
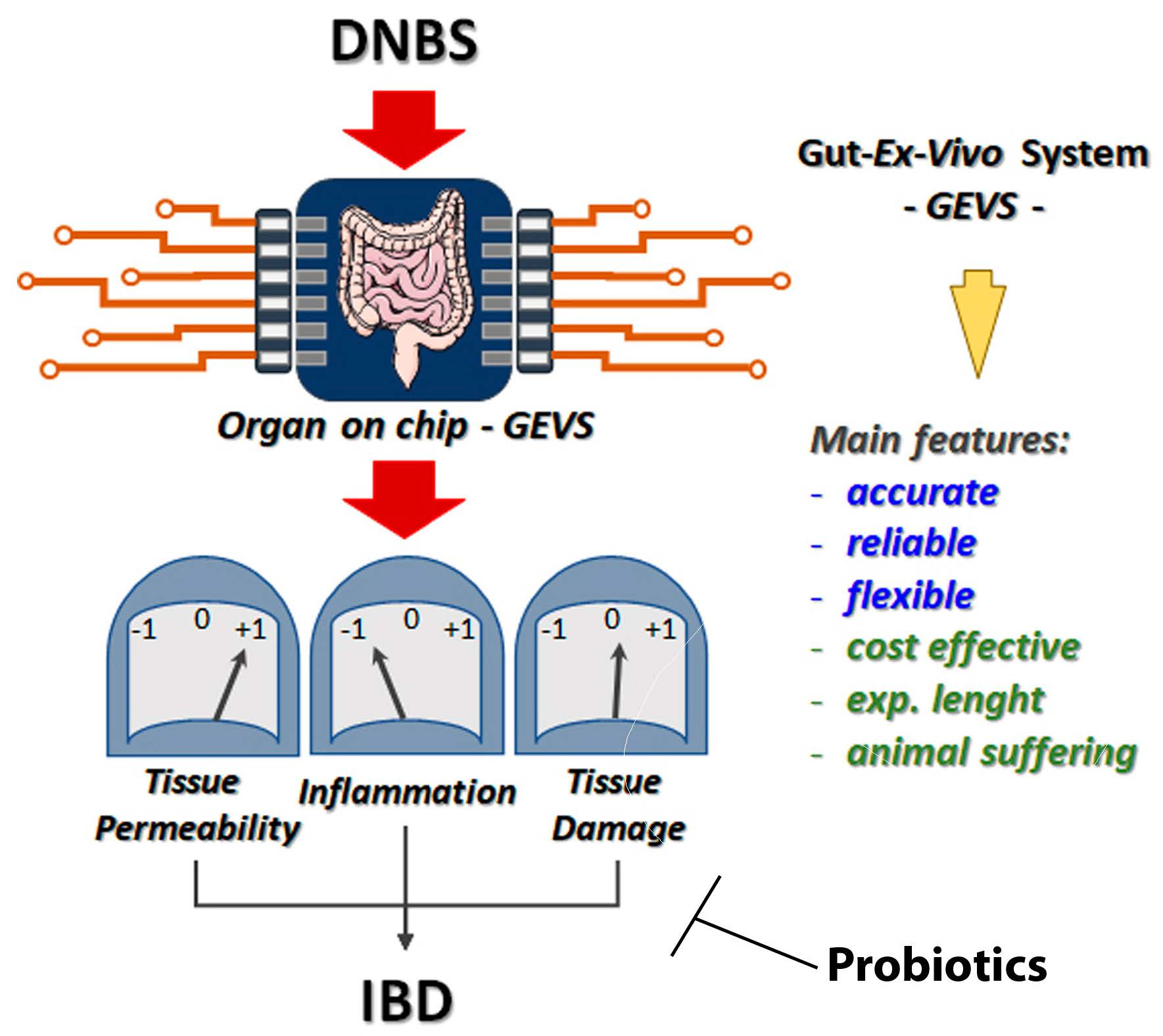
The Gut-Ex-Vivo System (GEVS) Is a Dynamic and Versatile Tool for the Study of DNBS-Induced IBD in BALB/C and C57BL/6 Mice, Highlighting the Protective Role of Probiotics
 , , ,
, , ,
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Silicone-Based Device and Colon Culture
2.3. Colon Cultures and Treatments
2.4. Probiotics Formulations and Treatments
2.5. Quantitative PCR (qPCR)
2.6. Western Blotting Analysis
2.7. Immunofluorescence Analysis
2.8. ELISA
2.9. Statistical Analysis
3. Results
3.1. DNBS Triggers UC in the Colon of C57BL/6 Mice by Using a Gut-Ex-Vivo System (GEVS)
3.2. Apoptotic and Ferroptotic Cell Death Modalities Associated with DNBS-Induced UC Can Be Studied by Using a GEVS, in Colon from Both C57BL/6 and BALB/c Mice
3.3. Probiotic Administration Prevents IBD Onset
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferrari, E.; Monzani, R.; Saverio, V.; Gagliardi, M.; Pańczyszyn, E.; Raia, V.; Villella, V.R.; Bona, G.; Pane, M.; Amoruso, A.; et al. Probiotics Supplements Reduce ER Stress and Gut Inflammation Associated with Gliadin Intake in a Mouse Model of Gluten Sensitivity. Nutrients 2021, 13, 1221. [Google Scholar] [CrossRef] [PubMed]
- Tam, R.Y.; van Dorst, J.M.; McKay, I.; Coffey, M.; Ooi, C.Y. Intestinal Inflammation and Alterations in the Gut Microbiota in Cystic Fibrosis: A Review of the Current Evidence, Pathophysiology and Future Directions. J. Clin. Med. 2022, 11, 649. [Google Scholar] [CrossRef]
- Gagliardi, M.; Monzani, R.; Clemente, N.; Fusaro, L.; Saverio, V.; Grieco, G.; Pańczyszyn, E.; Yissachar, N.; Boccafoschi, F.; Corazzari, M. A Gut-Ex-Vivo System to Study Gut Inflammation Associated to Inflammatory Bowel Disease (IBD). Biology 2021, 10, 605. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, Y.; Jiang, L.; Zhang, J.; Tong, X.; Chen, D.; Le, W. Intestinal Inflammation and Parkinson’s Disease. Aging Dis. 2021, 12, 2052–2068. [Google Scholar] [CrossRef] [PubMed]
- Bessac, A.; Cani, P.D.; Meunier, E.; Dietrich, G.; Knauf, C. Inflammation and Gut-Brain Axis During Type 2 Diabetes: Focus on the Crosstalk Between Intestinal Immune Cells and Enteric Nervous System. Front. Neurosci. 2018, 12, 725. [Google Scholar] [CrossRef]
- Jensen, C.; Teng, Y. Is It Time to Start Transitioning From 2D to 3D Cell Culture? Front. Mol. Biosci. 2020, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Decarli, M.C.; Amaral, R.; dos Santos, D.P.; Tofani, L.B.; Katayama, E.; Rezende, R.A.; da Silva, J.V.L.; Swiech, K.; Suazo, C.A.T.; Mota, C.; et al. Cell Spheroids as a Versatile Research Platform: Formation Mechanisms, High Throughput Production, Characterization and Applications. Biofabrication 2021, 13, 032002. [Google Scholar] [CrossRef]
- Kakni, P.; Truckenmüller, R.; Habibović, P.; Giselbrecht, S. Challenges to, and Prospects for, Reverse Engineering the Gastrointestinal Tract Using Organoids. Trends Biotechnol. 2022, 40, 932–944. [Google Scholar] [CrossRef]
- Eichele, D.D.; Kharbanda, K.K. Dextran Sodium Sulfate Colitis Murine Model: An Indispensable Tool for Advancing Our Understanding of Inflammatory Bowel Diseases Pathogenesis. World J. Gastroenterol. 2017, 23, 6016–6029. [Google Scholar] [CrossRef]
- Antoniou, E.; Margonis, G.A.; Angelou, A.; Pikouli, A.; Argiri, P.; Karavokyros, I.; Papalois, A.; Pikoulis, E. The TNBS-Induced Colitis Animal Model: An Overview. Ann. Med. Surg. 2016, 11, 9–15. [Google Scholar] [CrossRef]
- Barone, M.; Chain, F.; Sokol, H.; Brigidi, P.; Bermúdez-Humarán, L.G.; Langella, P.; Martín, R. A Versatile New Model of Chemically Induced Chronic Colitis Using an Outbred Murine Strain. Front. Microbiol. 2018, 9, 565. [Google Scholar] [CrossRef] [Green Version]
- Yissachar, N.; Zhou, Y.; Ung, L.; Lai, N.Y.; Mohan, J.F.; Ehrlicher, A.; Weitz, D.A.; Kasper, D.L.; Chiu, I.M.; Mathis, D.; et al. An Intestinal Organ Culture System Uncovers a Role for the Nervous System in Microbe-Immune Crosstalk. Cell 2017, 168, 1135–1148.e12. [Google Scholar] [CrossRef] [Green Version]
- Gagliardi, M.; Clemente, N.; Monzani, R.; Fusaro, L.; Ferrari, E.; Saverio, V.; Grieco, G.; Pańczyszyn, E.; Carton, F.; Santoro, C.; et al. Gut-Ex-Vivo System as a Model to Study Gluten Response in Celiac Disease. Cell Death Discov. 2021, 7, 45. [Google Scholar] [CrossRef]
- Giglio, P.; Gagliardi, M.; Tumino, N.; Antunes, F.; Smaili, S.; Cotella, D.; Santoro, C.; Bernardini, R.; Mattei, M.; Piacentini, M.; et al. PKR and GCN2 Stress Kinases Promote an ER Stress-Independent EIF2α Phosphorylation Responsible for Calreticulin Exposure in Melanoma Cells. Oncoimmunology 2018, 7, e1466765. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Yang, L.; Jiang, S.; Qian, D.; Duan, J. Excessive Apoptosis in Ulcerative Colitis: Crosstalk Between Apoptosis, ROS, ER Stress, and Intestinal Homeostasis. Inflamm. Bowel Dis. 2021, 28, 639–648. [Google Scholar] [CrossRef]
- Xu, M.; Tao, J.; Yang, Y.; Tan, S.; Liu, H.; Jiang, J.; Zheng, F.; Wu, B. Ferroptosis Involves in Intestinal Epithelial Cell Death in Ulcerative Colitis. Cell Death Dis. 2020, 11, 86. [Google Scholar] [CrossRef] [Green Version]
- Corazzari, M.; Lovat, P.E.; Armstrong, J.L.; Fimia, G.M.; Hill, D.S.; Birch-Machin, M.; Redfern, C.P.F.; Piacentini, M. Targeting Homeostatic Mechanisms of Endoplasmic Reticulum Stress to Increase Susceptibility of Cancer Cells to Fenretinide-Induced Apoptosis: The Role of Stress Proteins ERdj5 and ERp57. Br. J. Cancer 2007, 96, 1062–1071. [Google Scholar] [CrossRef]
- Nunes, T.; Bernardazzi, C.; de Souza, H.S. Cell Death and Inflammatory Bowel Diseases: Apoptosis, Necrosis, and Autophagy in the Intestinal Epithelium. BioMed Res. Int. 2014, 2014, 218493. [Google Scholar] [CrossRef]
- Pagliarini, V.; Giglio, P.; Bernardoni, P.; Zio, D.D.; Fimia, G.M.; Piacentini, M.; Corazzari, M. Downregulation of E2F1 during ER Stress Is Required to Induce Apoptosis. J. Cell Sci. 2015, 128, 1166–1179. [Google Scholar] [CrossRef] [Green Version]
- Finucane, D.M.; Bossy-Wetzel, E.; Waterhouse, N.J.; Cotter, T.G.; Green, D.R. Bax-Induced Caspase Activation and Apoptosis via Cytochrome c Release from Mitochondria Is Inhibitable by Bcl-XL. J. Biol. Chem. 1999, 274, 2225–2233. [Google Scholar] [CrossRef]
- Qiu, W.; Wu, B.; Wang, X.; Buchanan, M.E.; Regueiro, M.D.; Hartman, D.J.; Schoen, R.E.; Yu, J.; Zhang, L. PUMA-Mediated Intestinal Epithelial Apoptosis Contributes to Ulcerative Colitis in Humans and Mice. J. Clin. Investig. 2011, 121, 1722–1732. [Google Scholar] [CrossRef] [PubMed]
- Gagliardi, M.; Cotella, D.; Santoro, C.; Corà, D.; NA, B.; Piacentini, M.; Corazzari, M. Aldo-Keto Reductases Protect Metastatic Melanoma from ER Stress-Independent Ferroptosis. Cell Death Dis. 2019, 10, 902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagliardi, M.; Saverio, V.; Monzani, R.; Ferrari, E.; Piacentini, M.; Corazzari, M. Ferroptosis: A New Unexpected Chance to Treat Metastatic Melanoma? Cell Cycle 2020, 19, 2411–2425. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Kim, K.J.; Gaschler, M.M.; Patel, M.; Shchepinov, M.S.; Stockwell, B.R. Peroxidation of Polyunsaturated Fatty Acids by Lipoxygenases Drives Ferroptosis. Proc. Natl. Acad. Sci. USA 2016, 113, E4966–E4975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doll, S.; Proneth, B.; Tyurina, Y.Y.; Panzilius, E.; Kobayashi, S.; Ingold, I.; Irmler, M.; Beckers, J.; Aichler, M.; Walch, A.; et al. ACSL4 Dictates Ferroptosis Sensitivity by Shaping Cellular Lipid Composition. Nat. Chem. Biol. 2017, 13, 91–98. [Google Scholar] [CrossRef]
- Feng, H.; Schorpp, K.; Jin, J.; Yozwiak, C.E.; Hoffstrom, B.G.; Decker, A.M.; Rajbhandari, P.; Stokes, M.E.; Bender, H.G.; Csuka, J.M.; et al. Transferrin Receptor Is a Specific Ferroptosis Marker. Cell Rep. 2020, 30, 3411–3423.e7. [Google Scholar] [CrossRef] [Green Version]
- Frandsen, N.E.; Saugmann, S.; Marcussen, N. Acute Interstitial Nephritis Associated with the Use of Mesalazine in Inflammatory Bowel Disease. Nephron 2002, 92, 200–202. [Google Scholar] [CrossRef]
- Darb Emamie, A.; Rajabpour, M.; Ghanavati, R.; Asadolahi, P.; Farzi, S.; Sobouti, B.; Darbandi, A. The Effects of Probiotics, Prebiotics and Synbiotics on the Reduction of IBD Complications, a Periodic Review during 2009-2020. J. Appl. Microbiol. 2021, 130, 1823–1838. [Google Scholar] [CrossRef]
- Mentella, M.C.; Scaldaferri, F.; Pizzoferrato, M.; Gasbarrini, A.; Miggiano, G.A.D. Nutrition, IBD and Gut Microbiota: A Review. Nutrients 2020, 12, 944. [Google Scholar] [CrossRef] [Green Version]
- Iacomino, G.; Rotondi Aufiero, V.; Iannaccone, N.; Melina, R.; Giardullo, N.; de Chiara, G.; Venezia, A.; Taccone, F.S.; Iaquinto, G.; Mazzarella, G. IBD: Role of Intestinal Compartments in the Mucosal Immune Response. Immunobiology 2020, 225, 151849. [Google Scholar] [CrossRef]
- Leung, G.; Petri, B.; Reyes, J.L.; Wang, A.; Iannuzzi, J.; McKay, D.M. Cryopreserved Interleukin-4-Treated Macrophages Attenuate Murine Colitis in an Integrin Β7—Dependent Manner. Mol. Med. 2016, 21, 924–936. [Google Scholar] [CrossRef]
- Reardon, C.; Wang, A.; McKay, D.M. Transient Local Depletion of Foxp3+ Regulatory T Cells during Recovery from Colitis via Fas/Fas Ligand-Induced Death. J. Immunol. 2008, 180, 8316–8326. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.S. Epithelial ER Stress in Crohn’s Disease and Ulcerative Colitis. Inflamm. Bowel Dis. 2016, 22, 984–993. [Google Scholar] [CrossRef]
- Ma, X.; Dai, Z.; Sun, K.; Zhang, Y.; Chen, J.; Yang, Y.; Tso, P.; Wu, G.; Wu, Z. Intestinal Epithelial Cell Endoplasmic Reticulum Stress and Inflammatory Bowel Disease Pathogenesis: An Update Review. Front. Immunol. 2017, 8, 1271. [Google Scholar] [CrossRef] [Green Version]
- Corazzari, M.; Gagliardi, M.; Fimia, G.M.; Piacentini, M. Endoplasmic Reticulum Stress, Unfolded Protein Response, and Cancer Cell Fate. Front. Oncol. 2017, 7, 78. [Google Scholar] [CrossRef] [Green Version]
- Atreya, R.; Neurath, M.F.; Siegmund, B. Personalizing Treatment in IBD: Hype or Reality in 2020? Can We Predict Response to Anti-TNF? Front. Med. 2020, 7, 517. [Google Scholar] [CrossRef]
- Martyniak, A.; Medyńska-Przęczek, A.; Wędrychowicz, A.; Skoczeń, S.; Tomasik, P.J. Prebiotics, Probiotics, Synbiotics, Paraprobiotics and Postbiotic Compounds in IBD. Biomolecules 2021, 11, 1903. [Google Scholar] [CrossRef]
- Cuevas-González, P.F.; Liceaga, A.M.; Aguilar-Toalá, J.E. Postbiotics and Paraprobiotics: From Concepts to Applications. Food Res. Int. 2020, 136, 109502. [Google Scholar] [CrossRef]
- Mack, D.R.; Ahrne, S.; Hyde, L.; Wei, S.; Hollingsworth, M.A. Extracellular MUC3 Mucin Secretion Follows Adherence of Lactobacillus Strains to Intestinal Epithelial Cells in Vitro. Gut 2003, 52, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Ewaschuk, J.B.; Diaz, H.; Meddings, L.; Diederichs, B.; Dmytrash, A.; Backer, J.; van Langen, M.L.; Madsen, K.L. Secreted Bioactive Factors from Bifidobacterium Infantis Enhance Epithelial Cell Barrier Function. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G1025–G1034. [Google Scholar] [CrossRef]
- Jakubczyk, D.; Leszczyńska, K.; Górska, S. The Effectiveness of Probiotics in the Treatment of Inflammatory Bowel Disease (IBD)-A Critical Review. Nutrients 2020, 12, 1973. [Google Scholar] [CrossRef] [PubMed]
- Fries, W.; Muja, C.; Crisafulli, C.; Cuzzocrea, S.; Mazzon, E. Dynamics of Enterocyte Tight Junctions: Effect of Experimental Colitis and Two Different Anti-TNF Strategies. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G938–G947. [Google Scholar] [CrossRef] [PubMed]
- Landy, J.; Ronde, E.; English, N.; Clark, S.K.; Hart, A.L.; Knight, S.C.; Ciclitira, P.J.; Al-Hassi, H.O. Tight Junctions in Inflammatory Bowel Diseases and Inflammatory Bowel Disease Associated Colorectal Cancer. World J. Gastroenterol. 2016, 22, 3117–3126. [Google Scholar] [CrossRef] [PubMed]
- Marlow, G.J.; van Gent, D.; Ferguson, L.R. Why Interleukin-10 Supplementation Does Not Work in Crohn’s Disease Patients. World J. Gastroenterol. 2013, 19, 3931–3941. [Google Scholar] [CrossRef]
- Laroux, F.S.; Grisham, M.B. Immunological Basis of Inflammatory Bowel Disease: Role of the Microcirculation. Microcirculation 2001, 8, 283–301. [Google Scholar] [CrossRef]
- Kaser, A.; Martínez-Naves, E.; Blumberg, R.S. Endoplasmic Reticulum Stress: Implications for Inflammatory Bowel Disease Pathogenesis. Curr. Opin. Gastroenterol. 2010, 26, 318–326. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Lee, S.; Lee, H.; Ju, S.; Park, S.; Kwon, D.; Yoo, J.W.; Yoon, I.S.; Min, D.S.; Jung, Y.S.; et al. A Colon-Targeted Prodrug, 4-Phenylbutyric Acid-Glutamic Acid Conjugate, Ameliorates 2,4-Dinitrobenzenesulfonic Acid-Induced Colitis in Rats. Pharmaceutics 2020, 12, 843. [Google Scholar] [CrossRef]
- Gao, M.; Jiang, X. To Eat or Not to Eat—The Metabolic Flavor of Ferroptosis. Curr. Opin. Cell Biol. 2018, 51, 58–64. [Google Scholar] [CrossRef]
- Xu, S.; He, Y.; Lin, L.; Chen, P.; Chen, M.; Zhang, S. The Emerging Role of Ferroptosis in Intestinal Disease. Cell Death Dis. 2021, 12, 289. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ATF4_F | GTTTAGAGCTAGGCAGTGAAG |
| ATF4_R | CCTTTACACATGGAGGGATTAG |
| ATF6_F | GATGGTGACAACCAGAAAGA |
| ATF6_R | TGGAGGTGGAGGCATATAA |
| XBP1_F | AGTCCGCAGCAGGTG |
| XBP1_R | GGTCCAACTTGTCCAGAATG |
| CLD2_F | CCTCGCTGGCTTGTATTATC |
| CLD2_R | AAAGACTCCACCCACTACA |
| CLD15_F | GGGACCCTCCACATACTT |
| CLD15_R | CATACTTGGTTCCAGCATACA |
| OCL_F | TCTTTGGAGGAAGCCTAAAC |
| OCL_R | CTGCTCTTGGGTCTGTATATC |
| IFNγ_F | CCACATCTATGCCACTTGAG |
| IFNγ_R | CTCTTCCTCATGGCTGTTTC |
| TNFα_F | CCTATGTCTCAGCCTCTTCT |
| TNFα_R | GGGAACTTCTCATCCCTTTG |
| IL-10_F | TGAATTCCCTGGGTGAGA |
| IL-10_R | CCACTGCCTTGCTCTTATT |
| BAX_F | GGTTGCCCTCTTCTACTTTG |
| BAX_R | AGTGTCCAGCCCATGAT |
| PUMA_F | GGTTGCCCTCTTCTACTTTG |
| PUMA_R | AGTGTCCAGCCCATGAT |
| NOXA_F | GGTTGCCCTCTTCTACTTTG |
| NOXA_R | AGTGTCCAGCCCATGAT |
| CHAC1_F | TCACAGCACTGGCCTAT |
| CHAC1_R | CAAGGTTGTGACCAGAGAAG |
| PTGS2_F | GCCTGGTCTGATGATGTATG |
| PTGS2_R | GTCTGCTGGTTTGGAATAGT |
| ACSL4_F | TAAGCCCAXTTCAGACAAAC |
| ACSL4_R | GGCTACAGCATGGTCAAATA |
| TFR1_F | GGGCTATTGTAAGCGTGTAG |
| TFR1_R | CCTCTGTTTCCATGGTTTCT |
| GAPDH_F | TTCAACGGCACAGTCAAG |
| GAPDH_R | CCAGTAGACTCCACGACATA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monzani, R.; Gagliardi, M.; Clemente, N.; Saverio, V.; Pańczyszyn, E.; Santoro, C.; Yissachar, N.; Visciglia, A.; Pane, M.; Amoruso, A.; et al. The Gut-Ex-Vivo System (GEVS) Is a Dynamic and Versatile Tool for the Study of DNBS-Induced IBD in BALB/C and C57BL/6 Mice, Highlighting the Protective Role of Probiotics. Biology 2022, 11, 1574. https://doi.org/10.3390/biology11111574
Monzani R, Gagliardi M, Clemente N, Saverio V, Pańczyszyn E, Santoro C, Yissachar N, Visciglia A, Pane M, Amoruso A, et al. The Gut-Ex-Vivo System (GEVS) Is a Dynamic and Versatile Tool for the Study of DNBS-Induced IBD in BALB/C and C57BL/6 Mice, Highlighting the Protective Role of Probiotics. Biology. 2022; 11(11):1574. https://doi.org/10.3390/biology11111574
Chicago/Turabian StyleMonzani, Romina, Mara Gagliardi, Nausicaa Clemente, Valentina Saverio, Elżbieta Pańczyszyn, Claudio Santoro, Nissan Yissachar, Annalisa Visciglia, Marco Pane, Angela Amoruso, and et al. 2022. "The Gut-Ex-Vivo System (GEVS) Is a Dynamic and Versatile Tool for the Study of DNBS-Induced IBD in BALB/C and C57BL/6 Mice, Highlighting the Protective Role of Probiotics" Biology 11, no. 11: 1574. https://doi.org/10.3390/biology11111574