization of serotonin1A receptors in the rat central nervous system. J. Comp. Neurol. 1996, 365, 289–305. [Google Scholar] [CrossRef]
- Talley, E.M.; Bayliss, D.A. Postnatal development of 5-HT(1A) receptor expression in rat somatic motoneurons. Dev. Brain Res. 2000, 122, 1–10. [Google Scholar] [CrossRef]
- Colpaert, F.C. 5-HT(1A) receptor activation: New molecular and neuroadaptive mechanisms of pain relief. Curr. Opin. Investig. Drugs 2006, 7, 40–47. [Google Scholar]
- Kayser, V.; Elfassi, I.E.; Aubel, B.; Melfort, M.; Julius, D.; Gingrich, J.A.; Hamon, M.; Bourgoin, S. Mechanical, thermal and formalin-induced nociception is differentially altered in 5-HT1A−/−, 5-HT1B−/−, 5-HT2A−/−, 5-HT3A−/− and 5-HTT−/− knock-out male mice. Pain 2007, 130, 235–248. [Google Scholar] [CrossRef]
- Ali, Z.; Wu, G.; Kozlov, A.; Barasi, S. The actions of 5-HT1 agonists and antagonists on nociceptive processing in the rat spinal cord: Results from behavioural and electrophysiological studies. Brain Res. 1994, 661, 83–90. [Google Scholar] [CrossRef]
- Alhaider, A.A.; Wilcox, G.L. Differential roles of 5-hydroxytryptamine1A and 5-hydroxytryptamine1B receptor subtypes in modulating spinal nociceptive transmission in mice. J. Pharmacol. Exp. Ther. 1993, 265, 378–385. [Google Scholar]
- Perrier, J.F.; Cotel, F. Serotonin differentially modulates the intrinsic properties of spinal motoneurons from the adult turtle. J. Physiol. 2008, 586, 1233–1238. [Google Scholar] [CrossRef]
- Lecci, A.; Giuliani, S.; Santicioli, P.; Maggi, C.A. Involvement of 5-hydroxytryptamine1A receptors in the modulation of micturition reflexes in the anesthetized rat. J. Pharmacol. Exp. Ther. 1992, 262, 181–189. [Google Scholar]
- Ishizuka, O.; Gu, B.; Igawa, Y.; Nishizawa, O.; Pehrson, R.; Andersson, K.E. Role of supraspinal serotonin receptors for micturition in normal conscious rats. Neurourol. Urodyn. 2002, 21, 225–230. [Google Scholar] [CrossRef]
- Testa, R.; Guarneri, L.; Poggesi, E.; Angelico, P.; Velasco, C.; Ibba, M.; Cilia, A.; Motta, G.; Riva, C.; Leonardi, A. Effect of several 5-hydroxytryptamine(1A) receptor ligands on the micturition reflex in rats: Comparison with WAY 100635. J. Pharmacol. Exp. Ther. 1999, 290, 1258–1269. [Google Scholar]
- Conley, R.K.; Williams, T.J.; Ford, A.P.D.W.; Ramage, A.G. The role of α1-adrenoceptors and 5-HT1A receptors in the control of the micturition reflex in male anaesthetized rats. Br. J. Pharmacol. 2001, 133, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Kakizaki, H.; Yoshiyama, M.; Koyanagi, T.; De Groat, W.C. Effects of WAY100635, a selective 5-HT1A-receptor antagonist on the micturition-reflex pathway in the rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R1407–R1413. [Google Scholar] [CrossRef]
- Pehrson, R.; Ojteg, G.; Ishizuka, O.; Andersson, K.E. Effects of NAD-299, a new, highly selective 5-HT1A receptor antagonist, on bladder function in rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2002, 366, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Beato, M.; Nistri, A. Serotonin-induced inhibition of locomotor rhythm of the rat isolated spinal cord is mediated by the 5-HT1 receptor class. Proc. Biol. Sci. 1998, 265, 2073–2080. [Google Scholar] [CrossRef] [Green Version]
- Dunbar, M.J.; Tran, M.A.; Whelan, P.J. Endogenous extracellular serotonin modulates the spinal locomotor network of the neonatal mouse. J. Physiol. 2010, 588, 139–156. [Google Scholar] [CrossRef] [PubMed]
- Antri, M.; Mouffle, C.; Orsal, D.; Barthe, J.Y. 5-HT1A receptors are involved in short- and long-term processes responsible for 5-HT-induced locomotor function recovery in chronic spinal rat. Eur. J. Neurosci. 2003, 18, 1963–1972. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.A.; White, S.R. Receptor subtypes mediating facilitation by serotonin of excitability of spinal motoneurons. Neuropharmacology 1990, 29, 787–797. [Google Scholar] [CrossRef]
- Landry, E.S.; Lapointe, N.P.; Rouillard, C.; Levesque, D.; Hedlund, P.B.; Guertin, P.A. Contribution of spinal 5-HT1A and 5-HT7 receptors to locomotor-like movement induced by 8-OH-DPAT in spinal cord-transected mice. Eur. J. Neurosci. 2006, 24, 535–546. [Google Scholar] [CrossRef]
- Thor, K.B.; Nickolaus, S.; Helke, C.J. Autoradiographic localization of 5-hydroxytryptamine1A, 5-hydroxytryptamine1B and 5-hydroxytryptamine1C/2 binding sites in the rat spinal cord. Neuroscience 1993, 55, 235–252. [Google Scholar] [CrossRef]
- Laporte, A.M.; Doyen, C.; Nevo, I.T.; Chauveau, J.; Hauw, J.J.; Hamon, M. Autoradiographic mapping of serotonin 5-HT1A, 5-HT1D, 5-HT2A and 5-HT3 receptors in the aged human spinal cord. J. Chem. Neuroanat. 1996, 11, 67–75. [Google Scholar] [CrossRef]
- Kayser, V.; Aubel, B.; Hamon, M.; Bourgoin, S. The antimigraine 5-HT 1B/1D receptor agonists, sumatriptan, zolmitriptan and dihydroergotamine, attenuate pain-related behaviour in a rat model of trigeminal neuropathic pain. Br. J. Pharmacol. 2002, 137, 1287–1297. [Google Scholar] [CrossRef] [Green Version]
- Murray, K.C.; Stephens, M.J.; Rank, M.; D’Amico, J.; Gorassini, M.A.; Bennett, D.J. Polysynaptic excitatory postsynaptic potentials that trigger spasms after spinal cord injury in rats are inhibited by 5-HT1B and 5-HT1F receptors. J. Neurophysiol. 2011, 106, 925–943. [Google Scholar] [CrossRef] [Green Version]
- Honda, M.; Tanabe, M.; Ono, H. Serotonergic depression of spinal monosynaptic transmission is mediated by 5-HT1B receptors. Eur. J. Pharmacol. 2003, 482, 155–161. [Google Scholar] [CrossRef]
- Potrebic, S.; Ahn, A.H.; Skinner, K.; Fields, H.L.; Basbaum, A.I. Peptidergic nociceptors of both trigeminal and dorsal root ganglia express serotonin 1D receptors: Implications for the selective antimigraine action of triptans. J. Neurosci. 2003, 23, 10988–10997. [Google Scholar] [CrossRef]
- Ahn, A.H.; Basbaum, A.I. Tissue injury regulates serotonin 1D receptor expression: Implications for the control of migraine and inflammatory pain. J. Neurosci. 2006, 26, 8332–8338. [Google Scholar] [CrossRef] [Green Version]
- Enjin, A.; Leão, K.E.; Mikulovic, S.; Le Merre, P.; Tourtellotte, W.G.; Kullander, K. Sensorimotor function is modulated by the serotonin receptor 1d, a novel marker for gamma motor neurons. Mol. Cell. Neurosci. 2012, 49, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Honda, M.; Imaida, K.; Tanabe, M.; Ono, H. Endogenously released 5-hydroxytryptamine depresses the spinal monosynaptic reflex via 5-HT1D receptors. Eur. J. Pharmacol. 2004, 503, 55–61. [Google Scholar] [CrossRef]
- Gu, B.; Olejar, K.J.; Reiter, J.P.; Thor, K.B.; Dolber, P.C. Inhibition of bladder activity by 5-hydroxytryptamine1 serotonin receptor agonists in cats with chronic spinal cord injury. J. Pharmacol. Exp. Ther. 2004, 310, 1266–1272. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, J.M.; Li, Y.; Bennett, D.J.; Gorassini, M.A. Reduction of spinal sensory transmission by facilitation of 5-HT1B/D receptors in noninjured and spinal cord-injured humans. J. Neurophysiol. 2013, 109, 1485–1493. [Google Scholar] [CrossRef] [Green Version]
- Classey, J.D.; Bartsch, T.; Goadsby, P.J. Distribution of 5-HT(1B), 5-HT(1D) and 5-HT(1F) receptor expression in rat trigeminal and dorsal root ganglia neurons: Relevance to the selective anti-migraine effect of triptans. Brain Res. 2010, 1361, 76–85. [Google Scholar] [CrossRef]
- Agosti, R.M. 5HT1F- and 5HT7-receptor agonists for the treatment of migraines. CNS Neurol. Disord. Drug Targets 2007, 6, 235–237. [Google Scholar] [CrossRef]
- Ferrari, M.D.; Färkkilä, M.; Reuter, U.; Pilgrim, A.; Davis, C.; Krauss, M.; Diener, H.C. Acute treatment of migraine with the selective 5-HT1F receptor agonist lasmiditan--a randomised proof-of-concept trial. Cephalalgia 2010, 30, 1170–1178. [Google Scholar] [CrossRef]
- Doly, S.; Madeira, A.; Fischer, J.; Brisorgueil, M.J.; Daval, G.; Bernard, R.; Vergé, D.; Conrath, M. The 5-HT2A receptor is widely distributed in the rat spinal cord and mainly localized at the plasma membrane of postsynaptic neurons. J. Comp. Neurol. 2004, 472, 496–511. [Google Scholar] [CrossRef]
- Pompeiano, M.; Palacios, J.M.; Mengod, G. Distribution of the serotonin 5-HT2 receptor family mRNAs: Comparison between 5-HT2A and 5-HT2C receptors. Brain Res. Mol. Brain Res. 1994, 23, 163–178. [Google Scholar] [CrossRef]
- Xie, D.J.; Uta, D.; Feng, P.Y.; Wakita, M.; Shin, M.C.; Furue, H.; Yoshimura, M. Identification of 5-HT receptor subtypes enhancing inhibitory transmission in the rat spinal dorsal horn in vitro. Mol. Pain 2012, 8, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, T.; Otsuguro, K.; Kobayashi, T.; Ohta, T.; Ito, S. Endogenously released 5-HT inhibits A and C fiber-evoked synaptic transmission in the rat spinal cord by the facilitation of GABA/glycine and 5-HT release via 5-HT(2A) and 5-HT(3) receptors. Eur. J. Pharmacol. 2013, 702, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Aira, Z.; Buesa, I.; Gallego, M.; García del Caño, G.; Mendiable, N.; Mingo, J.; Rada, D.; Bilbao, J.; Zimmermann, M.; Azkue, J.J. Time-dependent cross talk between spinal serotonin 5-HT2A receptor and mGluR1 subserves spinal hyperexcitability and neuropathic pain after nerve injury. J. Neurosci. 2012, 32, 13568–13581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obata, H.; Saito, S.; Ishizaki, K.; Goto, F. Antinociception in rat by sarpogrelate, a selective 5-HT(2A) receptor antagonist, is peripheral. Eur. J. Pharmacol. 2000, 404, 95–102. [Google Scholar] [CrossRef]
- Crown, E.D.; Grau, J.W. Evidence that descending serotonergic systems protect spinal cord plasticity against the disruptive effect of uncontrollable stimulation. Exp. Neurol. 2005, 196, 164–176. [Google Scholar] [CrossRef]
- Watson, N.V.; Gorzalka, B.B. DOI-induced inhibition of copulatory behavior in male rats: Reversal by 5-HT2 antagonists. Pharmacol. Biochem. Behav. 1991, 39, 605–612. [Google Scholar] [CrossRef]
- Rössler, A.S.; Bernabé, J.; Denys, P.; Alexandre, L.; Giuliano, F. Effect of the 5-HT receptor agonist DOI on female rat sexual behavior. J. Sex. Med. 2006, 3, 432–441. [Google Scholar] [CrossRef]
- Mbaki, Y.; Gardiner, J.; McMurray, G.; Ramage, A.G. 5-HT 2A receptor activation of the external urethral sphincter and 5-HT 2C receptor inhibition of micturition: A study based on pharmacokinetics in the anaesthetized female rat. Eur. J. Pharmacol. 2012, 682, 142–152. [Google Scholar] [CrossRef]
- Chen, J.; Gu, B.; Wu, G.; Tu, H.; Si, J.; Xu, Y.; Andersson, K.E. The effect of the 5-HT2A/2C receptor agonist DOI on micturition in rats with chronic spinal cord injury. J. Urol. 2013, 189, 1982–1988. [Google Scholar] [CrossRef]
- Gordon, I.T.; Whelan, P.J. Monoaminergic control of cauda-equina-evoked locomotion in the neonatal mouse spinal cord. J. Neurophysiol. 2006, 96, 3122–3129. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Jordan, L.M. Stimulation of the parapyramidal region of the neonatal rat brain stem produces locomotor-like activity involving spinal 5-HT7 and 5-HT2A receptors. J. Neurophysiol. 2005, 94, 1392–1404. [Google Scholar] [CrossRef] [Green Version]
- Pearlstein, E.; Ben Mabrouk, F.; Pflieger, J.F.; Vinay, L. Serotonin refines the locomotor-related alternations in the in vitro neonatal rat spinal cord. Eur. J. Neurosci. 2005, 21, 1338–1346. [Google Scholar] [CrossRef]
- Nardone, R.; Holler, Y.; Thomschewski, A.; Holler, P.; Lochner, P.; Golaszewski, S.; Brigo, F.; Trinka, E. Serotonergic transmission after spinal cord injury. J. Neural Transm. 2015, 122, 279–295. [Google Scholar] [CrossRef]
- Antri, M.; Orsal, D.; Barthe, J.Y. Locomotor recovery in the chronic spinal rat: Effects of long-term treatment with a 5-HT2 agonist. Eur. J. Neurosci. 2002, 16, 467–476. [Google Scholar] [CrossRef]
- Zhou, S.Y.; Basura, G.J.; Goshgarian, H.G. Serotonin(2) receptors mediate respiratory recovery after cervical spinal cord hemisection in adult rats. J. Appl. Physiol. 2001, 91, 2665–2673. [Google Scholar] [CrossRef] [Green Version]
- Pineda-Farias, J.B.; Velázquez-Lagunas, I.; Barragán-Iglesias, P.; Cervantes-Durán, C.; Granados-Soto, V. 5-HT(2B) Receptor Antagonists Reduce Nerve Injury-Induced Tactile Allodynia and Expression of 5-HT(2B) Receptors. Drug Dev. Res. 2015, 76, 31–39. [Google Scholar] [CrossRef]
- Lin, S.Y.; Chang, W.J.; Lin, C.S.; Huang, C.Y.; Wang, H.F.; Sun, W.H. Serotonin receptor 5-HT2B mediates serotonin-induced mechanical hyperalgesia. J. Neurosci. 2011, 31, 1410–1418. [Google Scholar] [CrossRef] [Green Version]
- MacFarlane, P.M.; Vinit, S.; Mitchell, G.S. Serotonin 2A and 2B receptor-induced phrenic motor facilitation: Differential requirement for spinal NADPH oxidase activity. Neuroscience 2011, 178, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Cervantes-Durán, C.; Vidal-Cantú, G.C.; Barragán-Iglesias, P.; Pineda-Farias, J.B.; Bravo-Hernández, M.; Murbartián, J.; Granados-Soto, V. Role of peripheral and spinal 5-HT2B receptors in formalin-induced nociception. Pharmacol. Biochem. Behav. 2012, 102, 30–35. [Google Scholar] [CrossRef]
- Murray, K.C.; Stephens, M.J.; Ballou, E.W.; Heckman, C.J.; Bennett, D.J. Motoneuron excitability and muscle spasms are regulated by 5-HT2B and 5-HT2C receptor activity. J. Neurophysiol. 2011, 105, 731–748. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, M.I.; Ni, Y.G.; Dunning, D.D.; Miledi, R. Distribution of serotonin 2A, 2C and 3 receptor mRNA in spinal cord and medulla oblongata. Mol. Brain Res. 2001, 89, 11–19. [Google Scholar] [CrossRef]
- Ren, L.Q.; Wienecke, J.; Chen, M.; Møller, M.; Hultborn, H.; Zhang, M. The time course of serotonin 2C receptor expression after spinal transection of rats: An immunohistochemical study. Neuroscience 2013, 236, 31–46. [Google Scholar] [CrossRef]
- Machacek, D.W.; Garraway, S.M.; Shay, B.L.; Hochman, S. Serotonin 5-HT(2) receptor activation induces a long-lasting amplification of spinal reflex actions in the rat. J. Physiol. 2001, 537, 201–207. [Google Scholar] [CrossRef]
- Halberstadt, A.L.; van der Heijden, I.; Ruderman, M.A.; Risbrough, V.B.; Gingrich, J.A.; Geyer, M.A.; Powell, S.B. 5-HT(2A) and 5-HT(2C) receptors exert opposing effects on locomotor activity in mice. Neuropsychopharmacology 2009, 34, 1958–1967. [Google Scholar] [CrossRef] [Green Version]
- Mbaki, Y.; Ramage, A.G. Investigation of the role of 5-HT2 receptor subtypes in the control of the bladder and the urethra in the anaesthetized female rat. Br. J. Pharmacol. 2008, 155, 343–356. [Google Scholar] [CrossRef] [Green Version]
- Conlon, K.; Miner, W.; McCleary, S.; McMurray, G. Identification of 5-HT(2C) mediated mechanisms involved in urethral sphincter reflexes in a guinea-pig model of urethral function. BJU Int. 2012, 110, E113–E117. [Google Scholar] [CrossRef]
- Murray, K.C.; Nakae, A.; Stephens, M.J.; Rank, M.; D’Amico, J.; Harvey, P.J.; Li, X.; Harris, R.L.; Ballou, E.W.; Anelli, R.; et al. Recovery of motoneuron and locomotor function after spinal cord injury depends on constitutive activity in 5-HT2C receptors. Nat. Med. 2010, 16, 694–700. [Google Scholar] [CrossRef] [Green Version]
- Morales, M.; Battenberg, E.; Bloom, F.E. Distribution of neurons expressing immunoreactivity for the 5HT3 receptor subtype in the rat brain and spinal cord. J. Comp. Neurol. 1998, 402, 385–401. [Google Scholar] [CrossRef]
- Morales, M.; McCollum, N.; Kirkness, E.F. 5-HT(3)-receptor subunits A and B are co-expressed in neurons of the dorsal root ganglion. J. Comp. Neurol. 2001, 438, 163–172. [Google Scholar] [CrossRef]
- Smith, M.I.; Banner, S.E.; Sanger, G.J. 5-HT4 receptor antagonism potentiates inhibition of intestinal allodynia by 5-HT3 receptor antagonism in conscious rats. Neurosci. Lett. 1999, 271, 61–64. [Google Scholar] [CrossRef]
- Doak, G.J.; Sawynok, J. Formalin-induced nociceptive behavior and edema: Involvement of multiple peripheral 5-hydroxytryptamine receptor subtypes. Neuroscience 1997, 80, 939–949. [Google Scholar] [CrossRef]
- Alhaider, A.A.; Lei, S.Z.; Wilcox, G.L. Spinal 5-HT3 receptor-mediated antinociception: Possible release of GABA. J. Neurosci. 1991, 11, 1881–1888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khasabov, S.G.; Lopez-Garcia, J.A.; Asghar, A.U.; King, A.E. Modulation of afferent-evoked neurotransmission by 5-HT3 receptors in young rat dorsal horn neurones in vitro: A putative mechanism of 5-HT3 induced anti-nociception. Br. J. Pharmacol. 1999, 127, 843–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espey, M.J.; Du, H.J.; Downie, J.W. Serotonergic modulation of spinal ascending activity and sacral reflex activity evoked by pelvic nerve stimulation in cats. Brain Res. 1998, 798, 101–108. [Google Scholar] [CrossRef]
- Guertin, P.A.; Steuer, I. Ionotropic 5-HT3 receptor agonist-induced motor responses in the hindlimbs of paraplegic mice. J. Neurophysiol. 2005, 94, 3397–3405. [Google Scholar] [CrossRef] [PubMed]
- Suwa, B.; Bock, N.; Preusse, S.; Rothenberger, A.; Manzke, T. Distribution of serotonin 4(a) receptors in the juvenile rat brain and spinal cord. J. Chem. Neuroanat. 2014, 55, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Godínez-Chaparro, B.; López-Santillán, F.J.; Orduña, P.; Granados-Soto, V. Secondary mechanical allodynia and hyperalgesia depend on descending facilitation mediated by spinal 5-HT₄, 5-HT₆ and 5-HT₇ receptors. Neuroscience 2012, 222, 379–391. [Google Scholar] [CrossRef]
- Sławińska, U.; Miazga, K.; Cabaj, A.M.; Leszczyńska, A.N.; Majczyński, H.; Nagy, J.I.; Jordan, L.M. Grafting of fetal brainstem 5-HT neurons into the sublesional spinal cord of paraplegic rats restores coordinated hindlimb locomotion. Exp. Neurol. 2013, 247, 572–581. [Google Scholar] [CrossRef]
- Sławińska, U.; Miazga, K.; Jordan, L.M. 5-HT₂ and 5-HT₇ receptor agonists facilitate plantar stepping in chronic spinal rats through actions on different populations of spinal neurons. Front. Neural Circuits 2014, 8, 95. [Google Scholar] [CrossRef] [Green Version]
- Doly, S.; Fischer, J.; Brisorgueil, M.J.; Vergé, D.; Conrath, M. 5-HT5A receptor localization in the rat spinal cord suggests a role in nociception and control of pelvic floor musculature. J. Comp. Neurol. 2004, 476, 316–329. [Google Scholar] [CrossRef]
- Cervantes-Durán, C.; Rocha-González, H.I.; Granados-Soto, V. Peripheral and spinal 5-HT receptors participate in the pronociceptive and antinociceptive effects of fluoxetine in rats. Neuroscience 2013, 252, 396–409. [Google Scholar] [CrossRef]
- Muñoz-Islas, E.; Vidal-Cantú, G.C.; Bravo-Hernández, M.; Cervantes-Durán, C.; Quiñonez-Bastidas, G.N.; Pineda-Farias, J.B.; Barragán-Iglesias, P.; Granados-Soto, V. Spinal 5-HT₅A receptors mediate 5-HT-induced antinociception in several pain models in rats. Pharmacol. Biochem. Behav. 2014, 120, 25–32. [Google Scholar] [CrossRef]
- Grailhe, R.; Grabtree, G.W.; Hen, R. Human 5-HT(5) receptors: The 5-HT(5A) receptor is functional but the 5-HT(5B) receptor was lost during mammalian evolution. Eur. J. Pharmacol. 2001, 418, 157–167. [Google Scholar] [CrossRef]
- Gérard, C.; Martres, M.P.; Lefèvre, K.; Miquel, M.C.; Vergé, D.; Lanfumey, L.; Doucet, E.; Hamon, M.; el Mestikawy, S. Immuno-localization of serotonin 5-HT6 receptor-like material in the rat central nervous system. Brain Res. 1997, 746, 207–219. [Google Scholar] [CrossRef]
- Gérard, C.; el Mestikawy, S.; Lebrand, C.; Adrien, J.; Ruat, M.; Traiffort, E.; Hamon, M.; Martres, M.P. Quantitative RT-PCR distribution of serotonin 5-HT6 receptor mRNA in the central nervous system of control or 5,7-dihydroxytryptamine-treated rats. Synapse 1996, 23, 164–173. [Google Scholar] [CrossRef]
- Castañeda-Corral, G.; Rocha-González, H.I.; Araiza-Saldaña, C.I.; Ambriz-Tututi, M.; Vidal-Cantú, G.C.; Granados-Soto, V. Role of peripheral and spinal 5-HT6 receptors according to the rat formalin test. Neuroscience 2009, 162, 444–452. [Google Scholar] [CrossRef]
- Doly, S.; Fischer, J.; Brisorgueil, M.J.; Vergé, D.; Conrath, M. Pre- and postsynaptic localization of the 5-HT7 receptor in rat dorsal spinal cord: Immunocytochemical evidence. J. Comp. Neurol. 2005, 490, 256–269. [Google Scholar] [CrossRef]
- Noga, B.R.; Johnson, D.M.; Riesgo, M.I.; Pinzon, A. Locomotor-activated neurons of the cat. I. Serotonergic innervation and co-localization of 5-HT7, 5-HT2A, and 5-HT1A receptors in the thoraco-lumbar spinal cord. J. Neurophysiol. 2009, 102, 1560–1576. [Google Scholar] [CrossRef] [Green Version]
- Yesilyurt, O.; Seyrek, M.; Tasdemir, S.; Kahraman, S.; Deveci, M.S.; Karakus, E.; Halici, Z.; Dogrul, A. The critical role of spinal 5-HT7 receptors in opioid and non-opioid type stress-induced analgesia. Eur. J. Pharmacol. 2015, 762, 402–410. [Google Scholar] [CrossRef]
- Brenchat, A.; Zamanillo, D.; Hamon, M.; Romero, L.; Vela, J.M. Role of peripheral versus spinal 5-HT(7) receptors in the modulation of pain undersensitizing conditions. Eur. J. Pain 2012, 16, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Viguier, F.; Michot, B.; Hamon, M.; Bourgoin, S. Multiple roles of serotonin in pain control mechanisms--implications of 5-HT₇ and other 5-HT receptor types. Eur. J. Pharmacol. 2013, 716, 8–16. [Google Scholar] [CrossRef]
- Read, K.E.; Sanger, G.J.; Ramage, A.G. Evidence for the involvement of central 5-HT7 receptors in the micturition reflex in anaesthetized female rats. Br. J. Pharmacol. 2003, 140, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Gang, W.; Hongjian, T.; Jasheng, C.; Jiemin, S.; Zhong, C.; Yuemin, X.; Baojun, G.; Andersson, K.E. The effect of the 5-HT7 serotonin receptor agonist, LP44, on micturition in rats with chronic spinal cord injury. Neurourol. Urodyn. 2014, 33, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Akay, T.; Hedlund, P.B.; Pearson, K.G.; Jordan, L.M. Spinal 5-HT7 receptors are critical for alternating activity during locomotion: In vitro neonatal and in vivo adult studies using 5-HT7 receptor knockout mice. J. Neurophysiol. 2009, 102, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Meuser, T.; Pietruck, C.; Gabriel, A.; Xie, G.X.; Lim, K.J.; Pierce Palmer, P. 5-HT7 receptors are involved in mediating 5-HT-induced activation of rat primary afferent neurons. Life Sci. 2002, 71, 2279–2289. [Google Scholar] [CrossRef]
- Barnes, N.M.; Sharp, T. A review of central 5-HT receptors and their function. Neuropharmacology 1999, 38, 1083–1152. [Google Scholar] [CrossRef]
- Huang, Y.J.; Grau, J.W. Ionic plasticity and pain: The loss of descending serotonergic fibers after spinal cord injury transforms how GABA affects pain. Exp. Neurol. 2018, 306, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Amedro, J.; Williams, G.; Smith, D. A pharmacological analysis of the rat spinal cord serotonin (5-HT) autoreceptor. Eur. J. Pharmacol. 1988, 145, 163–171. [Google Scholar] [CrossRef]
- Eide, P.K.; Joly, N.M.; Hole, K. The role of spinal cord 5-HT1A and 5-HT1B receptors in the modulation of a spinal nociceptive reflex. Brain Res. 1990, 536, 195–200. [Google Scholar] [CrossRef]
- Jeong, C.Y.; Choi, J.I.; Yoon, M.H. Roles of serotonin receptor subtypes for the antinociception of 5-HT in the spinal cord of rats. Eur. J. Pharmacol. 2004, 502, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Kjørsvik, A.; Tjølsen, A.; Hole, K. Activation of spinal serotonin(2A/2C) receptors augments nociceptive responses in the rat. Brain Res. 2001, 910, 179–181. [Google Scholar] [CrossRef]
- Landry, E.S.; Guertin, P.A. Differential effects of 5-HT1 and 5-HT2 receptor agonists on hindlimb movements in paraplegic mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2004, 28, 1053–1060. [Google Scholar] [CrossRef]
- Rahman, W.; Bannister, K.; Bee, L.A.; Dickenson, A.H. A pronociceptive role for the 5-HT2 receptor on spinal nociceptive transmission: An in vivo electrophysiological study in the rat. Brain Res. 2011, 1382, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Fone, K.C.; Robinson, A.J.; Marsden, C.A. Characterization of the 5-HT receptor subtypes involved in the motor behaviours produced by intrathecal administration of 5-HT agonists in rats. Br. J. Pharmacol. 1991, 103, 1547–1555. [Google Scholar] [CrossRef] [Green Version]
- Espey, M.J.; Downie, J.W.; Fine, A. Effect of 5-HT receptor and adrenoceptor antagonists on micturition in conscious cats. Eur. J. Pharmacol. 1992, 221, 167–170. [Google Scholar] [CrossRef]
- Holohean, A.M.; Hackman, J.C. Mechanisms intrinsic to 5-HT2B receptor-induced potentiation of NMDA receptor responses in frog motoneurones. Br. J. Pharmacol. 2004, 143, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Miyoshi, K.; Dubner, R.; Gu, M.; Li, M.; Liu, J.; Yang, J.; Zou, S.; Ren, K.; Noguchi, K.; et al. Spinal 5-HT3 receptors mediate descending facilitation and contribute to behavioral hypersensitivity via a reciprocal neuron-glial signaling cascade. Mol. Pain 2014, 10, 35. [Google Scholar] [CrossRef] [Green Version]
- Nichols, D.E.; Nichols, C.D. Serotonin receptors. Chem. Rev. 2008, 108, 1614–1641. [Google Scholar] [CrossRef]
- Finn, D.P.; Fone, K.C.; Beckett, S.R.; Baxter, J.A.; Ansell, L.; Marsden, C.A.; Chapman, V. The effects of pharmacological blockade of the 5-HT(6) receptor on formalin-evoked nociceptive behaviour, locomotor activity and hypothalamo-pituitary-adrenal axis activity in rats. Eur. J. Pharmacol. 2007, 569, 59–63. [Google Scholar] [CrossRef]
- Martínez-García, E.; Leopoldo, M.; Lacivita, E.; Terrón, J.A. Increase of capsaicin-induced trigeminal Fos-like immunoreactivity by 5-HT(7) receptors. Headache 2011, 51, 1511–1519. [Google Scholar] [CrossRef] [Green Version]
- Brenchat, A.; Nadal, X.; Romero, L.; Ovalle, S.; Muro, A.; Sánchez-Arroyos, R.; Portillo-Salido, E.; Pujol, M.; Montero, A.; Codony, X.; et al. Pharmacological activation of 5-HT7 receptors reduces nerve injury-induced mechanical and thermal hypersensitivity. Pain 2010, 149, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Brenchat, A.; Ejarque, M.; Zamanillo, D.; Vela, J.M.; Romero, L. Potentiation of morphine analgesia by adjuvant activation of 5-HT7 receptors. J. Pharmacol. Sci. 2011, 116, 388–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaya-Castellanos, E.; Pineda-Farias, J.B.; Castañeda-Corral, G.; Vidal-Cantú, G.C.; Murbartián, J.; Rocha-González, H.I.; Granados-Soto, V. Blockade of 5-HT7 receptors reduces tactile allodynia in the rat. Pharmacol. Biochem. Behav. 2011, 99, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Cervo, L.; Rossi, C.; Tatarczynska, E.; Samanin, R. Role of 5-HT1A receptors in the antinociceptive action of 8-hydroxy-2-(di-n- propylamino)tetralin in the rat. Eur. J. Pharmacol. 1994, 263, 187–191. [Google Scholar] [CrossRef]
- Bardin, L.; Tarayre, J.P.; Koek, W.; Colpaert, F.C. In the formalin model of tonic nociceptive pain, 8-OH-DPAT produces 5-HT1A receptor-mediated, behaviorally specific analgesia. Eur. J. Pharmacol. 2001, 421, 109–114. [Google Scholar] [CrossRef]
- El-Yassir, N.; Fleetwood-Walker, S.M.; Mitchell, R. Heterogeneous effects of serotonin in the dorsal horn of rat: The involvement of 5-HT1 receptor subtypes. Brain Res. 1988, 456, 147–158. [Google Scholar] [CrossRef]
- Garraway, S.M.; Hochman, S. Pharmacological characterization of serotonin receptor subtypes modulating primary afferent input to deep dorsal horn neurons in the neonatal rat. Br. J. Pharmacol. 2001, 132, 1789–1798. [Google Scholar] [CrossRef] [Green Version]
- Garraway, S.M.; Hochman, S. Serotonin increases the incidence of primary afferent-evoked long-term depression in rat deep dorsal horn neurons. J. Neurophysiol. 2001, 85, 1864–1872. [Google Scholar] [CrossRef]
- Gjerstad, J.; Tjolsen, A.; Hole, K. The effect of 5-HT1A receptor stimulation on nociceptive dorsal horn neurones in rats. Eur. J. Pharmacol. 1996, 318, 315–321. [Google Scholar] [CrossRef]
- Barbeau, H.; Rossignol, S. The effects of serotonergic drugs on the locomotor pattern and on cutaneous reflexes of the adult chronic spinal cat. Brain Res. 1990, 514, 55–67. [Google Scholar] [CrossRef]
- Barbeau, H.; Rossignol, S. Initiation and modulation of the locomotor pattern in the adult chronic spinal cat by noradrenergic, serotonergic and dopaminergic drugs. Brain Res. 1991, 546, 250–260. [Google Scholar] [CrossRef]
- Cazalets, J.R.; Sqalli-Houssaini, Y.; Clarac, F. Activation of the central pattern generators for locomotion by serotonin and excitatory amino acids in neonatal rat. J. Physiol. 1992, 455, 187–204. [Google Scholar] [CrossRef]
- Cowley, K.C.; Schmidt, B.J. A comparison of motor patterns induced by N-methyl-D-aspartate, acetylcholine and serotonin in the in vitro neonatal rat spinal cord. Neurosci. Lett. 1994, 171, 147–150. [Google Scholar] [CrossRef]
- Feraboli-Lohnherr, D.; Barthe, J.Y.; Orsal, D. Serotonin-induced activation of the network for locomotion in adult spinal rats. J. Neurosci. Res. 1999, 55, 87–98. [Google Scholar] [CrossRef]
- Kiehn, O.; Kjaerulff, O. Spatiotemporal characteristics of 5-HT and dopamine-induced rhythmic hindlimb activity in the in vitro neonatal rat. J. Neurophysiol. 1996, 75, 1472–1482. [Google Scholar] [CrossRef]
- Ung, R.V.; Landry, E.S.; Rouleau, P.; Lapointe, N.P.; Rouillard, C.; Guertin, P.A. Role of spinal 5-HT2 receptor subtypes in quipazine-induced hindlimb movements after a low-thoracic spinal cord transection. Eur. J. Neurosci. 2008, 28, 2231–2242. [Google Scholar] [CrossRef]
- Madriaga, M.A.; McPhee, L.C.; Chersa, T.; Christie, K.J.; Whelan, P.J. Modulation of locomotor activity by multiple 5-HT and dopaminergic receptor subtypes in the neonatal mouse spinal cord. J. Neurophysiol. 2004, 92, 1566–1576. [Google Scholar] [CrossRef]
- Nishimaru, H.; Takizawa, H.; Kudo, N. 5-Hydroxytryptamine-induced locomotor rhythm in the neonatal mouse spinal cord in vitro. Neurosci. Lett. 2000, 280, 187–190. [Google Scholar] [CrossRef]
- Slawinska, U.; Jordan, L.M. Serotonergic influences on locomotor circuits. Curr. Opin. Physiol. 2019, 8, 7. [Google Scholar] [CrossRef]
- Miles, G.B.; Sillar, K.T. Neuromodulation of Vertebrate Locomotor Control Networks. Physiology 2011, 26, 393–411. [Google Scholar] [CrossRef]
- Alvarez, F.J.; Pearson, J.C.; Harrington, D.; Dewey, D.; Torbeck, L.; Fyffe, R.E. Distribution of 5-hydroxytryptamine-immunoreactive boutons on alpha-motoneurons in the lumbar spinal cord of adult cats. J. Comp. Neurol. 1998, 393, 69–83. [Google Scholar] [CrossRef]
- Hammar, I.; Bannatyne, B.A.; Maxwell, D.J.; Edgley, S.A.; Jankowska, E. The actions of monoamines and distribution of noradrenergic and serotoninergic contacts on different subpopulations of commissural interneurons in the cat spinal cord. Eur. J. Neurosci. 2004, 19, 1305–1316. [Google Scholar] [CrossRef] [Green Version]
- Ballion, B.; Branchereau, P.; Chapron, J.; Viala, D. Ontogeny of descending serotonergic innervation and evidence for intraspinal 5-HT neurons in the mouse spinal cord. Dev. Brain Res. 2002, 137, 81–88. [Google Scholar] [CrossRef]
- Cabaj, A.M.; Majczynski, H.; Couto, E.; Gardiner, P.F.; Stecina, K.; Slawinska, U.; Jordan, L.M. Serotonin controls initiation of locomotion and afferent modulation of coordination via 5-HT7 receptors in adult rats. J. Physiol. 2017, 595, 301–320. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, B.J.; Jordan, L.M. The role of serotonin in reflex modulation and locomotor rhythm production in the mammalian spinal cord. Brain Res. Bull. 2000, 53, 689–710. [Google Scholar] [CrossRef]
- Courtine, G.; Gerasimenko, Y.; van den Brand, R.; Yew, A.; Musienko, P.; Zhong, H.; Song, B.; Ao, Y.; Ichiyama, R.M.; Lavrov, I.; et al. Transformation of nonfunctional spinal circuits into functional states after the loss of brain input. Nat. Neurosci. 2009, 12, 1333–1342. [Google Scholar] [CrossRef]
- Fouad, K.; Rank, M.M.; Vavrek, R.; Murray, K.C.; Sanelli, L.; Bennett, D.J. Locomotion after spinal cord injury depends on constitutive activity in serotonin receptors. J. Neurophysiol. 2010, 104, 2975–2984. [Google Scholar] [CrossRef]
- Calaresu, F.R.; Yardley, C.P. Medullary basal sympathetic tone. Annu. Rev. Physiol. 1988, 50, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Chalmers, J.; Arnolda, L.; Llewellyn-Smith, I.; Minson, J.; Pilowsky, P.; Suzuki, S. Central neurons and neurotransmitters in the control of blood pressure. Clin. Exp. Pharmacol. Physiol. 1994, 21, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Jansen, A.S.; Nguyen, X.V.; Karpitskiy, V.; Mettenleiter, T.C.; Loewy, A.D. Central command neurons of the sympathetic nervous system: Basis of the fight-or-flight response. Science 1995, 270, 644–646. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn-Smith, I.J. Anatomy of synaptic circuits controlling the activity of sympathetic preganglionic neurons. J. Chem. Neuroanat. 2009, 38, 231–239. [Google Scholar] [CrossRef]
- Pyner, S.; Coote, J.H. Evidence that sympathetic preganglionic neurones are arranged in target-specific columns in the thoracic spinal cord of the rat. J. Comp. Neurol. 1994, 342, 15–22. [Google Scholar] [CrossRef]
- Tang, F.R.; Tan, C.K.; Ling, E.A. A light-microscopic study of the intermediolateral nucleus following injection of CB-HRP and fluorogold into the superior cervical ganglion of the rat. J. Auton. Nerv. Syst. 1995, 50, 333–338. [Google Scholar] [CrossRef]
- Zagon, A.; Smith, A.D. Monosynaptic projections from the rostral ventrolateral medulla oblongata to identified sympathetic preganglionic neurons. Neuroscience 1993, 54, 729–743. [Google Scholar] [CrossRef]
- Granata, A.R.; Ruggiero, D.A. Evidence of disynaptic projections from the rostral ventrolateral medulla to the thoracic spinal cord. Brain Res. 1998, 781, 329–334. [Google Scholar] [CrossRef]
- Guyenet, P.G.; Haselton, J.R.; Sun, M.K. Sympathoexcitatory neurons of the rostroventrolateral medulla and the origin of the sympathetic vasomotor tone. Prog. Brain Res. 1989, 81, 105–116. [Google Scholar] [CrossRef]
- Furlan, J.C.; Fehlings, M.G.; Shannon, P.; Norenberg, M.D.; Krassioukov, A.V. Descending vasomotor pathways in humans: Correlation between axonal preservation and cardiovascular dysfunction after spinal cord injury. J. Neurotrauma 2003, 20, 1351–1363. [Google Scholar] [CrossRef]
- Kerr, F.W.; Alexander, S. Descending Autonomic Pathways in the Spinal Cord. Arch. Neurol. 1964, 10, 249–261. [Google Scholar] [CrossRef]
- Hou, S.; Lu, P.; Blesch, A. Characterization of supraspinal vasomotor pathways and autonomic dysreflexia after spinal cord injury in F344 rats. Auton. Neurosci. 2013, 176, 54–63. [Google Scholar] [CrossRef]
- Bootle, D.J.; Adcock, J.J.; Ramage, A.G. Involvement of central 5-HT1A receptors in the reflex activation of pulmonary vagal motoneurones by inhaled capsaicin in anaesthetized cats. Br. J. Pharmacol. 1996, 117, 724–728. [Google Scholar] [CrossRef] [Green Version]
- Bootle, D.J.; Adcock, J.J.; Ramage, A.G. The role of central 5-HT receptors in the bronchoconstriction evoked by inhaled capsaicin in anaesthetised guinea-pigs. Neuropharmacology 1998, 37, 243–250. [Google Scholar] [CrossRef]
- Bogle, R.G.; Pires, J.G.; Ramage, A.G. Evidence that central 5-HT1A-receptors play a role in the von Bezold-Jarisch reflex in the rat. Br. J. Pharmacol. 1990, 100, 757–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Futuro-Neto, H.A.; Pires, J.G.; Gilbey, M.P.; Ramage, A.G. Evidence for the ability of central 5-HT1A receptors to modulate the vagal bradycardia induced by stimulating the upper airways of anesthetized rabbits with smoke. Brain Res. 1993, 629, 349–354. [Google Scholar] [CrossRef]
- Dando, S.B.; Skinner, M.R.; Jordan, D.; Ramage, A.G. Modulation of the vagal bradycardia evoked by stimulation of upper airway receptors by central 5-HT1 receptors in anaesthetized rabbits. Br. J. Pharmacol. 1998, 125, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Skinner, M.R.; Ramage, A.G.; Jordan, D. Modulation of reflexly evoked vagal bradycardias by central 5-HT1A receptors in anaesthetized rabbits. Br. J. Pharmacol. 2002, 137, 861–873. [Google Scholar] [CrossRef]
- Wang, Y.; Ramage, A.G. The role of central 5-HT(1A) receptors in the control of B-fibre cardiac and bronchoconstrictor vagal preganglionic neurones in anaesthetized cats. J. Physiol. 2001, 536, 753–767. [Google Scholar] [CrossRef]
- Zhang, M. Normal Distribution and Plasticity of Serotonin Receptors after Spinal Cord Injury and Their Impacts on Motor Outputs. In Recovery of Motor Function Following Spinal Cord Injury; IntechOpen: Rijeka, Croatia, 2016. [Google Scholar] [CrossRef] [Green Version]
- Laporte, A.M.; Fattaccini, C.M.; Lombard, M.C.; Chauveau, J.; Hamon, M. Effects of dorsal rhizotomy and selective lesion of serotonergic and noradrenergic systems on 5-HT1A, 5-HT1B, and 5-HT3 receptors in the rat spinal cord. J. Neural Transm. Gen. Sect. 1995, 100, 207–223. [Google Scholar] [CrossRef]
- Giroux, N.; Rossignol, S.; Reader, T.A. Autoradiographic study of alpha1- and alpha2-noradrenergic and serotonin1A receptors in the spinal cord of normal and chronically transected cats. J. Comp. Neurol. 1999, 406, 402–414. [Google Scholar] [CrossRef]
- Lee, J.K.; Johnson, C.S.; Wrathall, J.R. Up-regulation of 5-HT2 receptors is involved in the increased H-reflex amplitude after contusive spinal cord injury. Exp. Neurol. 2007, 203, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Fuller, D.D.; Baker-Herman, T.L.; Golder, F.J.; Doperalski, N.J.; Watters, J.J.; Mitchell, G.S. Cervical spinal cord injury upregulates ventral spinal 5-HT2A receptors. J. Neurotrauma 2005, 22, 203–213. [Google Scholar] [CrossRef]
- Kong, X.Y.; Wienecke, J.; Chen, M.; Hultborn, H.; Zhang, M. The time course of serotonin 2A receptor expression after spinal transection of rats: An immunohistochemical study. Neuroscience 2011, 177, 114–126. [Google Scholar] [CrossRef]
- Kong, X.Y.; Wienecke, J.; Hultborn, H.; Zhang, M. Robust upregulation of serotonin 2A receptors after chronic spinal transection of rats: An immunohistochemical study. Brain Res. 2010, 1320, 60–68. [Google Scholar] [CrossRef]
- Kao, T.; Shumsky, J.S.; Jacob-Vadakot, S.; Himes, B.T.; Murray, M.; Moxon, K.A. Role of the 5-HT2C receptor in improving weight-supported stepping in adult rats spinalized as neonates. Brain Res. 2006, 1112, 159–168. [Google Scholar] [CrossRef]
- Hayashi, Y.; Jacob-Vadakot, S.; Dugan, E.A.; McBride, S.; Olexa, R.; Simansky, K.; Murray, M.; Shumsky, J.S. 5-HT precursor loading, but not 5-HT receptor agonists, increases motor function after spinal cord contusion in adult rats. Exp. Neurol. 2010, 221, 68–78. [Google Scholar] [CrossRef] [Green Version]
- Salzman, S.K.; Hirofuji, E.; Llados-Eckman, C.; MacEwen, G.D.; Beckman, A.L. Monoaminergic responses to spinal trauma. Participation of serotonin in posttraumatic progression of neural damage. J. Neurosurg. 1987, 66, 431–439. [Google Scholar] [CrossRef]
- Navarrett, S.; Collier, L.; Cardozo, C.; Dracheva, S. Alterations of serotonin 2C and 2A receptors in response to T10 spinal cord transection in rats. Neurosci. Lett. 2012, 506, 74–78. [Google Scholar] [CrossRef]
- Cornide-Petronio, M.E.; Fernández-López, B.; Barreiro-Iglesias, A.; Rodicio, M.C. Traumatic injury induces changes in the expression of the serotonin 1A receptor in the spinal cord of lampreys. Neuropharmacology 2014, 77, 369–378. [Google Scholar] [CrossRef]
- Sharma, H.S.; Olsson, Y.; Dey, P.K. Early accumulation of serotonin in rat spinal cord subjected to traumatic injury. Relation to edema and blood flow changes. Neuroscience 1990, 36, 725–730. [Google Scholar] [CrossRef]
- Siegal, T. Participation of Serotonergic Mechanisms in the Pathophysiology of Experimental Neoplastic Spinal Cord Compression. Neurology 1991, 41, 574–580. [Google Scholar] [CrossRef]
- Shapiro, S.; McBride, W.; Sartorius, C.; Chernet, E.; Sanders, S.; Hall, P. Quantification of Changes in Serotonin Uptake with Spinal Cord Injury. Neurosurgery 1990, 26, 424–428. [Google Scholar] [CrossRef]
- Oliveras, J.L.; Bourgoin, S.; Hery, F.; Besson, J.M.; Hamon, M. The topographical distribution of serotoninergic terminals in the spinal cord of the cat: Biochemical mapping by the combined use of microdissection and microassay procedures. Brain Res. 1977, 138, 393–406. [Google Scholar] [CrossRef]
- Faden, A.I.; Gannon, A.; Basbaum, A.I. Use of Serotonin Immunocytochemistry as a Marker of Injury Severity After Experimental Spinal Trauma in Rats. Brain Res. 1988, 450, 94–100. [Google Scholar] [CrossRef]
- Saruhashi, Y.; Matsusue, Y.; Fujimiya, M. The recovery of 5-HT transporter and 5-HT immunoreactivity in injured rat spinal cord. Arch. Orthop. Trauma Surg. 2009, 129, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Saruhashi, Y.; Young, W.; Perkins, R. The recovery of 5-HT immunoreactivity in lumbosacral spinal cord and locomotor function after thoracic hemisection. Exp. Neurol. 1996, 139, 203–213. [Google Scholar] [CrossRef]
- Hashimoto, T.; Fukuda, N. Contribution of serotonin neurons to the functional recovery after spinal cord injury in rats. Brain Res. 1991, 539, 263–270. [Google Scholar] [CrossRef]
- Gjerstad, J.; Tjolsen, A.; Hole, K. Induction of long-term potentiation of single wide dynamic range neurones in the dorsal horn is inhibited by descending pathways. Pain 2001, 91, 263–268. [Google Scholar] [CrossRef]
- Le Bars, D.; Gozariu, M.; Cadden, S.W. Animal models of nociception. Pharmacol. Rev. 2001, 53, 597–652. [Google Scholar]
- Willis, W.D. Mechanisms of central sensitization of nociceptive dorsal horn neurons. In Spinal Cord Plasticity; Springer: Boston, MA, USA, 2001; pp. 127–161. [Google Scholar]
- Ji, R.R.; Kohno, T.; Moore, K.A.; Woolf, C.J. Central sensitization and LTP: Do pain and memory share similar mechanisms? Trends Neurosci. 2003, 26, 696–705. [Google Scholar] [CrossRef]
- Latremoliere, A.; Woolf, C.J. Central Sensitization: A Generator of Pain Hypersensitivity by Central Neural Plasticity. J. Pain 2009, 10, 895–926. [Google Scholar] [CrossRef] [Green Version]
- Sandkühler, J. Models and mechanisms of hyperalgesia and allodynia. Physiol. Rev. 2009, 89, 707–758. [Google Scholar] [CrossRef]
- Gao, Y.J.; Ji, R.R. c-Fos and pERK, which is a better marker for neuronal activation and central sensitization after noxious stimulation and tissue injury? Open Pain J. 2009, 2, 11–17. [Google Scholar] [CrossRef]
- Woolf, C.J. Central sensitization: Implications for the diagnosis and treatment of pain. Pain 2011, 152, S2–S15. [Google Scholar] [CrossRef]
- Sandkuhler, J. Learning and memory in pain pathways. Pain 2000, 88, 113–118. [Google Scholar] [CrossRef]
- Sandkühler, J.; Liu, X. Induction of long-term potentiation at spinal synapses by noxious stimulation or nerve injury. Eur. J. Neurosci. 1998, 10, 2476–2480. [Google Scholar] [CrossRef]
- Liu, X.G.; Morton, C.R.; Azkue, J.J.; Zimmermann, M.; Sandkuhler, J. Long-term depression of C-fibre-evoked spinal field potentials by stimulation of primary afferent A delta-fibres in the adult rat. Eur. J. Neurosci. 1998, 10, 3069–3075. [Google Scholar] [CrossRef]
- Hains, B.C.; Fullwood, S.D.; Eaton, M.J.; Hulsebosch, C.E. Subdural engraftment of serotonergic neurons following spinal hemisection restores spinal serotonin, downregulates serotonin transporter, and increases BDNF tissue content in rat. Brain Res. 2001, 913, 35–46. [Google Scholar] [CrossRef]
- Hains, B.C.; Johnson, K.M.; McAdoo, D.J.; Eaton, M.J.; Hulsebosch, C.E. Engraftment of serotonergic precursors enhances locomotor function and attenuates chronic central pain behavior following spinal hemisection injury in the rat. Exp. Neurol. 2001, 171, 361–378. [Google Scholar] [CrossRef]
- Hains, B.C.; Yucra, J.A.; Eaton, M.J.; Hulsebosch, C.E. Intralesion transplantation of serotonergic precursors enhances locomotor recovery but has no effect on development of chronic central pain following hemisection injury in rats. Neurosci. Lett. 2002, 324, 222–226. [Google Scholar] [CrossRef]
- Bardin, L.; Schmidt, J.; Alloui, A.; Eschalier, A. Effect of intrathecal administration of serotonin in chronic pain models in rats. Eur. J. Pharmacol. 2000, 409, 37–43. [Google Scholar] [CrossRef]
- Hains, B.C.; Yucra, J.A.; Hulsebosch, C.E. Reduction of pathological and behavioral deficits following spinal cord contusion injury with the selective cyclooxygenase-2 inhibitor NS-398. J. Neurotrauma 2001, 18, 409–423. [Google Scholar] [CrossRef]
- Grau, J.W.; Huie, J.R.; Lee, K.H.; Hoy, K.C.; Huang, Y.J.; Turtle, J.D.; Strain, M.M.; Baumbauer, K.M.; Miranda, R.M.; Hook, M.A.; et al. Metaplasticity and behavior: How training and inflammation affect plastic potential within the spinal cord and recovery after injury. Front. Neural Circuits 2014, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.J.; Lee, K.H.; Murphy, L.; Garraway, S.M.; Grau, J.W. Acute spinal cord injury (SCI) transforms how GABA affects nociceptive sensitization. Exp. Neurol. 2016, 285, 82–95. [Google Scholar] [CrossRef] [Green Version]
- Grau, J.W.; Huie, J.R.; Garraway, S.M.; Hook, M.A.; Crown, E.D.; Baumbauer, K.M.; Lee, K.H.; Hoy, K.C.; Ferguson, A.R. Impact of behavioral control on the processing of nociceptive stimulation. Front. Physiol. 2012, 3, 21. [Google Scholar] [CrossRef] [Green Version]
- Grau, J.W.; Barstow, D.G.; Joynes, R.L. Instrumental learning within the spinal cord: I. Behavioral properties. Behav. Neurosci. 1998, 112, 1366–1386. [Google Scholar] [CrossRef]
- Grau, J.W.; Baine, R.E.; Bean, P.A.; Davis, J.A.; Fauss, G.N.; Henwood, M.K.; Hudson, K.E.; Johnston, D.T.; Tarbet, M.M.; Strain, M.M. Learning to promote recovery after spinal cord injury. Exp. Neurol. 2020, 330, 113334. [Google Scholar] [CrossRef]
- Joynes, R.L.; Ferguson, A.R.; Crown, E.D.; Patton, B.C.; Grau, J.W. Instrumental learning within the spinal cord: V. Evidence the behavioral deficit observed after noncontingent nociceptive stimulation reflects an intraspinal modification. Behav. Brain Res. 2003, 141, 159–170. [Google Scholar] [CrossRef]
- Joynes, R.L.; Grau, J.W. Instrumental learning within the spinal cord: III. Prior exposure to noncontingent shock induces a behavioral deficit that is blocked by an opioid antagonist. Neurobiol. Learn. Mem. 2004, 82, 35–51. [Google Scholar] [CrossRef]
- Crown, E.D.; Ferguson, A.R.; Joynes, R.L.; Grau, J.W. Instrumental learning within the spinal cord: IV. Induction and retention of the behavioral deficit observed after noncontingent shock. Behav. Neurosci. 2002, 116, 1032–1051. [Google Scholar] [CrossRef]
- Ferguson, A.R.; Crown, E.D.; Grau, J.W. Nociceptive plasticity inhibits adaptive learning in the spinal cord. Neuroscience 2006, 141, 421–431. [Google Scholar] [CrossRef]
- Hook, M.A.; Huie, J.R.; Grau, J.W. Peripheral inflammation undermines the plasticity of the isolated spinal cord. Behav. Neurosci. 2008, 122, 233–249. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, A.R.; Huie, J.R.; Crown, E.D.; Grau, J.W. Central nociceptive sensitization vs. spinal cord training: Opposing forms of plasticity that dictate function after complete spinal cord injury. Front. Physiol. 2012, 3, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huie, J.R.; Stuck, E.D.; Lee, K.H.; Irvine, K.A.; Beattie, M.S.; Bresnahan, J.C.; Grau, J.W.; Ferguson, A.R. AMPA Receptor Phosphorylation and Synaptic Colocalization on Motor Neurons Drive Maladaptive Plasticity below Complete Spinal Cord Injury. eNeuro 2015, 2, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huie, J.R.; Baumbauer, K.M.; Lee, K.H.; Bresnahan, J.C.; Beattie, M.S.; Ferguson, A.R.; Grau, J.W. Glial Tumor Necrosis Factor Alpha (TNF alpha) Generates Metaplastic Inhibition of Spinal Learning. PLoS ONE 2012, 7, e39751. [Google Scholar] [CrossRef] [PubMed]
- Beattie, M.S.; Hermann, G.E.; Rogers, R.C.; Bresnahan, J.C. Cell death in models of spinal cord injury. In Spinal Cord Trauma: Regeneration, Neural Repair and Functional Recovery; McKerracher, L., Doucet, G., Rossignol, S., Eds.; Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2002; Volume 137, pp. 37–47. [Google Scholar]
- Grau, J.W.; Washburn, S.N.; Hook, M.A.; Ferguson, A.R.; Crown, E.D.; Garcia, G.; Bolding, K.A.; Miranda, R.C. Uncontrollable stimulation undermines recovery after spinal cord injury. J. Neurotrauma 2004, 21, 1795–1817. [Google Scholar] [CrossRef]
- Turtle, J.D.; Strain, M.M.; Reynolds, J.A.; Huang, Y.J.; Lee, K.H.; Henwood, M.K.; Garraway, S.M.; Grau, J.W. Pain Input After Spinal Cord Injury (SCI) Undermines Long-Term Recovery and Engages Signal Pathways That Promote Cell Death. Front. Syst. Neurosci. 2018, 12, 14. [Google Scholar] [CrossRef] [Green Version]
- Garraway, S.M.; Woller, S.A.; Huie, J.R.; Hartman, J.J.; Hook, M.A.; Miranda, R.C.; Huang, Y.J.; Ferguson, A.R.; Grau, J.W. Peripheral noxious stimulation reduces withdrawal threshold to mechanical stimuli after spinal cord injury: Role of tumor necrosis factor alpha and apoptosis. Pain 2014, 155, 2344–2359. [Google Scholar] [CrossRef] [Green Version]
- Bharne, A.P.; Upadhya, M.A.; Kokare, D.M.; Subhedar, N.K. Effect of alpha-melanocyte stimulating hormone on locomotor recovery following spinal cord injury in mice: Role of serotonergic system. Neuropeptides 2011, 45, 25–31. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kim, H.S.; Choi, H.Y.; Oh, T.H.; Ju, B.G.; Yune, T.Y. Valproic acid attenuates blood-spinal cord barrier disruption by inhibiting matrix metalloprotease-9 activity and improves functional recovery after spinal cord injury. J. Neurochem. 2012, 121, 818–829. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kang, S.R.; Yune, T.Y. Fluoxetine prevents oligodendrocyte cell death by inhibiting microglia activation after spinal cord injury. J. Neurotrauma 2015, 32, 633–644. [Google Scholar] [CrossRef] [Green Version]
- Washburn, S.N.; Patton, B.C.; Ferguson, A.R.; Hudson, K.L.; Grau, J.W. Exposure to intermittent nociceptive stimulation under pentobarbital anesthesia disrupts spinal cord function in rats. Psychopharmacology 2007, 192, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Madhavan, L.; Freed, W.J.; Anantharam, V.; Kanthasamy, A.G. 5-hydroxytryptamine 1A receptor activation protects against N-methyl-D-aspartate-induced apoptotic cell death in striatal and mesencephalic cultures. J. Pharmacol. Exp. Ther. 2003, 304, 913–923. [Google Scholar] [CrossRef] [Green Version]
- Maura, G.; Marcoli, M.; Pepicelli, O.; Rosu, C.; Viola, C.; Raiteri, M. Serotonin inhibition of the NMDA receptor/nitric oxide/cyclic GMP pathway in human neocortex slices: Involvement of 5-HT(2C) and 5-HT(1A) receptors. Br. J. Pharmacol. 2000, 130, 1853–1858. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, A.R.; Huie, J.R.; Crown, E.D.; Baumbauer, K.M.; Hook, M.A.; Garraway, S.M.; Lee, K.H.; Hoy, K.C.; Grau, J.W. Maladaptive spinal plasticity opposes spinal learning and recovery in spinal cord injury. Front. Physiol. 2012, 3, 17. [Google Scholar] [CrossRef] [Green Version]
- Grau, J.W.; Huang, Y.J.; Turtle, J.D.; Strain, M.M.; Miranda, R.C.; Garraway, S.M.; Hook, M.A. When Pain Hurts: Nociceptive Stimulation Induces a State of Maladaptive Plasticity and Impairs Recovery after Spinal Cord Injury. J. Neurotrauma 2017, 34, 1873–1890. [Google Scholar] [CrossRef] [Green Version]
- Crown, E.D.; Grau, J.W. Preserving and restoring behavioral potential within the spinal cord using an instrumental training paradigm. J. Neurophysiol. 2001, 86, 845–855. [Google Scholar] [CrossRef]
- Gomez-Pinilla, F.; Huie, J.R.; Ying, Z.; Ferguson, A.R.; Crown, E.D.; Baumbauer, K.M.; Edgerton, V.R.; Grau, J.W. BDNF and learning: Evidence that instrumental training promotes learning within the spinal cord by up-regulating BDNF expression. Neuroscience 2007, 148, 893–906. [Google Scholar] [CrossRef] [Green Version]
- Huie, J.R.; Garraway, S.M.; Baumbauer, K.M.; Hoy, K.C.; Beas, B.S.; Montgomery, K.S.; Bizon, J.L.; Grau, J.W. Brain-derived neurotrophic factor promotes adaptive plasticity within the spinal cord and mediates the beneficial effects of controllable stimulation. Neuroscience 2012, 200, 74–90. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.J.; Lee, K.H.; Grau, J.W. Complete spinal cord injury (SCI) transforms how brain derived neurotrophic factor (BDNF) affects nociceptive sensitization. Exp. Neurol. 2017, 288, 38–50. [Google Scholar] [CrossRef]
- Weishaupt, N.; Blesch, A.; Fouad, K. BDNF: The career of a multifaceted neurotrophin in spinal cord injury. Exp. Neurol. 2012, 238, 254–264. [Google Scholar] [CrossRef]
- Ying, Z.; Roy, R.R.; Edgerton, V.R.; Gomez-Pinilla, F. Exercise restores levels of neurotrophins and synaptic plasticity following spinal cord injury. Exp. Neurol. 2005, 193, 411–419. [Google Scholar] [CrossRef]
- Boyce, V.S.; Mendell, L.M. Neurotrophins and spinal circuit function. Front. Neural Circuits 2014, 8, 59. [Google Scholar] [CrossRef] [Green Version]
- Tashiro, S.; Shinozaki, M.; Mukaino, M.; Renault-Mihara, F.; Toyama, Y.; Liu, M.G.; Nakamura, M.; Okano, H. BDNF Induced by Treadmill Training Contributes to the Suppression of Spasticity and Allodynia After Spinal Cord Injury via Upregulation of KCC2. Neurorehabilit. Neural Repair 2015, 29, 677–689. [Google Scholar] [CrossRef]
- Merighi, A.; Bardoni, R.; Salio, C.; Lossi, L.; Ferrini, F.; Prandini, M.; Zonta, M.; Gustincich, S.; Carmignoto, G. Presynaptic functional trkB receptors mediate the release of excitatory neurotransmitters from primary afferent terminals in lamina II (substantia gelatinosa) of postnatal rat spinal cord. Dev. Neurobiol. 2008, 68, 457–475. [Google Scholar] [CrossRef]
- Pezet, S.; Cunningham, J.; Patel, J.; Grist, J.; Gavazzi, I.; Lever, I.J.; Malcangio, M. BDNF modulates sensory neuron synaptic activity by a facilitation of GABA transmission in the dorsal horn. Mol. Cell. Neurosci. 2002, 21, 51–62. [Google Scholar] [CrossRef]
- Smith, P.A. BDNF: No gain without pain? Neuroscience 2014, 283, 107–123. [Google Scholar] [CrossRef]
- Zhou, L.-J.; Zhong, Y.; Ren, W.-J.; Li, Y.-Y.; Zhang, T.; Liu, X.-G. BDNF induces late-phase LTP of C-fiber evoked field potentials in rat spinal dorsal horn. Exp. Neurol. 2008, 212, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Coull, J.A.M.; Beggs, S.; Boudreau, D.; Boivin, D.; Tsuda, M.; Inoue, K.; Gravel, C.; Salter, M.W.; De Koninck, Y. BDNF from microglia causes the shift in neuronal anion gradient underlying neuropathic pain. Nature 2005, 438, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.; Brambilla, R.; Thomas, K.L. A simple role for BDNF in learning and memory? Front. Mol. Neurosci. 2010, 3, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turrigiano, G.G. The Self-Tuning Neuron: Synaptic Scaling of Excitatory Synapses. Cell 2008, 135, 422–435. [Google Scholar] [CrossRef] [Green Version]
- Rutherford, L.C.; Nelson, S.B.; Turrigiano, G.G. BDNF has opposite effects on the quantal amplitude of pyramidal neuron and interneuron excitatory synapses. Neuron 1998, 21, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Desai, N.S.; Rutherford, L.C.; Turrigiano, G.G. BDNF regulates the intrinsic excitability of cortical neurons. Learn Mem. 1999, 6, 284–291. [Google Scholar] [CrossRef]
- Schinder, A.F.; Berninger, B.; Poo, M. Postsynaptic target specificity of neurotrophin-induced presynaptic potentiation. Neuron 2000, 25, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Rivera, C.; Voipio, J.; Thomas-Crusells, J.; Li, H.; Emri, Z.; Sipila, S.; Payne, J.A.; Minichiello, L.; Saarma, M.; Kaila, K. Mechanism of activity-dependent downregulation of the neuron-specific K-Cl cotransporter KCC2. J. Neurosci. 2004, 24, 4683–4691. [Google Scholar] [CrossRef] [Green Version]
- Garraway, S.M.; Petruska, J.C.; Mendell, L.M. BDNF sensitizes the response of lamina II neurons to high threshold primary afferent inputs. Eur. J. Neurosci. 2003, 18, 2467–2476. [Google Scholar] [CrossRef]
- Ferguson, A.R.; Bolding, K.A.; Huie, J.R.; Hook, M.A.; Santillano, D.R.; Miranda, R.C.; Grau, J.W. Group I metabotropic glutamate receptors control metaplasticity of spinal cord learning through a protein kinase C-dependent mechanism. J. Neurosci. 2008, 28, 11939–11949. [Google Scholar] [CrossRef]
- Abraham, W.C. Metaplasticity: Tuning synapses and networks for plasticity. Nat. Rev. Neurosci. 2008, 9, 387. [Google Scholar] [CrossRef]
- Bruce, J.C.; Oatway, M.A.; Weaver, L.C. Chronic pain after clip-compression injury of the rat spinal cord. Exp. Neurol. 2002, 178, 33–48. [Google Scholar] [CrossRef]
- Hains, B.C.; Everhart, A.W.; Fullwood, S.D.; Hulsebosch, C.E. Changes in serotonin, serotonin transporter expression and serotonin denervation supersensitivity: Involvement in chronic central pain after spinal hemisection in the rat. Exp. Neurol. 2002, 175, 347–362. [Google Scholar] [CrossRef]
- Oatway, M.A.; Chen, Y.; Weaver, L.C. The 5-HT3 receptor facilitates at-level mechanical allodynia following spinal cord injury. Pain 2004, 110, 259–268. [Google Scholar] [CrossRef]
- Chen, Y.; Oatway, M.A.; Weaver, L.C. Blockade of the 5-HT3 receptor for days causes sustained relief from mechanical allodynia following spinal cord injury. J. Neurosci. Res. 2009, 87, 418–424. [Google Scholar] [CrossRef]
- Wang, Y.X.; Bowersox, S.S.; Pettus, M.; Gao, D. Antinociceptive properties of fenfluramine, a serotonin reuptake inhibitor, in a rat model of neuropathy. J. Pharmacol. Exp. Ther. 1999, 291, 1008–1016. [Google Scholar] [PubMed]
- Irvine, K.A.; Sahbaie, P.; Ferguson, A.R.; Clark, J.D. Enhanced descending pain facilitation in acute traumatic brain injury. Exp. Neurol. 2019, 320, 112976. [Google Scholar] [CrossRef] [PubMed]
- Hultborn, H.; Denton, M.E.; Wienecke, J.; Nielsen, J.B. Variable amplification of synaptic input to cat spinal motoneurones by dendritic persistent inward current. J. Physiol. 2003, 552, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, B.L.; Martín-Cora, F.J.; Fornal, C.A. Activity of medullary serotonergic neurons in freely moving animals. Brain Res. Rev. 2002, 40, 45–52. [Google Scholar] [CrossRef]
- Perrier, J.F.; Delgado-Lezama, R. Synaptic release of serotonin induced by stimulation of the raphe nucleus promotes plateau potentials in spinal motoneurons of the adult turtle. J. Neurosci. 2005, 25, 7993–7999. [Google Scholar] [CrossRef] [Green Version]
- Hounsgaard, J.; Hultborn, H.; Jespersen, B.; Kiehn, O. Bistability of alpha-motoneurones in the decerebrate cat and in the acute spinal cat after intravenous 5-hydroxytryptophan. J. Physiol. 1988, 405, 345–367. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.R.; Gorassini, M.A.; Bennett, D.J. Role of persistent sodium and calcium currents in motoneuron firing and spasticity in chronic spinal rats. J. Neurophysiol. 2004, 91, 767–783. [Google Scholar] [CrossRef] [Green Version]
- Bennett, D.J.; Sanelli, L.; Cooke, C.L.; Harvey, P.J.; Gorassini, M.A. Spastic long-lasting reflexes in the awake rat after sacral spinal cord injury. J. Neurophysiol. 2004, 91, 2247–2258. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, J.M.; Murray, K.C.; Li, Y.; Chan, K.M.; Finlay, M.G.; Bennett, D.J.; Gorassini, M.A. Constitutively active 5-HT2/α1 receptors facilitate muscle spasms after human spinal cord injury. J. Neurophysiol. 2013, 109, 1473–1484. [Google Scholar] [CrossRef] [Green Version]
- Wainberg, M.; Barbeau, H.; Gauthier, S. The effects of cyproheptadine on locomotion and on spasticity in patients with spinal cord injuries. J. Neurol. Neurosurg. Psychiatry 1990, 53, 754–763. [Google Scholar] [CrossRef] [Green Version]
- Ramage, A.G. Central cardiovascular regulation and 5-hydroxytryptamine receptors. Brain Res. Bull. 2001, 56, 425–439. [Google Scholar] [CrossRef]
- Ramage, A.G.; Villalon, C.M. 5-hydroxytryptamine and cardiovascular regulation. Trends Pharmacol. Sci. 2008, 29, 472–481. [Google Scholar] [CrossRef]
- Coote, J.H. Bulbospinal serotonergic pathways in the control of blood pressure. J. Cardiovasc. Pharmacol. 1990, 15 (Suppl. S7), S35–S41. [Google Scholar] [CrossRef]
- Lewis, D.I.; Coote, J.H. The influence of 5-hydroxytryptamine agonists and antagonists on identified sympathetic preganglionic neurones in the rat, in vivo. Br. J. Pharmacol. 1990, 99, 667–672. [Google Scholar] [CrossRef] [Green Version]
- Chalmers, J.P.; Pilowsky, P.M.; Minson, J.B.; Kapoor, V.; Mills, E.; West, M.J. Central serotonergic mechanisms in hypertension. Am. J. Hypertens. 1988, 1, 79–83. [Google Scholar] [CrossRef]
- Brock, J.A.; Yeoh, M.; McLachlan, E.M. Enhanced neurally evoked responses and inhibition of norepinephrine reuptake in rat mesenteric arteries after spinal transection. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H398–H405. [Google Scholar] [CrossRef]
- Hou, S.; Duale, H.; Rabchevsky, A.G. Intraspinal sprouting of unmyelinated pelvic afferents after complete spinal cord injury is correlated with autonomic dysreflexia induced by visceral pain. Neuroscience 2009, 159, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Cameron, A.A.; Smith, G.M.; Randall, D.C.; Brown, D.R.; Rabchevsky, A.G. Genetic manipulation of intraspinal plasticity after spinal cord injury alters the severity of autonomic dysreflexia. J. Neurosci. 2006, 26, 2923–2932. [Google Scholar] [CrossRef] [Green Version]
- Weaver, L.C.; Verghese, P.; Bruce, J.C.; Fehlings, M.G.; Krenz, N.R.; Marsh, D.R. Autonomic dysreflexia and primary afferent sprouting after clip-compression injury of the rat spinal cord. J. Neurotrauma 2001, 18, 1107–1119. [Google Scholar] [CrossRef]
- Llewellyn-Smith, I.J.; Weaver, L.C. Changes in synaptic inputs to sympathetic preganglionic neurons after spinal cord injury. J. Comp. Neurol. 2001, 435, 226–240. [Google Scholar] [CrossRef]
- Krassioukov, A.V.; Johns, D.G.; Schramm, L.P. Sensitivity of sympathetically correlated spinal interneurons, renal sympathetic nerve activity, and arterial pressure to somatic and visceral stimuli after chronic spinal injury. J. Neurotrauma 2002, 19, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Rabchevsky, A.G. Segmental organization of spinal reflexes mediating autonomic dysreflexia after spinal cord injury. Prog. Brain Res. 2006, 152, 265–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, F.M.; Patel, S.P.; Rabchevsky, A.G. Intraspinal Plasticity Associated With the Development of Autonomic Dysreflexia After Complete Spinal Cord Injury. Front. Cell. Neurosci. 2019, 13, 10. [Google Scholar] [CrossRef] [PubMed]
- Cormier, C.M.; Mukhida, K.; Walker, G.; Marsh, D.R. Development of autonomic dysreflexia after spinal cord injury is associated with a lack of serotonergic axons in the intermediolateral cell column. J. Neurotrauma 2010, 27, 1805–1818. [Google Scholar] [CrossRef]
- Hou, S.; Tom, V.J.; Graham, L.; Lu, P.; Blesch, A. Partial restoration of cardiovascular function by embryonic neural stem cell grafts after complete spinal cord transection. J. Neurosci. 2013, 33, 17138–17149. [Google Scholar] [CrossRef] [Green Version]
- Hou, S.P.; Saltos, T.M.; Mironets, E.; Trueblood, C.T.; Connors, T.M.; Tom, V.J. Grafting Embryonic Raphe Neurons Reestablishes Serotonergic Regulation of Sympathetic Activity to Improve Cardiovascular Function after Spinal Cord Injury. J. Neurosci. 2020, 40, 1248–1264. [Google Scholar] [CrossRef]
- Trueblood, C.T.; Iredia, I.W.; Collyer, E.S.; Tom, V.J.; Hou, S. Development of Cardiovascular Dysfunction in a Rat Spinal Cord Crush Model and Responses to Serotonergic Interventions. J. Neurotrauma 2019, 36, 1478–1486. [Google Scholar] [CrossRef]
- Lujan, H.L.; DiCarlo, S.E. Direct comparison of cervical and high thoracic spinal cord injury reveals distinct autonomic and cardiovascular consequences. J. Appl. Physiol. 2020, 128, 554–564. [Google Scholar] [CrossRef]
- Mathias, C.J.; Christensen, N.J.; Corbett, J.L.; Frankel, H.L.; Spalding, J.M. Plasma catecholamines during paroxysmal neurogenic hypertension in quadriplegic man. Circ. Res. 1976, 39, 204–208. [Google Scholar] [CrossRef] [Green Version]
- Leman, S.; Bernet, F.; Sequeira, H. Autonomic dysreflexia increases plasma adrenaline level in the chronic spinal cord-injured rat. Neurosci. Lett. 2000, 286, 159–162. [Google Scholar] [CrossRef]
- Garshick, E.; Kelley, A.; Cohen, S.A.; Garrison, A.; Tun, C.G.; Gagnon, D.; Brown, R. A prospective assessment of mortality in chronic spinal cord injury. Spinal Cord 2005, 43, 408–416. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.C.; Chen, Y.C.; Liu, L.; Chen, T.J.; Huang, W.C.; Cheng, H.; Tung-Ping, S. Increased risk of stroke after spinal cord injury: A nationwide 4-year follow-up cohort study. Neurology 2012, 78, 1051–1057. [Google Scholar] [CrossRef]
- Lucin, K.M.; Sanders, V.M.; Jones, T.B.; Malarkey, W.B.; Popovich, P.G. Impaired antibody synthesis after spinal cord injury is level dependent and is due to sympathetic nervous system dysregulation. Exp. Neurol. 2007, 207, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Lucin, K.M.; Sanders, V.M.; Popovich, P.G. Stress hormones collaborate to induce lymphocyte apoptosis after high level spinal cord injury. J. Neurochem. 2009, 110, 1409–1421. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Guan, Z.; Reader, B.; Shawler, T.; Mandrekar-Colucci, S.; Huang, K.; Weil, Z.; Bratasz, A.; Wells, J.; Powell, N.D.; et al. Autonomic Dysreflexia Causes Chronic Immune Suppression after Spinal Cord Injury. J. Neurosci. 2013, 33, 12970–12981. [Google Scholar] [CrossRef]
- Karunakaran, K.D.; Yuan, R.; He, J.; Zhao, J.; Cui, J.L.; Zang, Y.F.; Zhang, Z.; Alvarez, T.L.; Biswal, B.B. Resting-State Functional Connectivity of the Thalamus in Complete Spinal Cord Injury. Neurorehabilit. Neural Repair 2020, 34, 122–133. [Google Scholar] [CrossRef]
- Chou, X.L.; Fang, Q.; Yan, L.; Zhong, W.; Peng, B.; Li, H.; Wei, J.; Tao, H.W.; Zhang, L.I. Contextual and cross-modality modulation of auditory cortical processing through pulvinar mediated suppression. eLife 2020, 9, e54157. [Google Scholar] [CrossRef]
- Fang, Q.; Chou, X.L.; Peng, B.; Zhong, W.; Zhang, L.I.; Tao, H.W. A Differential Circuit via Retino-Colliculo-Pulvinar Pathway Enhances Feature Selectivity in Visual Cortex through Surround Suppression. Neuron 2020, 105, 355–369.e6. [Google Scholar] [CrossRef]
- Ibrahim, L.A.; Mesik, L.; Ji, X.Y.; Fang, Q.; Li, H.F.; Li, Y.T.; Zingg, B.; Zhang, L.I.; Tao, H.W. Cross-Modality Sharpening of Visual Cortical Processing through Layer-1-Mediated Inhibition and Disinhibition. Neuron 2016, 89, 1031–1045. [Google Scholar] [CrossRef] [Green Version]
- Monckton, J.E.; McCormick, D.A. Neuromodulatory role of serotonin in the ferret thalamus. J. Neurophysiol. 2002, 87, 2124–2136. [Google Scholar] [CrossRef] [Green Version]
- Goodman, L.S.; Gillman, A.; Brunton Laurence, L.; Lazo John, S.; Parker Keith, L. Goodman & Gilman’s: The Pharmacological Basis of Therapeutics; McGraw-Hill Education: New York, NY, USA, 2006. [Google Scholar]
- Lu, Y.; Zheng, J.; Xiong, L.; Zimmermann, M.; Yang, J. Spinal cord injury-induced attenuation of GABAergic inhibition in spinal dorsal horn circuits is associated with down-regulation of the chloride transporter KCC2 in rat. J. Physiol. 2008, 586, 5701–5715. [Google Scholar] [CrossRef]
- Baba, H.; Ji, R.R.; Kohno, T.; Moore, K.A.; Ataka, T.; Wakai, A.; Okamoto, M.; Woolf, C.J. Removal of GABAergic inhibition facilitates polysynaptic A fiber-mediated excitatory transmission to the superficial spinal dorsal horn. Mol. Cell. Neurosci. 2003, 24, 818–830. [Google Scholar] [CrossRef]
- Dougherty, K.J.; Hochman, S. Spinal cord injury causes plasticity in a subpopulation of lamina I GABAergic interneurons. J. Neurophysiol. 2008, 100, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Roberts, L.A.; Beyer, C.; Komisaruk, B.R. Nociceptive responses to altered GABAergic activity at the spinal cord. Life Sci. 1986, 39, 1667–1674. [Google Scholar] [CrossRef]
- Sivilotti, L.; Woolf, C.J. The contribution of GABAA and glycine receptors to central sensitization: Disinhibition and touch-evoked allodynia in the spinal cord. J. Neurophysiol. 1994, 72, 169–179. [Google Scholar] [CrossRef]
- Zhang, Z.; Hefferan, M.P.; Loomis, C.W. Topical bicuculline to the rat spinal cord induces highly localized allodynia that is mediated by spinal prostaglandins. Pain 2001, 92, 351–361. [Google Scholar] [CrossRef]
- Sorkin, L.S.; Puig, S.; Jones, D.L. Spinal bicuculline produces hypersensitivity of dorsal horn neurons: Effects of excitatory amino acid antagonists. Pain 1998, 77, 181–190. [Google Scholar] [CrossRef]
- Hwang, J.H.; Yaksh, T.L. The effect of spinal GABA receptor agonists on tactile allodynia in a surgically-induced neuropathic pain model in the rat. Pain 1997, 70, 15–22. [Google Scholar] [CrossRef]
- Kaneko, M.; Hammond, D.L. Role of spinal gamma-aminobutyric acidA receptors in formalin-induced nociception in the rat. J. Pharmacol. Exp. Ther. 1997, 282, 928–938. [Google Scholar]
- Jergova, S.; Hentall, I.D.; Gajavelli, S.; Varghese, M.S.; Sagen, J. Intraspinal transplantation of GABAergic neural progenitors attenuates neuropathic pain in rats: A pharmacologic and neurophysiological evaluation. Exp. Neurol. 2012, 234, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Jolivalt, C.G.; Lee, C.A.; Ramos, K.M.; Calcutt, N.A. Allodynia and hyperalgesia in diabetic rats are mediated by GABA and depletion of spinal potassium-chloride co-transporters. Pain 2008, 140, 48–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaila, K.; Price, T.J.; Payne, J.A.; Puskarjov, M.; Voipio, J. Cation-chloride cotransporters in neuronal development, plasticity and disease. Nat. Rev. Neurosci. 2014, 15, 637–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiumelli, H.; Woodin, M.A. Role of activity-dependent regulation of neuronal chloride homeostasis in development. Curr. Opin. Neurobiol. 2007, 17, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Cramer, S.W.; Baggott, C.; Cain, J.; Tilghman, J.; Allcock, B.; Miranpuri, G.; Rajpal, S.; Sun, D.; Resnick, D. The role of cation-dependent chloride transporters in neuropathic pain following spinal cord injury. Mol. Pain 2008, 4, 36. [Google Scholar] [CrossRef] [Green Version]
- Medina, I.; Friedel, P.; Rivera, C.; Kahle, K.T.; Kourdougli, N.; Uvarov, P.; Pellegrino, C. Current view on the functional regulation of the neuronal K+–Cl− cotransporter KCC2. Front. Cell. Neurosci. 2014, 8, 18. [Google Scholar] [CrossRef] [Green Version]
- Kahle, K.T.; Deeb, T.Z.; Puskarjov, M.; Silayeva, L.; Liang, B.; Kaila, K.; Moss, S.J. Modulation of neuronal activity by phosphorylation of the K–CI cotransporter KCC2. Trends Neurosci. 2013, 36, 726–737. [Google Scholar] [CrossRef] [Green Version]
- Ben-Ari, Y. Excitatory actions of GABA during development: The nature of the nurture. Nat. Rev. Neurosci. 2002, 3, 728–739. [Google Scholar] [CrossRef]
- Rivera, C.; Voipio, J.; Kaila, K. Two developmental switches in GABAergic signalling: The K+–Cl− cotransporter KCC2 and carbonic anhydrase CAVII. J. Physiol. 2005, 562, 27–36. [Google Scholar] [CrossRef]
- Ferrini, F.; De Koninck, Y. Microglia control neuronal network excitability via BDNF signalling. Neural Plast. 2013, 2013, 429815. [Google Scholar] [CrossRef]
- Ostroumov, A.; Thomas, A.M.; Kimmey, B.A.; Karsch, J.S.; Doyon, W.M.; Dani, J.A. Stress Increases Ethanol Self-Administration via a Shift toward Excitatory GABA Signaling in the Ventral Tegmental Area. Neuron 2016, 92, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Ostroumov, A.; Dani, J.A. Convergent Neuronal Plasticity and Metaplasticity Mechanisms of Stress, Nicotine, and Alcohol. Annu. Rev. Pharmacol. Toxicol. 2018, 58, 547–566. [Google Scholar] [CrossRef]
- Wang, W.; Gong, N.; Xu, T.L. Downregulation of KCC2 following LTP-contributes to EPSP-spike potentiation in rat hippocampus. Biochem. Biophys. Res. Commun. 2006, 343, 1209–1215. [Google Scholar] [CrossRef]
- Ostroumov, A.; Dani, J.A. Inhibitory Plasticity of Mesocorticolimbic Circuits in Addiction and Mental Illness. Trends Neurosci. 2018, 41, 898–910. [Google Scholar] [CrossRef]
- Drew, G.M.; Siddall, P.J.; Duggan, A.W. Mechanical allodynia following contusion injury of the rat spinal cord is associated with loss of GABAergic inhibition in the dorsal horn. Pain 2004, 109, 379–388. [Google Scholar] [CrossRef]
- Boulenguez, P.; Liabeuf, S.; Bos, R.; Bras, H.; Jean-Xavier, C.; Brocard, C.; Stil, A.; Darbon, P.; Cattaert, D.; Delpire, E.; et al. Down-regulation of the potassium-chloride cotransporter KCC2 contributes to spasticity after spinal cord injury. Nat. Med. 2010, 16, 302–307. [Google Scholar] [CrossRef]
- Hasbargen, T.; Ahmed, M.M.; Miranpuri, G.; Li, L.; Kahle, K.T.; Resnick, D.; Sun, D. Role of NKCC1 and KCC2 in the development of chronic neuropathic pain following spinal cord injury. Ann. N. Y. Acad. Sci. 2010, 1198, 168–172. [Google Scholar] [CrossRef]
- Gackière, F.; Vinay, L. Serotonergic modulation of post-synaptic inhibition and locomotor alternating pattern in the spinal cord. Front. Neural Circuits 2014, 8, 102. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, A.R.; Washburn, S.N.; Crown, E.D.; Grau, J.W. GABA(A) receptor activation is involved in noncontingent shock inhibition of instrumental conditioning in spinal rats. Behav. Neurosci. 2003, 117, 799–812. [Google Scholar] [CrossRef]
- Chen, B.; Li, Y.; Yu, B.; Zhang, Z.; Brommer, B.; Williams, P.R.; Liu, Y.; Hegarty, S.V.; Zhou, S.; Zhu, J.; et al. Reactivation of Dormant Relay Pathways in Injured Spinal Cord by KCC2 Manipulations. Cell 2018, 174, 1599. [Google Scholar] [CrossRef]
- Beverungen, H.; Klaszky, S.C.; Klaszky, M.; Cote, M.P. Rehabilitation Decreases Spasticity by Restoring Chloride Homeostasis through the Brain-Derived Neurotrophic Factor-KCC2 Pathway after Spinal Cord Injury. J. Neurotrauma 2020, 37, 846–859. [Google Scholar] [CrossRef]
- Cote, M.P.; Gandhi, S.; Zambrotta, M.; Houle, J.D. Exercise Modulates Chloride Homeostasis after Spinal Cord Injury. J. Neurosci. 2014, 34, 8976–8987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee-Hotta, S.; Uchiyama, Y.; Kametaka, S. Role of the BDNF-TrkB pathway in KCC2 regulation and rehabilitation following neuronal injury: A mini review. Neurochem. Int. 2019, 128, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, L.Y.; Xu, T.L. Reduced potassium-chloride co-transporter expression in spinal cord dorsal horn neurons contributes to inflammatory pain hypersensitivity in rats. Neuroscience 2008, 152, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Beggs, S.; Salter, M.W. Microglia-neuronal signalling in neuropathic pain hypersensitivity 2.0. Curr. Opin. Neurobiol. 2010, 20, 474–480. [Google Scholar] [CrossRef] [Green Version]
- Shulga, A.; Thomas-Crusells, J.; Sigl, T.; Blaesse, A.; Mestres, P.; Meyer, M.; Yan, Q.; Kaila, K.; Saarma, M.; Rivera, C.; et al. Posttraumatic GABA(A)-mediated [Ca2+](i) increase is essential for the induction of brain-derived neurotrophic factor-dependent survival of mature central neurons. J. Neurosci. 2008, 28, 6996–7005. [Google Scholar] [CrossRef]
- King, T.; Vera-Portocarrero, L.; Gutierrez, T.; Vanderah, T.W.; Dussor, G.; Lai, J.; Fields, H.L.; Porreca, F. Unmasking the tonic-aversive state in neuropathic pain. Nat. Neurosci. 2009, 12, 1364–1366. [Google Scholar] [CrossRef] [Green Version]
- Bos, R.; Sadlaoud, K.; Boulenguez, P.; Buttigieg, D.; Liabeuf, S.; Brocard, C.; Haase, G.; Bras, H.; Vinay, L. Activation of 5-HT2A receptors upregulates the function of the neuronal K–Cl cotransporter KCC2. Proc. Natl. Acad. Sci. USA 2013, 110, 348–353. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Brualla, I.; Boulenguez, P.; Brocard, C.; Liabeuf, S.; Viallat-Lieutaud, A.; Navarro, X.; Udina, E.; Brocard, F. Activation of 5-HT(2A) Receptors Restores KCC2 Function and Reduces Neuropathic Pain after Spinal Cord Injury. Neuroscience 2018, 387, 48–57. [Google Scholar] [CrossRef]
- Kimmey, B.A.; Ostroumov, A.; Dani, J.A. 5-HT2A receptor activation normalizes stress-induced dysregulation of GABAergic signaling in the ventral tegmental area. Proc. Natl. Acad. Sci. USA 2019, 116, 27028–27034. [Google Scholar] [CrossRef]
- Jean-Xavier, C.; Sharples, S.A.; Mayr, K.A.; Lognon, A.P.; Whelan, P.J. Retracing your footsteps: Developmental insights to spinal network plasticity following injury. J. Neurophysiol. 2018, 119, 521–536. [Google Scholar] [CrossRef]
- Jean-Xavier, C.; Pflieger, J.F.; Liabeuf, S.; Vinay, L. Inhibitory postsynaptic potentials in lumbar motoneurons remain depolarizing after neonatal spinal cord transection in the rat. J. Neurophysiol. 2006, 96, 2274–2281. [Google Scholar] [CrossRef]
- Rekling, J.C.; Funk, G.D.; Bayliss, D.A.; Dong, X.W.; Feldman, J.L. Synaptic control of motoneuronal excitability. Physiol. Rev. 2000, 80, 767–852. [Google Scholar] [CrossRef]
- Rank, M.M.; Murray, K.C.; Stephens, M.J.; D’Amico, J.; Gorassini, M.A.; Bennett, D.J. Adrenergic receptors modulate motoneuron excitability, sensory synaptic transmission and muscle spasms after chronic spinal cord injury. J. Neurophysiol. 2011, 105, 410–422. [Google Scholar] [CrossRef] [Green Version]
- Al Dera, H.; Brock, J.A. Changes in sympathetic neurovascular function following spinal cord injury. Auton. Neurosci. 2018, 209, 25–36. [Google Scholar] [CrossRef]
- Streijger, F.; So, K.; Manouchehri, N.; Gheorghe, A.; Okon, E.B.; Chan, R.M.; Ng, B.; Shortt, K.; Sekhon, M.S.; Griesdale, D.E.; et al. A Direct Comparison between Norepinephrine and Phenylephrine for Augmenting Spinal Cord Perfusion in a Porcine Model of Spinal Cord Injury. J. Neurotrauma 2018, 35, 1345–1357. [Google Scholar] [CrossRef]
- Miyazato, M.; Oshiro, T.; Chancellor, M.B.; de Groat, W.C.; Yoshimura, N.; Saito, S. An alpha1-adrenoceptor blocker terazosin improves urine storage function in the spinal cord in spinal cord injured rats. Life Sci. 2013, 92, 125–130. [Google Scholar] [CrossRef]
- Mitsui, T.; Shumsky, J.S.; Lepore, A.C.; Murray, M.; Fischer, I. Transplantation of neuronal and glial restricted precursors into contused spinal cord improves bladder and motor functions, decreases thermal hypersensitivity, and modifies intraspinal circuitry. J. Neurosci. 2005, 25, 9624–9636. [Google Scholar] [CrossRef] [Green Version]
- Pertovaara, A. Noradrenergic pain modulation. Prog. Neurobiol. 2006, 80, 53–83. [Google Scholar] [CrossRef]
- Pertovaara, A.; Kauppila, T.; Jyväsjärvi, E.; Kalso, E. Involvement of supraspinal and spinal segmental alpha-2-adrenergic mechanisms in the medetomidine-induced antinociception. Neuroscience 1991, 44, 705–714. [Google Scholar] [CrossRef]
- Kwaśniewska, A.; Miazga, K.; Majczyński, H.; Jordan, L.M.; Zawadzka, M.; Sławińska, U. Noradrenergic Components of Locomotor Recovery Induced by Intraspinal Grafting of the Embryonic Brainstem in Adult Paraplegic Rats. Int. J. Mol. Sci. 2020, 21, 5520. [Google Scholar] [CrossRef]
- Xu, P.; Guo, S.; Xie, Y.; Liu, Z.; Liu, C.; Zhang, X.; Yang, D.; Gong, H.; Chen, Y.; Du, L.; et al. Effects of highly selective sympathectomy on neurogenic bowel dysfunction in spinal cord injury rats. Sci. Rep. 2021, 11, 15892. [Google Scholar] [CrossRef]
- Li, Y.; Harvey, P.J.; Li, X.; Bennett, D.J. Spastic long-lasting reflexes of the chronic spinal rat studied in vitro. J. Neurophysiol. 2004, 91, 2236–2246. [Google Scholar] [CrossRef] [Green Version]
- Corleto, J.A.; Bravo-Hernández, M.; Kamizato, K.; Kakinohana, O.; Santucci, C.; Navarro, M.R.; Platoshyn, O.; Cizkova, D.; Lukacova, N.; Taylor, J.; et al. Thoracic 9 Spinal Transection-Induced Model of Muscle Spasticity in the Rat: A Systematic Electrophysiological and Histopathological Characterization. PLoS ONE 2015, 10, e0144642. [Google Scholar] [CrossRef]
- Gao, J.; Sun, Z.; Xiao, Z.; Du, Q.; Niu, X.; Wang, G.; Chang, Y.W.; Sun, Y.; Sun, W.; Lin, A.; et al. Dexmedetomidine modulates neuroinflammation and improves outcome via alpha2-adrenergic receptor signaling after rat spinal cord injury. Br. J. Anaesth. 2019, 123, 827–838. [Google Scholar] [CrossRef] [Green Version]
- DeVeau, K.M.; Harman, K.A.; Squair, J.W.; Krassioukov, A.V.; Magnuson, D.S.K.; West, C.R. A comparison of passive hindlimb cycling and active upper-limb exercise provides new insights into systolic dysfunction after spinal cord injury. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H861–H870. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, N.; Gotoh, D.; Nishimoto, M.; Hashimoto, M.; Saito, T.; Fujita, K.; Hirayama, A.; Yoshimura, N.; Uemura, H. Efficacy of vibegron, a novel β3-adrenoreceptor agonist, for lower urinary tract dysfunction in mice with spinal cord injury. Int. J. Urol. 2021, 28, 1068–1072. [Google Scholar] [CrossRef]
- Scholpa, N.E.; Simmons, E.C.; Tilley, D.G.; Schnellmann, R.G. β(2)-adrenergic receptor-mediated mitochondrial biogenesis improves skeletal muscle recovery following spinal cord injury. Exp. Neurol. 2019, 322, 113064. [Google Scholar] [CrossRef]
- Scholpa, N.E.; Williams, H.; Wang, W.; Corum, D.; Narang, A.; Tomlinson, S.; Sullivan, P.G.; Rabchevsky, A.G.; Schnellmann, R.G. Pharmacological Stimulation of Mitochondrial Biogenesis Using the Food and Drug Administration-Approved β(2)-Adrenoreceptor Agonist Formoterol for the Treatment of Spinal Cord Injury. J. Neurotrauma 2019, 36, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Fuchigami, T.; Kakinohana, O.; Hefferan, M.P.; Lukacova, N.; Marsala, S.; Platoshyn, O.; Sugahara, K.; Yaksh, T.L.; Marsala, M. Potent suppression of stretch reflex activity after systemic or spinal delivery of tizanidine in rats with spinal ischemia-induced chronic spastic paraplegia. Neuroscience 2011, 194, 160–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida, H.; Yamauchi, H.; Ito, H.; Akino, H.; Yokoyama, O. α1D-Adrenoceptor blockade increases voiding efficiency by improving external urethral sphincter activity in rats with spinal cord injury. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R971–R978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas, D.A.; Chancellor, M.B.; Huang, B.; Salzman, S.K. Autonomic dysreflexia in a rat model spinal cord injury and the effect of pharmacologic agents. Neurourol. Urodyn. 1995, 14, 141–152. [Google Scholar] [CrossRef]
- Morrison, S.F.; Ge, Y.Z. Adrenergic modulation of a spinal sympathetic reflex in the rat. J. Pharmacol. Exp. Ther. 1995, 273, 380–385. [Google Scholar]
- Yaksh, T.L.; Pogrel, J.W.; Lee, Y.W.; Chaplan, S.R. Reversal of nerve ligation-induced allodynia by spinal alpha-2 adrenoceptor agonists. J. Pharmacol. Exp. Ther. 1995, 272, 207–214. [Google Scholar]
- Rong, H.; Zhao, Z.; Feng, J.; Lei, Y.; Wu, H.; Sun, R.; Zhang, Z.; Hou, B.; Zhang, W.; Sun, Y.; et al. The effects of dexmedetomidine pretreatment on the pro- and anti-inflammation systems after spinal cord injury in rats. Brain Behav. Immun. 2017, 64, 195–207. [Google Scholar] [CrossRef]
- Molina, C.; Herrero, J.F. The influence of the time course of inflammation and spinalization on the antinociceptive activity of the alpha2-adrenoceptor agonist medetomidine. Eur. J. Pharmacol. 2006, 532, 50–60. [Google Scholar] [CrossRef]
- Sakitani, N.; Iwasawa, H.; Nomura, M.; Miura, Y.; Kuroki, H.; Ozawa, J.; Moriyama, H. Mechanical Stress by Spasticity Accelerates Fracture Healing After Spinal Cord Injury. Calcif. Tissue Int. 2017, 101, 384–395. [Google Scholar] [CrossRef]
- Lemmens, S.; Nelissen, S.; Dooley, D.; Geurts, N.; Peters, E.M.J.; Hendrix, S. Stress Pathway Modulation Is Detrimental or Ineffective for Functional Recovery after Spinal Cord Injury in Mice. J. Neurotrauma 2020, 37, 564–571. [Google Scholar] [CrossRef]
- Karatas, Y.; Cengiz, S.L.; Esen, H.; Toker, A.; Savas, C. Effect of Carvedilol on Secondary Damage in Experimental Spinal Cord Injury in Rats. Turk. Neurosurg. 2015, 25, 930–935. [Google Scholar] [CrossRef]
- Liu, D.; Huang, Y.; Li, B.; Jia, C.; Liang, F.; Fu, Q. Carvedilol promotes neurological function, reduces bone loss and attenuates cell damage after acute spinal cord injury in rats. Clin. Exp. Pharmacol. Physiol. 2015, 42, 202–212. [Google Scholar] [CrossRef]
- Vega, J.L.; Ganea, D.; Jonakait, G.M. Acute down-regulation of antibody production following spinal cord injury: Role of systemic catecholamines. J. Neuropathol. Exp. Neurol. 2003, 62, 848–854. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Hartung, J.E.; Bortsov, A.V.; Kim, S.; O’Buckley, S.C.; Kozlowski, J.; Nackley, A.G. Sustained stimulation of β(2)- and β(3)-adrenergic receptors leads to persistent functional pain and neuroinflammation. Brain Behav. Immun. 2018, 73, 520–532. [Google Scholar] [CrossRef]
- Schoultz, T.W.; Deluca, D.C.; Reding, D.L. Norepinephrine Levels in the Traumatized Spinal Cord of Catecholamine-Depleted Cats. Brain Res. 1976, 109, 367–374. [Google Scholar] [CrossRef]
- Osterholm, J.L.; Mathews, G.J. Altered norepinephrine metabolism, following experimental spinal cord injury. 2. Protection against traumatic spinal cord hemorrhagic necrosis by norepinephrine synthesis blockade with alpha methyl tyrosine. J. Neurosurg. 1972, 36, 395–401. [Google Scholar] [CrossRef]
- Vise, W.M.; Yashon, D.; Hunt, W.E. Mechanisms of norepinephrine accumulation within sites of spinal cord injury. J. Neurosurg. 1974, 40, 76–82. [Google Scholar] [CrossRef]
- Alderman, J.L.; Osterholm, J.L.; Damore, B.R.; Williams, H.D.; Irvin, J.D. The Influence of the Adrenal Gland Upon Actue Spinal Cord Injury. Life Sci. 1980, 26, 1627–1632. [Google Scholar] [CrossRef]
- Crawford, R.A.; Griffiths, I.R.; McCulloch, J. The Effect of Norepinephrine on Spinal Cord Circulation and its Possible Implications in Pathogenesis of Acute Spinal Trauma. J. Neurosurg. 1977, 47, 567–576. [Google Scholar] [CrossRef]
- Rawe, S.E.; Lee, W.A.; Perot, P.L., Jr. The histopathology of experimental spinal cord trauma. The effect of systemic blood pressure. J. Neurosurg. 1978, 48, 1002–1007. [Google Scholar] [CrossRef]
- Rawe, S.E.; Roth, R.H.; Collins, W.F. Norepinephrine levels in experimental spinal cord trauma. Part 2: Histopathological study of hemorrhagic necrosis. J. Neurosurg. 1977, 46, 350–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurihara, M. Role of monoamines in experimental spinal cord injury in rats. Relationship between Na+–K+-ATPase and lipid peroxidation. J. Neurosurg. 1985, 62, 743–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osterholm, J.L.; Mathews, G.J. A proposed biochemical mechanism for traumatic spinal cord hemorrhagic necrosis. Successful therapy for severe injuries by metabolic blockade. Trans. Am. Neurol. Assoc. 1971, 96, 187–191. [Google Scholar] [PubMed]
- Osterholm, J.L. The pathophysiological response to spinal cord injury. The current status of related research. J. Neurosurg. 1974, 40, 5–33. [Google Scholar] [CrossRef]
- Kalous, A.; Osborne, P.B.; Keast, J.R. Acute and chronic changes in dorsal horn innervation by primary afferents and descending supraspinal pathways after spinal cord injury. J. Comp. Neurol. 2007, 504, 238–253. [Google Scholar] [CrossRef]
- Kalous, A.; Osborne, P.B.; Keast, J.R. Spinal cord compression injury in adult rats initiates changes in dorsal horn remodeling that may correlate with development of neuropathic pain. J. Comp. Neurol. 2009, 513, 668–684. [Google Scholar] [CrossRef]
- Hou, S.; DeFinis, J.H.; Daugherty, S.L.; Tang, C.; Weinberger, J.; de Groat, W.C. Deciphering Spinal Endogenous Dopaminergic Mechanisms That Modulate Micturition Reflexes in Rats with Spinal Cord Injury. eNeuro 2021, 8, 1–20. [Google Scholar] [CrossRef]
- Qiao, Y.; Brodnik, Z.D.; Zhao, S.; Trueblood, C.T.; Li, Z.; Tom, V.J.; España, R.A.; Hou, S. Spinal Dopaminergic Mechanisms Regulating the Micturition Reflex in Male Rats with Complete Spinal Cord Injury. J. Neurotrauma 2021, 38, 803–817. [Google Scholar] [CrossRef]
- Lahlou, S. Enhanced hypotensive response to intravenous apomorphine in chronic spinalized, conscious rats: Role of spinal dopamine D(1) and D(2) receptors. Neurosci. Lett. 2003, 349, 115–119. [Google Scholar] [CrossRef]
- Rodgers, H.M.; Patton, R.; Yow, J.; Zeczycki, T.N.; Kew, K.; Clemens, S.; Brewer, K.L. Morphine resistance in spinal cord injury-related neuropathic pain in rats is associated with alterations in dopamine and dopamine-related metabolomics. J. Pain 2021, in press. [Google Scholar] [CrossRef]
- Bao, Y.N.; Dai, W.L.; Fan, J.F.; Ma, B.; Li, S.S.; Zhao, W.L.; Yu, B.Y.; Liu, J.H. The dopamine D1-D2DR complex in the rat spinal cord promotes neuropathic pain by increasing neuronal excitability after chronic constriction injury. Exp. Mol. Med. 2021, 53, 235–249. [Google Scholar] [CrossRef]
- Rooney, K.F.; Sewell, R.D. Evaluation of selective actions of dopamine D-1 and D-2 receptor agonists and antagonists on opioid antinociception. Eur. J. Pharmacol. 1989, 168, 329–336. [Google Scholar] [CrossRef]
- Rodgers, H.M.; Yow, J.; Evans, E.; Clemens, S.; Brewer, K.L. Dopamine D1 and D3 receptor modulators restore morphine analgesia and prevent opioid preference in a model of neuropathic pain. Neuroscience 2019, 406, 376–388. [Google Scholar] [CrossRef]
- Khalilzadeh, M.; Hassanzadeh, F.; Aghamiri, H.; Dehpour, A.R.; Shafaroodi, H. Aripiprazole prevents from development of vincristine-induced neuropathic nociception by limiting neural NOS overexpression and NF-kB hyperactivation. Cancer Chemother. Pharm. 2020, 86, 393–404. [Google Scholar] [CrossRef]
- Guertin, P.A.; Ung, R.V.; Rouleau, P.; Steuer, I. Effects on locomotion, muscle, bone, and blood induced by a combination therapy eliciting weight-bearing stepping in nonassisted spinal cord-transected mice. Neurorehabilit. Neural Repair 2011, 25, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Mei, W.; Wang, Q.; Guo, R.; Liu, P.; Wang, Y.; Zhang, Z.; Wang, L. Role of Dehydrocorybulbine in Neuropathic Pain After Spinal Cord Injury Mediated by P2X4 Receptor. Mol. Cells 2019, 42, 143–150. [Google Scholar] [CrossRef]
- Qu, S.; Le, W.; Zhang, X.; Xie, W.; Zhang, A.; Ondo, W.G. Locomotion is increased in a11-lesioned mice with iron deprivation: A possible animal model for restless legs syndrome. J. Neuropathol. Exp. Neurol. 2007, 66, 383–388. [Google Scholar] [CrossRef] [Green Version]
- Haranishi, Y.; Hara, K.; Terada, T. Antihyperalgesic effects of intrathecal perospirone in a rat model of neuropathic pain. Pharmacol. Biochem. Behav. 2020, 195, 172964. [Google Scholar] [CrossRef]
- Zarrindast, M.R.; Moghaddampour, E. Opposing influences of D-1 and D-2 dopamine receptors activation on morphine-induced antinociception. Arch. Int. Pharmacodyn. Ther. 1989, 300, 37–50. [Google Scholar] [PubMed]
- Verma, A.; Kulkarni, S.K. Modulatory role of D-1 and D-2 dopamine receptor subtypes in nociception in mice. J. Psychopharmacol. 1993, 7, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Ben-Sreti, M.M.; Gonzalez, J.P.; Sewell, R.D. Differential effects of SKF 38393 and LY 141865 on nociception and morphine analgesia. Life Sci. 1983, 33 (Suppl. S1), 665–668. [Google Scholar] [CrossRef]
- Zarrindast, M.R.; Nassiri-Rad, S.; Pazouki, M. Effects of dopaminergic agents on antinociception in formalin test. Gen. Pharm. 1999, 32, 517–522. [Google Scholar] [CrossRef]
- Yang, P.; Arnold, S.A.; Habas, A.; Hetman, M.; Hagg, T. Ciliary neurotrophic factor mediates dopamine D2 receptor-induced CNS neurogenesis in adult mice. J. Neurosci. 2008, 28, 2231–2241. [Google Scholar] [CrossRef] [Green Version]
- Naftchi, N.E.; Demeny, M.; Decrescito, V.; Tomasula, J.J.; Flamm, E.S.; Campbell, J.B. Biogenic Amine Concecentrations in Traumatized Spinal Cords of Cats—Effect of Drug Therapy. J. Neurosurg. 1974, 40, 52–57. [Google Scholar] [CrossRef]
- Hedeman, L.S.; Sil, R. Studies in experimental spinal cord trauma. 2. Comparison of treatment with steroids, low molecular weight dextran, and catecholamine blockade. J. Neurosurg. 1974, 40, 44–51. [Google Scholar] [CrossRef]
- Chu, T.H.; Cummins, K.; Stys, P.K. The triple monoamine re-uptake inhibitor DOV 216,303 promotes functional recovery after spinal cord contusion injury in mice. Neurosci. Lett. 2018, 675, 1–6. [Google Scholar] [CrossRef]
- Lahlou, S. Involvement of spinal dopamine receptors in mediation of the hypotensive and bradycardic effects of systemic quinpirole in anaesthetised rats. Eur. J. Pharmacol. 1998, 353, 227–237. [Google Scholar] [CrossRef]
- Pellissier, G.; Demenge, P. Hypotensive and bradycardic effects elicited by spinal dopamine receptor stimulation: Effects of D1 and D2 receptor agonists and antagonists. J. Cardiovasc. Pharmacol. 1991, 18, 548–555. [Google Scholar] [CrossRef]
- Petitjean, P.; Mouchet, P.; Pellissier, G.; Manier, M.; Feuerstein, C.; Demenge, P. Cardiovascular effects in the rat of intrathecal injections of apomorphine at the thoracic spinal cord level. Eur. J. Pharmacol. 1984, 105, 355–359. [Google Scholar] [CrossRef]
- Turtle, J.D.; Henwood, M.K.; Strain, M.M.; Huang, Y.J.; Miranda, R.C.; Grau, J.W. Engaging pain fibers after a spinal cord injury fosters hemorrhage and expands the area of secondary injury. Exp. Neurol. 2019, 311, 115–124. [Google Scholar] [CrossRef]
- Mautes, A.E.; Weinzierl, M.R.; Donovan, F.; Noble, L.J. Vascular events after spinal cord injury: Contribution to secondary pathogenesis. Phys. Ther. 2000, 80, 673–687. [Google Scholar] [CrossRef]
- Davis, J.A.; Bopp, A.C.; Henwood, M.K.; Baine, R.E.; Cox, C.C.; Grau, J.W. Pharmacological transection of brain-spinal cord communication blocks pain-induced hemorrhage and locomotor deficits after spinal cord injury in rats. J. Neurotrauma 2020, 37, 1729–1739. [Google Scholar] [CrossRef]
- Reynolds, J.; Turtle, J.; Leal, G.; Huang, Y.J.; Strain, M.; Grau, J. Inflammatory response to peripheral pain input after SCI is blocked by a spinal transection. J. Neurotrauma 2017, 34, A6. [Google Scholar]
- Fauss, G.N.K.; Strain, M.M.; Huang, Y.-J.; Reynolds, J.A.; Davis, J.A.; Henwood, M.K.; West, C.R.; Grau, J.W. Contribution of Brain Processes to Tissue Loss After Spinal Cord Injury: Does a Pain-Induced Rise in Blood Pressure Fuel Hemorrhage? Front. Syst. Neurosci. 2021, 15, 733056. [Google Scholar] [CrossRef] [PubMed]
- Strain, M.M.; Johnston, D.T.; Baine, R.E.; Reynolds, J.; Huang, Y.J.; Henwood, M.K.; Fauss, G.N.; Davis, J.A.; Miranda, R.C.; West, C.R.; et al. Hemorrhage and locomotor deficits induced by pain input after spinal cord injury are partially mediated by changes in hemodynamics. J. Neurotrauma 2021, 38, 3406–3430. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.D.; Everhart, A.W.; Pickelman, J.T.; Hulsebosch, C.E. Mechanical and thermal allodynia in chronic central pain following spinal cord injury. Pain 1996, 68, 97–107. [Google Scholar] [CrossRef]
- Hook, M.A.; Liu, G.T.; Washburn, S.N.; Ferguson, A.R.; Bopp, A.C.; Huie, J.R.; Grau, J.W. The impact of morphine after a spinal cord injury. Behav. Brain Res. 2007, 179, 281–293. [Google Scholar] [CrossRef] [Green Version]
- Hook, M.A.; Moreno, G.; Woller, S.; Puga, D.; Hoy, K.; Balden, R.; Grau, J.W. Intrathecal Morphine Attenuates Recovery of Function after a Spinal Cord Injury. J. Neurotrauma 2009, 26, 741–752. [Google Scholar] [CrossRef]
- Dale, E.A.; Ben Mabrouk, F.; Mitchell, G.S. Unexpected benefits of intermittent hypoxia: Enhanced respiratory and nonrespiratory motor function. Physiology 2014, 29, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Allen, L.L.; Seven, Y.B.; Baker, T.L.; Mitchell, G.S. Cervical spinal contusion alters Na+–K+–2Cl− and K+–Cl− cation-chloride cotransporter expression in phrenic motor neurons. Respir. Physiol. Neurobiol. 2019, 261, 15–23. [Google Scholar] [CrossRef]


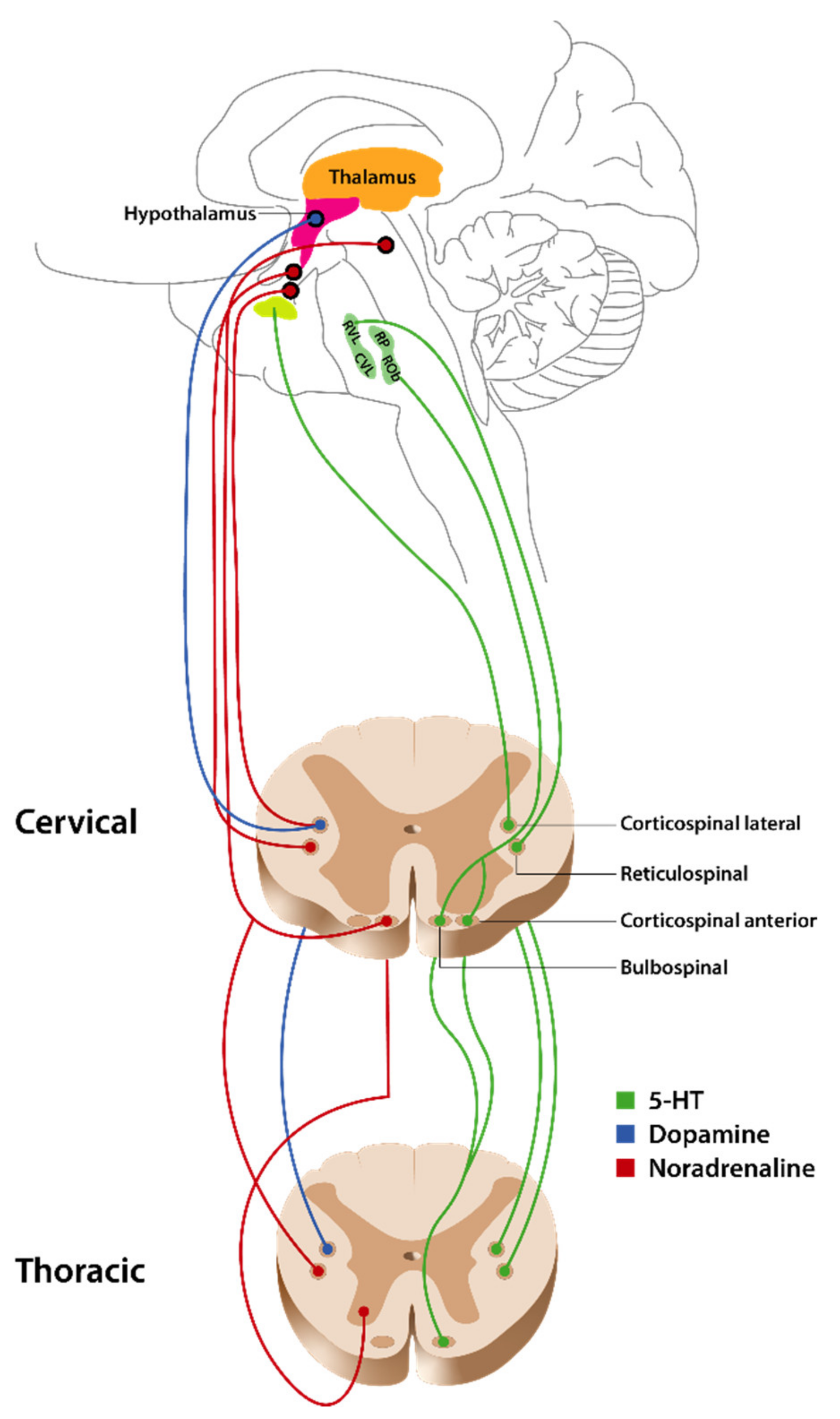{kind=link}
| Receptor | Receptor Type | Location in SC | Normal Function | Function after SCI | |
|---|---|---|---|---|---|
| 5-HT1 | 1A | Gi/o | Primarily in laminae I and II [35]; Cell bodies in dorsal and ventral horns and intermediate zone [36,37,38] | Antinociception [39,40]; Pronociception [41,42]; Enhances motoneurons [43]; Micturition reflex facilitation [44,45,46,47,48,49]; Inhibits motor function [50,51] | Locomotor recovery [52,53,54]; Antinociception [39] |
| 1B | Intermediate zone [35,55]; Dorsal horn (laminae I and IV) [35,55,56] | Antinociception [40,57] | Mitigating spasms [58]; Inhibits mono- and polysynaptic reflexes [58,59] | ||
| 1D | Superficial dorsal horn [60,61]; γ motoneurons in ventral horn [62] | Antinociception [57]; Inhibits monosynaptic reflexes [63] | Inhibits bladder activity [64]; Inhibits mono- and polysynaptic reflexes [65] | ||
| 1E | |||||
| 1F | DRG [66] | Antinociception [67,68] | Mitigating Spasms [58] | ||
| 5-HT2 | 2A | Gαq | Laminae II and II of dorsal horn [69]; Ventral horn [70] | Antinociception [71,72]; Pronociception [40,73,74]; Protects adaptive learning [75]; Sexual behavior [76,77]; Micturition reflex facilitation [78,79]; Motor function [50,51,80,81,82] | Functional motor recovery [83,84]; Respiratory recovery [85]; Bladder recovery [79] |
| 2B | Dorsal horn [86]; DRG [86,87]; Motoneurons [88] | Pronociception [86,87,89] | Functional motor recovery [83]; Mitigates spasms [83,90]; Respiration [88] | ||
| 2C | Most parts of spinal gray (except lamina II) [91] and superficial dorsal horn [92] | Spinal reflexes [93]; Inhibit motor activity [94]; Micturition reflex inhibition [78,79,95,96] | Functional motor recovery [83,97]; Mitigates spasms [83,90] | ||
| 5-HT3 | 3A | Ligand-gated ion channel | In spinal gray matter [91,98]; Laminae VI through X in dorsal horn [91]; DRG [99] | Pronociception [40,100,101]; Antinociception [102,103]; Micturition facilitation [104] | Motor recovery [105] |
| 3B | |||||
| 3C | |||||
| 3D | |||||
| 3E | |||||
| 5-HT4 | GαS | Ventral horn [106] | Pronociception [107]; Micturition reflex facilitation [45] | Locomotor recovery [54,108,109] | |
| 5-HT5 | 5A | Gi/o | Laminae I and II of dorsal horn [110] | Antinociception [111,112]; Micturition function [110] | |
| 5B | Not expressed in humans [113] | ||||
| 5-HT6 | GαS | Superficial dorsal horn and lamina IX [114]; DRG [115] | Pronociception [116] | ||
| 5-HT7 | GαS | Superficial laminae [117]; Laminae VII and VIII [118] | Pronociception [119,120]; Antinociception [120,121]; Micturition reflex facilitation [122,123]; Motor function [51,81,82,124] | ||
| Receptor | Agonists | Antagonists | Non-Selective Agonists | Non-Selective Antagonists | |
|---|---|---|---|---|---|
| 5-HT1 | 1A | 8-OH-DPAT (5-HT1A/7) [125]; Diprpyl-5-CT and Gepirone [126] | WAY-100635 [127]; BMY 7378, NAN-190, MDL 75005 EF; SDZ 216525 [126]; NAD-299 [49] | Propranolol [116]; Spiperone and Pindolol [126] | |
| 1B | TFMPP and mCPP [128]; L-694247, RU 24969 [129], 5-CT, CP 93129 [126] | Quipazine [128], Methiothepin, SB-244289 and SB-216641 [126] | |||
| 1D | Gr-46611 [130] | BRL-15572 [126,130]; Ketanserin and Ritanserin [126] | |||
| 1E | |||||
| 1F | Lasmiditan (COL-144; LY573144) [68]; LY344864 and LY334370 [126] | ||||
| 5-HT2 | 2A | DOI (5-HT2A/2C) [131]; TCB-2 [71]; Quipazine [132] | Ketanserin; Ritanserin (5-HT2A/2c) [133]; MDL 100907, SB 200646A, SB 206553 [126] | DOM [134]; SB 200646 (5-HT2B/2C) and SB 206553 (5-HT2B/2C) [126] | Ketanserin [116]; Methysergide (5-HT1/2) [135] |
| 2B | α-methey-5-HT [136]; SB 204741, Yohimbine [126] | RS-127445 [95]; SB 204741 [126] | |||
| 2C | MK-212 [130]; WAY-161503 [71]; RO-600175 [96] | D-MC-5-H-dibenzo [130]; N-desmethylclozapine [71]; SB-242084 [96,126], RS-102221 [126] | |||
| 5-HT3 | 3A | SR-57227 [137]; 2-methyl-5-HT [134]; PBG [126] | Ondansetron (Zofran3), Alosetron [138], Granisetron, Tropisetron, MDL 72 222 [126] | Tropisetron [116] | |
| 3B | |||||
| 3C | |||||
| 3D | |||||
| 3E | |||||
| 5-HT4 | GR 113808 and SB204070 [126] | ||||
| 5-HT5 | 5A | ||||
| 5B | |||||
| 5-HT6 | EMD-386088 [117] | SB-271046 [139]; SB-399885, SB-258585 [116]; Ro 04-6790 and Ro 63-0563 [126] | |||
| 5-H71 | LP-211 [140]; E-57431, AS-19 [141]; E-55888 [142] | SB-269970 [119,143]; SB-656104 [140]; SB-258719 [141,142]; LP44 [123] | |||
| Receptor | Receptor Type | Location in SC | Normal Function | Function after SCI | |
|---|---|---|---|---|---|
| α1 | α1A | Gq/11 | Dorsal horn, intermediate cell column, and ventral horn [17]; motoneurons [356] | Antinociception [17]; motor behavior, pronociception, autonomic processing [17] | Spontaneous motoneuron activity7; spasticity [357]; Sympathetic neurovascular function [358,359]; micturition [360,361] |
| α1B | |||||
| α1C | |||||
| α2 | α2A | Gi/o | Superficial dorsal horn and deeper laminae, and lamina X [17]; motoneurons [356] | Antinociception [17,362,363]; inhibits sympathetic outflow [17] | Locomotor recovery [364]; mediates bowel dysfunction [365]; reflex/muscle spasticity [366,367]; neurological recovery [368] |
| α2B | Dorsal horn [17] | ||||
| α2C | Dorsal horn and DRG [17]; motoneurons [356] | ||||
| β | β1 | Gs | Cardiac function [369] | Micturition [370], locomotor recovery [371,372], cardiac function [369] | |
| β2 | |||||
| β3 | |||||
| β4 | |||||
| Receptor | Agonists | Antagonists | Non-Selective Agonists | Non-Selective Antagonists | |
|---|---|---|---|---|---|
| α1 | α1A | Methoxamine (A61603) [357] | WB4010 [357], prozosin [357], BRL44408 [373], silodosin, naftopidil [374], tamsulosin [361] | REC15/2739 [357]; methoxamine [358], phenylephrine [358,359,360] | Terazosin [360,375] |
| α1B | |||||
| α1C | |||||
| α2 | α2A | Clonidine, UK14303 [357,376], Guanfacine [377] | Atipamezole [373] | Dexmedetomidine [368,378], guanabenz, UK-14304 [376], tianidine [367]; medetomidine [379] | Yohimbine, RX821001(2) [357], rauwolscine, idazoxan [376], efaroxan [373] |
| α2B | ARC239 [373] | ||||
| α2C | |||||
| β | β1 | Dobutamine [369] | Propranolol [380,381], carvedilol [382,383], nadolol [384] | ||
| β2 | Formoterol [371,372] | ICI118551 [385] | |||
| β3 | Vibegron [370] | SR59230A [385] | |||
| β4 | |||||
| Receptor | Receptor Type | Location in SC | Normal Function | Function after SCI | |
|---|---|---|---|---|---|
| D1-like | D1 | Gq | Throughout the spinal cord [17] | Pronociception [17] | Micturition [398,399], cardiovascular function [400], pronociception [401] |
| D5 | |||||
| D2-like | D2 | Gi/o | Superficial laminae and lamina X [17] | Antinociception, pronociception [17] | Micturition [398,399], cardiovascular function [400], |
| D3 | Dorsal horn [17] | antinociception [401] | |||
| D4 | Dorsal horn (check this one) [17] | ||||
| Receptor | Agonists | Antagonists | Non-Selective Agonists | Non-Selective Antagonists | |
|---|---|---|---|---|---|
| D1-like | D1 | SKF 38393 [398,399,402] [403] | SCH 23390 [398,399,400,402,403], SCH 39166 [401,404] | Aripiprazole [405], apomorphine [300,398,400,406], SKF 83959 [402] | |
| D5 | |||||
| D2 | Quinpirole [399,402,403,407], Ropinirole [408], sumanirole [409], B-HT 920, bromocriptine [410,411], LY 141865 [412] | Remoxipride [398,399], domperidone [400,413], metoclopramide [400], eticlopride [414], L-741,626 [402], (−)-sulpiride [403,410], haloperidol [411] | |||
| D2-like | D3 | Pramipexole [401,404], ropinirole [408] | |||
| D4 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fauss, G.N.K.; Hudson, K.E.; Grau, J.W. Role of Descending Serotonergic Fibers in the Development of Pathophysiology after Spinal Cord Injury (SCI): Contribution to Chronic Pain, Spasticity, and Autonomic Dysreflexia. Biology 2022, 11, 234. https://doi.org/10.3390/biology11020234
Fauss GNK, Hudson KE, Grau JW. Role of Descending Serotonergic Fibers in the Development of Pathophysiology after Spinal Cord Injury (SCI): Contribution to Chronic Pain, Spasticity, and Autonomic Dysreflexia. Biology. 2022; 11(2):234. https://doi.org/10.3390/biology11020234
Chicago/Turabian StyleFauss, Gizelle N. K., Kelsey E. Hudson, and James W. Grau. 2022. "Role of Descending Serotonergic Fibers in the Development of Pathophysiology after Spinal Cord Injury (SCI): Contribution to Chronic Pain, Spasticity, and Autonomic Dysreflexia" Biology 11, no. 2: 234. https://doi.org/10.3390/biology11020234