
Oral Processing of Three Guenon Species in Taï National Park, Côte d’Ivoire
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Species
2.2. Oral-Processing Data Collection
2.3. Analytical Methods
2.4. Data Availability and Ethical Statement
3. Results
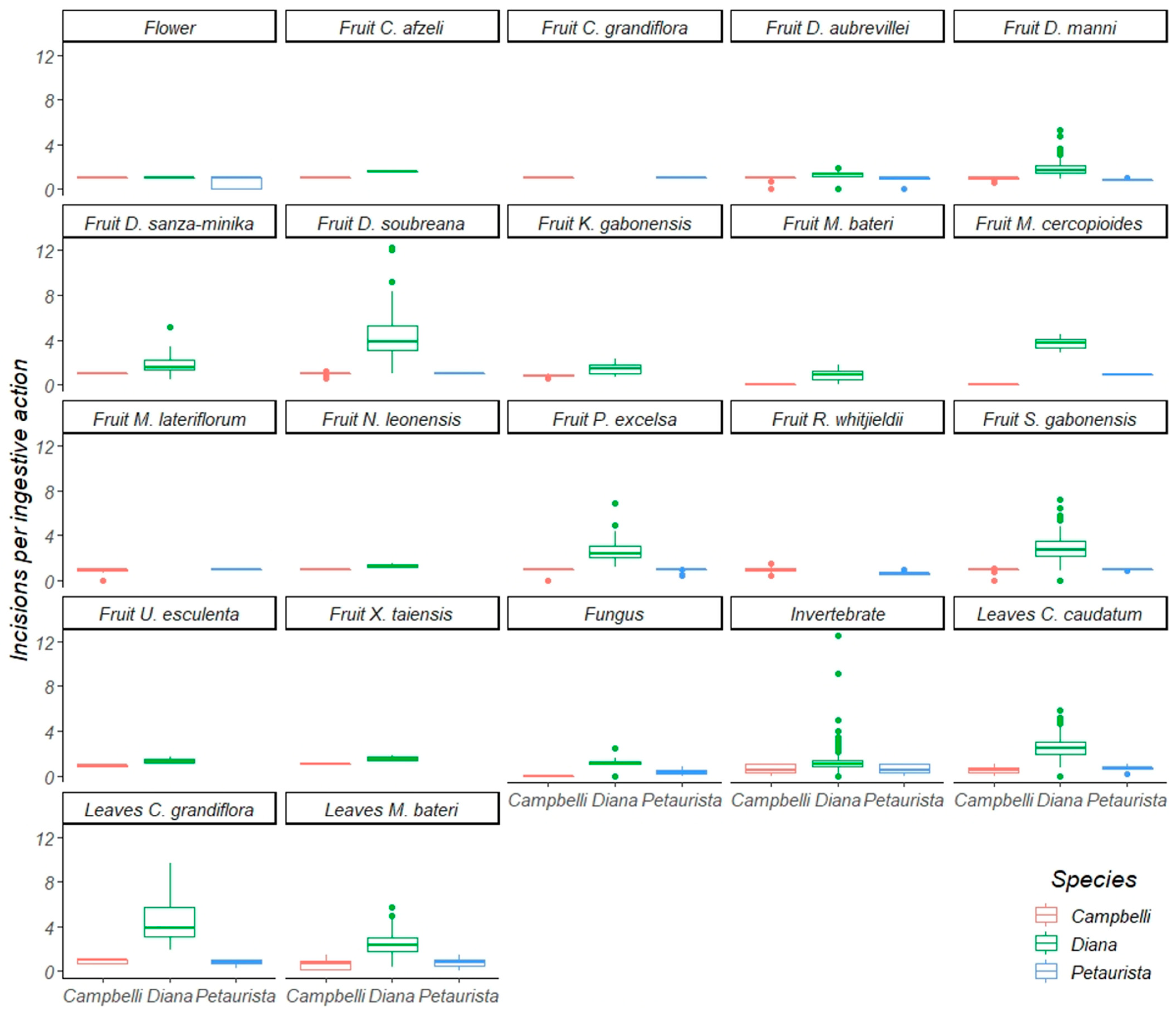
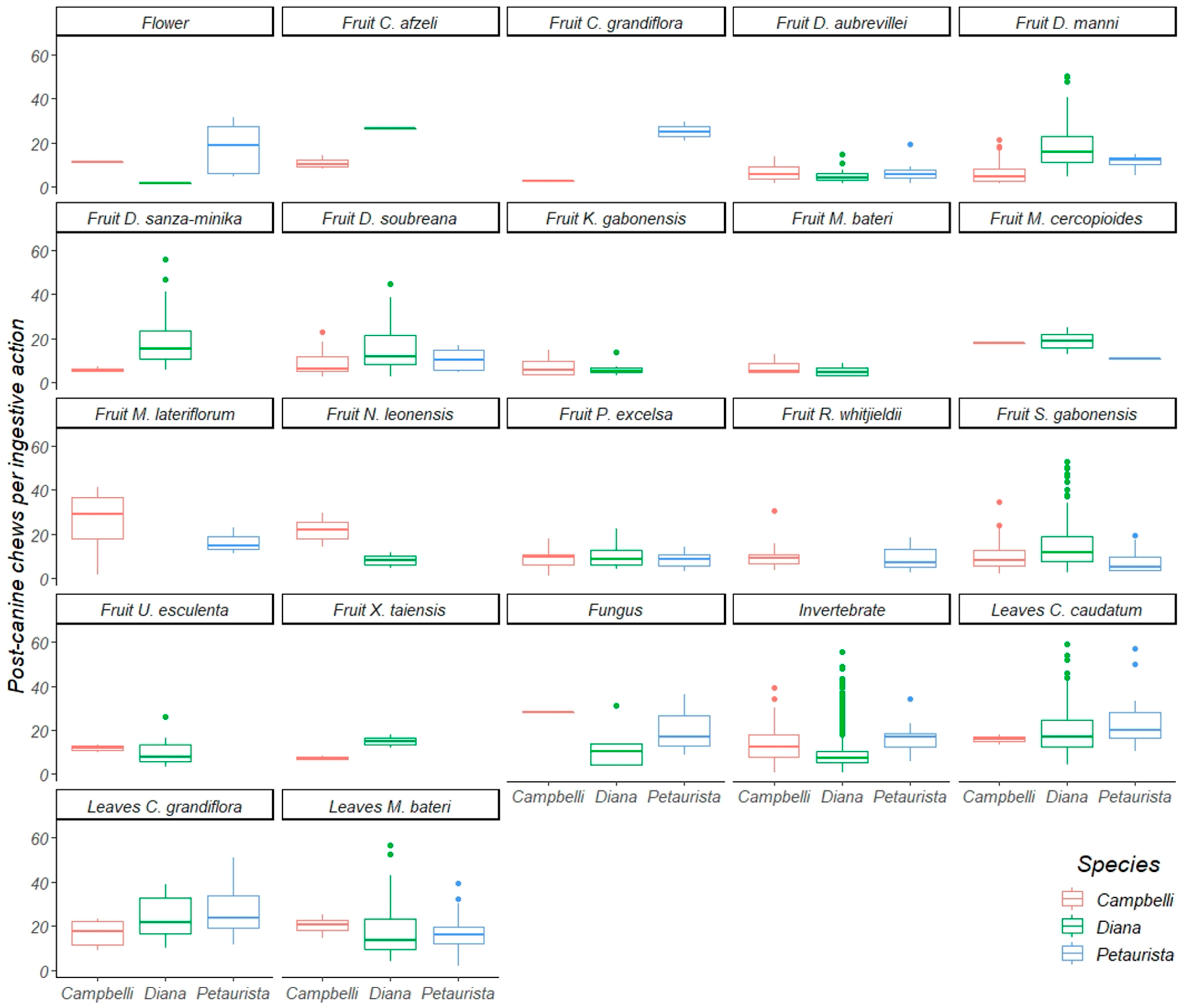
3.1. Oral Processing of Fruit
3.2. Oral Processing of Foliage
3.3. Oral Processing of Invertebrates, Fungi, and Flowers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bshary, R.; Noe, R. Red colobus and Diana monkeys provide mutual protection against predators. Anim. Behav. 1997, 54, 1461–1474. [Google Scholar] [CrossRef] [PubMed]
- Bitty, E.A.; McGraw, W.S. Locomotion and habitat use of Stampflii’s putty-nosed monkey (Cercopithecus nictitans stampflii) in Taï National Park, Ivory Coast. Am. J. Phys. Anthropol. 2007, 134, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Buzzard, P.J. Cheek pouch use in relation to interspecific competition and predator risk for three guenon monkeys (Cercopithecus spp.). Primates 2006, 47, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Buzzard, P.J. Ecological Partitioning of Cercopithecus campbelli, C. petaurista, and C. diana in the Taï Forest. Inter. J. Primatol. 2006, 27, 529–558. [Google Scholar] [CrossRef]
- Buzzard, P.J. Polyspecific associations of Cercopithecus campbelli and C. petaurista with C. diana: What are the costs and benefits? Primates 2010, 51, 307–314. [Google Scholar] [CrossRef] [PubMed]
- McGraw, W.S.; Bshary, R. Association of terrestrial mangabeys (Cercocebus atys) with arboreal monkeys: Experimental evidence for the effects of reduced ground predator pressure on habitat use. Int. J. Primatol. 2002, 23, 311–325. [Google Scholar] [CrossRef]
- Eckardt, W.; Zuberbühler, K. Cooperation and competition in two forest monkeys. Behav. Ecol. 2004, 15, 400–411. [Google Scholar] [CrossRef] [Green Version]
- Wachter, B.; Schabel, M.; Noë, R. Diet Overlap and Polyspecific Associations of Red Colobus and Diana Monkeys in the Taï National Park, Ivory Coast. Ethology 1997, 103, 514–526. [Google Scholar] [CrossRef]
- Wolters, S.; Zuberbühler, K. Mixed-Species Associations of Diana and Campbell’s Monkeys: The Costs and Benefits of a Forest Phenomenon. Behaviour 2003, 140, 371–385. [Google Scholar] [CrossRef] [Green Version]
- Buzzard, P.J. Interspecific Competition among Cercopithecus campbelli, C. petaurista, and C. diana at Taï Forest, Cote d’Ivoire. Ph.D. Thesis, Columbia University, New York, NY, USA, 2004. [Google Scholar]
- Buzzard, P.J.; Eckardt, W. The social systems of the guenons. In Monkeys of the Taï Forest: An African Primate Community; McGraw, W.S., Zuberbühler, K., Noe, R., Eds.; Cambridge University Press: New York, NY, USA, 2007; pp. 51–71. [Google Scholar]
- Cope, D.A. Variation as Indicators of Multiple Taxa in Samples of Sympatric Cercopithecus Species. In Species, Species Concepts, and Primate Evolution; Kimbel, W.H., Martin, L.B., Eds.; Springer Science+Business Media: New York, NY, USA, 1993; pp. 211–237. [Google Scholar]
- Cardini, A.; Elton, S. Variation in guenon skulls (I): Species divergence, ecological and genetic differences. J. Hum. Evol. 2008, 54, 615–637. [Google Scholar] [CrossRef]
- Martin, R.D.; MacLarnon, A.M. Quantitative comparisons of the skulls and teeth in guenons. In A Primate Radiation: Evolutionary Biology of the African Guenons; Gautier-Hion, A., Bourliere, J.P., Gautier, A., Kingdon, J., Eds.; Cambridge University Press: New York, NY, USA, 1988; pp. 160–183. [Google Scholar]
- Kay, R.F. Molar structure and diet in extant cercopithecidae. In Development, Function and Evolution of Teeth; Butler, P.M., Joysey, K.A., Eds.; Academic Press: London, UK, 1978; pp. 309–340. [Google Scholar]
- Oates, J.F.; Whitesides, G.H.; Davies, A.G.; Waterman, P.G.; Green, S.M.; Dasilva, G.L.; Mole, S. Determinants of Variation in Tropical Forest Primate Biomass: New Evidence from West Africa. Ecology 1990, 71, 328–343. [Google Scholar] [CrossRef]
- Fannin, L.D.; Guatelli-Steinberg, D.; Geissler, E.; Constantino, P.; McGraw, W.S. Frequencies of enamel chipping in Taï Forest Cercopithecoids: Insights for paleoanthropology. J. Hum. Evol. 2020, 141, 102742. [Google Scholar] [CrossRef] [PubMed]
- Koyabu, D.B.; Endo, H.; Shimizu, D.; Hashimoto, C.; Furuichi, T.; Tashiro, Y.; Go, M.; Ihobe, H. Food toughness and craniodental morphology in three sympatric guenon species in Kalinzu Forest, Uganda. In Proceedings of the Congress of the International Primatological Society, Kyoto, Japan, 12–18 September 2010. Abstract # 798. [Google Scholar]
- Gautier-Hion, A. Polyspecific associations among forest guenons: Ecological, behavioural, and evolutionary aspects. In A Primate Radiation: Evolutionary Biology of the African Guenons; Gautier-Hion, A., Bourliere, F., Gautier, J.P., Kingdon, J., Eds.; Cambridge University Press: New York, NY, USA, 1988; pp. 452–476. [Google Scholar]
- Chapman, C.A.; Chapman, L.J.; Cords, M.; Gathua, J.M.; Gautier-Hion, A.; Lambert, J.E.; Rode, K.; Tutin, C.E.G.; White, L.J.T. Variation in the diets of Cercopithecus species: Differences within forests, among forests, and across species. In:The Guenons: Diversity and Adaptation in African Monkeys; Cords, M., Glenn, M.E., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2002; pp. 325–350. [Google Scholar]
- Enstam, K.L.; Isbell, L.A. The guenons (genus Cercopithecus) and their allies: Behavioral ecology of polyspecific associations. In Primates in Perspective; Campbell, C.J., Fuentes, A., MacKinnon, K.C., Beard, S., Stumpf, R., Eds.; Oxford University Press: New York, NY, USA, 2007; pp. 252–274. [Google Scholar]
- Kane, E.E.; McGraw, W.S. Dietary Variation in Diana Monkeys (Cercopithecus diana): The Effects of Polyspecific Associations. Folia Primatol. 2007, 88, 455–482. [Google Scholar] [CrossRef]
- Ungar, P.S. Patterns of ingestive behavior and anterior tooth use differences in sympatric anthropoid primates. Am. J. Phys. Anthropol. 1994, 95, 197–219. [Google Scholar] [CrossRef] [PubMed]
- Wright, B.W.; Ulibarri, L.; O’Brien, J.; Sadler, B.; Prodhan, R.; Covert, H.H.; Nadler, T. It’s tough out there: Variation in the toughness of ingested leaves and feeding behavior among four colobinae in Vietnam. Int. J. Primatol. 2008, 29, 1455–1466. [Google Scholar] [CrossRef]
- Ross, C.F.; Iriarte-Diaz, J.; Nunn, C.L. Innovative approaches to the relationship between diet and mandibular morphology in primates. Int. J. Primatol. 2012, 33, 632–660. [Google Scholar] [CrossRef]
- Vogel, E.R.; Zulfa, A.; Hardus, M.; Wich, S.A.; Dominy, N.J.; Taylor, A.B. Food mechanical properties, feeding ecology, and the mandibular morphology of wild orangutans. J. Hum. Evol. 2014, 75, 110–124. [Google Scholar] [CrossRef]
- Yamashita, N. Diets of two lemur species in different microhabitats in Beza Mahafaly Special Reserve, Madagascar. Int. J. Primatol. 2002, 23, 1025–1051. [Google Scholar] [CrossRef]
- Yamashita, N. Food procurement and tooth use in two sympatric lemur species. Am. J. Phys. Anthropol. 2003, 121, 125–133. [Google Scholar] [CrossRef]
- Cuozzo, F.P.; Sauther, M.L. Severe wear and tooth loss in wild ring-tailed lemurs (Lemur catta): A function of feeding ecology, dental structure, and individual life history. J. Hum. Evol. 2006, 51, 490–505. [Google Scholar] [CrossRef]
- Yamashita, N. Mechanical threshholds as a criterion for food selection in two prosimian primate species. In Proceedings of the 3rd Plant Biomechanics Conference, Freiburg/Badenweiler, Germany, 27 August–2 September 2000; Spatz, H.-C., Speck, T., Eds.; Thieme-Verlag: Stuttgart, Germany, 2000; pp. 590–595. [Google Scholar]
- Yamashita, N. Chemical properties of the diets of two lemur species in southwestern Madagascar. Int. J. Priamtol. 2008, 29, 339–364. [Google Scholar] [CrossRef]
- Yamashita, N.; Vinyard, C.J.; Tan, C.L. Food mechanical properties in three sympatric species of Hapalemur in Ranomafana National Park, Madagascar. Am. J. Phys. Anthropol. 2009, 139, 368–381. [Google Scholar] [CrossRef]
- Laird, M.F.; Wright, B.W.; Rivera, A.O.; Fogaça, M.D.; van Casteren, A.; Fragaszy, D.M.; Izar, P.; Visalberghi, E.; Scott, R.S.; Strait, D.S.; et al. Ingestive behaviors in bearded capuchins (Sapajus libidinosus). Sci. Rep. 2020, 10, 20850. [Google Scholar] [CrossRef] [PubMed]
- Laird, M.F.; Punjani, Z.; Oshay, R.R.; Wright, B.W.; Fogaça, M.D.; van Casteren, A.; Izar, P.; Visalberghi, E.; Fragaszy, D.; Strait, D.S.; et al. Feeding postural behaviors and food gemetric and material properties in bearded capuchin monkeys (Sapajust libidinosus). Am. J. Biol. Anthropol. 2022, 178, 3–16. [Google Scholar] [CrossRef]
- Chalk-Wilayto, J.; Fogaça, M.D.; Wright, B.W.; van Casteren, A.; Fragaszy, D.M.; Izar, P.; Visalberghi, E.; Strait, D.S.; Ross, C.F.; Wright, K.A.; et al. Effects of food material properties and embedded status on food processing efficiency in bearded capuchins. Am. J. Biol. Anthropol. 2022, 178, 617–635. [Google Scholar] [CrossRef]
- Taylor, A.B.; Vinyard, C.J. The relationships among jaw-muscle fiber architecture, jaw morphology, and feeding in extant apes and modern humans. Am. J. Phys. Anthropol. 2013, 151, 120–134. [Google Scholar] [CrossRef]
- Ross, C.F.; Iriarte-Diaz, J.; Reed, D.A.; Stewart, T.A.; Taylor, A.B. In vivo bone strain in the mandibular corpus of Sapajus during a range of oral food processing behaviors. J. Hum. Evol. 2016, 98, 36–65. [Google Scholar] [CrossRef] [Green Version]
- van Casteren, A.; Wright, E.; Kupczik, K.; Robbins, M.M. Unexpected hard-object feeding in Western lowland gorillas. Am. J. Phys. Anthropol. 2019, 170, 433–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, E.E.; Traff, J.N.; Daegling, D.J.; McGraw, W.S. Oral Processing Behavior of Diana Monkeys (Cercopithecus diana) in Taï National Park, Côte d’Ivoire. Folia Primatol. 2020, 91, 541–557. [Google Scholar] [CrossRef]
- McGraw, W.S.; Vick, A.E.; Daegling, D.J. Sex and age differences in the diet and ingestive behaviors of sooty mangabeys (Cercocebus atys) in the Taï forest, Ivory Coast. Am. J. Phys. Anthropol. 2011, 144, 140–153. [Google Scholar] [CrossRef]
- McGraw, W.S.; Daegling, D.J. Primate Feeding and Foraging: Integrating Studies of Behavior and Morphology. Annu. Rev. Anthropol. 2012, 41, 203–219. [Google Scholar] [CrossRef]
- Daegling, D.J.; Hotzman, J.L.; McGraw, W.S.; Rapoff, A.J. Material property variation of mandibular symphyseal bone in colobine monkeys. J. Morphol. 2009, 270, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Daegling, D.J.; McGraw, W.S. Masticatory stress and the mechanics of “wishboning” in colobines jaws. Am. J. Phys. Anthropol. 2009, 138, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Daegling, D.J.; Granatosky, M.; McGraw, W.S.; Rapoff, A.J. Reduced stiffness in the alveolar bone of the colobine mandible. Am. J. Phys. Anthropol. 2011, 144, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Daegling, D.J.; McGraw, W.S.; Ungar, P.S.; Pampush, J.D.; Vick, A.E.; Bitty, E.A. Hard-Object Feeding in Sooty Mangabeys (Cercocebus atys) and Interpretation of Early Hominin Feeding Ecology. PLoS ONE 2011, 6, e23095. [Google Scholar] [CrossRef]
- Daegling, D.J.; Granatosky, M.C.; McGraw, W.S. Spatial patterning of bone stiffness in the anterior mandibular corpus of Macaca fascicularis: Implications for models of bone adaptation. Am. J. Phys. Anthropol. 2015, 156, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Lad, S.E.; Daegling, D.J.; McGraw, W.S. Bone remodeling is reduced in high stress regions of the cercopithecoid mandible. Am. J. Phys. Anthropol. 2016, 161, 426–435. [Google Scholar] [CrossRef]
- McGraw, W.S.; Daegling, D.J. Diet, feeding behavior, and jaw architecture of Taï monkeys: Congruence and chaos in the realm of functional morphology. Evol. Anthropol. 2020, 29, 14–28. [Google Scholar] [CrossRef]
- McGraw, W.S.; Pampush, J.D.; Daegling, D.J. Brief communication: Enamel thickness and durophagy in mangabeys revisited. Am. J. Phys. Anthropol. 2012, 147, 326–333. [Google Scholar] [CrossRef]
- McGraw, W.S.; Vick, A.E.; Daegling, D.J. Dietary variation and food hardness in sooty mangabeys (Cercocebus atys): Implications for fallback foods and dental adaptation: Seasonality Of Feeding In Sooty Mangabeys. Am. J. Phys. Anthropol. 2014, 154, 413–423. [Google Scholar] [CrossRef]
- McGraw, W.S.; van Casteren, A.; Kane, E.; Geissler, E.; Burrows, B.; Daegling, D.J. Feeding and oral processing behaviors of two colobine monkeys in Taï Forest, Ivory Coast. J. Hum. Evol. 2016, 98, 90–102. [Google Scholar] [CrossRef] [Green Version]
- Ray, T.J.; McGraw, W.S.; Sun, Z.; Jeon, M.; Johnson, T.; Cheffins, K.; Daegling, D.J.; Kim, D.-G. Mandibular bone mineral density variation in three West African cercopithecoid monkey species: Associations with diet and feeding behavior. Arch. Oral Biol. 2015, 60, 1714–1720. [Google Scholar] [CrossRef]
- Traff, J.N.; McGraw, W.S.; Daegling, D.J. Masticatory and ingestive effort in Procolobus verus, a small-bodied African colobine. Primates 2022, 63, 271–282. [Google Scholar] [CrossRef]
- Morse, P.E.; Daegling, D.J.; McGraw, W.S.; Pampush, J.D. Dental wear among cercopithecid monkeys of the Taï Forest, Cote d’Ivoire. Am. J. Phys. Anthropol. 2013, 150, 655–665. [Google Scholar] [CrossRef]
- Calhoun, G.; Guatelli-Steinberg, D.; Morse, P.; McGraw, W.S. Dental microwear, diet and anterior tooth use in Colobus and Piliocolobus. J. Hum. Evol. 2022, 163, 103123. [Google Scholar] [CrossRef]
- Boesch, C.; Boesch, H. Optimisation of nut cracking with natural hammers by wild chimpanzees. Behaviour 1983, 83, 265–286. [Google Scholar] [CrossRef] [Green Version]
- McGraw, W.S.; Zuberbühler, K. The Monkeys of the Taï forest: An introduction. In Monkeys of the Taï Forest: An African Primate Community; McGraw, W.S., Zuberbühler, K., Noë, R., Eds.; Cambridge Studies in Biological and Evolutionary Anthropology: Volume 51; Cambridge University Press: Cambridge, UK, 2007; pp. 1–48. [Google Scholar]
- McGraw, W.S.; Zuberbühler, K. Socioecology, predation, and cognition in a community of West African monkeys. Evol. Anthropol. 2008, 17, 254–266. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.P.; Nordheim, E.V.; Moermond, T.C.; Gone Bi, Z.B.; Boesch, C. Factors Influencing Tree Phenology in Taï National Park, Cote d’Ivoire. Biotropica 2005, 37, 631–640. [Google Scholar] [CrossRef]
- Lambert, J.E. Digestive retention times in Forest Guenons (Cercopithecus spp.) with reference to chimpanzees (Pan troglodtyes). Int. J. Primatol. 2002, 23, 1169–1185. [Google Scholar] [CrossRef]
- Lambert, J.E. Competition, predation, and the evolutionary significance of the cercopithecine cheek pouch: The case of Cercopithecus and Lophocebus. Am. J. Phys. Anthropol. 2005, 126, 183–192. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Hylander, W.L. In-vivo bone strain as an indicator of masticatory bite force in Macaca fascicularis. Arch. Oral Biol. 1986, 31, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Bunn, J.M.; Ungar, P.S. Dental topography and diets of four Old World monkey species. Am. J. Primatol. 2009, 71, 466–477. [Google Scholar] [CrossRef]
- Sheine, W.S.; Kay, R.F. An analysis of chewed food particle size and its relationship to molar structure in the primates Cheirogaleus medius and Galago senegalensis and the insectivoran Tupaia glis. Am. J. Phys. Anthropol. 1977, 47, 15–20. [Google Scholar] [CrossRef]
- St Clair, E.M.; Boyer, D.M. Lower molar shape and size in prosimian and platyrrhine primates. Am. J. Phys. Anthropol. 2016, 161, 237–258. [Google Scholar] [CrossRef]
- Robinson, B.W.; Wilson, D.S. Optimal foraging, specialization, and a solution to Liem’s paradox. Am. Nat. 1998, 151, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Ungar, P.S. Tooth form and function: Insights into adaptation through the analysis of dental microwear. Front. Oral Biol. 2009, 13, 38–43. [Google Scholar] [PubMed]
- Ungar, P.S.; Healy, C.; Karme, A.; Teaford, M.; Fortelius, M. Dental topography and diets of platyrrhine primates. Hist. Biol. 2018, 30, 64–75. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Age–Sex Class | Individual ID | Number of Focals |
|---|---|---|---|
| C. diana | Adult female | Unknown | 300 |
| C. diana | Adult female | Agnes | 207 |
| C. diana | Adult female | Cheri | 203 |
| C. diana | Adult female | Emi | 165 |
| C. diana | Adult female | Eva | 321 |
| C. diana | Adult female | Nina | 201 |
| C. diana | Adult female | Sonia | 202 |
| C. diana | Adult female | Veronique | 239 |
| C. campbelli | Adult female | Agathe | 75 |
| C. campbelli | Adult female | Caro | 11 |
| C. campbelli | Adult female | Florence | 126 |
| C. campbelli | Adult female | Lucie | 73 |
| C. campbelli | Adult female | Ode | 70 |
| C. campbelli | Adult male | Domi | 40 |
| C. petaurista | Adult female | Unknown | 198 |
| C. petaurista | Adult male | Unknown | 118 |
| Species | Focals | Flowers | Fruit | Invertebrates | Leaves | Fungi | Seeds | Stems |
|---|---|---|---|---|---|---|---|---|
| C. diana | 1838 | 1 | 435 | 1161 | 227 | 8 | 0 | 6 |
| C. campbelli | 395 | 1 | 239 | 104 | 21 | 1 | 14 | 14 |
| C. petaurista | 316 | 7 | 67 | 24 | 192 | 3 | 3 | 20 |
| Diana Monkeys (C. diana) | Campbell’s Monkeys (C. campbelli) | Lesser Spot-Nosed Monkeys (C. petaurista) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Focals (N) | Action (N) | I/A 1 | C/A 2 | PC/A 3 | Focals (N) | Action (N) | I/A | C/A | PC/A | Focals (N) | Action (N) | I/A | C/A | PC/A | |
| All focals | 1838 | 21,275 | 1.27 | 0.00 | 8.82 | 395 | 4468 | 1.00 | 0.00 | 9.58 | 315 | 3099 | 0.82 | 0.00 | 14.39 |
| Fruit | 582 | 11,657 | 2.07 | 0.00 | 19.48 | 223 | 2574 | 0.86 | 0.01 | 10.15 | 73 | 1156 | 0.88 | 0.05 | 9.91 |
| Cersalia afzeli | 1 | 8 | 1.63 | 0.00 | 26.38 | 4 | 13 | 0.88 | 0.00 | 10.43 | |||||
| Coelecarion oxycarpum | 1 | 10 | 1.10 | 0.00 | 4.30 | 1 | 12 | 1.00 | 0.00 | 5.50 | |||||
| Culcasia grandiflora | 1 | 21 | 1.00 | 0.00 | 2.80 | 2 | 11 | 1.00 | 0.00 | 25.13 | |||||
| Dialium aubrevillei | 27 | 789 | 1.30 | 0.00 | 8.90 | 15 | 156 | 0.91 | 0.00 | 6.96 | 7 | 92 | 0.85 | 0.00 | 7.21 |
| Diospyros manni | 148 | 2547 | 1.92 | 0.00 | 37.74 | 25 | 241 | 0.95 | 0.02 | 7.05 | 7 | 78 | 0.85 | 0.15 | 11.32 |
| Diospyros sanza-minika | 22 | 305 | 1.90 | 0.00 | 21.14 | 3 | 33 | 1.00 | 0.00 | 5.75 | |||||
| Diospyros soubreana | 61 | 936 | 4.58 | 0.00 | 16.24 | 8 | 61 | 0.95 | 0.03 | 9.42 | 4 | 33 | 1.00 | 0.00 | 10.37 |
| Klainodoxa gabonensis | 4 | 54 | 0.8 | 0.16 | 7.41 | 19 | 532 | 1.48 | 0.00 | 5.64 | |||||
| Maesobotria bateri | 13 | 602 | 0.83 | 0.00 | 5.17 | 3 | 26 | 0.00 | 0.00 | 7.32 | |||||
| Musanga cercopioides | 2 | 13 | 3.69 | 0.00 | 18.76 | 1 | 2 | 0.00 | 0.00 | 17.67 | 1 | 7 | 0.86 | 0.00 | 10.71 |
| Napoleona leonensis | 2 | 30 | 1.33 | 0.00 | 8.29 | 2 | 9 | 1.00 | 0.00 | 21.93 | |||||
| Parinari excelsa | 25 | 536 | 2.77 | 0.00 | 10.93 | 9 | 184 | 0.88 | 0.00 | 9.28 | 14 | 348 | 0.92 | 0.08 | 8.64 |
| Rothmannia whitjieldii | 13 | 181 | 0.97 | 0.04 | 10.43 | 6 | 138 | 0.66 | 0.25 | 9.33 | |||||
| Sacoglottis gabonensis | 117 | 2057 | 2.98 | 0.00 | 17.36 | 59 | 781 | 0.96 | 0.00 | 9.73 | 15 | 338 | 0.70 | 0.00 | 17.65 |
| Uapaca esculenta | 11 | 360 | 1.36 | 0.00 | 10.31 | 2 | 37 | 0.91 | 0.09 | 11.72 | |||||
| Xylopia taiensis | 2 | 30 | 1.53 | 0.00 | 15.02 | 2 | 28 | 1.00 | 0.00 | 7.31 | |||||
| Leaves (young) | 235 | 2809 | 3.09 | 0.00 | 15.73 | 17 | 71 | 0.73 | 0.00 | 14.69 | 142 | 1136 | 0.56 | 0.00 | 17.24 |
| Craterispermum caudatum | 60 | 774 | 2.64 | 0.00 | 14.96 | 1 | 6 | 1.00 | 0.00 | 13.60 | 13 | 75 | 0.69 | 0.00 | 18.42 |
| Culcasia grandiflora | 16 | 143 | 4.84 | 0.00 | 23.69 | 5 | 18 | 0.86 | 0.00 | 16.53 | 16 | 87 | 0.77 | 0.00 | 26.53 |
| Maesobotria bateri | 48 | 575 | 2.76 | 0.00 | 14.69 | 4 | 15 | 0.79 | 0.00 | 18.33 | 36 | 311 | 0.67 | 0.00 | 15.77 |
| Pauridiantha sylvicola | 1 | 1 | 4.00 | 0.00 | 57.00 | 2 | 21 | 0.75 | 0.00 | 20.05 | |||||
| Leaves (mature) | 350 | 4157 | 2.49 | 0.00 | 24.15 | 4 | 25 | 0.29 | 0.00 | 20.56 | 50 | 428 | 0.82 | 0.00 | 23.17 |
| Craterispermum caudatum | 74 | 848 | 2.57 | 0.00 | 23.86 | 1 | 5 | 0.00 | 0.00 | 18.00 | 5 | 28 | 0.60 | 0.00 | 38.40 |
| Maesobotria bateri | 30 | 440 | 2.11 | 0.00 | 22.12 | 2 | 10 | 0.38 | 0.00 | 24.06 | 22 | 230 | 0.78 | 0.00 | 18.31 |
| Invertebrates | 1164 | 9459 | 1.14 | 0.00 | 9.24 | 104 | 306 | 0.58 | 0.04 | 13.22 | 24 | 36 | 0.60 | 0.00 | 15.81 |
| All other | 14 | 376 | 10 | 43 | 5 | 28 | |||||||||
| Fungi | 8 | 160 | 1.17 | 0.00 | 11.44 | 1 | 1 | 0.00 | 0.00 | 28.00 | 3 | 20 | 0.25 | 0.00 | 17.20 |
| Diospyros manni seeds | 9 | 42 | 7.15 | 0.00 | 24.27 | 2 | 3 | 5.00 | 0.00 | 73.25 | |||||
| Flowers | 1 | 159 | 1.06 | 0.00 | 1.45 | 1 | 2 | 1.00 | 0.00 | 11.00 | 1 | 17 | 0.06 | 0.00 | 4.53 |
| Trichosypha arborea stems | 5 | 80 | 1.51 | 0.00 | 7.39 | 1 | 6 | 1.00 | 0.00 | 12.33 | |||||
| Incisions per Ingestive Action: Resampled Mean Pairwise Comparisons | Postcanine Chews per Ingestive Action: Resampled Mean Pairwise Comparisons | |||
|---|---|---|---|---|
| Species Comparison | p-Value | Species Comparison | p-Value | |
| Fruit | C. diana > C. campbelli | 0 | C. diana > C. campbelli | 0 |
| C. diana > C. petaurista | 0 | C. diana > C. petaurista | 0 | |
| C. campbelli < C. petaurista | 0 | C. campbelli < C. petaurista | 0.04 | |
| Cersalia afzeli (ripe) | C. diana > C. campbelli | 0 | C. diana > C. campbelli | 0.02 |
| C. diana, C. petaurista | NA | C. diana, C. petaurista | NA | |
| C. campbelli, C. petaurista | NA | C. campbelli, C. petaurista | NA | |
| Coelecarion oxycarpum (ripe) | C. diana > C. campbelli | 0 | C. diana = C. campbelli | 0.56 |
| C. diana, C. petaurista | NA | C. diana, C. petaurista | NA | |
| C. campbelli, C. petaurista | NA | C. campbelli, C. petaurista | NA | |
| Culcasia grandiflora (ripe) | C. diana, C. campbelli | NA | C. diana, C. campbelli | NA |
| C. diana, C. petaurista | NA | C. diana, C. petaurista | NA | |
| C. campbelli < C. petaurista | 0 | C. campbelli < C. petaurista | 0 | |
| Dialium aubrevillei (ripe) | C. diana > C. campbelli | 0 | C. diana > C. campbelli | 0 |
| C. diana > C. petaurista | 0 | C. diana > C. petaurista | 0.01 | |
| C. campbelli = C. petaurista | 0.11 | C. campbelli < C. petaurista | 0.02 | |
| Diospyros manni (ripe) | C. diana > C. campbelli | 0 | C. diana > C. campbelli | 0 |
| C. diana > C. petaurista | 0 | C. diana > C. petaurista | 0 | |
| C. campbelli < C. petaurista | 0 | C. campbelli < C. petaurista | 0 | |
| Diospyros sanza-minika (ripe) | C. diana > C. campbelli | 0 | C. diana > C. campbelli | 0 |
| C. diana, C. petaurista | NA | C. diana, C. petaurista | NA | |
| C. campbelli, C. petaurista | NA | C. campbelli, C. petaurista | NA | |
| Diospyros soubreana (ripe) | C. diana > C. campbelli | 0 | C. diana > C. campbelli | 0 |
| C. diana > C. petaurista | 0 | C. diana > C. petaurista | 0 | |
| C. campbelli = C. petaurista | 0.49 | C. campbelli = C. petaurista | 0.14 | |
| Klainodoxa gabonensis (ripe) | C. diana = C. campbelli | 0.12 | C. diana = C. campbelli | 0.12 |
| C. diana, C. petaurista | NA | C. diana, C. petaurista | NA | |
| C. campbelli, C. petaurista | NA | C. campbelli, C. petaurista | NA | |
| Maesobotria bateri (ripe) | C. diana > C. campbelli | 0 | C. diana > C. campbelli | 0 |
| C. diana, C. petaurista | NA | C. diana, C. petaurista | NA | |
| C. campbelli, C. petaurista | NA | C. campbelli, C. petaurista | NA | |
| Memecylon lateriflorum (Unripe) | C. diana, C. campbelli | NA | C. diana, C. campbelli | NA |
| C. diana, C. petaurista | NA | C. diana, C. petaurista | NA | |
| C. campbelli < C. petaurista | 0 | C. campbelli < C. petaurista | 0 | |
| Musanga cercopioides (ripe) | C. diana > C. campbelli | 0 | C. diana = C. campbelli | 0.15 |
| C. diana > C. petaurista | 0 | C. diana = C. petaurista | 0.1 | |
| C. campbelli < C. petaurista | 0 | C. campbelli = C. petaurista | 0.48 | |
| Napoleona leonensis (ripe) | C. diana > C. campbelli | 0 | C. diana > C. campbelli | 0 |
| C. diana, C. petaurista | NA | C. diana, C. petaurista | NA | |
| C. campbelli, C. petaurista | NA | C. campbelli, C. petaurista | NA | |
| Parinari excelsa (ripe) | C. diana > C. campbelli | 0 | C. diana = C. campbelli | 0.42 |
| C. diana > C. petaurista | 0 | C. diana > C. petaurista | 0 | |
| C. campbelli = C. petaurista | 0.1 | C. campbelli > C. petaurista | 0 | |
| Rothmannia whitjieldii (ripe) | C. diana, C. campbelli | NA | C. diana, C. campbelli | NA |
| C. diana, C. petaurista | NA | C. diana, C. petaurista | NA | |
| C. campbelli = C. petaurista | 0.44 | C. cambelli < C. petaurista | 0 | |
| Sacoglottis gabonensis (ripe) | C. diana > C. campbelli | 0 | C. diana > C. campbelli | 0 |
| C. diana > C. petaurista | 0 | C. diana > C. petaurista | 0 | |
| C. cambelli < C. petaurista | 0 | C. campbelli = C. petaurista | 0.37 | |
| Uapaca esculenta (ripe) | C. diana > C. campbelli | 0 | C. diana > C. campbelli | 0.01 |
| C. diana > C. petaurista | NA | C. diana > C. petaurista | NA | |
| C. campbelli, C. petaurista | NA | C. campbelli, C. petaurista | NA | |
| Xylopia taiensis (ripe) | C. diana = C. campbelli | 0.34 | C. diana = C. campbelli | 0.07 |
| C. diana, C. petaurista | NA | C. diana > C. petaurista | NA | |
| C. campbelli, C. petaurista | NA | C. campbelli, C. petaurista | NA | |
| Foliage | C. diana > C. campbelli | 0 | C. diana > C. campbelli | 0 |
| C. diana > C. petaurista | 0 | C. diana > C. petaurista | 0 | |
| C. campbelli = C. petaurista | 0.23 | C. campbelli < C. petaurista | 0 | |
| Craterispermum caudatum (Mature) | C. diana > C. campbelli | 0 | C. diana = C. campbelli | 0.06 |
| C. diana > C. petaurista | 0 | C. diana = C. petaurista | 0.18 | |
| C. campbelli = C. petaurista | 0.44 | C. campbelli < C. petaurista | 0.04 | |
| Culcasia grandiflora (Young) | C. diana > C. campbelli | 0 | C. diana > C. petaurista | 0.01 |
| C. diana > C. petaurista | 0 | C. diana > C. petaurista | 0 | |
| C. campbelli = C. petaurista | 0.22 | C. campbelli < C. petaurista | 0.04 | |
| Maesobotria bateri (Young) | C. diana > C. campbelli | 0 | C. diana > C. campbelli | 0.02 |
| C. diana > C. petaurista | 0 | C. diana > C. petaurista | 0 | |
| C. campbelli = C. petaurista | 0.23 | C. campbelli < C. petaurista | 0.04 | |
| Maesobotria bateri (Mature) | C. diana > C. campbelli | 0 | C. diana = C. campbelli | 0.40 |
| C. diana > C. petaurista | 0 | C. diana > C. petaurista | 0 | |
| C. campbelli = C. petaurista | 0.15 | C. campbelli = C. petaurista | 0.07 | |
| Trichosypha arborea (petioles) | C. diana, C. campbelli | NA | C. diana, C. campbelli | NA |
| C. diana = C. petaurista | 0.59 | C. diana > C. petaurista | 0 | |
| C. campbelli, C. petaurista | NA | C. campbelli, C. petaurista | NA | |
| Invertebrates | C. diana > C. campbelli | 0 | C. diana = C. campbelli | 0.22 |
| C. diana > C. petaurista | 0 | C. diana = C. petaurista | 0.10 | |
| C. campbelli = C. petaurista | 0.38 | C. campbelli < C. petaurista | 0 | |
| Fungi | C. diana, C. campbelli | NA | C. diana, C. campbelli | NA |
| C. diana > C. petaurista | 0 | C. diana = C. petaurista | 0.17 | |
| C. campbelli, C. petaurista | NA | C. campbelli, C. petaurista | NA | |
| Flowers | C. diana > C. campbelli | 0 | C. diana > C. campbelli | 0 |
| C. diana > C. petaurista | 0 | C. diana = C. petaurista | 0.22 | |
| C. campbelli = C. petaurista | 0.41 | C. campbelli < C. petaurista | 0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kane, E.E.; Polvadore, T.; Bele, F.O.; Bitty, E.A.; Kamy, E.; Mehon, F.G.; Daegling, D.J.; McGraw, W.S. Oral Processing of Three Guenon Species in Taï National Park, Côte d’Ivoire. Biology 2022, 11, 1850. https://doi.org/10.3390/biology11121850
Kane EE, Polvadore T, Bele FO, Bitty EA, Kamy E, Mehon FG, Daegling DJ, McGraw WS. Oral Processing of Three Guenon Species in Taï National Park, Côte d’Ivoire. Biology. 2022; 11(12):1850. https://doi.org/10.3390/biology11121850
Chicago/Turabian StyleKane, Erin E., Taylor Polvadore, Ferdinand Ouro Bele, Eloi Anderson Bitty, Ernest Kamy, Frederic Gnepa Mehon, David J. Daegling, and William Scott McGraw. 2022. "Oral Processing of Three Guenon Species in Taï National Park, Côte d’Ivoire" Biology 11, no. 12: 1850. https://doi.org/10.3390/biology11121850