
Osteopenia in a Mouse Model of Spinal Cord Injury: Effects of Age, Sex and Motor Function

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Subjects
2.2. Spinal Contusion Injury
2.3. Assessment of Motor Recovery
2.4. Tibial MicroCT Analysis
2.5. Statistical Analyses
3. Results
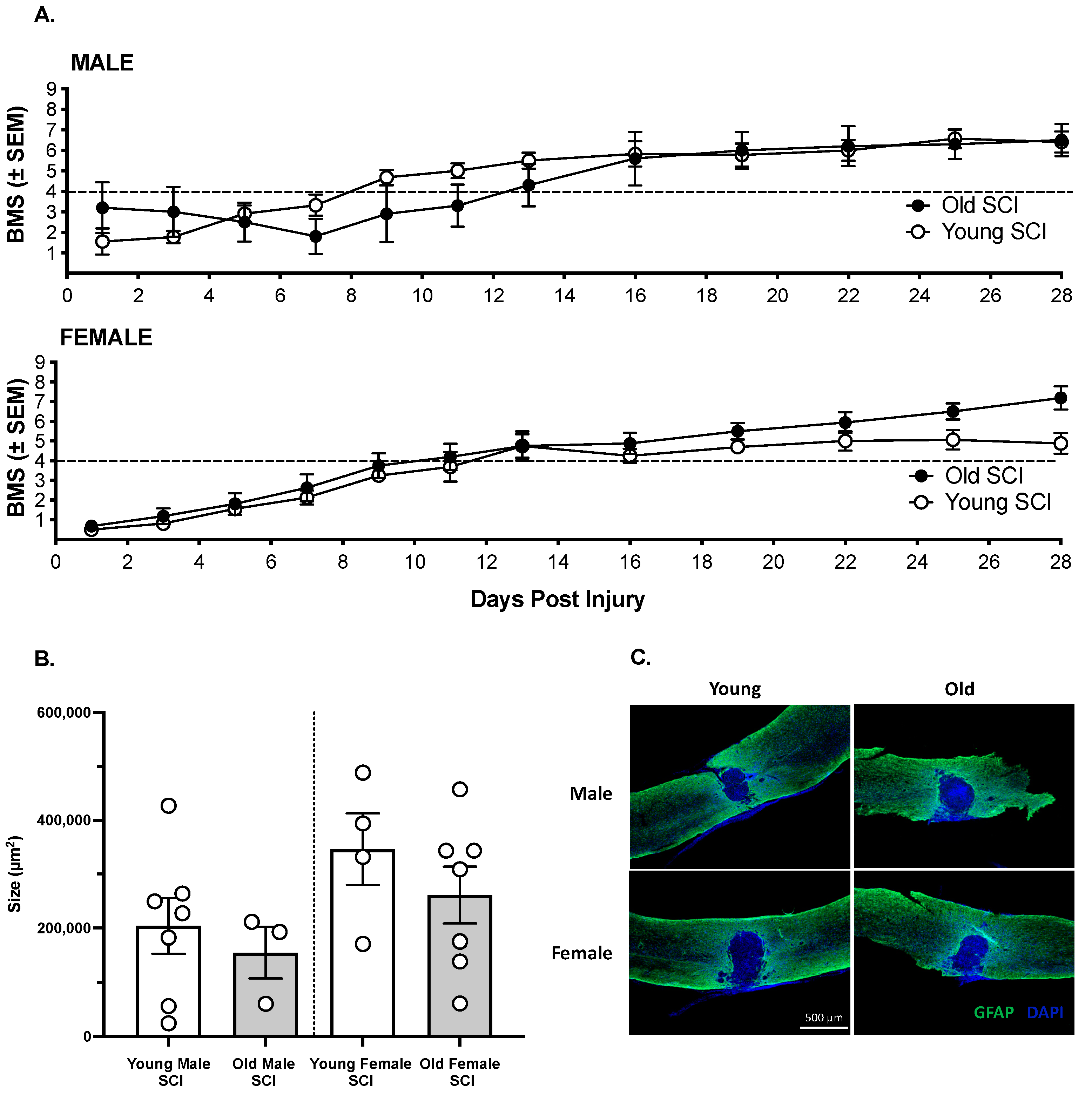
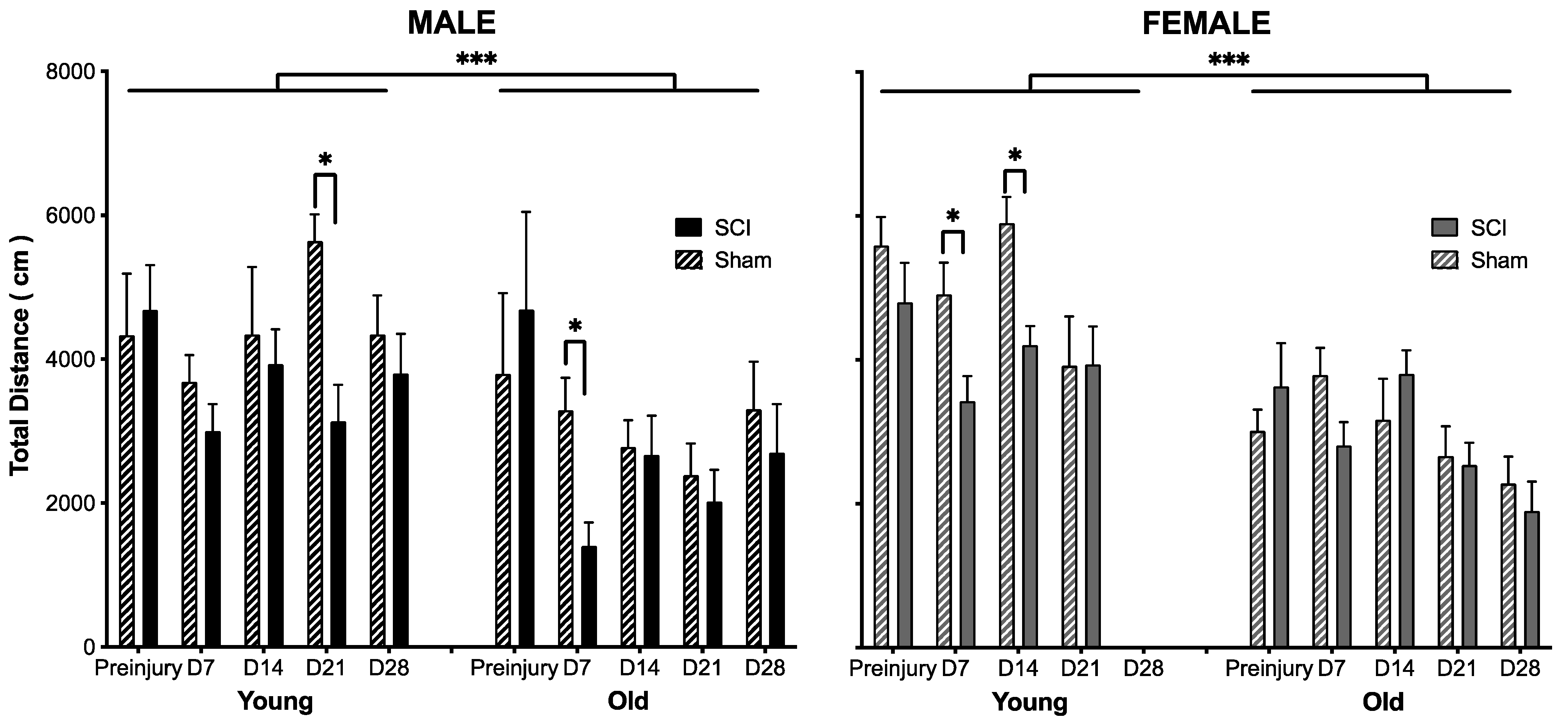
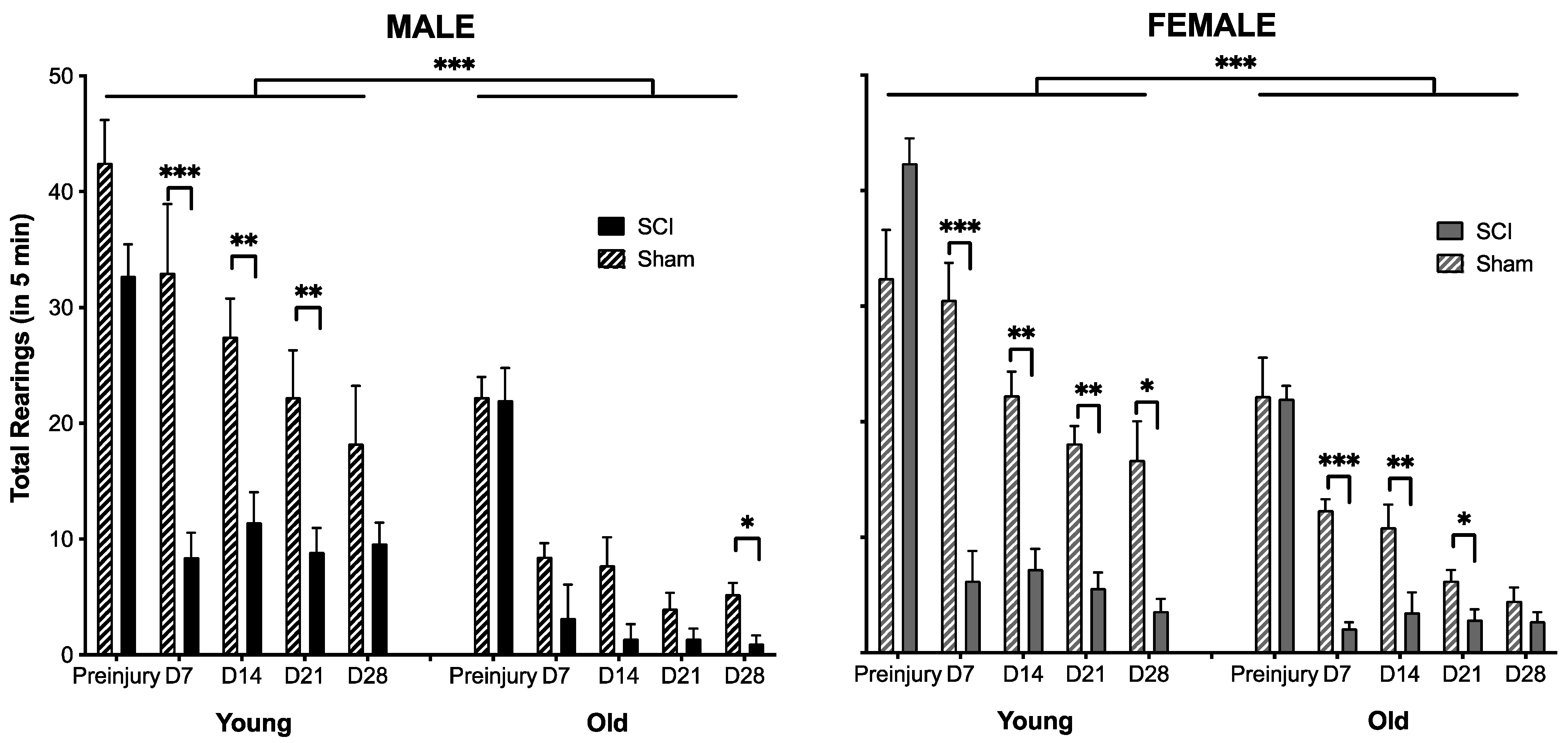
3.1. The Effects of SCI on Motor Recovery Are Task Dependent
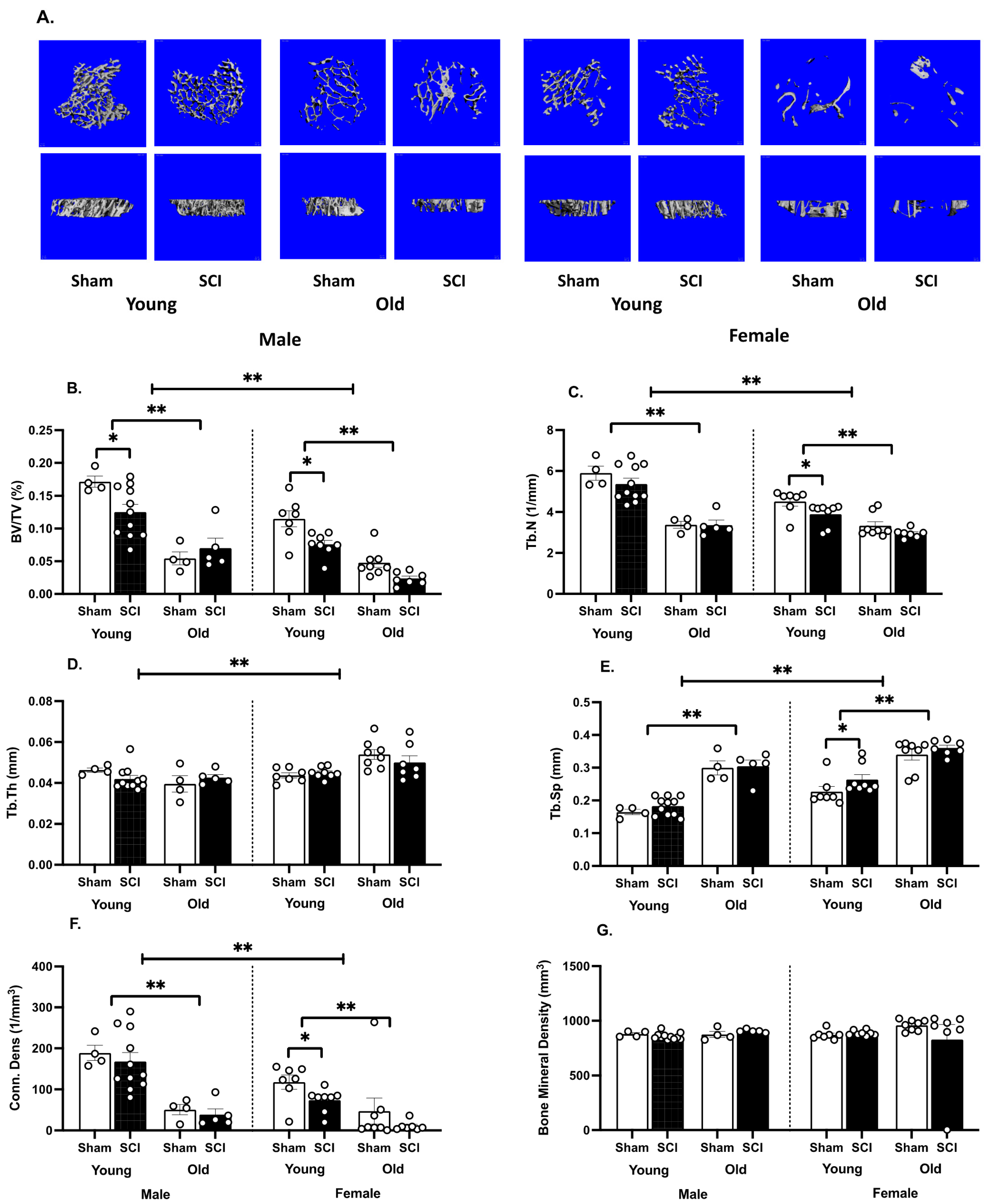
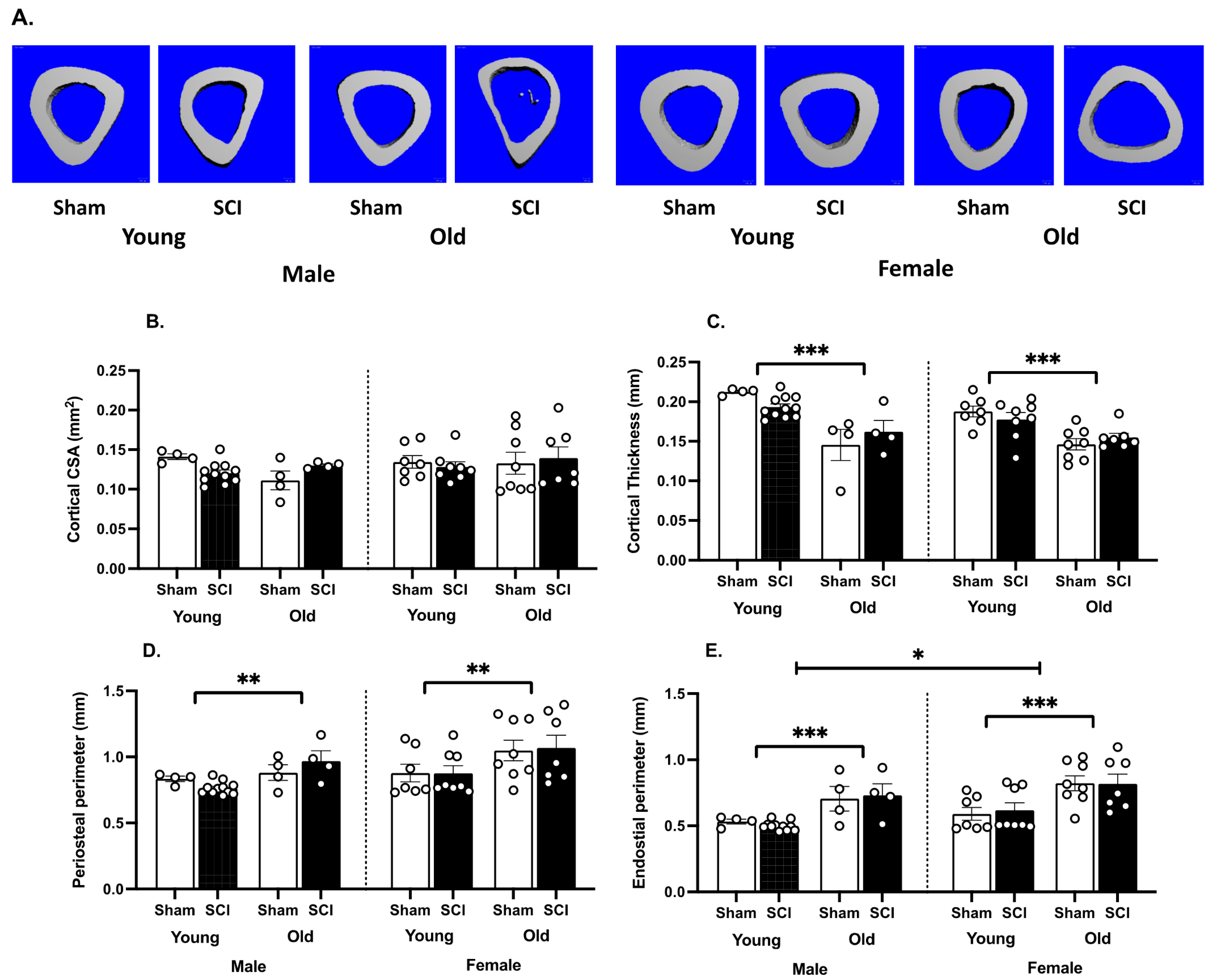
3.2. SCI Significantly Increases Bone Loss
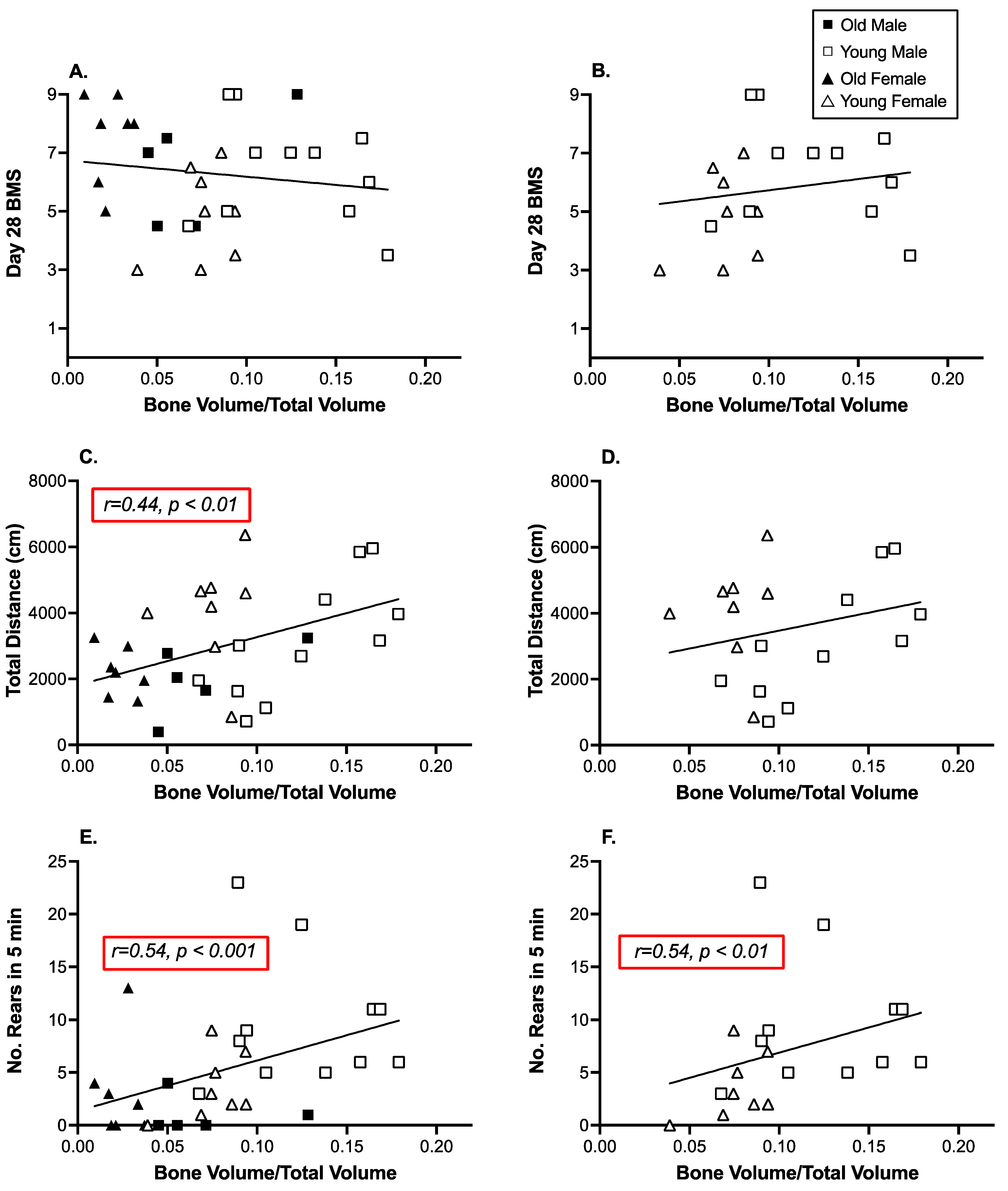
3.3. Open Field Activity and Rearing Were Significantly Correlated with Decreases in Tibial Trabecular Bone Volume
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garland, D.E.; Adkins, R.H.; Stewart, C.A.; Ashford, R.; Vigil, D. Regional osteoporosis in women who have a complete spinal cord injury. J. Bone Jt. Surg Am. 2001, 83, 1195–1200. [Google Scholar] [CrossRef] [PubMed]
- Garland, D.E.; Stewart, C.A.; Adkins, R.H.; Hu, S.S.; Rosen, C.; Liotta, F.J.; Weinstein, D.A. Osteoporosis after spinal cord injury. J. Orthop. Res. 1992, 10, 371–378. [Google Scholar] [CrossRef] [PubMed]
- de Bruin, E.D.; Dietz, V.; Dambacher, M.A.; Stussi, E. Longitudinal changes in bone in men with spinal cord injury. Clin. Rehabil. 2000, 14, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Giangregorio, L.; McCartney, N. Bone loss and muscle atrophy in spinal cord injury: Epidemiology, fracture prediction, and rehabilitation strategies. J. Spinal Cord. Med. 2006, 29, 489–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biering-Sorensen, F.; Bohr, H.H.; Schaadt, O.P. Longitudinal study of bone mineral content in the lumbar spine, the forearm and the lower extremities after spinal cord injury. Eur. J. Clin. Invest. 1990, 20, 330–335. [Google Scholar] [CrossRef]
- Modlesky, C.M.; Majumdar, S.; Narasimhan, A.; Dudley, G.A. Trabecular bone microarchitecture is deteriorated in men with spinal cord injury. J. Bone Miner. Res. 2004, 19, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Shields, R.K.; Dudley-Javoroski, S. Musculoskeletal plasticity after acute spinal cord injury: Effects of long-term neuromuscular electrical stimulation training. J. Neurophysiol. 2006, 95, 2380–2390. [Google Scholar] [CrossRef] [Green Version]
- Modlesky, C.M.; Slade, J.M.; Bickel, C.S.; Meyer, R.A.; Dudley, G.A. Deteriorated geometric structure and strength of the midfemur in men with complete spinal cord injury. Bone 2005, 36, 331–339. [Google Scholar] [CrossRef]
- Eser, P.; Frotzler, A.; Zehnder, Y.; Wick, L.; Knecht, H.; Denoth, J.; Schiessl, H. Relationship between the duration of paralysis and bone structure: A pQCT study of spinal cord injured individuals. Bone 2004, 34, 869–880. [Google Scholar] [CrossRef]
- Coupaud, S.; McLean, A.N.; Purcell, M.; Fraser, M.H.; Allan, D.B. Decreases in bone mineral density at cortical and trabecular sites in the tibia and femur during the first year of spinal cord injury. Bone 2015, 74, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Ragnarsson, K.T.; Sell, G.H. Lower extremity fractures after spinal cord injury: A retrospective study. Arch. Phys. Med. Rehabil. 1981, 62, 418–423. [Google Scholar] [PubMed]
- Vestergaard, P.; Krogh, K.; Rejnmark, L.; Mosekilde, L. Fracture rates and risk factors for fractures in patients with spinal cord injury. Spinal Cord. 1998, 36, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Lazo, M.G.; Shirazi, P.; Sam, M.; Giobbie-Hurder, A.; Blacconiere, M.J.; Muppidi, M. Osteoporosis and risk of fracture in men with spinal cord injury. Spinal Cord. 2001, 39, 208–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, L.D.; Chin, A.S.; Burns, S.P.; Svircev, J.N.; Hoenig, H.; Heggeness, M.; Bailey, L.; Weaver, F. Mortality after lower extremity fractures in men with spinal cord injury. J. Bone Miner. Res. 2014, 29, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Gifre, L.; Vidal, J.; Carrasco, J.; Portell, E.; Puig, J.; Monegal, A.; Guanabens, N.; Peris, P. Incidence of skeletal fractures after traumatic spinal cord injury: A 10-year follow-up study. Clin. Rehabil. 2014, 28, 361–369. [Google Scholar] [CrossRef]
- Edwards, W.B.; Schnitzer, T.J. Bone Imaging and Fracture Risk after Spinal Cord. Injury. Curr. Osteoporos. Rep. 2015, 13, 310–317. [Google Scholar] [CrossRef]
- Keating, J.F.; Kerr, M.; Delargy, M. Minimal trauma causing fractures in patients with spinal cord injury. Disabil. Rehabil. 1992, 14, 108–109. [Google Scholar] [CrossRef]
- Morse, L.R.; Giangregorio, L.; Battaglino, R.A.; Holland, R.; Craven, B.C.; Stolzmann, K.L.; Lazzari, A.A.; Sabharwal, S.; Garshick, E. VA-based survey of osteoporosis management in spinal cord injury. PM R 2009, 1, 240–244. [Google Scholar] [CrossRef] [Green Version]
- Abdelrahman, S.; Ireland, A.; Winter, E.M.; Purcell, M.; Coupaud, S. Osteoporosis after spinal cord injury: Aetiology, effects and therapeutic approaches. J. Musculoskelet. Neuronal. Interact. 2021, 21, 26–50. [Google Scholar]
- Zeman, R.J.; Zhao, J.; Zhang, Y.; Zhao, W.; Wen, X.; Wu, Y.; Pan, J.; Bauman, W.A.; Cardozo, C. Differential skeletal muscle gene expression after upper or lower motor neuron transection. Pflug. Arch. 2009, 458, 525–535. [Google Scholar] [CrossRef]
- Dudley-Javoroski, S.; Shields, R.K. Muscle and bone plasticity after spinal cord injury: Review of adaptations to disuse and to electrical muscle stimulation. J. Rehabil. Res. Dev. 2008, 45, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Bauman, W.A.; Cardozo, C. Bone and muscle loss after spinal cord injury: Organ interactions. Ann. N. Y. Acad. Sci. 2010, 1211, 66–84. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.M.; Findlay, D.M. Musculoskeletal Health in the Context of Spinal Cord. Injury. Curr. Osteoporos. Rep. 2017, 15, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Colaianni, G.; Cuscito, C.; Mongelli, T.; Oranger, A.; Mori, G.; Brunetti, G.; Colucci, S.; Cinti, S.; Grano, M. Irisin enhances osteoblast differentiation in vitro. Int. J. Endocrinol. 2014, 2014, 902186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colaianni, G.; Grano, M. Role of Irisin on the bone-muscle functional unit. Bonekey Rep. 2015, 4, 765. [Google Scholar] [CrossRef] [Green Version]
- Kawao, N.; Moritake, A.; Tatsumi, K.; Kaji, H. Roles of Irisin in the Linkage from Muscle to Bone During Mechanical Unloading in Mice. Calcif. Tissue Int. 2018, 103, 24–34. [Google Scholar] [CrossRef]
- Estell, E.G.; Le, P.T.; Vegting, Y.; Kim, H.; Wrann, C.; Bouxsein, M.L.; Nagano, K.; Baron, R.; Spiegelman, B.M.; Rosen, C.J. Irisin directly stimulates osteoclastogenesis and bone resorption in vitro and in vivo. Elife 2020, 9, e58172. [Google Scholar] [CrossRef]
- Shams, R.; Drasites, K.P.; Zaman, V.; Matzelle, D.; Shields, D.C.; Garner, D.P.; Sole, C.J.; Haque, A.; Banik, N.L. The Pathophysiology of Osteoporosis after Spinal Cord. Injury. Int. J. Mol. Sci. 2021, 22, 3057. [Google Scholar] [CrossRef]
- Huang, T.S.; Wang, Y.H.; Lee, S.H.; Lai, J.S. Impaired hypothalamus-pituitary-adrenal axis in men with spinal Cord. injuries. Am. J. Phys. Med. Rehabil. 1998, 77, 108–112. [Google Scholar] [CrossRef]
- Jiang, S.D.; Jiang, L.S.; Dai, L.Y. Mechanisms of osteoporosis in spinal cord injury. Clin. Endocrinol. 2006, 65, 555–565. [Google Scholar] [CrossRef]
- del Rivero, T.; Bethea, J.R. The effects of spinal cord injury on bone loss and dysregulation of the calcium/parathyroid hormone loop in mice. Osteoporos. Sarcopenia 2016, 2, 164–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minaire, P.; Edouard, C.; Arlot, M.; Meunier, P.J. Marrow changes in paraplegic patients. Calcif. Tissue Int. 1984, 36, 338–340. [Google Scholar] [CrossRef] [PubMed]
- Elder, C.P.; Apple, D.F.; Bickel, C.S.; Meyer, R.A.; Dudley, G.A. Intramuscular fat and glucose tolerance after spinal cord injury—A cross-sectional study. Spinal Cord. 2004, 42, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Gorgey, A.S.; Dudley, G.A. Skeletal muscle atrophy and increased intramuscular fat after incomplete spinal cord injury. Spinal Cord. 2007, 45, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Li, B.; Chen, J.W.; Jiang, S.D.; Jiang, L.S. Spinal cord injury causes bone loss through peroxisome proliferator-activated receptor-gamma and Wnt signalling. J. Cell Mol. Med. 2012, 16, 2968–2977. [Google Scholar] [CrossRef] [PubMed]
- Metzger, C.E.; Gong, S.; Aceves, M.; Bloomfield, S.A.; Hook, M.A. Osteocytes reflect a pro-inflammatory state following spinal cord injury in a rodent model. Bone 2019, 120, 465–475. [Google Scholar] [CrossRef]
- Bauman, W.A.; Cardozo, C.P. Osteoporosis in individuals with spinal cord injury. PM R 2015, 7, 188–201, quiz 201. [Google Scholar] [CrossRef]
- Maimoun, L.; Ben Bouallegue, F.; Gelis, A.; Aouinti, S.; Mura, T.; Philibert, P.; Souberbielle, J.C.; Piketty, M.; Garnero, P.; Mariano-Goulart, D.; et al. Periostin and sclerostin levels in individuals with spinal cord injury and their relationship with bone mass, bone turnover, fracture and osteoporosis status. Bone 2019, 127, 612–619. [Google Scholar] [CrossRef]
- Zleik, N.; Weaver, F.; Harmon, R.L.; Le, B.; Radhakrishnan, R.; Jirau-Rosaly, W.D.; Craven, B.C.; Raiford, M.; Hill, J.N.; Etingen, B.; et al. Prevention and management of osteoporosis and osteoporotic fractures in persons with a spinal cord injury or disorder: A systematic scoping review. J. Spinal Cord. Med. 2019, 42, 735–759. [Google Scholar] [CrossRef]
- Exton-Smith, A.N.; Millard, P.H.; Payne, P.R.; Wheeler, E.F. Pattern of development and loss of bone with age. Lancet 1969, 2, 1154–1157. [Google Scholar] [CrossRef]
- Firooznia, H.; Golimbu, C.; Rafii, M.; Schwartz, M.S.; Alterman, E.R. Quantitative computed tomography assessment of spinal trabecular bone. I. Age-related regression in normal men and women. J. Comput. Tomogr. 1984, 8, 91–97. [Google Scholar] [CrossRef]
- Wright, N.C.; Looker, A.C.; Saag, K.G.; Curtis, J.R.; Delzell, E.S.; Randall, S.; Dawson-Hughes, B. The recent prevalence of osteoporosis and low bone mass in the United States based on bone mineral density at the femoral neck or lumbar spine. J. Bone Miner. Res. 2014, 29, 2520–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Berg, M.E.; Castellote, J.M.; Mahillo-Fernandez, I.; de Pedro-Cuesta, J. Incidence of spinal cord injury worldwide: A systematic review. Neuroepidemiology 2010, 34, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Knutsdottir, S.; Thorisdottir, H.; Sigvaldason, K.; Jonsson, H., Jr.; Bjornsson, A.; Ingvarsson, P. Epidemiology of traumatic spinal Cord. injuries in Iceland from 1975 to 2009. Spinal Cord. 2012, 50, 123–126. [Google Scholar] [CrossRef] [Green Version]
- DeVivo, M.J.; Chen, Y. Trends in new injuries, prevalent cases, and aging with spinal cord injury. Arch. Phys. Med. Rehabil. 2011, 92, 332–338. [Google Scholar] [CrossRef]
- Nijendijk, J.H.; Post, M.W.; van Asbeck, F.W. Epidemiology of traumatic spinal Cord. injuries in The Netherlands in 2010. Spinal Cord. 2014, 52, 258–263. [Google Scholar] [CrossRef]
- McCaughey, E.J.; Purcell, M.; McLean, A.N.; Fraser, M.H.; Bewick, A.; Borotkanics, R.J.; Allan, D.B. Changing demographics of spinal cord injury over a 20-year period: A longitudinal population-based study in Scotland. Spinal Cord. 2016, 54, 270–276. [Google Scholar] [CrossRef] [Green Version]
- Barbara-Bataller, E.; Mendez-Suarez, J.L.; Aleman-Sanchez, C.; Sanchez-Enriquez, J.; Sosa-Henriquez, M. Change in the profile of traumatic spinal cord injury over 15 years in Spain. Scand. J. Trauma Resusc. Emerg. Med. 2018, 26, 27. [Google Scholar] [CrossRef] [Green Version]
- Hernlund, E.; Svedbom, A.; Ivergard, M.; Compston, J.; Cooper, C.; Stenmark, J.; McCloskey, E.V.; Jonsson, B.; Kanis, J.A. Osteoporosis in the European Union: Medical management, epidemiology and economic burden. A report prepared in collaboration with the International Osteoporosis Foundation (IOF) and the European Federation of Pharmaceutical Industry Associations (EFPIA). Arch. Osteoporos. 2013, 8, 136. [Google Scholar] [CrossRef] [Green Version]
- Alswat, K.A. Gender Disparities in Osteoporosis. J. Clin. Med. Res. 2017, 9, 382–387. [Google Scholar] [CrossRef] [Green Version]
- Krolner, B.; Tondevold, E.; Toft, B.; Berthelsen, B.; Nielsen, S.P. Bone mass of the axial and the appendicular skeleton in women with Colles’ fracture: Its relation to physical activity. Clin. Physiol. 1982, 2, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Nilas, L.; Christiansen, C. Rates of bone loss in normal women: Evidence of accelerated trabecular bone loss after the menopause. Eur. J. Clin. Invest. 1988, 18, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Stepan, J.J.; Tesarova, A.; Havranek, T.; Jodl, J.; Formankova, J.; Pacovsky, V. Age and sex dependency of the biochemical indices of bone remodelling. Clin. Chim Acta 1985, 151, 273–283. [Google Scholar] [CrossRef]
- Dannels, A.; Charlifue, S. The perimenopause experience for women with spinal Cord. injuries. SCI. Nurs. 2004, 21, 9–13. [Google Scholar]
- Hubscher, C.H.; Armstrong, J.E.; Johnson, J.R. Effects of spinal cord injury on the rat estrous cycle. Brain Res. 2006, 1100, 118–124. [Google Scholar] [CrossRef]
- Rutberg, L.; Friden, B.; Karlsson, A.K. Amenorrhoea in newly spinal Cord. injured women: An effect of hyperprolactinaemia? Spinal Cord. 2008, 46, 189–191. [Google Scholar] [CrossRef] [Green Version]
- Furlan, J.C.; Krassioukov, A.V.; Fehlings, M.G. The effects of gender on clinical and neurological outcomes after acute cervical spinal cord injury. J. Neurotrauma 2005, 22, 368–381. [Google Scholar] [CrossRef]
- Ng, J.S.; Chin, K.Y. Potential mechanisms linking psychological stress to bone health. Int. J. Med. Sci. 2021, 18, 604–614. [Google Scholar] [CrossRef]
- Baker-LePain, J.C.; Nakamura, M.C.; Lane, N.E. Effects of inflammation on bone: An update. Curr. Opin. Rheumatol. 2011, 23, 389–395. [Google Scholar] [CrossRef]
- Basso, D.M.; Fisher, L.C.; Anderson, A.J.; Jakeman, L.B.; McTigue, D.M.; Popovich, P.G. Basso Mouse Scale for locomotion detects differences in recovery after spinal cord injury in five common mouse strains. J. Neurotrauma 2006, 23, 635–659. [Google Scholar] [CrossRef]
- Perrien, D.S.; Akel, N.S.; Edwards, P.K.; Carver, A.A.; Bendre, M.S.; Swain, F.L.; Skinner, R.A.; Hogue, W.R.; Nicks, K.M.; Pierson, T.M.; et al. Inhibin A is an endocrine stimulator of bone mass and strength. Endocrinology 2007, 148, 1654–1665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouxsein, M.L.; Boyd, S.K.; Christiansen, B.A.; Guldberg, R.E.; Jepsen, K.J.; Muller, R. Guidelines for assessment of bone microstructure in rodents using micro-computed tomography. J. Bone Miner. Res. 2010, 25, 1468–1486. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Bouchard, S.; Peters, K.; Woller, S.A.; Madahian, B.; Faghihi, U.; Patel, S.; Bake, S.; Hook, M.A. Inflammation is increased with anxiety- and depression-like signs in a rat model of spinal cord injury. Brain Behav. Immun. 2016, 51, 176–195. [Google Scholar] [CrossRef] [Green Version]
- Luedtke, K.; Bouchard, S.M.; Woller, S.A.; Funk, M.K.; Aceves, M.; Hook, M.A. Assessment of depression in a rodent model of spinal cord injury. J. Neurotrauma 2014, 31, 1107–1121. [Google Scholar] [CrossRef] [Green Version]
- Brakel, K.; Aceves, A.R.; Aceves, M.; Hierholzer, A.; Nguyen, Q.N.; Hook, M.A. Depression-like behavior corresponds with cardiac changes in a rodent model of spinal cord injury. Exp. Neurol. 2019, 320, 112969. [Google Scholar] [CrossRef] [PubMed]
- Brakel, K.; Aceves, M.; Garza, A.; Yoo, C.; Escobedo, G.; Jr Panchani, N.; Shapiro, L.; Hook, M. Inflammation increases the development of depression behaviors in male rats after spinal cord injury. Brain Behav Immun Health 2021, 14, 100258. [Google Scholar] [CrossRef]
- Wu, J.; Zhao, Z.; Sabirzhanov, B.; Stoica, B.A.; Kumar, A.; Luo, T.; Skovira, J.; Faden, A.I. Spinal cord injury causes brain inflammation associated with cognitive and affective changes: Role of cell cycle pathways. J. Neurosci. Off. J. Soc. Neurosci. 2014, 34, 10989–11006. [Google Scholar] [CrossRef]
- Williams, R.; Murray, A. Prevalence of depression after spinal cord injury: A meta-analysis. Arch. Phys. Med. Rehabil. 2015, 96, 133–140. [Google Scholar] [CrossRef]
- Farrell, K.; Houle, J.D. Systemic Inhibition of Soluble Tumor Necrosis Factor with XPro1595 Exacerbates a Post-Spinal Cord. Injury Depressive Phenotype in Female Rats. J. Neurotrauma 2019, 36, 2964–2976. [Google Scholar] [CrossRef]
- do Espirito Santo, C.C.; da Silva Fiorin, F.; Ilha, J.; Duarte, M.; Duarte, T.; Santos, A.R.S. Spinal cord injury by clip-compression induces anxiety and depression-like behaviours in female rats: The role of the inflammatory response. Brain Behav. Immun. 2019, 78, 91–104. [Google Scholar] [CrossRef]
- Elliott, T.R.; Frank, R.G. Depression following spinal cord injury. Arch. Phys. Med. Rehabil. 1996, 77, 816–823. [Google Scholar] [CrossRef]
- Dolbow, D.R.; Gorgey, A.S.; Cifu, D.X.; Moore, J.R.; Gater, D.R. Feasibility of home-based functional electrical stimulation cycling: Case report. Spinal Cord. 2012, 50, 170–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolbow, D.R.; Gorgey, A.S.; Ketchum, J.M.; Moore, J.R.; Hackett, L.A.; Gater, D.R. Exercise adherence during home-based functional electrical stimulation cycling by individuals with spinal cord injury. Am. J. Phys. Med. Rehabil. 2012, 91, 922–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morse, L.R.; Nguyen, N.; Battaglino, R.A.; Guarino, A.J.; Gagnon, D.R.; Zafonte, R.; Garshick, E. Wheelchair use and lipophilic statin medications may influence bone loss in chronic spinal cord injury: Findings from the FRASCI-bone loss study. Osteoporos. Int. 2016, 27, 3503–3511. [Google Scholar] [CrossRef] [Green Version]
- Turner, C.H.; Takano, Y.; Owan, I. Aging changes mechanical loading thresholds for bone formation in rats. J. Bone Miner. Res. 1995, 10, 1544–1549. [Google Scholar] [CrossRef]
- Hughes, J.M.; Charkoudian, N.; Barnes, J.N.; Morgan, B.J. Revisiting the Debate: Does Exercise Build Strong Bones in the Mature and Senescent Skeleton? Front. Physiol. 2016, 7, 369. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, T.; Hughes, F.J.; McCulloch, C.A.; Melcher, A.H. Effects of donor age on osteogenic cells of rat bone marrow in vitro. Mech. Ageing Dev. 1990, 51, 121–132. [Google Scholar] [CrossRef]
- Quarto, R.; Thomas, D.; Liang, C.T. Bone progenitor cell deficits and the age-associated decline in bone repair capacity. Calcif. Tissue Int. 1995, 56, 123–129. [Google Scholar] [CrossRef]
- Majors, A.K.; Boehm, C.A.; Nitto, H.; Midura, R.J.; Muschler, G.F. Characterization of human bone marrow stromal cells with respect to osteoblastic differentiation. J. Orthop. Res. 1997, 15, 546–557. [Google Scholar] [CrossRef]
- Nishida, S.; Endo, N.; Yamagiwa, H.; Tanizawa, T.; Takahashi, H.E. Number of osteoprogenitor cells in human bone marrow markedly decreases after skeletal maturation. J. Bone Miner. Metab 1999, 17, 171–177. [Google Scholar] [CrossRef]
- Muschler, G.F.; Nitto, H.; Boehm, C.A.; Easley, K.A. Age- and gender-related changes in the cellularity of human bone marrow and the prevalence of osteoblastic progenitors. J. Orthop. Res. 2001, 19, 117–125. [Google Scholar] [CrossRef]
- Yeh, L.C.; Wilkerson, M.; Lee, J.C.; Adamo, M.L. IGF-1 Receptor Insufficiency Leads to Age-Dependent Attenuation of Osteoblast Differentiation. Endocrinology 2015, 156, 2872–2879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, K.N.Z.; Thyfault, J.P. Barriers in translating preclinical rodent exercise metabolism findings to human health. J. Appl. Physiol. 2021, 130, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Finch, C.E.; Felicio, L.S.; Mobbs, C.V.; Nelson, J.F. Ovarian and steroidal influences on neuroendocrine aging processes in female rodents. Endocr. Rev. 1984, 5, 467–497. [Google Scholar] [CrossRef] [Green Version]
- Almeida, M.; Han, L.; Martin-Millan, M.; Plotkin, L.I.; Stewart, S.A.; Roberson, P.K.; Kousteni, S.; O’Brien, C.A.; Bellido, T.; Parfitt, A.M.; et al. Skeletal involution by age-associated oxidative stress and its acceleration by loss of sex steroids. J. Biol. Chem. 2007, 282, 27285–27297. [Google Scholar] [CrossRef] [Green Version]
- Gelderd, J.B.; Peppler, R.D. Effect of spinal Cord. transection on the reproductive system in the female rat. Neuroendocrinology 1979, 29, 293–299. [Google Scholar] [CrossRef]
- Shah, P.K.; Song, J.; Kim, S.; Zhong, H.; Roy, R.R.; Edgerton, V.R. Rodent estrous cycle response to incomplete spinal cord injury, surgical interventions, and locomotor training. Behav. NeuroSci. 2011, 125, 996–1002. [Google Scholar] [CrossRef] [Green Version]
- Yarrow, J.F.; Phillips, E.G.; Conover, C.F.; Bassett, T.E.; Chen, C.; Teurlings, T.; Vasconez, A.; Alerte, J.; Prock, H.; Jiron, J.M.; et al. Testosterone Plus Finasteride Prevents Bone Loss without Prostate Growth in a Rodent Spinal Cord. Injury Model. J. Neurotrauma 2017, 34, 2972–2981. [Google Scholar] [CrossRef]
- Yarrow, J.F.; Ye, F.; Balaez, A.; Mantione, J.M.; Otzel, D.M.; Chen, C.; Beggs, L.A.; Baligand, C.; Keener, J.E.; Lim, W.; et al. Bone loss in a new rodent model combining spinal cord injury and cast immobilization. J. Musculoskelet. Neuronal. Interact. 2014, 14, 255–266. [Google Scholar]
- Stewart, A.N.; MacLean, S.M.; Stromberg, A.J.; Whelan, J.P.; Bailey, W.M.; Gensel, J.C.; Wilson, M.E. Considerations for Studying Sex as a Biological Variable in Spinal Cord. Injury. Front. Neurol. 2020, 11, 802. [Google Scholar] [CrossRef]
- Boot, A.M.; de Ridder, M.A.; van der Sluis, I.M.; van Slobbe, I.; Krenning, E.P.; Keizer-Schrama, S.M. Peak bone mineral density, lean body mass and fractures. Bone 2010, 46, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Laurent, M.R.; Dubois, V.; Claessens, F.; O’Brien, C.A.; Bouillon, R.; Vanderschueren, D.; Manolagas, S.C. Estrogens and Androgens in Skeletal Physiology and Pathophysiology. Physiol. Rev. 2017, 97, 135–187. [Google Scholar] [CrossRef] [PubMed]
- Burghardt, A.J.; Kazakia, G.J.; Ramachandran, S.; Link, T.M.; Majumdar, S. Age- and gender-related differences in the geometric properties and biomechanical significance of intracortical porosity in the distal radius and tibia. J. Bone Miner. Res. 2010, 25, 983–993. [Google Scholar] [PubMed]
- Dalzell, N.; Kaptoge, S.; Morris, N.; Berthier, A.; Koller, B.; Braak, L.; van Rietbergen, B.; Reeve, J. Bone micro-architecture and determinants of strength in the radius and tibia: Age-related changes in a population-based study of normal adults measured with high-resolution pQCT. Osteoporos. Int. 2009, 20, 1683–1694. [Google Scholar] [CrossRef]
- Khosla, S.; Riggs, B.L.; Atkinson, E.J.; Oberg, A.L.; McDaniel, L.J.; Holets, M.; Peterson, J.M.; Melton, L.J., 3rd. Effects of sex and age on bone microstructure at the ultradistal radius: A population-based noninvasive in vivo assessment. J. Bone Miner. Res. 2006, 21, 124–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macdonald, H.M.; Nishiyama, K.K.; Kang, J.; Hanley, D.A.; Boyd, S.K. Age-related patterns of trabecular and cortical bone loss differ between sexes and skeletal sites: A population-based HR-pQCT study. J. Bone Miner. Res. 2011, 26, 50–62. [Google Scholar] [CrossRef]
- Nieves, J.W.; Formica, C.; Ruffing, J.; Zion, M.; Garrett, P.; Lindsay, R.; Cosman, F. Males have larger skeletal size and bone mass than females, despite comparable body size. J. Bone Miner. Res. 2005, 20, 529–535. [Google Scholar] [CrossRef]
- Qin, W.; Sun, L.; Cao, J.; Peng, Y.; Collier, L.; Wu, Y.; Creasey, G.; Li, J.; Qin, Y.; Jarvis, J.; et al. The central nervous system (CNS)-independent anti-bone-resorptive activity of muscle contraction and the underlying molecular and cellular signatures. J. Biol. Chem. 2013, 288, 13511–13521. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Peng, Y.; Hu, Y.; Guo, X.E.; Li, J.; Cao, J.; Pan, J.; Feng, J.Q.; Cardozo, C.; Jarvis, J.; et al. Electrical stimulation of hindlimb skeletal muscle has beneficial effects on sublesional bone in a rat model of spinal cord injury. Bone 2021, 144, 115825. [Google Scholar] [CrossRef]
- Baunsgaard, C.B.; Nissen, U.V.; Brust, A.K.; Frotzler, A.; Ribeill, C.; Kalke, Y.B.; Leon, N.; Gomez, B.; Samuelsson, K.; Antepohl, W.; et al. Exoskeleton gait training after spinal cord injury: An exploratory study on secondary health conditions. J. Rehabil. Med. 2018, 50, 806–813. [Google Scholar] [CrossRef] [Green Version]
- Juszczak, M.; Gallo, E.; Bushnik, T. Examining the Effects of a Powered Exoskeleton on Quality of Life and Secondary Impairments in People Living With Spinal Cord Injury. Top. Spinal Cord. Inj. Rehabil. 2018, 24, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Kim, H.S.; Jang, S.H.; Hyun, D.J.; Park, S.I.; Yoon, J.; Lim, H.; Kim, M.J. Cardiorespiratory Responses to 10 Weeks of Exoskeleton-Assisted Overground Walking Training in Chronic Nonambulatory Patients with Spinal Cord. Injury. Sensors 2021, 21, 5022. [Google Scholar] [CrossRef] [PubMed]
- Asselin, P.; Cirnigliaro, C.M.; Kornfeld, S.; Knezevic, S.; Lackow, R.; Elliott, M.; Bauman, W.A.; Spungen, A.M. Effect of Exoskeletal-Assisted Walking on Soft Tissue Body Composition in Persons With Spinal Cord. Injury. Arch. Phys. Med. Rehabil. 2021, 102, 196–202. [Google Scholar] [CrossRef]
- Xiang, X.N.; Zong, H.Y.; Ou, Y.; Yu, X.; Cheng, H.; Du, C.P.; He, H.C. Exoskeleton-assisted walking improves pulmonary function and walking parameters among individuals with spinal cord injury: A randomized controlled pilot study. J. Neuroeng. Rehabil. 2021, 18, 86. [Google Scholar] [CrossRef] [PubMed]
- Needham-Shropshire, B.M.; Broton, J.G.; Klose, K.J.; Lebwohl, N.; Guest, R.S.; Jacobs, P.L. Evaluation of a training program for persons with SCI. paraplegia using the Parastep 1 ambulation system: Part 3. Lack of effect on bone mineral density. Arch. Phys. Med. Rehabil. 1997, 78, 799–803. [Google Scholar] [CrossRef]
- Thoumie, P.; Le Claire, G.; Beillot, J.; Dassonville, J.; Chevalier, T.; Perrouin-Verbe, B.; Bedoiseau, M.; Busnel, M.; Cormerais, A.; Courtillon, A.; et al. Restoration of functional gait in paraplegic patients with the RGO-II hybrid orthosis. A multicenter controlled study. II: Physiological evaluation. Paraplegia 1995, 33, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karelis, A.D.; Carvalho, L.P.; Castillo, M.J.; Gagnon, D.H.; Aubertin-Leheudre, M. Effect on body composition and bone mineral density of walking with a robotic exoskeleton in adults with chronic spinal cord injury. J. Rehabil. Med. 2017, 49, 84–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheffler, L.R.; Chae, J. Neuromuscular electrical stimulation in neurorehabilitation. Muscle Nerve 2007, 35, 562–590. [Google Scholar] [CrossRef]
- Feskanich, D.; Willett, W.; Colditz, G. Walking and leisure-time activity and risk of hip fracture in postmenopausal women. JAMA 2002, 288, 2300–2306. [Google Scholar] [CrossRef] [Green Version]
- Frost, H.M. Bone “mass” and the “mechanostat”: A proposal. Anat. Rec. 1987, 219, 1–9. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hook, M.A.; Falck, A.; Dundumulla, R.; Terminel, M.; Cunningham, R.; Sefiani, A.; Callaway, K.; Gaddy, D.; Geoffroy, C.G. Osteopenia in a Mouse Model of Spinal Cord Injury: Effects of Age, Sex and Motor Function. Biology 2022, 11, 189. https://doi.org/10.3390/biology11020189
Hook MA, Falck A, Dundumulla R, Terminel M, Cunningham R, Sefiani A, Callaway K, Gaddy D, Geoffroy CG. Osteopenia in a Mouse Model of Spinal Cord Injury: Effects of Age, Sex and Motor Function. Biology. 2022; 11(2):189. https://doi.org/10.3390/biology11020189
Chicago/Turabian StyleHook, Michelle A., Alyssa Falck, Ravali Dundumulla, Mabel Terminel, Rachel Cunningham, Arthur Sefiani, Kayla Callaway, Dana Gaddy, and Cédric G. Geoffroy. 2022. "Osteopenia in a Mouse Model of Spinal Cord Injury: Effects of Age, Sex and Motor Function" Biology 11, no. 2: 189. https://doi.org/10.3390/biology11020189