
Occurrence of Anthropogenic Debris in Three Commercial Shrimp Species from South-Western Ionian Sea
 ,
,  , ,
, ,  and
and
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
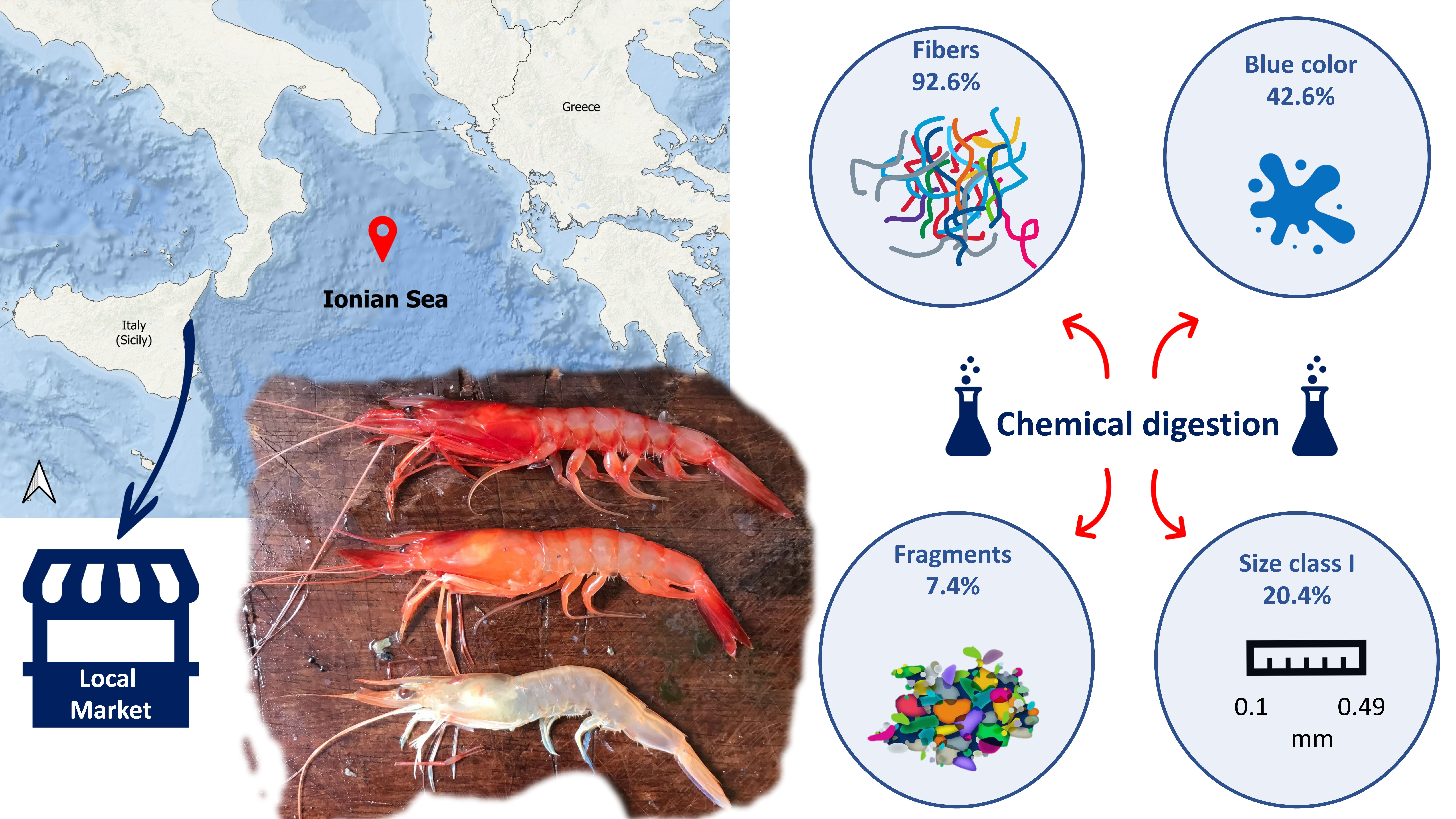
2.1. Sampling Area and Samples Processing
2.2. Anthropogenic Debris Extraction Protocol
2.3. Contamination Prevention
2.4. Data Analysis
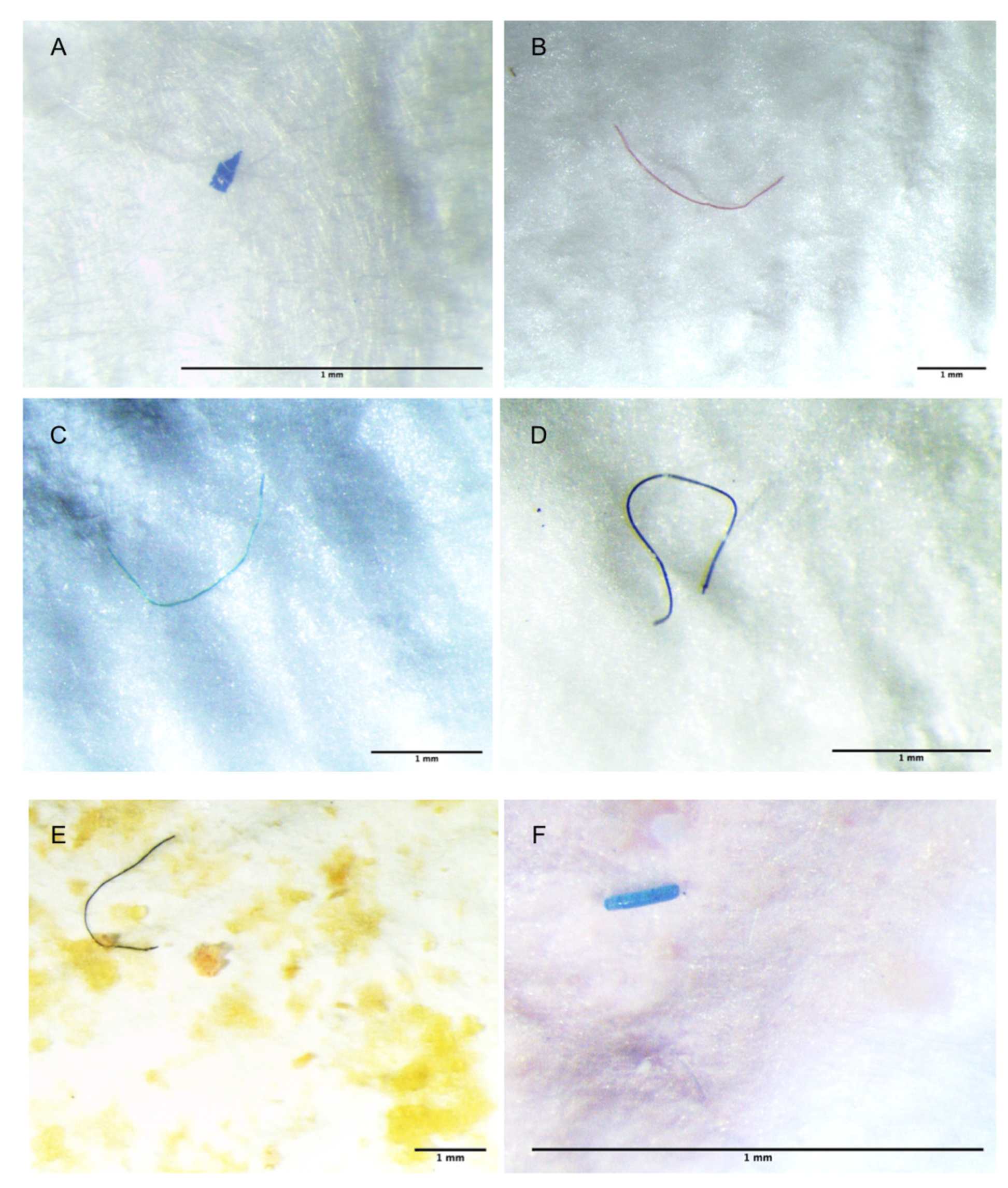
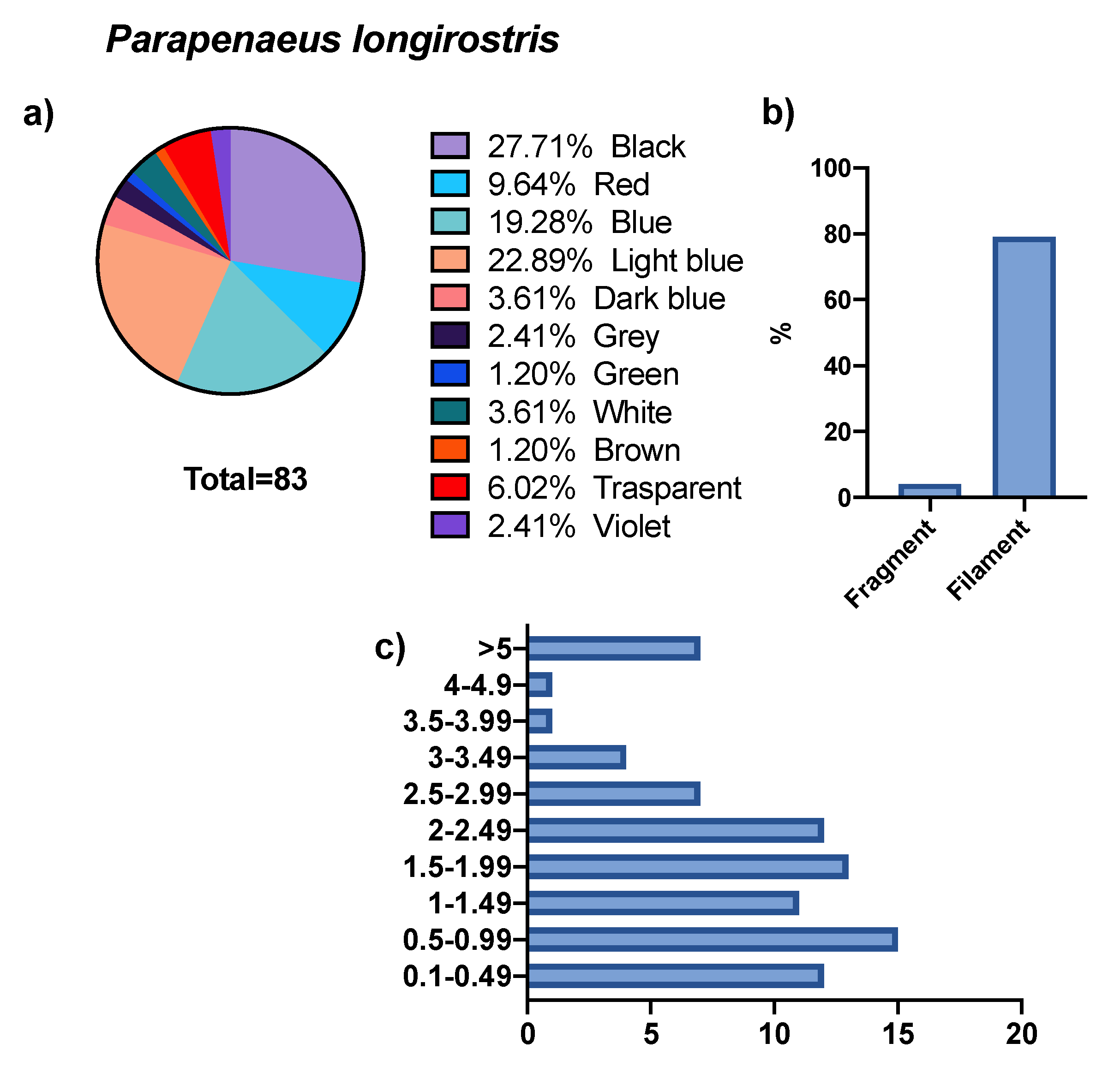
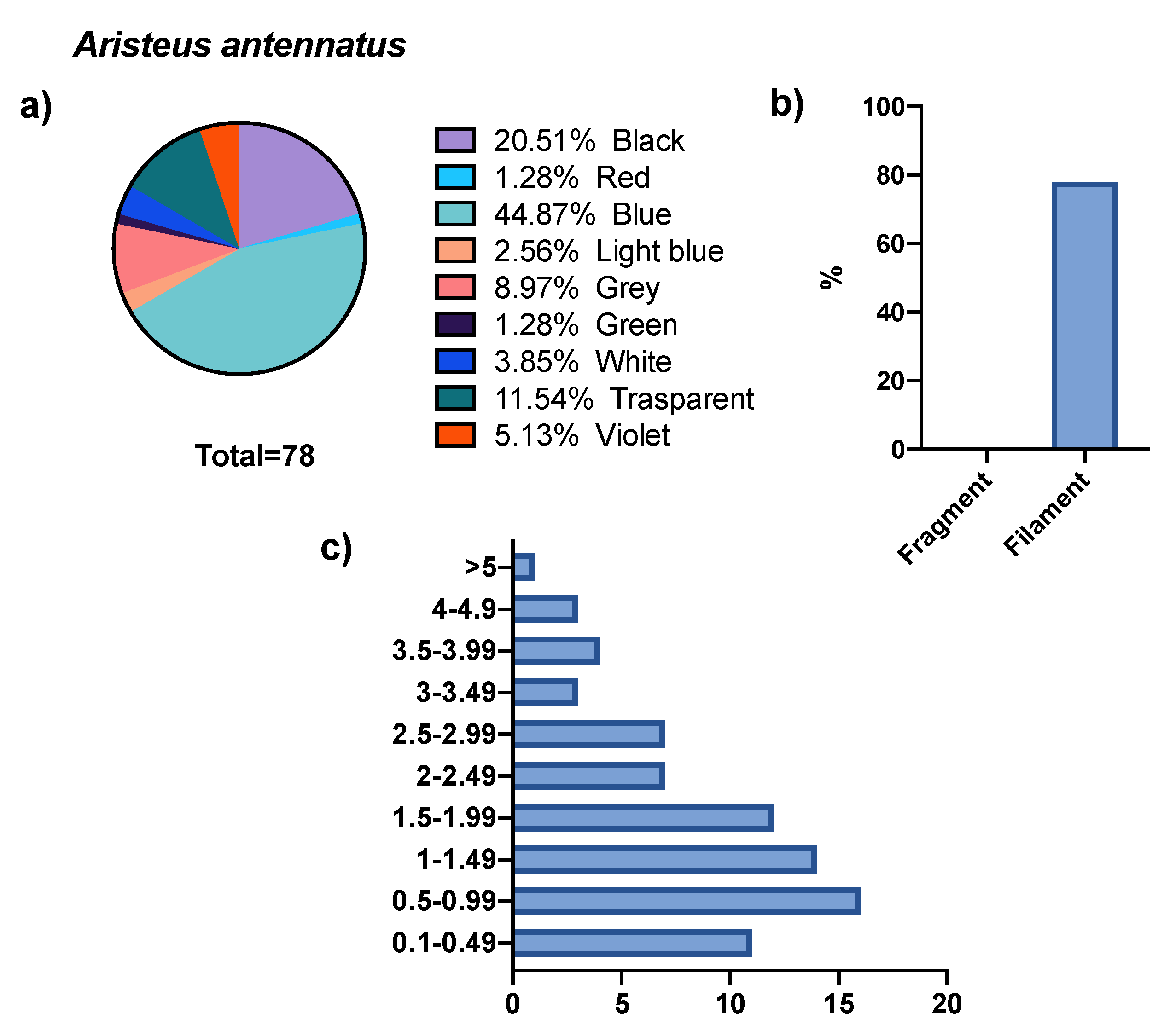
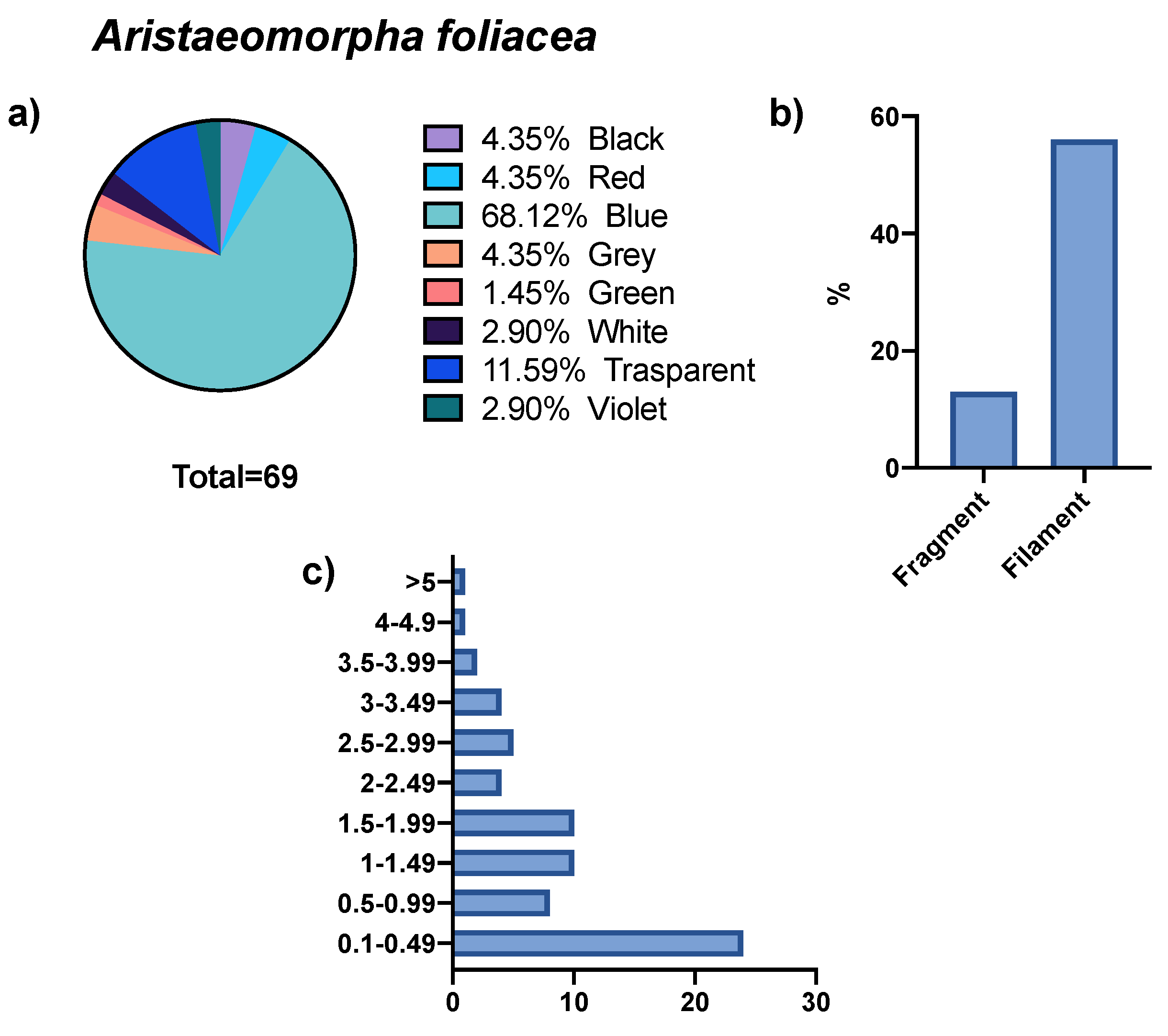
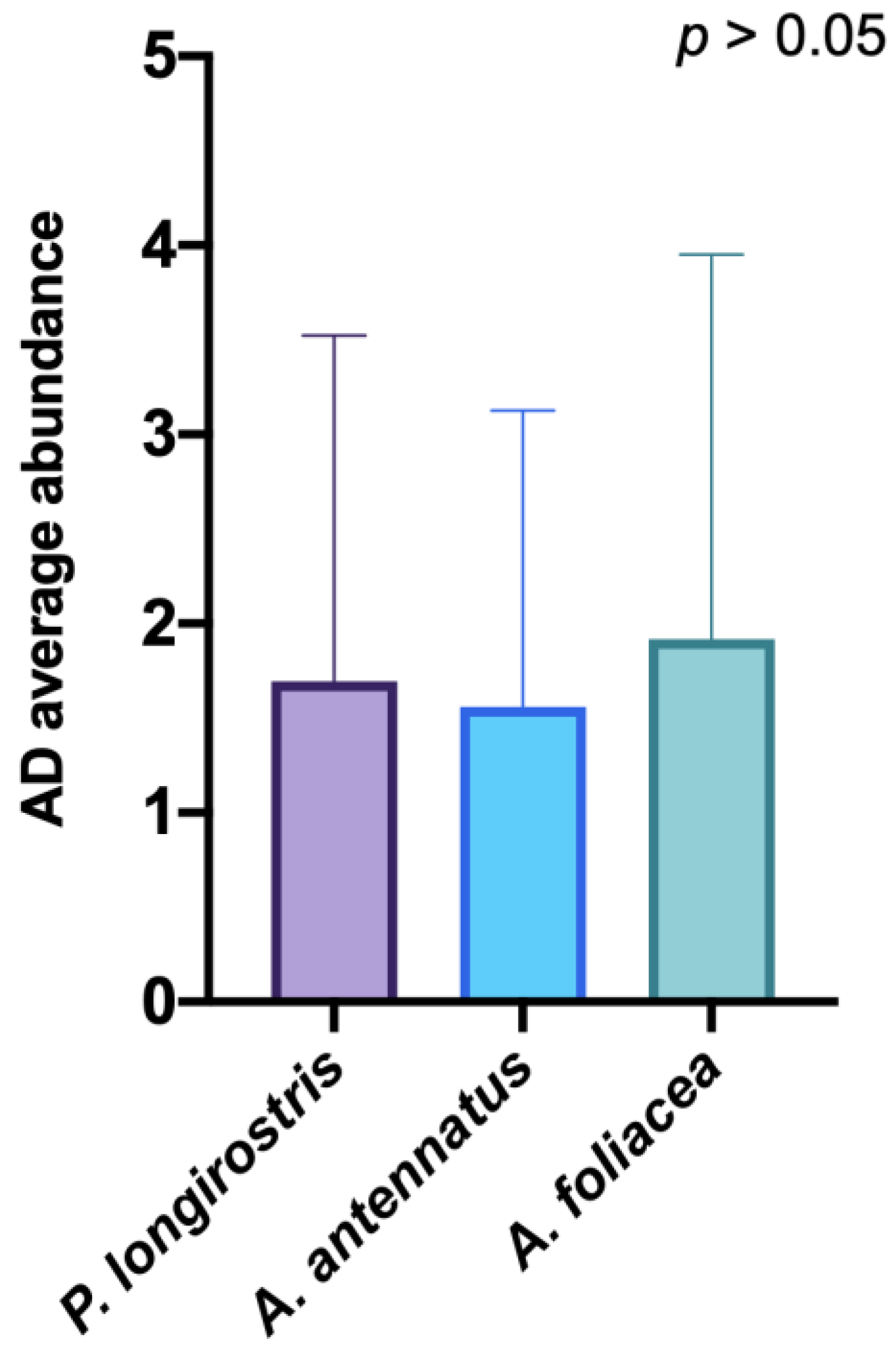
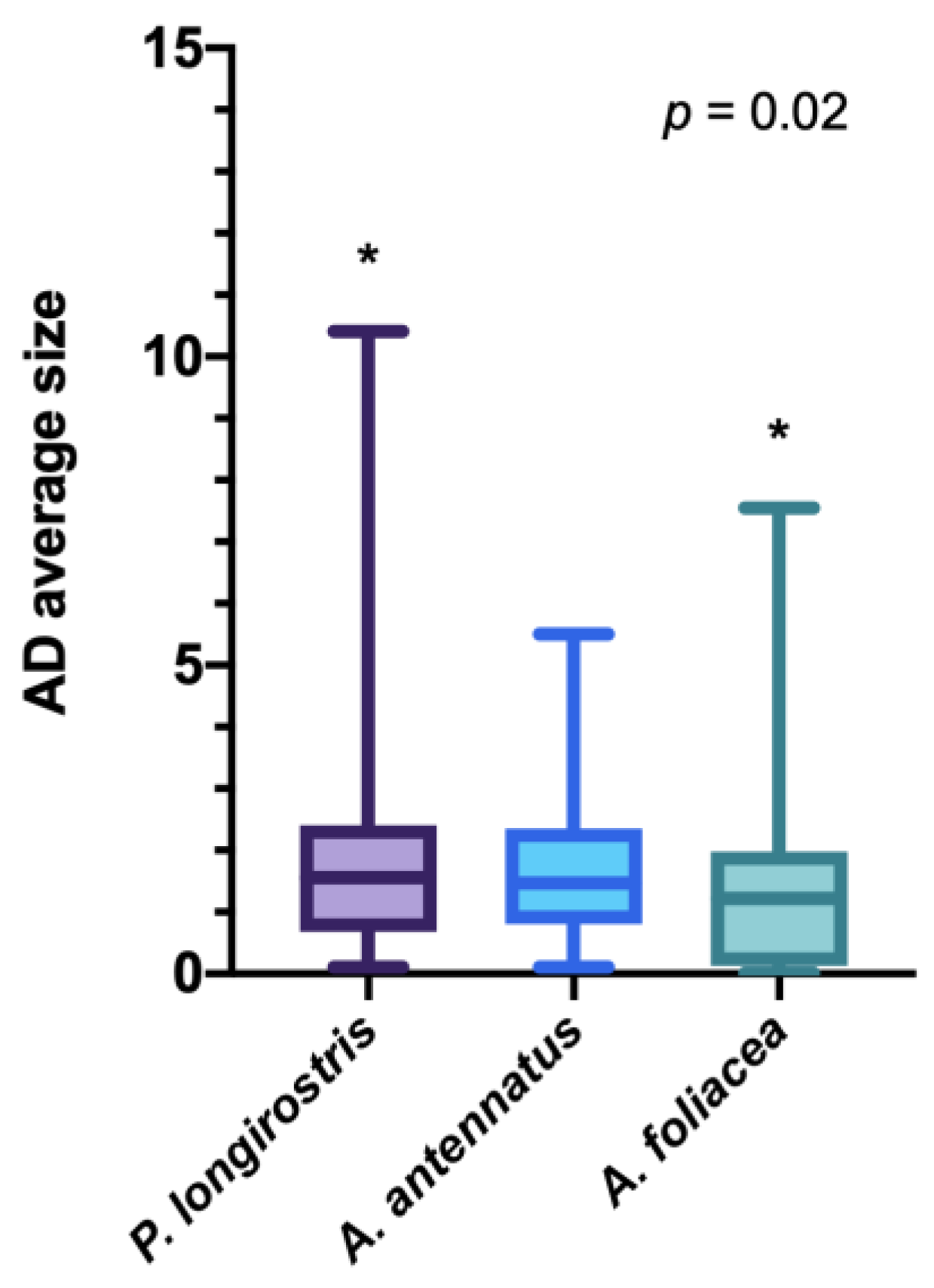
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hardesty, B.D.; Wilcox, C. A risk framework for tackling marine debris. Anal. Methods 2017, 9, 1429–1436. [Google Scholar] [CrossRef]
- Hardesty, B.D.; Wilcox, C.A. Presence of microplastics and nanoplastics in food, with particular focus on seafood. EFSA J. 2016, 14, e04501. [Google Scholar] [CrossRef] [Green Version]
- Jeftic, L.; Sheavly, S.; Adler, E. Marine Litter: A Global Challenge Marine Litter: A Global Challenge; UNEP: Nairobi, Kenya, 2009; ISBN 9789280730296. [Google Scholar]
- Jambeck, J.R.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A.; Narayan, R.; Law, K.L. Plastic waste inputs from land into the ocean. Science 2015, 347, 768–771. [Google Scholar] [CrossRef] [PubMed]
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [Green Version]
- Andrady, A.L. Microplastics in the marine environment. Mar. Pollut. Bull. 2011, 62, 1596–1605. [Google Scholar] [CrossRef]
- Consoli, P.; Falautano, M.; Sinopoli, M.; Perzia, P.; Canese, S.; Esposito, V.; Battaglia, P.; Romeo, T.; Andaloro, F.; Galgani, F.; et al. Composition and abundance of benthic marine litter in a coastal area of the central Mediterranean Sea. Mar. Pollut. Bull. 2018, 136, 243–247. [Google Scholar] [CrossRef]
- Ryan, P.G.; Moore, C.J.; Van Franeker, J.A.; Moloney, C.L. Monitoring the abundance of plastic debris in the marine environment. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1999–2012. [Google Scholar] [CrossRef] [Green Version]
- Kane, I.A.; Clare, M.A.; Miramontes, E.; Wogelius, R.; Rothwell, J.J.; Garreau, P.; Pohl, F. Seafloor microplastic hotspots controlle by deep-sea circulation. Science 2020, 368, 1140–1145. [Google Scholar] [CrossRef]
- Ballent, A.; Purser, A.; de Jesus Mendes, P.; Pando, S.; Thomsen, L. Physical transport properties of marine microplastic pollution. Biogeosci. Discuss. 2012, 9, 18755–18798. [Google Scholar] [CrossRef] [Green Version]
- Ballent, A.; Pando, S.; Purser, A.; Juliano, M.F.; Thomsen, L. Modelled transport of benthic marine microplastic pollution in the Nazaré Canyon. Biogeosciences 2013, 10, 7957–7970. [Google Scholar] [CrossRef]
- Kowalski, N.; Reichardt, A.M.; Waniek, J.J. Sinking rates of microplastics and potential implications of their alteration by physical, biological, and chemical factors. Mar. Pollut. Bull. 2016, 109, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Cózar, A.; Sanz-Martín, M.; Martí, E.; González-Gordillo, J.I.; Ubeda, B.; Gálvez, J.Á.; Irigoien, X.; Duarte, C.M. Plastic accumulation in the mediterranean sea. PLoS ONE 2015, 10, e0121762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, X.; Chen, M.; Chen, S.; Dasgupta, S.; Xu, H.; Ta, K.; Du, M.; Li, J.; Guo, Z.; Bai, S. Microplastics contaminate the deepest part of the world’s ocean. Geochem. Perspect. Lett. 2018, 9, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Kako, S.; Isobe, A.; Kataoka, T.; Hinata, H. A decadal prediction of the quantity of plastic marine debris littered on beaches of the East Asian marginal seas. Mar. Pollut. Bull. 2014, 81, 174–184. [Google Scholar] [CrossRef]
- Liubartseva, S.; Coppini, G.; Lecci, R.; Creti, S. Regional approach to modeling the transport of floating plastic debris in the Adriatic Sea. Mar. Pollut. Bull. 2016, 103, 115–127. [Google Scholar] [CrossRef]
- Cózar, A.; Echevarría, F.; González-Gordillo, J.I.; Irigoien, X.; Úbeda, B.; Hernández-León, S.; Palma, Á.T.; Navarro, S.; García-de-Lomas, J.; Ruiz, A.; et al. Plastic debris in the open ocean. Proc. Natl. Acad. Sci. USA 2014, 111, 10239–10244. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, M.; Lebreton, L.C.M.; Carson, H.S.; Thiel, M.; Moore, C.J.; Borerro, J.C.; Galgani, F.; Ryan, P.G.; Reisser, J. Plastic Pollution in the World’s Oceans: More than 5 Trillion Plastic Pieces Weighing over 250,000 Tons Afloat at Sea. PLoS ONE 2014, 9, e0111913. [Google Scholar] [CrossRef] [Green Version]
- Woodall, L.C.; Robinson, L.F.; Rogers, A.D.; Narayanaswamy, B.E.; Paterson, G.L.J. Deep-sea litter: A comparison of seamounts, banks and a ridge in the Atlantic and Indian Oceans reveals both environmental and anthropogenic factors impact accumulation and composition. Front. Mar. Sci. 2015, 2, 3. [Google Scholar] [CrossRef]
- Chiba, S.; Saito, H.; Fletcher, R.; Yogi, T.; Kayo, M.; Miyagi, S.; Ogido, M.; Fujikura, K. Human footprint in the abyss: 30 year records of deep-sea plastic debris. Mar. Policy 2018, 96, 204–212. [Google Scholar] [CrossRef]
- Choy, C.A.; Robison, B.H.; Gagne, T.O.; Erwin, B.; Firl, E.; Halden, R.U.; Hamilton, J.A.; Katija, K.; Lisin, S.E.; Rolsky, C.; et al. The vertical distribution and biological transport of marine microplastics across the epipelagic and mesopelagic water column. Sci. Rep. 2019, 9, 7843. [Google Scholar] [CrossRef]
- Lusher, A.L.; Welden, N.A.; Sobral, P.; Cole, M. Sampling, isolating and identifying microplastics ingested by fish and invertebrates. Anal. Methods 2017, 9, 1346–1360. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, N.B.; Hüffer, T.; Thompson, R.C.; Hassellöv, M.; Verschoor, A.; Daugaard, A.E.; Rist, S.; Karlsson, T.; Brennholt, N.; Cole, M.; et al. Are We Speaking the Same Language? Recommendations for a Definition and Categorization Framework for Plastic Debris. Environ. Sci. Technol. 2019, 53, 1039–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imhof, H.K.; Sigl, R.; Brauer, E.; Feyl, S.; Giesemann, P.; Klink, S.; Leupolz, K.; Löder, M.G.J.; Löschel, L.A.; Missun, J.; et al. Spatial and temporal variation of macro-, meso- and microplastic abundance on a remote coral island of the Maldives, Indian Ocean. Mar. Pollut. Bull. 2017, 116, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Alomar, C.; Deudero, S. Evidence of microplastic ingestion in the shark Galeus melastomus Rafinesque, 1810 in the continental shelf off the western Mediterranean Sea. Environ. Pollut. 2017, 223, 223–229. [Google Scholar] [CrossRef]
- Albano, M.; Panarello, G.; Di Paola, D.; D’Angelo, G.; Granata, A.; Savoca, S.; Capillo, G. The mauve stinger Pelagia noctiluca (Cnidaria, Scyphozoa) plastics contamination, the Strait of Messina case. Int. J. Environ. Stud. 2021, 78, 977–982. [Google Scholar] [CrossRef]
- D’Iglio, C.; Savoca, S.; Rinelli, P.; Spanò, N. Diet of the Deep-Sea Shark Galeus melastomus Rafinesque, 1810, in the Mediterranean Sea: What We Know and What We Should Know. Sustainability 2021, 13, 3962. [Google Scholar] [CrossRef]
- Ladewig, S.M.; Bao, S.; Chow, A.T. Natural Fibers: A Missing Link to Chemical Pollution Dispersion in Aquatic Environments. Environ. Sci. Technol. 2015, 49, 12609–12610. [Google Scholar] [CrossRef]
- Li, L.; Frey, M.; Browning, K.J. Biodegradability study on cotton and polyester fabrics. J. Eng. Fibers Fabr. 2010, 5, 42–53. [Google Scholar] [CrossRef]
- García-Rivera, S.; Lizaso, J.L.S.; Millán, J.M.B. Spatial and temporal trends of marine litter in the Spanish Mediterranean seafloor. Mar. Pollut. Bull. 2018, 137, 252–261. [Google Scholar] [CrossRef]
- UNEP/MAP. Updated Report on Marine Litter Assessment in the Mediterranean; UNEP(DEPI)/MED WG.421/Inf.18; European Environment Agency: Athens, Greece, 2015. [Google Scholar]
- Suaria, G.; Aliani, S. Floating debris in the Mediterranean Sea. Mar. Pollut. Bull. 2014, 86, 494–504. [Google Scholar] [CrossRef]
- Suaria, G.; Avio, C.G.; Mineo, A.; Lattin, G.L.; Magaldi, M.G.; Belmonte, G.; Moore, C.J.; Regoli, F.; Aliani, S. The Mediterranean Plastic Soup: Synthetic polymers in Mediterranean surface waters. Sci. Rep. 2016, 6, 37551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalberg Advisors, W.M.M. Initiative Stop the Flood of Plastic: How Mediterranean Countries Can Save Their Sea. A Guide for Policy Makers in Morocco; WWF: Gland, Switzerland, 2019. [Google Scholar]
- Costa, R.; Albergamo, A.; Piparo, M.; Zaccone, G.; Capillo, G.; Manganaro, A.; Dugo, P.; Mondello, L. Multidimensional gas chromatographic techniques applied to the analysis of lipids from wild-caught and farmed marine species. Eur. J. Lipid Sci. Technol. 2017, 119, 1600043. [Google Scholar] [CrossRef]
- Alimba, C.G.; Faggio, C. Microplastics in the marine environment: Current trends in environmental pollution and mechanisms of toxicological profile. Environ. Toxicol. Pharmacol. 2019, 68, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Prevenios, M.; Zeri, C.; Tsangaris, C.; Liubartseva, S.; Fakiris, E.; Papatheodorou, G. Beach litter dynamics on Mediterranean coasts: Distinguishing sources and pathways. Mar. Pollut. Bull. 2018, 129, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Politikos, D.V.; Tsiaras, K.; Papatheodorou, G.; Anastasopoulou, A. Modeling of floating marine litter originated from the Eastern Ionian Sea: Transport, residence time and connectivity. Mar. Pollut. Bull. 2020, 150, 110727. [Google Scholar] [CrossRef]
- Vlachogianni, T.; Fortibuoni, T.; Ronchi, F.; Zeri, C.; Mazziotti, C.; Tutman, P.; Varezić, D.B.; Palatinus, A.; Trdan, Š.; Peterlin, M.; et al. Marine litter on the beaches of the Adriatic and Ionian Seas: An assessment of their abundance, composition and sources. Mar. Pollut. Bull. 2018, 131, 745–756. [Google Scholar] [CrossRef]
- Galgani, F.; Leaute, J.P.; Moguedet, P.; Souplet, A.; Verin, Y.; Carpentier, A.; Goraguer, H.; Latrouite, D.; Andral, B.; Cadiou, Y.; et al. Litter on the sea floor along European coasts. Mar. Pollut. Bull. 2000, 40, 516–527. [Google Scholar] [CrossRef]
- Angiolillo, M.; di Lorenzo, B.; Farcomeni, A.; Bo, M.; Bavestrello, G.; Santangelo, G.; Cau, A.; Mastascusa, V.; Cau, A.; Sacco, F.; et al. Distribution and assessment of marine debris in the deep Tyrrhenian Sea (NW Mediterranean Sea, Italy). Mar. Pollut. Bull. 2015, 92, 149–159. [Google Scholar] [CrossRef]
- Tubau, X.; Canals, M.; Lastras, G.; Rayo, X.; Rivera, J.; Amblas, D. Marine litter on the floor of deep submarine canyons of the Northwestern Mediterranean Sea: The role of hydrodynamic processes. Prog. Oceanogr. 2015, 134, 379–403. [Google Scholar] [CrossRef]
- Cau, A.; Alvito, A.; Moccia, D.; Canese, S.; Pusceddu, A.; Rita, C.; Angiolillo, M.; Follesa, M.C. Submarine canyons along the upper Sardinian slope (Central Western Mediterranean) as repositories for derelict fishing gears. Mar. Pollut. Bull. 2017, 123, 357–364. [Google Scholar] [CrossRef]
- García-Rivera, S.; Lizaso, J.L.S.; Millán, J.M.B. Composition, spatial distribution and sources of macro-marine litter on the Gulf of Alicante seafloor (Spanish Mediterranean). Mar. Pollut. Bull. 2017, 121, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Pierdomenico, M.; Casalbore, D.; Chiocci, F.L. The key role of canyons in funnelling litter to the deep sea: A study of the Gioia Canyon (Southern Tyrrhenian Sea). Anthropocene 2020, 30, 100237. [Google Scholar] [CrossRef]
- Cau, A.; Avio, C.G.; Dessì, C.; Follesa, M.C.; Moccia, D.; Regoli, F.; Pusceddu, A. Microplastics in the crustaceans Nephrops norvegicus and Aristeus antennatus: Flagship species for deep-sea environments? Environ. Pollut. 2019, 255, 113107. [Google Scholar] [CrossRef] [PubMed]
- Cau, A.; Avio, C.G.; Dessì, C.; Moccia, D.; Pusceddu, A.; Regoli, F.; Cannas, R.; Follesa, M.C. Benthic Crustacean Digestion Can Modulate the Environmental Fate of Microplastics in the Deep Sea. Environ. Sci. Technol. 2020, 54, 4886–4892. [Google Scholar] [CrossRef] [PubMed]
- STECF. Mediterranean Assessments Part 1 (STECF-15-18); JRC 98676; EUR 27638; Publications Office of the European Union: Luxembourg, 2015; Volume 1, ISBN 978-92-79-54141-4. [Google Scholar]
- STECF. Scientific, Technical and Economic Committee for Fisheries (STECF)—Stock Assessments: Demersal Stocks in the Western Mediterranean Sea (STECF-20-09); EUR 28359; Publications Office of the European Union: Luxembourg, 2020; ISBN 978-92-76-11288-4. [Google Scholar]
- Carvalho, N.; Doerner, H. Scientific, Technical and Economic Committee for Fisheries (STECF) Balance Indicators; Publications Office of the European Union: Luxembourg, 2014. [Google Scholar] [CrossRef]
- Perdichizzi, A.; D’Iglio, C.; Giordano, D.; Profeta, A.; Ragonese, S.; Rinelli, P. Comparing life-history traits in two contiguous stocks of the deep-water rose shrimp Parapenaeus longirostris (H. Lucas, 1846) (Crustacea: Decapoda) in the Southern Tyrrhenian Sea (Central Mediterranean Sea). Fish. Res. 2022, 248, 106206. [Google Scholar] [CrossRef]
- Cartes, J.E.; Company, J.B.; Maynou, F. Deep-water decapod crustacean communities in the Northwestern Mediterranean: Influence of submarine canyons and season. Mar. Biol. 1994, 120, 221–229. [Google Scholar] [CrossRef]
- García, T.; Sobrino, I. Biology and fishery of the deepwater Rose shrimp, Parapenaeus longirostris (Lucas, 1846), from the Atlantic Moroccan coast. Sci. Mar. 1994, 58, 299–305. [Google Scholar]
- Modica, L.; Cartes, J.E.; Velasco, F.; Bozzano, A. Juvenile hake predation on Myctophidae and Sternoptychidae: Quantifying an energy transfer between mesopelagic and neritic communities. J. Sea Res. 2015, 95, 217–225. [Google Scholar] [CrossRef]
- Cartes, J.E. Diets of, and trophic resources exploited by, bathyal penaeoidean shrimps from the western Mediterranean. Mar. Freshw. Res. 1995, 46, 889–896. [Google Scholar] [CrossRef]
- D’Iglio, C.; Porcino, N.; Savoca, S.; Profeta, A.; Perdichizzi, A.; Armeli, E.; Davide, M.; Francesco, S.; Rinelli, P.; Giordano, D. Ontogenetic shift and feeding habits of the European hake (Merluccius merluccius L., 1758) in Central and Southern Tyrrhenian Sea (Western Mediterranean Sea): A comparison between past and present data. Ecol. Evol. 2022, 12, e8634. [Google Scholar] [CrossRef]
- D’Iglio, C.; Albano, M.; Tiralongo, F.; Famulari, S.; Rinelli, P.; Savoca, S.; Spanò, N.; Capillo, G. Biological and Ecological Aspects of the Blackmouth Catshark (Galeus melastomus Rafinesque, 1810) in the Southern Tyrrhenian Sea. J. Mar. Sci. Eng. 2021, 9, 967. [Google Scholar] [CrossRef]
- D’Iglio, C.; Famulari, S.; Albano, M.; Giordano, D.; Rinelli, P.; Capillo, G.; Spanò, N.; Savoca, S. Time-Scale Analysis of Prey Preferences and Ontogenetic Shift in the Diet of European Hake Merluccius merluccius (Linnaeus, 1758) in Southern and Central Tyrrhenian Sea. Fishes 2022, 7, 167. [Google Scholar] [CrossRef]
- Carreras-Colom, E.; Constenla, M.; Soler-Membrives, A.; Cartes, J.E.; Baeza, M.; Carrassón, M. A closer look at anthropogenic fiber ingestion in Aristeus antennatus in the NW Mediterranean Sea: Differences among years and locations and impact on health condition. Environ. Pollut. 2020, 263, 114567. [Google Scholar] [CrossRef] [PubMed]
- Bono, G.; Falsone, F.; Falco, F.; Di Maio, F.; Gabriele, M.; Gancitano, V.; Geraci, M.L.; Mancuso, M.; Okpala, C.; Luisa, P.; et al. Microplastics and Alien Black Particles as Contaminants of Deep-Water Rose Shrimp (Parapenaeus longistroris Lucas, 1846) in the Central Mediterranean Sea. J. Adv. Biotechnol. Bioeng. 2020, 8, 23–28. [Google Scholar] [CrossRef]
- Carreras-Colom, E. Unravelling the (Micro)Plastic Threat: The Case Study of Plastic Ingestion in Aristeus antennatus and Nephrops norvegicus from the NW Mediterranean Sea and Its Potential Impact on Health Condition. Ph.D. Thesis, Universitat Autònoma de Barcelona, Barcelona, Spain, 2021. [Google Scholar]
- Kapiris, K.; Thessalou-Legaki, M.; Petrakis, G.; Conides, A. Ontogenetic shifts and temporal changes in the trophic patterns of the deep-sea red shrimp, Aristaeomorpha foliacea (Decapods: Aristeidae), in the Eastern Ionian Sea (Eastern Mediterranean). Mar. Ecol. 2010, 31, 341–354. [Google Scholar] [CrossRef]
- Mangano, M.C.; Kaiser, M.J.; Porporato, E.M.D.; Lambert, G.I.; Spanò, N. Trawling disturbance effects on the trophic ecology of two co-generic Astropectinid species. Mediterr. Mar. Sci. 2015, 16, 538–549. [Google Scholar] [CrossRef]
- Porporato, E.M.D.; Lo Giudice, A.; Michaud, L.; de Domenico, E.; Spanò, N. Diversity and Antibacterial Activity of the Bacterial Communities Associated with Two Mediterranean Sea Pens, Pennatula phosphorea and Pteroeides spinosum (Anthozoa: Octocorallia). Microb. Ecol. 2013, 66, 701–714. [Google Scholar] [CrossRef]
- Follesa, M.C.; Carbonara, P. Atlas of the Maturity Stages of Mediterranean Fishery Resources. Studies and Reviews n. 99; FAO: Rome, Italy, 2019; ISBN 9789251319758. [Google Scholar]
- Savoca, S.; Matanović, K.; D’Angelo, G.; Vetri, V.; Anselmo, S.; Bottari, T.; Mancuso, M.; Kužir, S.; Spanò, N.; Capillo, G.; et al. Ingestion of plastic and non-plastic microfibers by farmed gilthead sea bream (Sparus aurata) and common carp (Cyprinus carpio) at different life stages. Sci. Total Environ. 2021, 782, 146851. [Google Scholar] [CrossRef]
- Besley, A.; Vijver, M.G.; Behrens, P.; Bosker, T. A standardized method for sampling and extraction methods for quantifying microplastics in beach sand. Mar. Pollut. Bull. 2017, 114, 77–83. [Google Scholar] [CrossRef]
- Silva, A.B.; Bastos, A.S.; Justino, C.I.L.; da Costa, J.P.; Duarte, A.C.; Rocha-Santos, T.A.P. Microplastics in the environment: Challenges in analytical chemistry—A review. Anal. Chim. Acta 2018, 1017, 1–19. [Google Scholar] [CrossRef]
- Giani, D.; Baini, M.; Galli, M.; Casini, S.; Fossi, M.C. Microplastics occurrence in edible fish species (Mullus barbatus and Merluccius merluccius) collected in three different geographical sub-areas of the Mediterranean Sea. Mar. Pollut. Bull. 2019, 140, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Li, J.Y.; Craig, N.J.; Su, L. Microplastic pollution in wild populations of decapod crustaceans: A review. Chemosphere 2022, 291, 132985. [Google Scholar] [CrossRef] [PubMed]
- Collignon, A.; Hecq, J.H.; Galgani, F.; Collard, F.; Goffart, A. Annual variation in neustonic micro- and meso-plastic particles and zooplankton in the Bay of Calvi (Mediterranean-Corsica). Mar. Pollut. Bull. 2014, 79, 293–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fastelli, P.; Blašković, A.; Bernardi, G.; Romeo, T.; Čižmek, H.; Andaloro, F.; Russo, G.F.; Guerranti, C.; Renzi, M. Plastic litter in sediments from a marine area likely to become protected (Aeolian Archipelago’s islands, Tyrrhenian sea). Mar. Pollut. Bull. 2016, 113, 526–529. [Google Scholar] [CrossRef]
- Cau, A.; Bellodi, A.; Moccia, D.; Mulas, A.; Pesci, P.; Cannas, R.; Pusceddu, A.; Follesa, M.C. Dumping to the abyss: Single-use marine litter invading bathyal plains of the Sardinian margin (Tyrrhenian Sea). Mar. Pollut. Bull. 2018, 135, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Mistri, M.; Scoponi, M.; Granata, T.; Moruzzi, L.; Massara, F.; Munari, C. Types, occurrence and distribution of microplastics in sediments from the northern Tyrrhenian Sea. Mar. Pollut. Bull. 2020, 153, 111016. [Google Scholar] [CrossRef]
- Rossi, S.; Gabbianelli, G. Geomorfologia del Golfo di Taranto. Bull. Soc Geol. 1978, 97, 423–437. [Google Scholar]
- Cataudella, S.; Spagnolo, M. The State of Italian Marine Fisheries and Aquaculture; MiPAAF: Rome, Italy, 2011. [Google Scholar]
- Theocharis, A.; Georgopoulos, D.; Lascaratos, A.; Nittis, K. Water masses and circulation in the central region of the Eastern Mediterranean: Eastern Ionian, South Aegean and Northwest Levantine, 1986–1987. Deep Sea Res. Part II 1993, 40, 1121–1142. [Google Scholar] [CrossRef]
- Canals, M.; Danovaro, R.; Heussner, S.; Lykousis, V.; Puig, P.; Trincardi, F.; Calafat, A.M.; de Madron, X.D.; Palanques, A.; Sànchez-Vidal, A. Cascades in mediterranean submarine grand canyons. Oceanography 2009, 22, 26–43. [Google Scholar] [CrossRef] [Green Version]
- Klein, B.; Roether, W.; Manca, B.B.; Bregant, D.; Beitzel, V.; Kovacevic, V.; Luchetta, A. The large deep water transient in the Eastern Mediterranean. Deep Sea Res. Part I Oceanogr. Res. Pap. 1999, 46, 371–414. [Google Scholar] [CrossRef]
- Manca, B.B.; Ursella, L.; Scarazzato, P. New development of eastern mediterranean circulation based on hydrological observations and current measurements. Mar. Ecol. 2002, 23, 237–257. [Google Scholar] [CrossRef]
- Watts, A.J.R.; Urbina, M.A.; Goodhead, R.; Moger, J.; Lewis, C.; Galloway, T.S. Effect of Microplastic on the Gills of the Shore Crab Carcinus maenas. Environ. Sci. Technol. 2016, 50, 5364–5369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Rath, C.C.; Das, A.P. Marine microfiber pollution: A review on present status and future challenges. Mar. Pollut. Bull. 2019, 140, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Desforges, J.P.W.; Galbraith, M.; Ross, P.S. Ingestion of Microplastics by Zooplankton in the Northeast Pacific Ocean. Arch. Environ. Contam. Toxicol. 2015, 69, 320–330. [Google Scholar] [CrossRef]
- Cartes, J.E.; Fanelli, E.; Kapiris, K.; Bayhan, Y.K.; Ligas, A.; López-Pérez, C.; Murenu, M.; Papiol, V.; Rumolo, P.; Scarcella, G. Spatial variability in the trophic ecology and biology of the deep-sea shrimp Aristaeomorpha foliacea in the Mediterranean Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 2014, 87, 1–13. [Google Scholar] [CrossRef]
- Kapiris, K. Feeding ecology of Parapenaeus longirostris (Lucas, 1846) (Decapoda: Penaeidae) from the Ionian Sea (Central and Eastern Mediterranean Sea). Sci. Mar. 2004, 68, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Sbrana, M.; Viva, C.; Belcari, P. Fishery of the deep-water rose shrimp Parapenaeus longirostris (Lucas, 1846) (Crustacea: Decapoda) in the northern Tyrrhenian Sea (western Mediterranean). Hydrobiologia 2006, 557, 135–144. [Google Scholar] [CrossRef]
- Burukovskii, R.N. On the bathymetric distribution and feeding of the shrimp Parapenaeus longirostris (Lucas). Ices C. M. 1969, 1–7. [Google Scholar]
- Lagardère, J.P. Recherches sur la distribution et sur l’alimentation des crustacés décapodes benthiques de la pente continentale du Golfe de Gascogne. Analyse des groupements carcinologiques. Bull. Cent. Étud. Rech. Sci. Biarritz 1977, 11, 367–440. [Google Scholar]
- Cartes, J.E. Influence of depth and season on the diet of the deep-water aristeid Aristeus antennatus along the continental slope (400 to 2300 m) in the Catalan Sea (western Mediterranean). Mar. Biol. 1994, 120, 639–648. [Google Scholar] [CrossRef]
- Cartes, J.; Sarda, F. Feeding ecology of the deep-water aristeid crustacean Aristeus antennatus. Mar. Ecol. Prog. Ser. 1989, 54, 229–238. [Google Scholar] [CrossRef]
- Rainer, S.F. Diet of prawns from the continental slope of north-western Australia. Bull. Mar. Sci. 1992, 50, 258–274. [Google Scholar]
- Lagardere, J.P. Recherches sur l’ alimentation des crevettes de la pente continentale marocaine. Tethys 1972, 3, 655–675. [Google Scholar]
- Carreras-Colom, E.; Constenla, M.; Soler-Membrives, A.; Cartes, J.E.; Baeza, M.; Padrós, F.; Carrassón, M. Spatial occurrence and effects of microplastic ingestion on the deep-water shrimp Aristeus antennatus. Mar. Pollut. Bull. 2018, 133, 44–52. [Google Scholar] [CrossRef] [PubMed]
- D’Costa, A.H. Microplastics in decapod crustaceans: Accumulation, toxicity and impacts, a review. Sci. Total Environ. 2022, 832, 154963. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Length (mm) | Weight (g) | N° of Specimens | Np | Items/Specimen |
|---|---|---|---|---|---|
| Means ± SD | Means ± SD | ||||
| Parapenaues longirostris | 23.3 ± 1.6 | 8.1 ± 1.5 | 50 | 37 | 2.24 |
| Aristeus antennatus | 45.8 ± 5 | 31.5 ± 8 | 50 | 35 | 2.22 |
| Aristaeomorpha foliacea | 38.4 ± 6.7 | 19.1 ± 11 | 36 | 30 | 2.30 |
| Species | Sample | Length (mm) | Weight (g) | Sex | Maturity | N° AD |
|---|---|---|---|---|---|---|
| Parapenaues longirostris | 8 | 27.20 | 7.60 | M | 2E | 0 |
| 10 | 27.60 | 9.10 | M | 2B | 0 | |
| 14 | 26.40 | 9.90 | M | 2B | 0 | |
| 16 | 29.00 | 8.40 | M | 2E | 0 | |
| 19 | 26.00 | 6.70 | M | 2E | 0 | |
| 26 | 27.20 | 9.70 | F | 2B | 0 | |
| 32 | 26.20 | 7.40 | F | 2B | 0 | |
| 34 | 25.50 | 7.80 | F | 2B | 0 | |
| 35 | 25.00 | 7.40 | F | 2B | 0 | |
| 36 | 26.00 | 4.70 | F | 2B | 0 | |
| 38 | 26.80 | 8.70 | F | 2B | 0 | |
| 41 | 27.10 | 9.20 | F | 2B | 0 | |
| 44 | 26.00 | 8.40 | F | 2B | 0 | |
| Aristeus antennatus | 51 | 49.60 | 36.50 | F | 2E | 0 |
| 53 | 51.50 | 41.70 | F | 2E | 0 | |
| 56 | 50.30 | 40.80 | F | 2E | 0 | |
| 60 | 37.50 | 19.00 | F | 2E | 0 | |
| 61 | 42.50 | 25.10 | F | 2E | 0 | |
| 76 | 50.80 | 41.50 | F | 2E | 0 | |
| 77 | 49.30 | 38.80 | F | 2E | 0 | |
| 78 | 41.90 | 23.70 | F | 2B | 0 | |
| 80 | 43.00 | 30.70 | F | 2B | 0 | |
| 86 | 50.50 | 34.30 | F | 2B | 0 | |
| 87 | 55.50 | 45.90 | F | 2B | 0 | |
| 89 | 48.10 | 33.60 | F | 2B | 0 | |
| 93 | 56.00 | 45.90 | F | 2B | 0 | |
| 94 | 52.00 | 41.60 | F | 2B | 0 | |
| 98 | 46.80 | 37.40 | F | 2B | 0 | |
| Aristaeomorpha foliacea | 105 | 36.00 | 17.11 | F | 2E | 0 |
| 107 | 40.0 | 19.64 | F | 2E | 0 | |
| 109 | 34.80 | 11.10 | F | 2E | 0 | |
| 122 | 37.50 | 18.99 | F | 2E | 0 | |
| 125 | 52.00 | 42.23 | F | 2E | 0 | |
| 132 | 31.00 | 9.00 | F | 2E | 0 |
| Size Classes | Size Range | P. longirostris | A. antennatus | A. foliacea |
|---|---|---|---|---|
| I | 0.10–0.49 | 12 | 11 | 24 |
| II | 0.50–0.99 | 15 | 16 | 8 |
| III | 1.00–1.49 | 11 | 14 | 10 |
| IV | 1.50–1.99 | 13 | 12 | 10 |
| V | 2.00–2.49 | 12 | 7 | 4 |
| VI | 2.50–2.99 | 7 | 7 | 5 |
| VII | 3.00–3.49 | 4 | 3 | 4 |
| VIII | 3.50–3.99 | 1 | 4 | 2 |
| IX | 4.00–4.99 | 1 | 3 | 1 |
| X | ≥5.00 | 7 | 1 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Iglio, C.; Di Fresco, D.; Spanò, N.; Albano, M.; Panarello, G.; Laface, F.; Faggio, C.; Capillo, G.; Savoca, S. Occurrence of Anthropogenic Debris in Three Commercial Shrimp Species from South-Western Ionian Sea. Biology 2022, 11, 1616. https://doi.org/10.3390/biology11111616
D’Iglio C, Di Fresco D, Spanò N, Albano M, Panarello G, Laface F, Faggio C, Capillo G, Savoca S. Occurrence of Anthropogenic Debris in Three Commercial Shrimp Species from South-Western Ionian Sea. Biology. 2022; 11(11):1616. https://doi.org/10.3390/biology11111616
Chicago/Turabian StyleD’Iglio, Claudio, Dario Di Fresco, Nunziacarla Spanò, Marco Albano, Giuseppe Panarello, Federica Laface, Caterina Faggio, Gioele Capillo, and Serena Savoca. 2022. "Occurrence of Anthropogenic Debris in Three Commercial Shrimp Species from South-Western Ionian Sea" Biology 11, no. 11: 1616. https://doi.org/10.3390/biology11111616