
Effect of Diet on the Midgut Microbial Composition and Host Immunity of the Fall Armyworm, Spodoptera frugiperda
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collections
2.2. Midgut DNA Extraction
2.3. Bioinformatics Analysis
2.4. Antibiotic Treatment and Removal of the Gut Bacterial Communities
2.5. Quantitative Real-Time PCR Analysis of the Immune Genes
2.6. Isolation of Gut Microbiota from S. frugiperda
2.7. Removal of Gut Microbiota from S. frugiperda Using Antibiotic Treatment
2.8. Statistical Analysis
3. Results
3.1. Illumina Sequencing Statistics
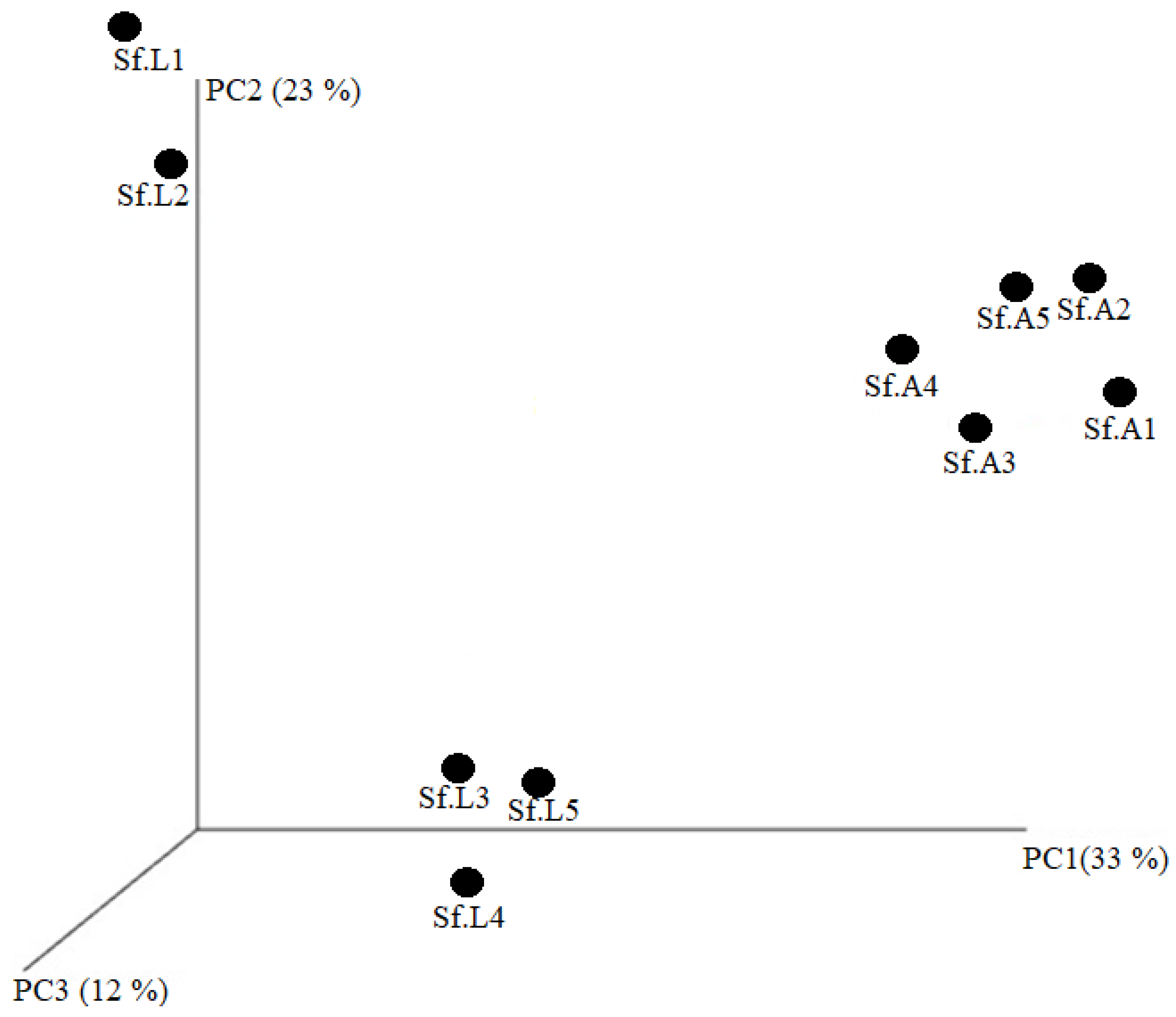
3.2. Diversity Analysis of Midgut Microbiota in “Lab Diet” and “Field Diet” Groups
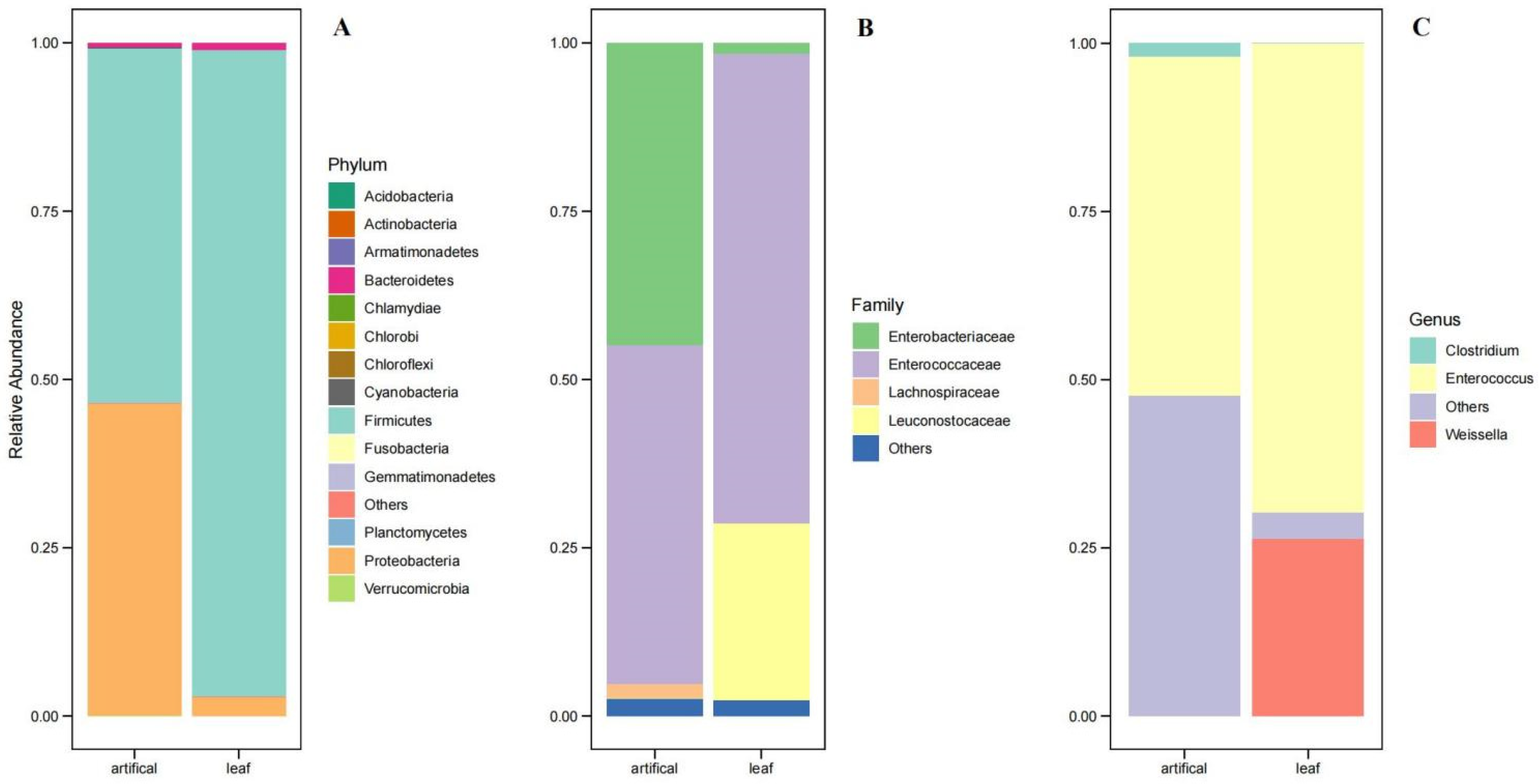
3.3. Bacterial Community of S. frugiperda Reared in “Lab Diet” and “Field Diet”
Comparison of the Gut Microbiota of S. frugiperda Reared in “Lab Diet” and “Field Diet”
3.4. Predictions of Metabolic Potentials
3.5. Effect of the Gut Microbiome on the Host Immune Gene Expression
3.6. Isolation of Midgut Microbiota from S. frugiperda
3.7. Effect of Gut Microbiota on Autographa Californica Nucleopolyhedrovirus (AcMNPV) Pathogenesis in S. frugiperda Larvae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PICRUSt | Phylogenetic Investigation of Communities by Reconstruction of Unobserved States |
| HTS | High throughput sequencing |
| OTUs | Operational taxonomic units |
| ACE | Abundance-based coverage estimator |
References
- Douglas, A.E. Multiorganismal insects: Diversity and function of resident microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yek, S.H.; Mueller, U.G. The metapleural gland of ants. Biol. Rev. 2011, 86, 774–791. [Google Scholar] [CrossRef] [PubMed]
- Ruokolainen, L.; Ikonen, S.; Makkonen, H.; Hanski, I. Larval growth rate is associated with the composition of the gut microbiota in the Glanville fritillary butterfly. Oecologia 2016, 181, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Leitão-Gonçalves, R.; Carvalho-Santos, Z.; Francisco, A.P.; Fioreze, G.T.; Anjos, M.; Baltazar, C.; Elias, A.P.; Itskov, P.M.; Piper, M.D.W.; Ribeiro, C. Commensal bacteria and essential amino acids control food choice behavior and reproduction. PLoS Biol. 2017, 15, e2000862. [Google Scholar] [CrossRef] [Green Version]
- Ankrah, N.Y.D.; Douglas, A.E. Nutrient factories: Metabolic function of beneficial microorganisms associated with insects. Environ. Microbiol. 2018, 20, 2002–2011. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Chen, B.; Sun, C.; Ishida, K.; Hertweck, C.; Boland, W. Symbiont-derived antimicrobials contribute to the control of the Lepidopteran gut microbiota. Cell Chem. Biol. 2017, 24, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.G.; Mason, C.J.; Felton, G.W.; Hoover, K. Host plant and population source drive diversity of microbial gut communities in two polyphagous insects. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Broderick, N.A.; Raffa, K.F.; Handelsman, J. Midgut bacteria required for Bacillus thuringiensis insecticidal activity. Proc. Natl. Acad. Sci. USA 2006, 103, 15196–15199. [Google Scholar] [CrossRef] [Green Version]
- Pinto-Tomás, A.; Uribe-Lorío, L.; Blanco, J.; Fontecha, G.; Rodríguez, C.; Mora, M.; Janzen, D.; Chavarría, F.; Díaz, J.; Sittenfeld, A. Actividades enzimáticas en aislamientos bacterianos de tractos digestivos de larvas y del contenido de pupas de Automeris zugana y Rothschildia lebeau (Lepidoptera: Saturniidae). Rev. De Biol. Trop. 2007, 55, 401–415. [Google Scholar] [CrossRef] [Green Version]
- Prem Anand, A.A.; Vennison, S.J.; Sankar, S.G.; Gilwax Prabhu, D.I.; Vasan, P.T.; Raghuraman, T.; Jerome Geoffrey, C.; Vendan, S.E. Isolation and characterization of bacteria from the gut of Bombyx mori that degrade cellulose, xylan, pectin and starch and their impact on digestion. J. Insect Sci. 2010, 10, 107. [Google Scholar]
- Wang, J.; Peiffer, M.; Hoover, K.; Rosa, C.; Zeng, R.; Felton, G.W. Helicoverpa zea gut-associated bacteria indirectly induce defenses in tomato by triggering a salivary elicitor(s). New Phytol. 2017, 214, 1294–1306. [Google Scholar] [CrossRef] [PubMed]
- Hammer, T.J.; Janzen, D.H.; Hallwachs, W.; Jaffe, S.P.; Fierer, N. Caterpillars lack a resident gut microbiome. Proc. Natl. Acad. Sci. USA 2017, 114, 9641–9646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparks, A.N. A review of the biology of the fall armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.d.; Peterson, J.A.; Hunt, T. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Feldmann, F.; Rieckmann, U.; Winter, S. The spread of the fall armyworm Spodoptera frugiperda in Africa—What should be done next? J. Plant Dis. Prot. 2019, 126, 97–101. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (JE Smith)(Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [Green Version]
- Sharanabasappa; Kalleshwaraswamy, C.M.; Asokan, R.; Swamy, H.M.M.; Maruthi, M.S.; Pavithra, H.B.; Hegbe, K.; Navi, S.; Prabhu, S.T.; Goergen, G.E. First report of the fall armyworm, Spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest Manag. Hortic. Ecosyst. 2018, 24, 23–29. [Google Scholar]
- Guo, J.; Zhao, J.; He, K.; Zhang, F.; Wang, Z. Potential invasion of the crop-devastating insect pest fall armyworm Spodoptera frugiperda to China. Plant Prot. 2018, 44, 1–10. [Google Scholar]
- Sharanabasappa; Kalleshwaraswamy, C.M.; Maruthi, M.S.; Pavithra, H.B. Biology of invasive fall army worm Spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae) on maize. Indian J. Entomol. 2018, 80, 540–543. [Google Scholar]
- Sun, X.-x.; Hu, C.-x.; Jia, H.-r.; Wu, Q.-l.; Shen, X.-j.; Zhao, S.-y.; Jiang, Y.-y.; Wu, K.-m. Case study on the first immigration of fall armyworm Spodoptera frugiperda invading into China. J. Integr. Agric. 2020, 19, 664–672. [Google Scholar] [CrossRef]
- De Groote, H.; Kimenju, S.C.; Munyua, B.; Palmas, S.; Kassie, M.; Bruce, A. Spread and impact of fall armyworm (Spodoptera frugiperda JE Smith) in maize production areas of Kenya. Agric. Ecosyst. Environ. 2020, 292, 106804. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, A.C.; Staten, R.T. The Sterile Insect Release Method and Other Genetic Control Strategies. In Radcliffe’s IPM World Textbook; University of Minnesota: St. Paul, MN, USA, 1996; Available online: https://ipmworld.umn.edu/bartlett (accessed on 30 July 2022).
- Wright, R.J. Evaluation of crop rotation for control of Colorado potato beetles (Coleoptera: Chrysomelidae) in commercial potato fields on Long Island. J. Econ. Entomol. 1984, 77, 1254–1259. [Google Scholar] [CrossRef] [Green Version]
- Jakka, S.R.K.; Gong, L.; Hasler, J.; Banerjee, R.; Sheets, J.J.; Narva, K.; Blanco, C.A.; Jurat-Fuentes, J.L. Field-evolved mode 1 resistance of the fall armyworm to transgenic Cry1Fa-expressing corn associated with reduced Cry1Fa toxin binding and midgut alkaline phosphatase expression. Appl. Environ. Microbiol. 2016, 82, 1023–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, R.; Hasler, J.; Meagher, R.; Nagoshi, R.; Hietala, L.; Huang, F.; Narva, K.; Jurat-Fuentes, J.L. Mechanism and DNA-based detection of field-evolved resistance to transgenic Bt corn in fall armyworm (Spodoptera frugiperda). Sci. Rep. 2017, 7, 1–10. [Google Scholar]
- Arora, A.K.; Douglas, A.E. Hype or opportunity? Using microbial symbionts in novel strategies for insect pest control. J. Insect Physiol. 2017, 103, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Ceja-Navarro, J.A.; Karaoz, U.; Bill, M.; Hao, Z.; White, R.A.; Arellano, A.; Ramanculova, L.; Filley, T.R.; Berry, T.D.; Conrad, M.E.; et al. Gut anatomical properties and microbial functional assembly promote lignocellulose deconstruction and colony subsistence of a wood-feeding beetle. Nat. Microbiol. 2019, 4, 864. [Google Scholar] [CrossRef] [Green Version]
- Vilanova, C.; Baixeras, J.; Latorre, A.; Porcar, M. The generalist inside the specialist: Gut bacterial communities of two insect species feeding on toxic plants are dominated by Enterococcus sp. Front. Microbiol. 2016, 7, 1005. [Google Scholar] [CrossRef] [Green Version]
- Ceja-Navarro, J.A.; Vega, F.E.; Karaoz, U.; Hao, Z.; Jenkins, S.; Lim, H.C.; Kosina, P.; Infante, F.; Northen, T.R.; Brodie, E.L. Gut microbiota mediate caffeine detoxification in the primary insect pest of coffee. Nat. Commun. 2015, 6, 7618. [Google Scholar] [CrossRef] [Green Version]
- Fadrosh, D.W.; Ma, B.; Gajer, P.; Sengamalay, N.; Ott, S.; Brotman, R.M.; Ravel, J. An improved dual-indexing approach for multiplexed 16S rRNA gene sequencing on the Illumina MiSeq platform. Microbiome 2014, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Lv, D.; Liu, X.; Dong, Y.; Yan, Z.; Zhang, X.; Wang, P.; Yuan, X.; Li, Y. Comparison of Gut Bacterial Communities of Fall Armyworm (Spodoptera frugiperda) Reared on Different Host Plants. Int. J. Mol. Sci. 2021, 22, 11266. [Google Scholar] [CrossRef]
- Chen, Y.-P.; Li, Y.-H.; Sun, Z.-X.; Du, E.W.; Lu, Z.-H.; Li, H.; Gui, F.-R. Effects of Host Plants on Bacterial Community Structure in Larvae Midgut of Spodoptera frugiperda. Insects 2022, 13, 373. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, F.E.; Peiffer, M.; Tan, C.-W.; Stanley, B.A.; Stanley, A.; Wang, J.; Jones, A.G.; Hoover, K.; Rosa, C.; Luthe, D.; et al. Fall armyworm-associated gut bacteria modulate plant defense responses. Mol. Plant-Microbe Interact. 2017, 30, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Almeida, L.G.d.; Moraes, L.A.B.d.; Trigo, J.R.; Omoto, C.; Consoli, F.L. The gut microbiota of insecticide-resistant insects houses insecticide-degrading bacteria: A potential source for biotechnological exploitation. PLoS ONE 2017, 12, e0174754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozadilla, G.; Cabrera, N.A.; Virla, E.G.; Greco, N.M.; McCarthy, C.B. Gut microbiota of Spodoptera frugiperda (JE Smith) larvae as revealed by metatranscriptomic analysis. J. Appl. Entomol. 2020, 144, 351–363. [Google Scholar] [CrossRef]
- Gichuhi, J.; Sevgan, S.; Khamis, F.; Van den Berg, J.; du Plessis, H.; Ekesi, S.; Herren, J.K. Diversity of fall armyworm, Spodoptera frugiperda and their gut bacterial community in Kenya. PeerJ 2020, 8, e8701. [Google Scholar] [CrossRef]
- Ugwu, J.A.; Liu, M.; Sun, H.; Asiegbu, F.O. Microbiome of the larvae of Spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae) from maize plants. J. Appl. Entomol. 2020, 144, 764–776. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Babraham Bioinformatics; Babraham Institute: Cambridge, UK, 2010. [Google Scholar]
- De Mandal, S.; Chatterjee, R.; Kumar, N.S. Dominant bacterial phyla in caves and their predicted functional roles in C and N cycle. BMC Microbiol. 2017, 17, 90. [Google Scholar] [CrossRef] [Green Version]
- De Mandal, S.; Panda, A.K.; Bisht, S.S.; Senthil Kumar, N. MiSeq HV4 16S rRNA gene analysis of bacterial community composition among the cave sediments of Indo-Burma biodiversity hotspot. Environ. Sci. Pollut. Res. 2016, 23, 12216–12226. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Liu, Y.-X.; Huang, L. ImageGP: An easy-to-use data visualization web server for scientific researchers. iMeta 2022, 1, e5. [Google Scholar] [CrossRef]
- Appel, H.M.; Martin, M.M. Gut redox conditions in herbivorous lepidopteran larvae. J. Chem. Ecol. 1990, 16, 3277–3290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, J.F. Insect acid-base physiology. Annu. Rev. Entomol. 2001, 46, 221–250. [Google Scholar] [CrossRef] [PubMed]
- Colman, D.R.; Toolson, E.C.; Takacs-Vesbach, C.D. Do diet and taxonomy influence insect gut bacterial communities? Mol. Ecol. 2012, 21, 5124–5137. [Google Scholar] [CrossRef]
- Mikaelyan, A.; Dietrich, C.; Köhler, T.; Poulsen, M.; Sillam-Dussès, D.; Brune, A. Diet is the primary determinant of bacterial community structure in the guts of higher termites. Mol. Ecol. 2015, 24, 5284–5295. [Google Scholar] [CrossRef]
- Sugio, A.; Dubreuil, G.; Giron, D.; Simon, J.-C. Plant–insect interactions under bacterial influence: Ecological implications and underlying mechanisms. J. Exp. Bot. 2015, 66, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Zheng, D.; Zhong, H.; Qin, B.; Gurr, G.M.; Vasseur, L.; Lin, H.; Bai, J.; He, W.; You, M. DNA sequencing reveals the midgut microbiota of diamondback moth, Plutella xylostella (L.) and a possible relationship with insecticide resistance. PLoS ONE 2013, 8, e68852. [Google Scholar] [CrossRef]
- Snyman, M.; Gupta, A.K.; Bezuidenhout, C.C.; Claassens, S.; Van den Berg, J. Gut microbiota of Busseola fusca (Lepidoptera: Noctuidae). World J. Microbiol. Biotechnol. 2016, 32, 1–9. [Google Scholar] [CrossRef]
- Landry, M.; Comeau, A.M.; Derome, N.; Cusson, M.; Levesque, R.C. Composition of the spruce budworm (Choristoneura fumiferana) midgut microbiota as affected by rearing conditions. PLoS ONE 2015, 10, e0144077. [Google Scholar] [CrossRef] [PubMed]
- Belda, E.; Pedrola, L.; Peretó, J.; Martínez-Blanch, J.F.; Montagud, A.; Navarro, E.; Urchueguía, J.; Ramón, D.; Moya, A.; Porcar, M. Microbial diversity in the midguts of field and lab-reared populations of the European corn borer Ostrinia nubilalis. PLoS ONE 2011, 6, e21751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Hu, X.; Yang, S.; Zhou, J.; Zhang, T.; Qi, L.; Sun, X.; Fan, M.; Xu, S.; Cha, M.; et al. Comparative analysis of the gut microbiota composition between captive and wild forest musk deer. Front. Microbiol. 2017, 8, 1705. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Teh, B.-S.; Sun, C.; Hu, S.; Lu, X.; Boland, W.; Shao, Y. Biodiversity and activity of the gut microbiota across the life history of the insect herbivore Spodoptera littoralis. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonknechten, N.; Chaussonnerie, S.; Tricot, S.; Lajus, A.; Andreesen, J.R.; Perchat, N.; Pelletier, E.; Gouyvenoux, M.; Barbe, V.; Salanoubat, M.; et al. Clostridium sticklandii, a specialist in amino acid degradation: Revisiting its metabolism through its genome sequence. BMC Genom. 2010, 11, 555. [Google Scholar] [CrossRef] [PubMed]
- Lü, J.; Guo, W.; Chen, S.; Guo, M.; Qiu, B.; Yang, C.; Lian, T.; Pan, H. Host plants influence the composition of the gut bacteria in Henosepilachna vigintioctopunctata. PLoS ONE 2019, 14, e0224213. [Google Scholar] [CrossRef] [Green Version]
- Björkroth, J.; Holzapfel, W. Genera Leuconostoc, Oenococcus and Weissella. In The Prokaryotes: A Handbook on the Biology of Bacteria: Firmicutes, Cyanobacteria; Dworkin, M., Ed.; Springer: New York, NY, USA, 2006; Volume 4. [Google Scholar]
- Chelo, I.M.; Ze-Ze, L.; Tenreiro, R. Congruence of evolutionary relationships inside the Leuconostoc–Oenococcus–Weissella clade assessed by phylogenetic analysis of the 16S rRNA gene, dnaA, gyrB, rpoC and dnaK. Int. J. Syst. Evol. Microbiol. 2007, 57, 276–286. [Google Scholar] [CrossRef]
- Scully, E.D.; Geib, S.M.; Hoover, K.; Tien, M.; Tringe, S.G.; Barry, K.W.; del Rio, T.G.; Chovatia, M.; Herr, J.R.; Carlson, J.E. Metagenomic profiling reveals lignocellulose degrading system in a microbial community associated with a wood-feeding beetle. PLoS ONE 2013, 8, e73827. [Google Scholar] [CrossRef] [Green Version]
- Mereghetti, V.; Chouaia, B.; Montagna, M. New Insights into the Microbiota of Moth Pests. Int. J. Mol. Sci. 2017, 18, 2450. [Google Scholar] [CrossRef] [Green Version]
- Broderick, N.A.; Raffa, K.F.; Goodman, R.M.; Handelsman, J. Census of the bacterial community of the gypsy moth larval midgut by using culturing and culture-independent methods. Appl. Environ. Microbiol. 2004, 70, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Arias-Cordero, E.; Guo, H.; Bartram, S.; Boland, W. In vivo Pyro-SIP assessing active gut microbiota of the cotton leafworm, Spodoptera littoralis. PLoS ONE 2014, 9, e85948. [Google Scholar]
- Shao, Y.; Spiteller, D.; Tang, X.; Ping, L.; Colesie, C.; Münchberg, U.; Bartram, S.; Schneider, B.; Büdel, B.; Popp, J.; et al. Crystallization of α-and β-carotene in the foregut of Spodoptera larvae feeding on a toxic food plant. Insect Biochem. Mol. Biol. 2011, 41, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, N.; Martens, R.; Tebbe, C.C. Origin and diversity of metabolically active gut bacteria from laboratory-bred larvae of Manduca sexta (Sphingidae, Lepidoptera, Insecta). Appl. Environ. Microbiol. 2008, 74, 7189–7196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madhusudan, S.; Jalali, S.; Venkatesan, T.; Lalitha, Y.; Srinivas, R. 16S rRNA gene based identification of gut bacteria from laboratory and wild larvae of Helicoverpa armigera (Lepidoptera: Noctuidae) from tomato farm. Bioscan 2011, 6, 175–183. [Google Scholar]
- Staudacher, H.; Kaltenpoth, M.; Breeuwer, J.A.; Menken, S.B.; Heckel, D.G.; Groot, A.T. Variability of bacterial communities in the moth Heliothis virescens indicates transient association with the host. PLoS ONE 2016, 11, e0154514. [Google Scholar] [CrossRef] [Green Version]
- Srionnual, S.; Yanagida, F.; Lin, L.-H.; Hsiao, K.-N.; Chen, Y.-s. Weissellicin 110, a newly discovered bacteriocin from Weissella cibaria 110, isolated from plaa-som, a fermented fish product from Thailand. Appl. Environ. Microbiol. 2007, 73, 2247–2250. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Cho, S.-M.; Kim, M.; Ko, Y.-G.; Yong, D.; Lee, K. Weissella confusa bacteremia in an immune-competent patient with underlying intramural hematomas of the aorta. Ann. Lab. Med. 2013, 33, 459–462. [Google Scholar] [CrossRef]
- Kamboj, K.; Vasquez, A.; Balada-Llasat, J.-M. Identification and significance of Weissella species infections. Front. Microbiol. 2015, 6, 1204. [Google Scholar] [CrossRef]
- Kang, M.-S.; Yeu, J.-E.; Hong, S.-P. Safety Evaluation of Oral Care Probiotics Weissella cibaria CMU and CMS1 by Phenotypic and Genotypic Analysis. Int. J. Mol. Sci. 2019, 20, 2693. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.; Hamada, M.; Cho, H.; Weon, H.-Y.; Kim, J.-S.; Hong, S.-B.; Kim, S.-J.; Kwon, S.-W. Weissella cryptocerci sp. nov., isolated from gut of the insect Cryptocercus kyebangensis. Int. J. Syst. Evol. Microbiol. 2019, 69, 2801–2806. [Google Scholar] [CrossRef]
- Praet, J.; Meeus, I.; Cnockaert, M.; Houf, K.; Smagghe, G.; Vandamme, P. Novel lactic acid bacteria isolated from the bumble bee gut: Convivina intestini gen. nov., sp. nov., Lactobacillus bombicola sp. nov., and Weissella bombi sp. nov. Antonie Van Leeuwenhoek 2015, 107, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Freitak, D.; Vogel, H.; Ping, L.; Shao, Y.; Cordero, E.A.; Andersen, G.; Westermann, M.; Heckel, D.G.; Boland, W. Complexity and variability of gut commensal microbiota in polyphagous lepidopteran larvae. PLoS ONE 2012, 7, e36978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwong, W.K.; Mancenido, A.L.; Moran, N.A. Immune system stimulation by the native gut microbiota of honey bees. R. Soc. Open Sci. 2017, 4, 170003. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Zhao, B.a.; Jin, X.; Cheng, X.; Huang, S.; Li, J. Antibiotic Treatment Decrease the Fitness of Honeybee (Apis mellifera) Larvae. Insects 2021, 12, 301. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, H.; Lai, Y.; Chen, Q.; Yu, X.-Q.; Wang, X. Gut Microbiota Dysbiosis Influences Metabolic Homeostasis in Spodoptera frugiperda. Front. Microbiol. 2021, 12, 2803. [Google Scholar] [CrossRef]
- Li, G.; Xia, X.; Zhao, S.; Shi, M.; Liu, F.; Zhu, Y. The physiological and toxicological effects of antibiotics on an interspecies insect model. Chemosphere 2020, 248, 126019. [Google Scholar] [CrossRef]
- Thakur, A.; Dhammi, P.; Saini, H.S.; Kaur, S. Effect of antibiotic on survival and development of Spodoptera litura (Lepidoptera: Noctuidae) and its gut microbial diversity. Bull. Entomol. Res. 2016, 106, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Krams, I.A.; Kecko, S.; Jõers, P.; Trakimas, G.; Elferts, D.; Krams, R.; Luoto, S.; Rantala, M.J.; Inashkina, I.; Gudrā, D.; et al. Microbiome symbionts and diet diversity incur costs on the immune system of insect larvae. J. Exp. Biol. 2017, 220, 4204–4212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fellous, S.; Lazzaro, B.P. Larval food quality affects adult (but not larval) immune gene expression independent of effects on general condition. Mol. Ecol. 2010, 19, 1462–1468. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, A.; He, J.; Yu, T.; Sun, C.; Shi, D.; Jiang, Y.; Xianyu, Y.; Shao, Y. Dietary exposure of copper and zinc oxides nanoparticles affect the fitness, enzyme activity, and microbial community of the model insect, silkworm Bombyx mori. Sci. Total Environ. 2022, 813, 152608. [Google Scholar] [CrossRef]
- Muhammad, A.; Zhou, X.; He, J.; Zhang, N.; Shen, X.; Sun, C.; Yan, B.; Shao, Y. Toxic effects of acute exposure to polystyrene microplastics and nanoplastics on the model insect, silkworm Bombyx mori. Environ. Pollut. 2021, 285, 117255. [Google Scholar] [CrossRef] [PubMed]
- Koch, H.; Schmid-Hempel, P. Socially transmitted gut microbiota protect bumble bees against an intestinal parasite. Proc. Natl. Acad. Sci. USA 2011, 108, 19288–19292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Lu, Y.; Zhang, H.; Kumar, D.; Liu, B.; Gong, Y.; Zhu, M.; Zhu, L.; Liang, Z.; Kuang, S.; et al. Effects of BmCPV infection on silkworm Bombyx mori intestinal bacteria. PLoS ONE 2016, 11, e0146313. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Zhang, N.; Xie, S.; Zhang, X.; He, J.; Muhammad, A.; Sun, C.; Lu, X.; Shao, Y. Gut bacteria of the silkworm Bombyx mori facilitate host resistance against the toxic effects of organophosphate insecticides. Environ. Int. 2020, 143, 105886. [Google Scholar] [CrossRef] [PubMed]
- Mikonranta, L.; Mappes, J.; Kaukoniitty, M.; Freitak, D. Insect immunity: Oral exposure to a bacterial pathogen elicits free radical response and protects from a recurring infection. Front. Zool. 2014, 11, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. The B vitamin nutrition of insects: The contributions of diet, microbiome and horizontally acquired genes. Curr. Opin. Insect Sci. 2017, 23, 65–69. [Google Scholar] [CrossRef]
- Li, T.; Zhang, Q.; Zhang, X.; Wan, Q.; Wang, S.; Zhang, R.; Zhang, Z. Transcriptome and microbiome analyses of the mechanisms underlying antibiotic-mediated inhibition of larval development of the saprophagous insect Musca domestica (Diptera: Muscidae). Ecotoxicol. Environ. Saf. 2021, 223, 112602. [Google Scholar] [CrossRef]
- Hosokawa, T.; Kikuchi, Y.; Shimada, M.; Fukatsu, T. Obligate symbiont involved in pest status of host insect. Proc. R. Soc. B Biol. Sci. 2007, 274, 1979–1984. [Google Scholar] [CrossRef] [Green Version]
- Marcinkevicius, E.V.; Shirasu-Hiza, M.M. Message in a biota: Gut microbes signal to the circadian clock. Cell Host Microbe 2015, 17, 541–543. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alpha Diversity Index | Lab Diet | Field Diet | p-Value |
|---|---|---|---|
| Observed OTUs | 108.6 ± 11.43 | 75.80 ± 31.01 | 0.09 |
| Chao | 129.77 ± 21.78 | 85.19 ± 33.99 | 0.03 |
| Ace | 138.65 ± 26.67 | 88.04 ± 35.11 | 0.01 |
| Shannon | 0.93 ± 0.10 | 0.57 ± 0.30 | 0.05 |
| Simpson | 0.48 ± 0.01 | 0.73 ± 0.19 | 0.03 |
| Coverage | 0.999 ± 0.0001 | 0.999 ± 0.0001 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; De Mandal, S.; Wu, H.; Zhu, S.; Kong, J.; Lin, S.; Jin, F. Effect of Diet on the Midgut Microbial Composition and Host Immunity of the Fall Armyworm, Spodoptera frugiperda. Biology 2022, 11, 1602. https://doi.org/10.3390/biology11111602
Xu X, De Mandal S, Wu H, Zhu S, Kong J, Lin S, Jin F. Effect of Diet on the Midgut Microbial Composition and Host Immunity of the Fall Armyworm, Spodoptera frugiperda. Biology. 2022; 11(11):1602. https://doi.org/10.3390/biology11111602
Chicago/Turabian StyleXu, Xiaoxia, Surajit De Mandal, Hongxin Wu, Shaojie Zhu, Jinrong Kong, Sisi Lin, and Fengliang Jin. 2022. "Effect of Diet on the Midgut Microbial Composition and Host Immunity of the Fall Armyworm, Spodoptera frugiperda" Biology 11, no. 11: 1602. https://doi.org/10.3390/biology11111602