
SMAC Mimetic BV6 Co-Treatment Downregulates the Factors Involved in Resistance and Relapse of Cancer: IAPs and Autophagy
 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Maintenance
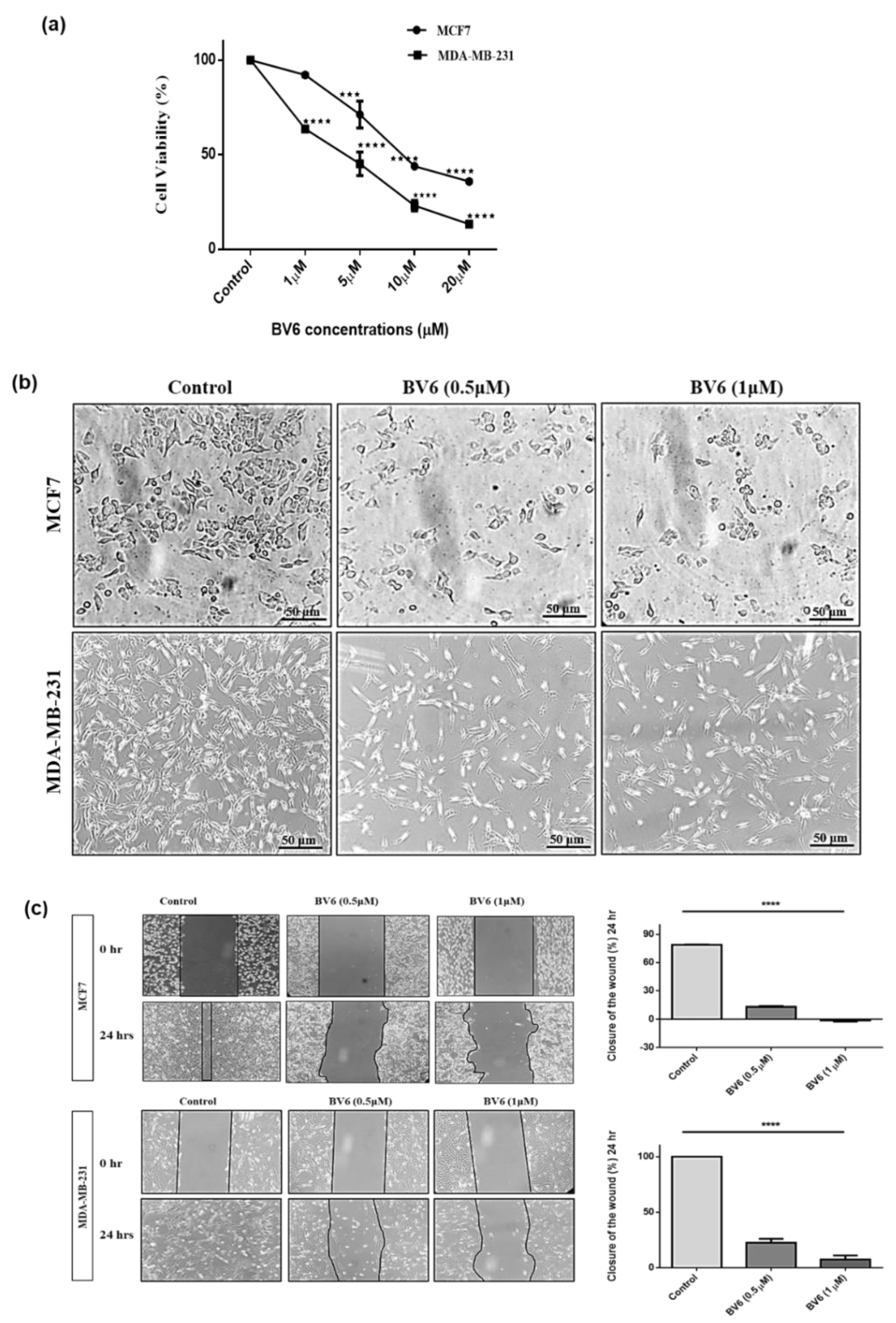
2.2. MTT Assay
2.3. Cellular Morphology
2.4. Wound Healing
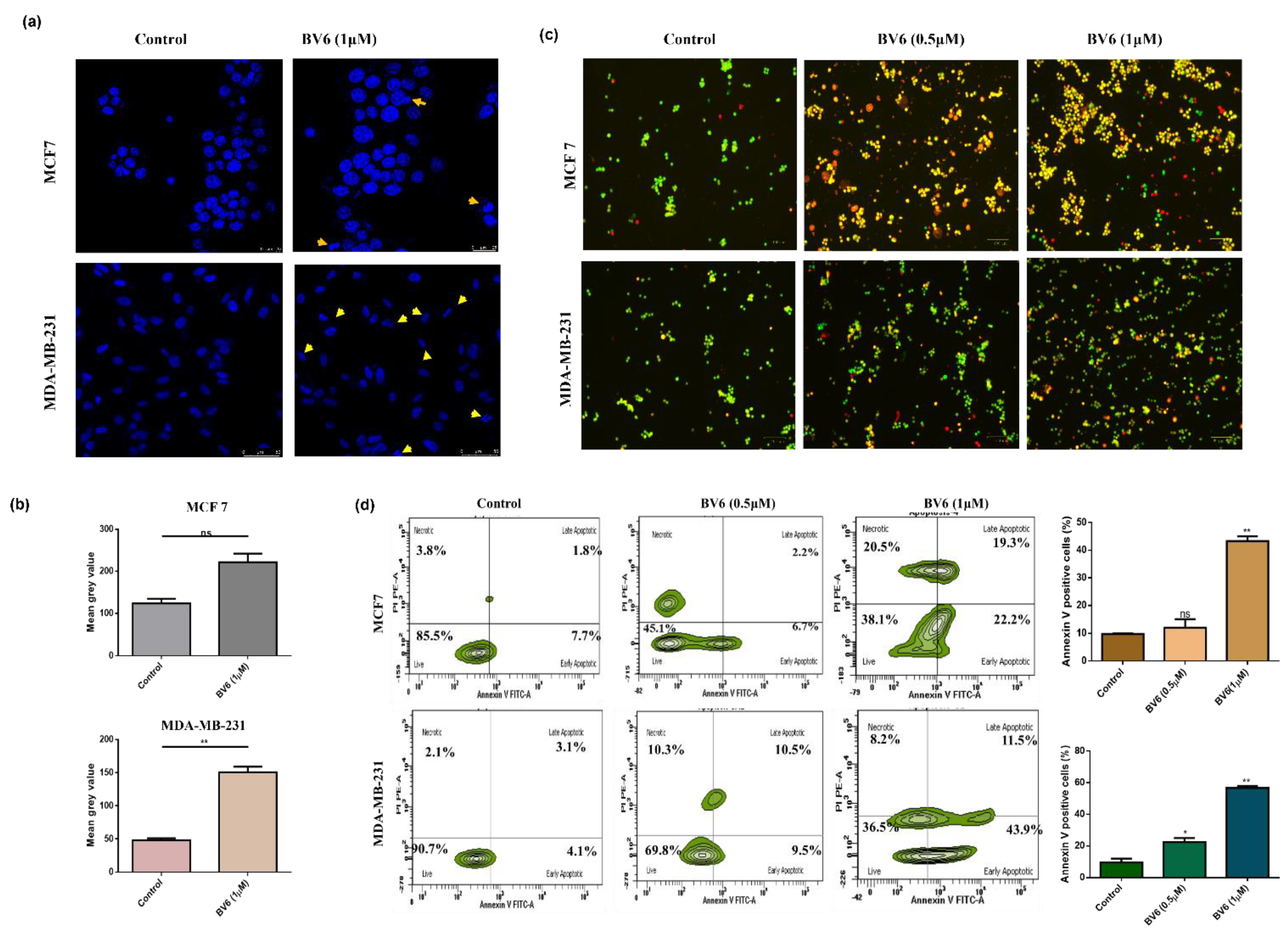
2.5. Acridine Orange and Ethidium Bromide Staining
2.6. DAPI Staining
2.7. Annexin V Staining Assay
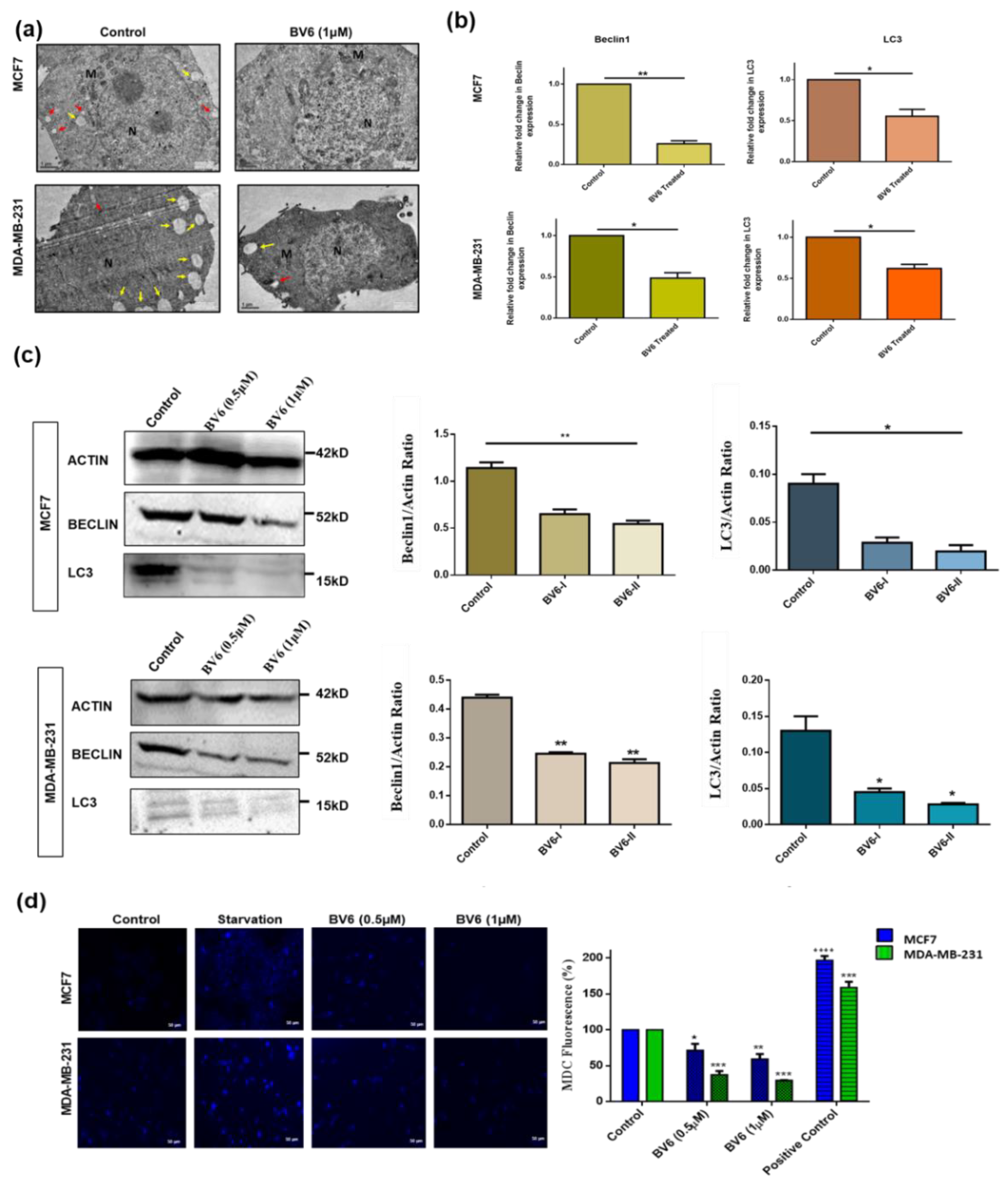
2.8. MDC Staining
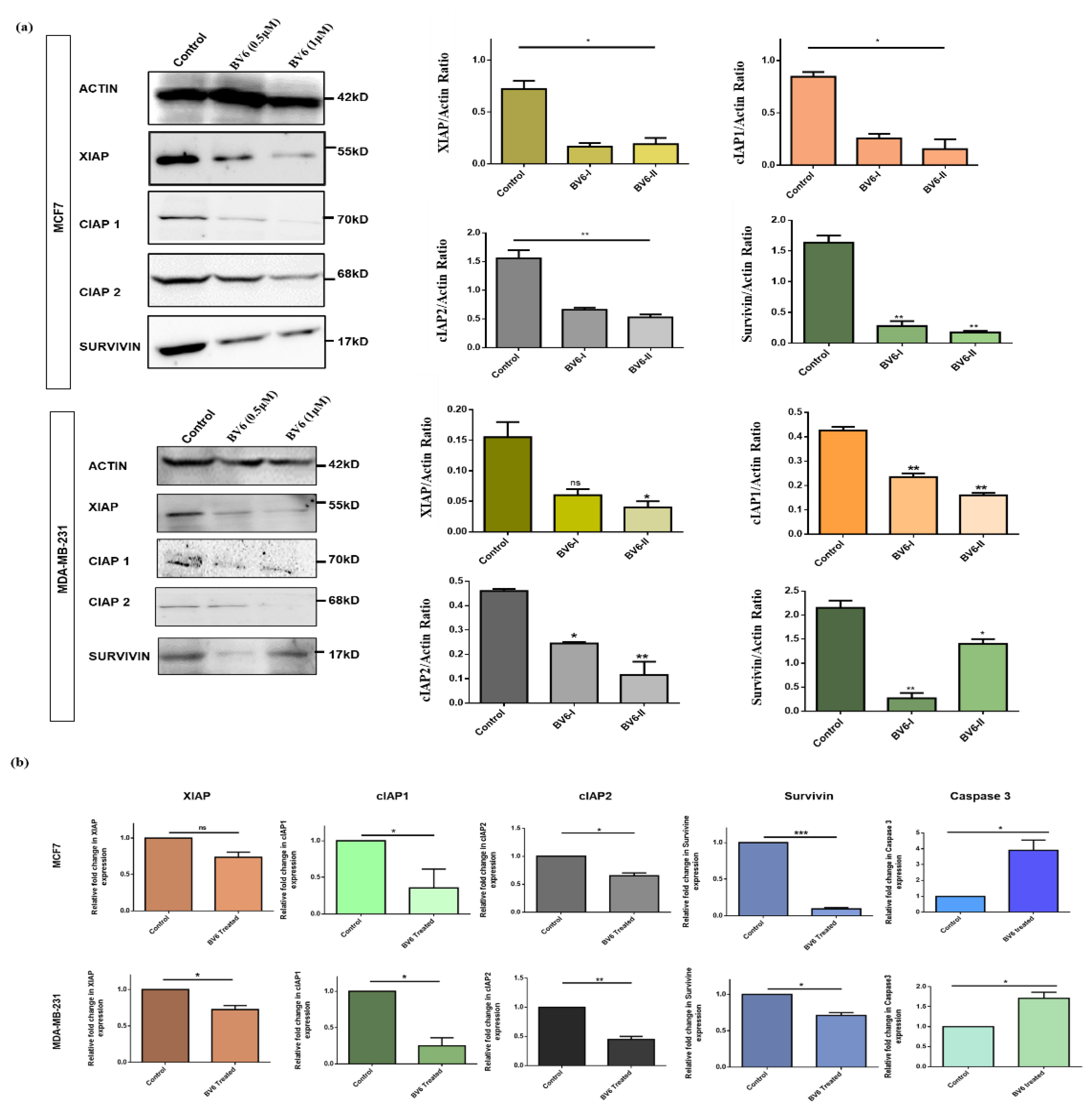
2.9. Real-Time PCR
2.10. Western Immunoblotting
2.11. Transmission Electron Microscope (TEM)
2.12. Statistical Analysis
3. Results
3.1. BV6 Effect on Cell Viability, Cell Morphology and Migration in MCF7 and MDA-MB-231
3.2. BV6 Induces Apoptosis in MCF7 and MDA-MB-231
3.3. BV6 Exposure Downregulated the Expression of IAPs
3.4. Inhibition of Autophagy in MCF7 and MDA-MB-231 Cells Treated with BV6
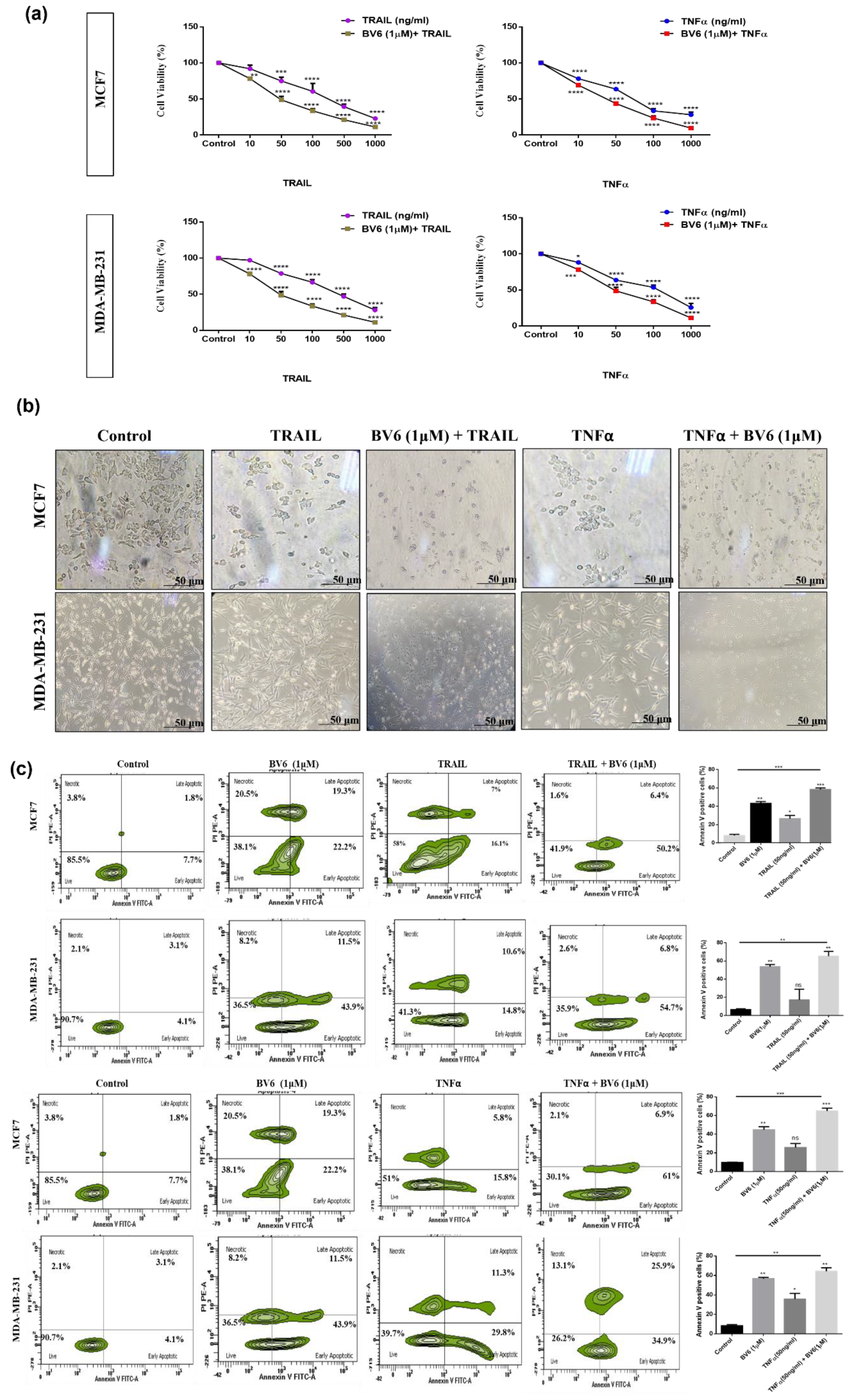
3.5. Effect of BV6 Cotreated with TRAIL and TNFα on MCF7 and MDA-MB-231 Cell Viability, Cell Morphology and Apoptosis
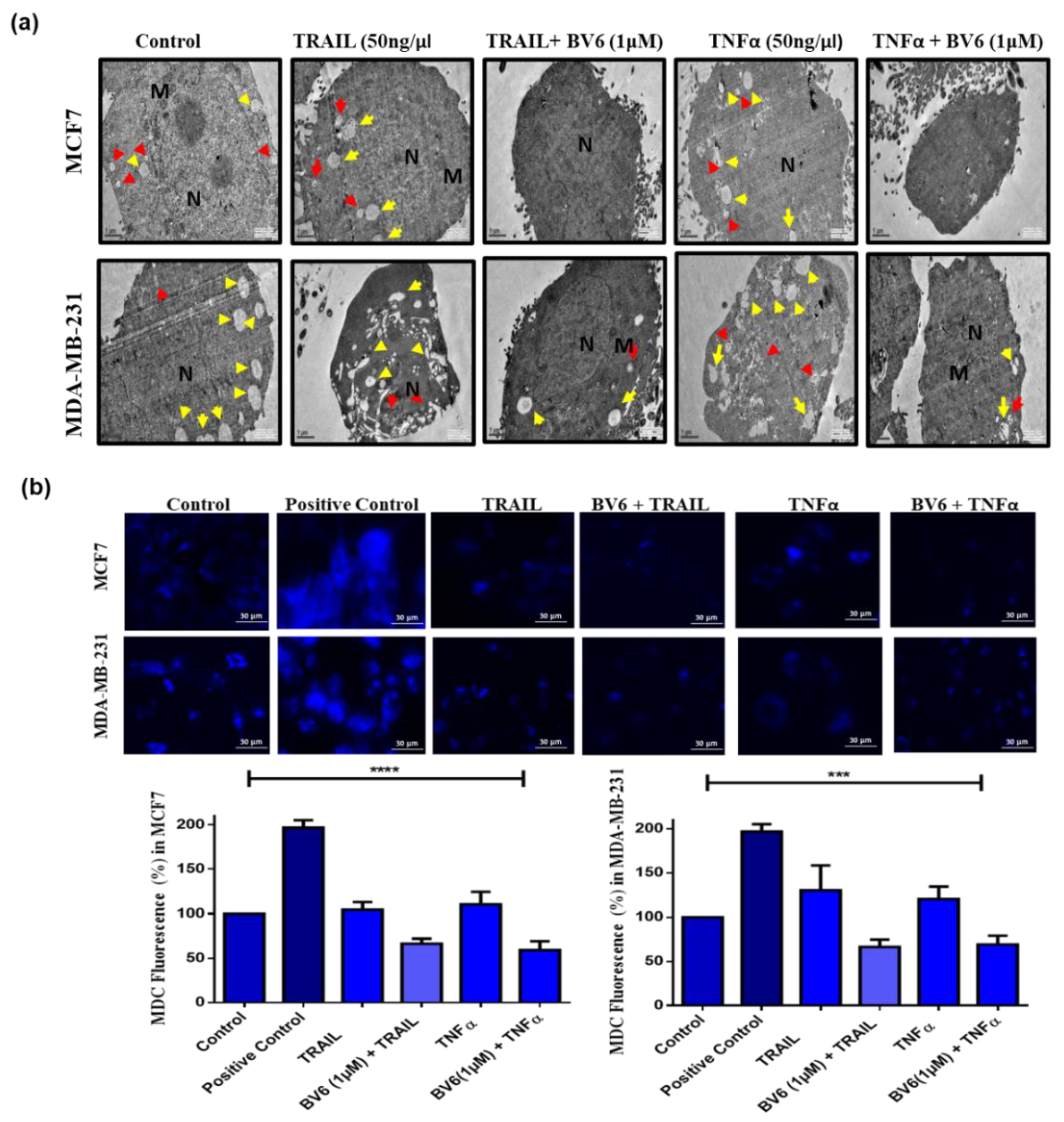
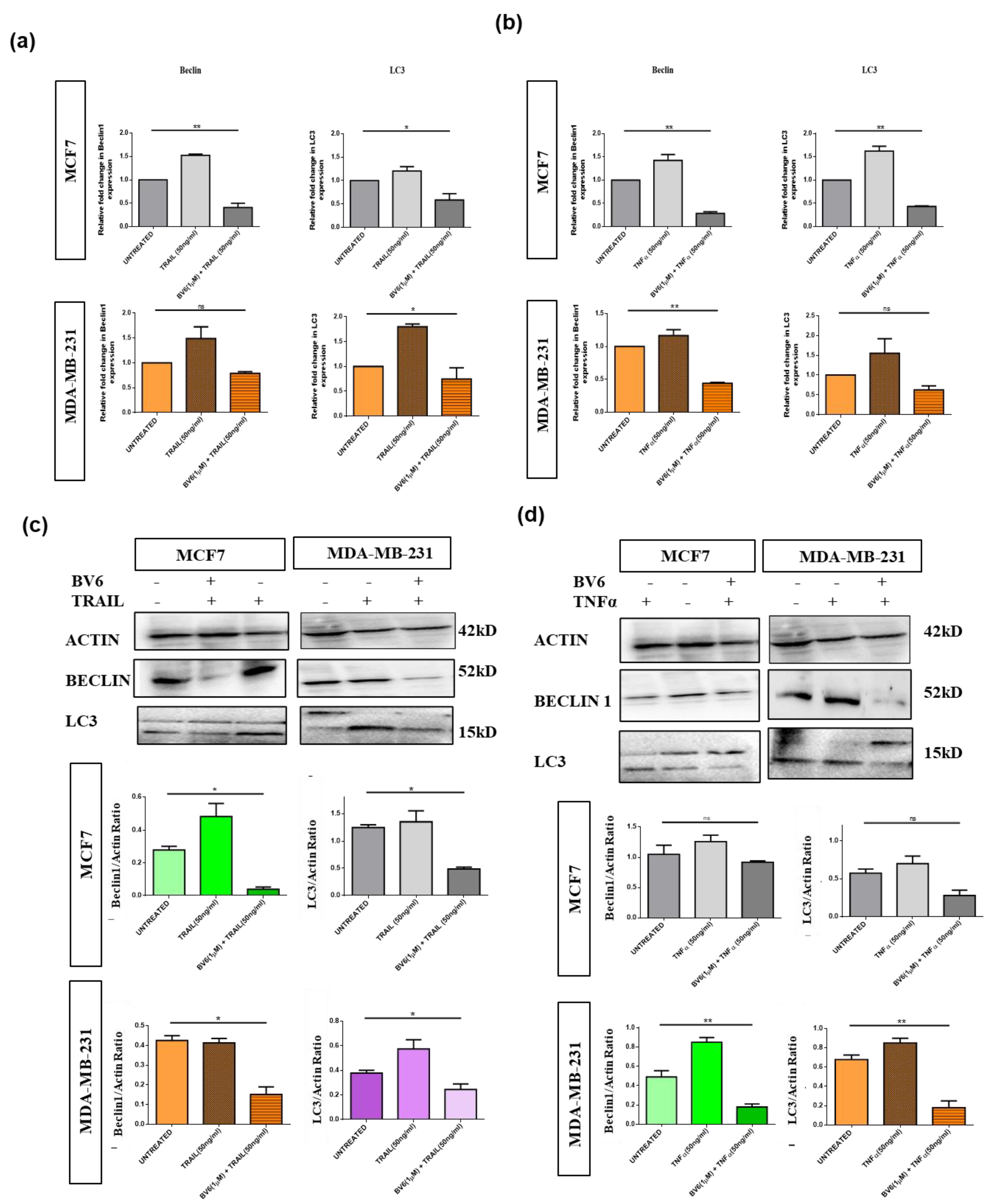
3.6. Autophagy in MCF7 and MDA-MB-231 Cells Treated with TRAIL, TNFα and in Synergy with BV6
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Projections in Breast and Lung Cancer Mortality among Women: A Bayesian Analysis of 52 Countries Worldwide|Cancer Research. Available online: https://cancerres.aacrjournals.org/content/78/15/4436.short (accessed on 3 January 2022).
- Nikkhoo, A.; Rostami, N.; Hojjat-Farsangi, M.; Azizi, G.; Yousefi, B.; Ghalamfarsa, G.; Jadidi-Niaragh, F. Smac mimetics as novel promising modulators of apoptosis in the treatment of breast cancer. J. Cell. Biochem. 2019, 120, 9300–9314. [Google Scholar] [CrossRef]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef]
- Death and Anti-Death: Tumour Resistance to Apoptosis|Nature Reviews Cancer. Available online: https://www.nature.com/articles/nrc776 (accessed on 7 August 2021).
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Fulda, S.; Vucic, D. Targeting IAP proteins for therapeutic intervention in cancer. Nat. Rev. Drug Discov. 2012, 11, 109–124. [Google Scholar] [CrossRef]
- Pluta, P.; Jeziorski, A.; Cebula-Obrzut, A.P.; Wierzbowska, A.; Piekarski, J.; Smolewski, P. Expression of IAP family proteins and its clinical importance in breast cancer patients. Neoplasma 2015, 62, 666–673. [Google Scholar] [CrossRef] [Green Version]
- Abbas, R.; Larisch, S. Targeting XIAP for Promoting Cancer Cell Death—The Story of ARTS and SMAC. Cells 2020, 9, 663. [Google Scholar] [CrossRef] [Green Version]
- Flygare, J.A.; Beresini, M.; Budha, N.; Chan, H.; Chan, I.T.; Cheeti, S.; Cohen, F.; Deshayes, K.; Doerner, K.; Eckhardt, S.G.; et al. Discovery of a potent small-molecule antagonist of inhibitor of apoptosis (IAP) proteins and clinical candidate for the treatment of cancer (GDC-0152). J. Med. Chem. 2012, 55, 4101–4113. [Google Scholar] [CrossRef] [Green Version]
- Hehlgans, S.; Oppermann, J.; Reichert, S.; Fulda, S.; Rödel, C.; Rödel, F. The SMAC mimetic BV6 sensitizes colorectal cancer cells to ionizing radiation by interfering with DNA repair processes and enhancing apoptosis. Radiat. Oncol. 2015, 10, 198. [Google Scholar] [CrossRef] [Green Version]
- El-Mesery, M.; Shaker, M.E.; Elgaml, A. The SMAC mimetic BV6 induces cell death and sensitizes different cell lines to TNF-α and TRAIL-induced apoptosis. Exp. Biol. Med. 2016, 241, 2015–2022. [Google Scholar] [CrossRef] [Green Version]
- Targeting Triple-Negative Breast Cancers with the Smac-Mimetic Birinapant|Cell Death & Differentiation. Available online: https://www.nature.com/articles/s41418-020-0541-0 (accessed on 8 August 2021).
- Wang, X.; Lu, M.; Gu, H.; Xiao, T.; Hu, G.; Luo, M.; Guo, X.; Xia, Y. Conjugation of the Fn14 Ligand to a SMAC Mimetic Selectively Suppresses Experimental Squamous Cell Carcinoma in Mice. J. Investig. Dermatol. 2022, in press. [CrossRef]
- Aredia, F.; Guamán Ortiz, L.M.; Giansanti, V.; Scovassi, A.I. Autophagy and Cancer. Cells 2012, 1, 520–534. [Google Scholar] [CrossRef] [Green Version]
- Amaravadi, R.; Kimmelman, A.C.; White, E. Recent insights into the function of autophagy in cancer. Genes Dev. 2016, 30, 1913–1930. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.K.; Bharti, P.S.; Rafat, S.; Bhatt, D.; Goyal, Y.; Pandey, K.K.; Ranjan, S.; Almatroodi, S.A.; Alsahli, M.A.; Rahmani, A.H.; et al. Autophagy Paradox of Cancer: Role, Regulation, and Duality. Oxid. Med. Cell. Longev. 2021, 2021, e8832541. [Google Scholar] [CrossRef]
- Galluzzi, L.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cecconi, F.; Choi, A.M.; Chu, C.T.; Codogno, P.; Colombo, M.I.; et al. Molecular definitions of autophagy and related processes. EMBO J. 2017, 36, 1811–1836. [Google Scholar] [CrossRef]
- Onorati, A.V.; Dyczynski, M.; Ojha, R.; Amaravadi, R.K. Targeting autophagy in cancer. Cancer 2018, 124, 3307–3318. [Google Scholar] [CrossRef] [Green Version]
- Cancer Drug Resistance: An Evolving Paradigm|Nature Reviews Cancer. Available online: https://www.nature.com/articles/nrc3599 (accessed on 8 August 2021).
- Targeting Autophagy in Cancer—Onorati—2018—Cancer—Wiley Online Library. Available online: https://acsjournals.onlinelibrary.wiley.com/doi/full/10.1002/cncr.31335 (accessed on 8 August 2021).
- Ke, B.; Tian, M.; Li, J.; Liu, B.; He, G. Targeting Programmed Cell Death Using Small-Molecule Compounds to Improve Potential Cancer Therapy: Anticancer Compounds Targeting Cell Death. Med. Res. Rev. 2016, 36, 983–1035. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, B.; Mani, A.M.; Wu, Z.; Fan, Y.; Li, W.; Wu, Z.-H. Survivin Inhibitors Mitigate Chemotherapeutic Resistance in Breast Cancer Cells by Suppressing Genotoxic Nuclear Factor-κB Activation. J. Pharmacol. Exp. Ther. 2018, 366, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Benetatos, C.A.; Mitsuuchi, Y.; Burns, J.M.; Neiman, E.M.; Condon, S.M.; Yu, G.; Seipel, M.E.; Kapoor, G.S.; Laporte, M.G.; Rippin, S.R.; et al. Birinapant (TL32711), a bivalent SMAC mimetic, targets TRAF2-associated cIAPs, abrogates TNF-induced NF-κB activation, and is active in patient-derived xenograft models. Mol. Cancer Ther. 2014, 13, 867–879. [Google Scholar] [CrossRef] [Green Version]
- Panayotopoulou, E.G.; Müller, A.-K.; Börries, M.; Busch, H.; Hu, G.; Lev, S. Targeting of apoptotic pathways by SMAC or BH3 mimetics distinctly sensitizes paclitaxel-resistant triple-negative breast cancer cells. Oncotarget 2017, 8, 45088–45104. [Google Scholar] [CrossRef] [Green Version]
- Ehrenschwender, M.; Bittner, S.; Seibold, K.; Wajant, H. XIAP-targeting drugs re-sensitize PIK3CA-mutated colorectal cancer cells for death receptor-induced apoptosis. Cell Death Dis. 2014, 5, e1570. [Google Scholar] [CrossRef]
- Lei, W.; Duan, R.; Li, J.; Liu, X.; Huston, A.; Boyce, B.F.; Yao, Z. The IAP Antagonist SM-164 Eliminates Triple-Negative Breast Cancer Metastasis to Bone and Lung in Mice. Sci. Rep. 2020, 10, 7004. [Google Scholar] [CrossRef] [Green Version]
- Allensworth, J.L.; Sauer, S.J.; Lyerly, H.K.; Morse, M.A.; Devi, G.R. Smac mimetic Birinapant induces apoptosis and enhances TRAIL potency in inflammatory breast cancer cells in an IAP-dependent and TNF-α-independent mechanism. Breast Cancer Res. Treat. 2013, 137, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Kimmelman, A.C.; White, E. Autophagy and Tumor Metabolism. Cell Metab. 2017, 25, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Brech, A.; Ahlquist, T.; Lothe, R.A.; Stenmark, H. Autophagy in tumour suppression and promotion. Mol. Oncol. 2009, 3, 366–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, Y.; Tam, K.Y. Pathological mechanisms and therapeutic strategies for Alzheimer’s disease. Neural Regen. Res. 2022, 17, 543–549. [Google Scholar] [CrossRef]
- Richard, P.; Feng, S.; Tsai, Y.-L.; Li, W.; Rinchetti, P.; Muhith, U.; Irizarry-Cole, J.; Stolz, K.; Sanz, L.A.; Hartono, S.; et al. SETX (senataxin), the helicase mutated in AOA2 and ALS4, functions in autophagy regulation. Autophagy 2021, 17, 1889–1906. [Google Scholar] [CrossRef]
- Richard, P.; Rosonina, E. Regulating autophagy: A novel role for SETX (Senataxin). Neural Regen. Res. 2021, 16, 2008–2009. [Google Scholar] [CrossRef]
- He, J.-D.; Wang, Z.; Li, S.-P.; Xu, Y.-J.; Yu, Y.; Ding, Y.-J.; Yu, W.-L.; Zhang, R.-X.; Zhang, H.-M.; Du, H.-Y. Vitexin suppresses autophagy to induce apoptosis in hepatocellular carcinoma via activation of the JNK signaling pathway. Oncotarget 2016, 7, 84520–84532. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-L.; Liu, J.-F.; Liu, Y.; Wang, Y.-X.; Fu, K.-F.; Yu, X.-J.; Pu, Q.; Chen, X.-X.; Zhou, L.-J. Beclin1 inhibition enhances paclitaxel-mediated cytotoxicity in breast cancer in vitro and in vivo. Int. J. Mol. Med. 2019, 43, 1866–1878. [Google Scholar] [CrossRef]
- Xi, G.; Hu, X.; Wu, B.; Jiang, H.; Young, C.Y.F.; Pang, Y.; Yuan, H. Autophagy inhibition promotes paclitaxel-induced apoptosis in cancer cells. Cancer Lett. 2011, 307, 141–148. [Google Scholar] [CrossRef]
- Xiao, M.; Benoit, A.; Hasmim, M.; Duhem, C.; Vogin, G.; Berchem, G.; Noman, M.Z.; Janji, B. Targeting Cytoprotective Autophagy to Enhance Anticancer Therapies. Front. Oncol. 2021, 11, 180. [Google Scholar] [CrossRef]
- Cheung, C.H.A.; Chang, Y.-C.; Lin, T.-Y.; Cheng, S.M.; Leung, E. Anti-apoptotic proteins in the autophagic world: An update on functions of XIAP, Survivin, and BRUCE. J. Biomed. Sci. 2020, 27, 31. [Google Scholar] [CrossRef] [PubMed]
- Li, B.-X.; Wang, H.-B.; Qiu, M.-Z.; Luo, Q.-Y.; Yi, H.-J.; Yan, X.-L.; Pan, W.-T.; Yuan, L.-P.; Zhang, Y.-X.; Xu, J.-H.; et al. Novel smac mimetic APG-1387 elicits ovarian cancer cell killing through TNF-alpha, Ripoptosome and autophagy mediated cell death pathway. J. Exp. Clin. Cancer Res. 2018, 37, 53. [Google Scholar] [CrossRef] [PubMed]
- Véquaud, E.; Séveno, C.; Loussouarn, D.; Engelhart, L.; Campone, M.; Juin, P.; Barillé-Nion, S. YM155 potently triggers cell death in breast cancer cells through an autophagy-NF-kB network. Oncotarget 2015, 6, 13476–13486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peiqi, L.; Rong, H.; Hongming, D.; Zhuogang, L.; Wei, J.; Miao, M. GDC-0152-induced autophagy promotes apoptosis in HL-60 cells. Mol. Cell. Biochem. 2018, 445, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Gradzka, S.; Thomas, O.S.; Kretz, O.; Haimovici, A.; Vasilikos, L.; Wong, W.W.-L.; Häcker, G.; Gentle, I.E. Inhibitor of apoptosis proteins are required for effective fusion of autophagosomes with lysosomes. Cell Death Dis. 2018, 9, 529. [Google Scholar] [CrossRef]
- Rafat, S.; Dar, M.I.; Sunita, K.; Khan, S.; Verma, A.K.; Ahmad, F.; Dev, K. Therapeutic potential and protective effect against induced ROS and autophagy inhibition of AT101 compound in human breast cancer cell line MCF7. Gene Rep. 2022, 27, 101560. [Google Scholar] [CrossRef]
- Lin, F.; Ghislat, G.; Luo, S.; Renna, M.; Siddiqi, F.; Rubinsztein, D.C. XIAP and cIAP1 amplifications induce Beclin 1-dependent autophagy through NFκB activation. Hum. Mol. Genet. 2015, 24, 2899–2913. [Google Scholar] [CrossRef] [Green Version]
- Qiao, X.; Wang, X.; Shang, Y.; Li, Y.; Chen, S.-Z. Azithromycin enhances the anticancer activity of TRAIL by inhibiting autophagy and up-regulating the protein levels of DR4/5 in colon cancer cells in vitro and in vivo. Cancer Commun. Lond. Engl. 2018, 38, 43. [Google Scholar] [CrossRef] [Green Version]
- Lv, S.; Wang, X.; Zhang, N.; Sun, M.; Qi, W.; Li, Y.; Yang, Q. Autophagy facilitates the development of resistance to the tumor necrosis factor superfamily member TRAIL in breast cancer. Int. J. Oncol. 2015, 46, 1286–1294. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rafat, S.; Singh, P.; Pandey, K.K.; Almatroodi, S.A.; Alsahli, M.A.; Almatroudi, A.; Rahmani, A.H.; Dev, K. SMAC Mimetic BV6 Co-Treatment Downregulates the Factors Involved in Resistance and Relapse of Cancer: IAPs and Autophagy. Biology 2022, 11, 1581. https://doi.org/10.3390/biology11111581
Rafat S, Singh P, Pandey KK, Almatroodi SA, Alsahli MA, Almatroudi A, Rahmani AH, Dev K. SMAC Mimetic BV6 Co-Treatment Downregulates the Factors Involved in Resistance and Relapse of Cancer: IAPs and Autophagy. Biology. 2022; 11(11):1581. https://doi.org/10.3390/biology11111581
Chicago/Turabian StyleRafat, Sahar, Prabhakar Singh, Kamlesh Kumar Pandey, Saleh A. Almatroodi, Mohammed A. Alsahli, Ahmad Almatroudi, Arshad Husain Rahmani, and Kapil Dev. 2022. "SMAC Mimetic BV6 Co-Treatment Downregulates the Factors Involved in Resistance and Relapse of Cancer: IAPs and Autophagy" Biology 11, no. 11: 1581. https://doi.org/10.3390/biology11111581