
A Modified Meiotic Recombination in Brassica napus Largely Improves Its Breeding Efficiency

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Flow Cytometry and Cytogenetic Studies
2.3. DNA Extraction and SNP Genotyping
2.4. Genetic Maps
2.5. Characterization of the B. rapa Introgressions
2.6. Localization of a QTL of Interest
2.7. Inferring the Position of Centromeric and Pericentromeric Regions
2.8. Statistical Analyses
3. Results
3.1. Impact of Brassica Hybrid Genomic Structure (Diploid, Allotriploid or Allotetraploid) on Homologous Recombination Frequency and Distribution
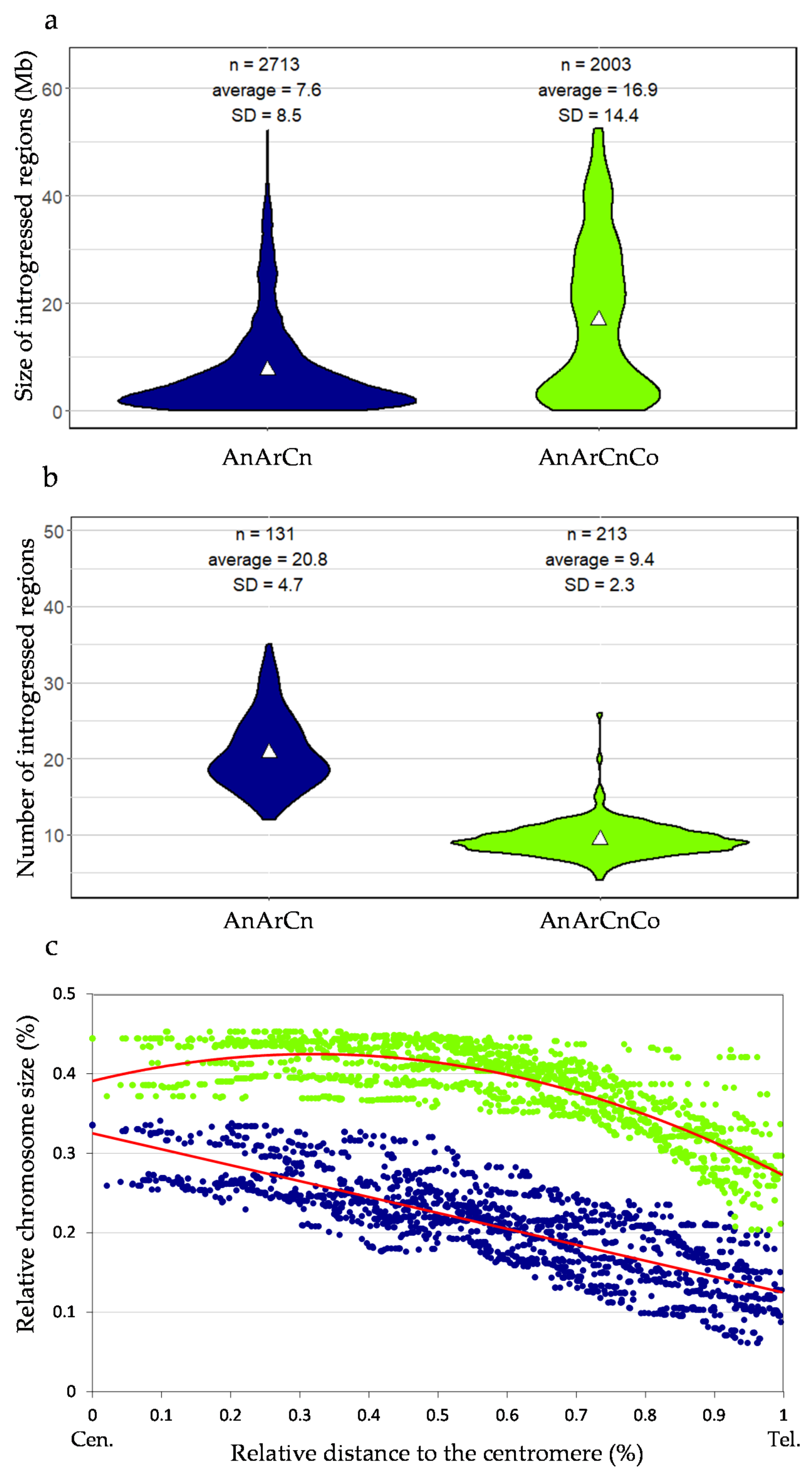
3.2. Impact of Recombination on the Size and Distribution of Introgression within Oilseed Rape
3.3. Interest in Using the Modified Recombination Pattern Oberved in AAC Allotriploid Hybrid to Reduce the Size of a QTL Present in a B. napus Pericentromeric Region
4. Discussion
4.1. Comparison of Breeding Strategies
4.2. Optimization of Breeding Strategies
4.3. Development of the Strategy on Other Models
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Felsenstein, J. The Evolutionary Advantage of Recombination. Genetics 1974, 78, 737–756. [Google Scholar] [CrossRef] [PubMed]
- Barton, N.H.; Charlesworth, B. Why Sex and Recombination? Science 1998, 281, 1986–1990. [Google Scholar] [CrossRef] [PubMed]
- Wijnker, E.; de Jong, H. Managing Meiotic Recombination in Plant Breeding. Trends Plant Sci. 2008, 13, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Mercier, R.; Mézard, C.; Jenczewski, E.; Macaisne, N.; Grelon, M. The Molecular Biology of Meiosis in Plants. Annu. Rev. Plant Biol. 2015, 66. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Moran, E.; Santos, J.-L.; Jones, G.H.; Franklin, F.C.H. ASY1 Mediates AtDMC1-Dependent Interhomolog Recombination during Meiosis in Arabidopsis. Genes Dev. 2007, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferdous, M.; Higgins, J.D.; Osman, K.; Lambing, C.; Roitinger, E.; Mechtler, K.; Armstrong, S.J.; Perry, R.; Pradillo, M.; Cuñado, N.; et al. Inter-Homolog Crossing-Over and Synapsis in Arabidopsis Meiosis Are Dependent on the Chromosome Axis Protein AtASY3. PLoS Genet. 2012, 8. [Google Scholar] [CrossRef] [Green Version]
- Crismani, W.; Girard, C.; Froger, N.; Pradillo, M.; Santos, J.L.; Chelysheva, L.; Copenhaver, G.P.; Horlow, C.; Mercier, R. FANCM Limits Meiotic Crossovers. Science 2012, 336, 1588–1590. [Google Scholar] [CrossRef] [Green Version]
- Girard, C.; Chelysheva, L.; Choinard, S.; Froger, N.; Macaisne, N.; Lehmemdi, A.; Mazel, J.; Crismani, W.; Mercier, R. AAA-ATPase FIDGETIN-LIKE 1 and Helicase FANCM Antagonize Meiotic Crossovers by Distinct Mechanisms. PLoS Genet. 2015, 11, e1005369. [Google Scholar] [CrossRef] [Green Version]
- Séguéla-Arnaud, M.; Crismani, W.; Larchevêque, C.; Mazel, J.; Froger, N.; Choinard, S.; Lemhemdi, A.; Macaisne, N.; Van Leene, J.; Gevaert, K.; et al. Multiple Mechanisms Limit Meiotic Crossovers: TOP3α and Two BLM Homologs Antagonize Crossovers in Parallel to FANCM. Proc. Natl. Acad. Sci. USA 2015, 112, 4713–4718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nageswaran, D.C.; Kim, J.; Lambing, C.; Kim, J.; Park, J.; Kim, E.-J.; Cho, H.S.; Kim, H.; Byun, D.; Park, Y.M.; et al. High Crossover Rate1 Encodes Protein Phosphatase X1 and Restricts Meiotic Crossovers in Arabidopsis. Nat. Plants 2021, 7, 452–467. [Google Scholar] [CrossRef]
- Saintenac, C.; Falque, M.; Martin, O.C.; Paux, E.; Feuillet, C.; Sourdille, P. Detailed Recombination Studies along Chromosome 3B Provide New Insights on Crossover Distribution in Wheat (Triticum Aestivum L.). Genetics 2009, 181. [Google Scholar] [CrossRef] [Green Version]
- Anderson, L.K.; Doyle, G.G.; Brigham, B.; Carter, J.; Hooker, K.D.; Lai, A.; Rice, M.; Stack, S.M. High-Resolution Crossover Maps for Each Bivalent of Zea Mays Using Recombination Nodules. Genetics 2003, 165, 849–865. [Google Scholar] [CrossRef] [PubMed]
- Felcher, K.J.; Coombs, J.J.; Massa, A.N.; Hansey, C.N.; Hamilton, J.P.; Veilleux, R.E.; Buell, C.R.; Douches, D.S. Integration of Two Diploid Potato Linkage Maps with the Potato Genome Sequence. PLoS ONE 2012, 7, e36347. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.; Zhao, X.; Kelly, K.A.; Venn, O.; Higgins, J.D.; Yelina, N.E.; Hardcastle, T.J.; Ziolkowski, P.A.; Copenhaver, G.P.; Franklin, F.C.H.; et al. Arabidopsis Meiotic Crossover Hotspots Overlap with H2A.Z Nucleosomes at Gene Promoters. Nat. Genet. 2013, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.; Henderson, I.R. Meiotic Recombination Hotspots—A Comparative View. Plant J. 2015, 83, 52–61. [Google Scholar] [CrossRef]
- Choi, K.; Zhao, X.; Tock, A.J.; Lambing, C.; Underwood, C.J.; Hardcastle, T.J.; Serra, H.; Kim, J.; Cho, H.S.; Kim, J.; et al. Nucleosomes and DNA Methylation Shape Meiotic DSB Frequency in Arabidopsis Thaliana Transposons and Gene Regulatory Regions. Genome Res. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marand, A.P.; Jansky, S.H.; Zhao, H.; Leisner, C.P.; Zhu, X.; Zeng, Z.; Crisovan, E.; Newton, L.; Hamernik, A.J.; Veilleux, R.E.; et al. Meiotic Crossovers Are Associated with Open Chromatin and Enriched with Stowaway Transposons in Potato. Genome Biol. 2017, 18, 203. [Google Scholar] [CrossRef] [Green Version]
- Colome-Tatche, M.; Cortijo, S.; Wardenaar, R.; Morgado, L.; Lahouze, B.; Sarazin, A.; Etcheverry, M.; Martin, A.; Feng, S.; Duvernois-Berthet, E.; et al. Features of the Arabidopsis Recombination Landscape Resulting from the Combined Loss of Sequence Variation and DNA Methylation. Proc. Natl. Acad. Sci. USA 2012, 109, 16240–16245. [Google Scholar] [CrossRef] [Green Version]
- Yelina, N.E.; Lambing, C.; Hardcastle, T.J.; Zhao, X.; Santos, B.; Henderson, I.R. DNA Methylation Epigenetically Silences Crossover Hot Spots and Controls Chromosomal Domains of Meiotic Recombination in Arabidopsis. Genes Dev. 2015, 29, 2183–2202. [Google Scholar] [CrossRef] [Green Version]
- Underwood, C.J.; Choi, K.; Lambing, C.; Zhao, X.; Serra, H.; Borges, F.; Simorowski, J.; Ernst, E.; Jacob, Y.; Henderson, I.R.; et al. Epigenetic Activation of Meiotic Recombination near Arabidopsis Thaliana Centromeres via Loss of H3K9me2 and Non-CG DNA Methylation. Genome Res. 2018, 28, 519–531. [Google Scholar] [CrossRef] [Green Version]
- Rowan, B.A.; Heavens, D.; Feuerborn, T.R.; Tock, A.J.; Henderson, I.R.; Weigel, D. An Ultra High-Density Arabidopsis Thaliana Crossover Map That Refines the Influences of Structural Variation and Epigenetic Features. Genetics 2019, 213, 771–787. [Google Scholar] [CrossRef] [Green Version]
- Smukowski, C.S.; Noor, M.A.F. Recombination Rate Variation in Closely Related Species. Heredity 2011, 107. [Google Scholar] [CrossRef] [Green Version]
- Haenel, Q.; Laurentino, T.G.; Roesti, M.; Berner, D. Meta-Analysis of Chromosome-Scale Crossover Rate Variation in Eukaryotes and Its Significance to Evolutionary Genomics. Mol. Ecol. 2018, 27, 2477–2497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M. Modification of Linkage Intensity by Natural Selection. Genetics 1967, 57, 625–641. [Google Scholar] [CrossRef]
- Feldman, M.; Liu, B.; Segal, G.; Abbo, S.; Levy, A.A.; Vega, J.M. Rapid Elimination of Low-Copy DNA Sequences in Polyploid Wheat: A Possible Mechanism for Differentiation of Homoeologous Chromosomes. Genetics 1997, 147, 1381–1387. [Google Scholar] [CrossRef]
- Coop, G.; Przeworski, M. An Evolutionary View of Human Recombination. Nat. Rev. Genet. 2007, 8, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Blary, A.; Jenczewski, E. Manipulation of Crossover Frequency and Distribution for Plant Breeding. Theor. Appl. Genet. 2019, 132, 575–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bomblies, K.; Higgins, J.D.; Yant, L. Meiosis Evolves: Adaptation to External and Internal Environments. New Phytol. 2015, 208, 306–323. [Google Scholar] [CrossRef]
- Drouaud, J.; Camilleri, C.; Bourguignon, P.-Y.; Canaguier, A.; Bérard, A.; Vezon, D.; Giancola, S.; Brunel, D.; Colot, V.; Prum, B.; et al. Variation in Crossing-over Rates across Chromosome 4 of Arabidopsis Thaliana Reveals the Presence of Meiotic Recombination “Hot Spots”. Genome Res. 2006, 16, 106–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraut, L.; Falque, M.; Drouaud, J.; Pereira, L.; Martin, O.C.; Mézard, C. Genome-Wide Crossover Distribution in Arabidopsis Thaliana Meiosis Reveals Sex-Specific Patterns along Chromosomes. PLoS Genet. 2011, 7, e1002354. [Google Scholar] [CrossRef] [Green Version]
- Bauer, E.; Falque, M.; Walter, H.; Bauland, C.; Camisan, C.; Campo, L.; Meyer, N.; Ranc, N.; Rincent, R.; Schipprack, W.; et al. Intraspecific Variation of Recombination Rate in Maize. Genome Biol. 2013, 14, R103. [Google Scholar] [CrossRef] [PubMed]
- Ziolkowski, P.A.; Underwood, C.J.; Lambing, C.; Martinez-Garcia, M.; Lawrence, E.J.; Ziolkowska, L.; Griffin, C.; Choi, K.; Franklin, F.C.H.; Martienssen, R.A.; et al. Natural Variation and Dosage of the HEI10 Meiotic E3 Ligase Control Arabidopsis Crossover Recombination. Genes Dev. 2017, 31, 306–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, E.J.; Gao, H.; Tock, A.J.; Lambing, C.; Blackwell, A.R.; Feng, X.; Henderson, I.R. Natural Variation in TBP-Associated Factor 4b Controls Meiotic Crossover and Germline Transcription in Arabidopsis. Curr. Biol. 2019, 29, 2676–2686.e3. [Google Scholar] [CrossRef] [PubMed]
- Pelé, A.; Rousseau-Gueutin, M.; Chèvre, A.-M. Speciation Success of Polyploid Plants Closely Relates to the Regulation of Meiotic Recombination. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Leflon, M.; Grandont, L.; Eber, F.; Huteau, V.; Coriton, O.; Chelysheva, L.; Jenczewski, E.; Chèvre, A.-M. Crossovers Get a Boost in Brassica Allotriploid and Allotetraploid Hybrids. Plant Cell 2010, 22, 2253–2264. [Google Scholar] [CrossRef] [Green Version]
- Pelé, A.; Falque, M.; Trotoux, G.; Eber, F.; Nègre, S.; Gilet, M.; Huteau, V.; Lodé, M.; Jousseaume, T.; Dechaumet, S.; et al. Amplifying Recombination Genome-Wide and Reshaping Crossover Landscapes in Brassicas. PLoS Genet. 2017, 13, e1006794. [Google Scholar] [CrossRef] [PubMed]
- Leflon, M.; Eber, F.; Letanneur, J.C.; Chelysheva, L.; Coriton, O.; Huteau, V.; Ryder, C.D.; Barker, G.; Jenczewski, E.; Chèvre, A.M. Pairing and Recombination at Meiosis of Brassica Rapa (AA) × Brassica Napus (AACC) Hybrids. Theor. Appl. Genet. Theor. Angew. Genet. 2006, 113, 1467–1480. [Google Scholar] [CrossRef]
- Snowdon, R.J.; Abbadi, A.; Kox, T.; Schmutzer, T.; Leckband, G. Heterotic Haplotype Capture: Precision Breeding for Hybrid Performance. Trends Plant Sci. 2015, 20, 410–413. [Google Scholar] [CrossRef]
- Tourrette, E.; Bernardo, R.; Falque, M.; Martin, O.C. Assessing by Modeling the Consequences of Increased Recombination in Recurrent Selection of Oryza Sativa and Brassica Rapa. G3 Genes Genomes Genet. 2019, 9, 4169–4181. [Google Scholar] [CrossRef] [Green Version]
- Tourrette, E.; Falque, M.; Martin, O.C. Enhancing Backcross Programs through Increased Recombination. Genet. Sel. Evol. 2021, 53, 25. [Google Scholar] [CrossRef]
- Kumar, V.; Paillard, S.; Fopa-Fomeju, B.; Falentin, C.; Deniot, G.; Baron, C.; Vallée, P.; Manzanares-Dauleux, M.J.; Delourme, R. Multi-Year Linkage and Association Mapping Confirm the High Number of Genomic Regions Involved in Oilseed Rape Quantitative Resistance to Blackleg. Theor. Appl. Genet. 2018, 131, 1627–1643. [Google Scholar] [CrossRef] [PubMed]
- Pelé, A.; Trotoux, G.; Eber, F.; Lodé, M.; Gilet, M.; Deniot, G.; Falentin, C.; Nègre, S.; Morice, J.; Rousseau-Gueutin, M.; et al. The Poor Lonesome A Subgenome of Brassica Napus Var. Darmor (AACC) May Not Survive without Its Mate. New Phytol. 2017, 213, 1886–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szadkowski, E.; Eber, F.; Huteau, V.; Lodé, M.; Huneau, C.; Belcram, H.; Coriton, O.; Manzanares-Dauleux, M.J.; Delourme, R.; King, G.J.; et al. The First Meiosis of Resynthesized Brassica Napus, a Genome Blender. New Phytol. 2010, 186, 102–112. [Google Scholar] [CrossRef]
- Howell, E.C.; Barker, G.C.; Jones, G.H.; Kearsey, M.J.; King, G.J.; Kop, E.P.; Ryder, C.D.; Teakle, G.R.; Vicente, J.G.; Armstrong, S.J. Integration of the Cytogenetic and Genetic Linkage Maps of Brassica Oleracea. Genetics 2002, 161, 1225–1234. [Google Scholar] [CrossRef]
- Wang, J.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.-H.; Bancroft, I.; Cheng, F.; Huang, S.; Li, X.; Hua, W.; et al. The Genome of the Mesopolyploid Crop Species Brassica Rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousseau-Gueutin, M.; Belser, C.; Da Silva, C.; Richard, G.; Istace, B.; Cruaud, C.; Falentin, C.; Boideau, F.; Boutte, J.; Delourme, R.; et al. Long-Read Assembly of the Brassica Napus Reference Genome Darmor-Bzh. GigaScience 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- De Givry, S.; Bouchez, M.; Chabrier, P.; Milan, D.; Schiex, T. Carhta Gene: Multipopulation Integrated Genetic and Radiation Hybrid Mapping. Bioinformatics 2005, 21, 1703–1704. [Google Scholar] [CrossRef] [Green Version]
- Kosambi, D.D. The Estimation of Map Distances from Recombination Values. Ann. Eugen. 1943, 12, 172–175. [Google Scholar] [CrossRef]
- Lim, K.-B.; de Jong, H.; Yang, T.-J.; Park, J.-Y.; Kwon, S.-J.; Kim, J.S.; Lim, M.-H.; Kim, J.A.; Jin, M.; Jin, Y.-M.; et al. Characterization of RDNAs and Tandem Repeats in the Heterochromatin of Brassica Rapa. Mol. Cells 2005, 19, 436–444. [Google Scholar]
- Lim, K.-B.; Yang, T.-J.; Hwang, Y.-J.; Kim, J.S.; Park, J.-Y.; Kwon, S.-J.; Kim, J.; Choi, B.-S.; Lim, M.-H.; Jin, M.; et al. Characterization of the Centromere and Peri-Centromere Retrotransposons in Brassica Rapa and Their Distribution in Related Brassica Species. Plant J. 2007, 49, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Yelina, N.E.; Choi, K.; Chelysheva, L.; Macaulay, M.; de Snoo, B.; Wijnker, E.; Miller, N.; Drouaud, J.; Grelon, M.; Copenhaver, G.P.; et al. Epigenetic Remodeling of Meiotic Crossover Frequency in Arabidopsis Thaliana DNA Methyltransferase Mutants. PLoS Genet. 2012, 8, e1002844. [Google Scholar] [CrossRef] [Green Version]
- Mason, A.S.; Rousseau-Gueutin, M.; Morice, J.; Bayer, P.E.; Besharat, N.; Cousin, A.; Pradhan, A.; Parkin, I.A.P.; Chevre, A.-M.; Batley, J.; et al. Centromere Locations in Brassica A and C Genomes Revealed Through Half-Tetrad Analysis. Genetics 2016, 202, 513–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Z.; Pires, J.C. Karyotype and Identification of All Homoeologous Chromosomes of Allopolyploid Brassica Napus and Its Diploid Progenitors. Genetics 2011, 187. [Google Scholar] [CrossRef] [Green Version]
- Song, K.; Lu, P.; Tang, K.; Osborn, T.C. Rapid Genome Change in Synthetic Polyploids of Brassica and Its Implications for Polyploid Evolution. Proc. Natl. Acad. Sci. USA 1995, 92, 7719–7723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaeta, R.T.; Pires, J.C.; Iniguez-Luy, F.; Leon, E.; Osborn, T.C. Genomic Changes in Resynthesized Brassica Napus and Their Effect on Gene Expression and Phenotype. Plant Cell 2007, 19. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Z.; Gaeta, R.T.; Pires, J.C. Homoeologous Shuffling and Chromosome Compensation Maintain Genome Balance in Resynthesized Allopolyploid Brassica Napus. Proc. Natl. Acad. Sci. USA 2011, 108, 7908–7913. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Z.; Gaeta, R.T.; Edger, P.P.; Cao, Y.; Zhao, K.; Zhang, S.; Pires, J.C. Chromosome Inheritance and Meiotic Stability in Allopolyploid Brassica Napus. G3 Genes Genomes Genet. 2021, 11. [Google Scholar] [CrossRef]
- Rousseau-Gueutin, M.; Morice, J.; Coriton, O.; Huteau, V.; Trotoux, G.; Nègre, S.; Falentin, C.; Deniot, G.; Gilet, M.; Eber, F.; et al. The Impact of Open Pollination on the Structural Evolutionary Dynamics, Meiotic Behavior, and Fertility of Resynthesized Allotetraploid Brassica Napus L. G3 Genes Genomes Genet. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Ferreira De Carvalho, J.; Stoeckel, S.; Eber, F.; Lodé-Taburel, M.; Gilet, M.-M.; Trotoux, G.; Morice, J.; Falentin, C.; Chèvre, A.-M.; Rousseau-Gueutin, M. Untangling Structural Factors Driving Genome Stabilization in Nascent Brassica Napus Allopolyploids. New Phytol. 2021, 230, 2072–2084. [Google Scholar] [CrossRef]
- Mason, A.S.; Snowdon, R.J. Oilseed Rape: Learning about Ancient and Recent Polyploid Evolution from a Recent Crop Species. Plant Biol. 2016, 18, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Kittipol, V.; He, Z.; Wang, L.; Doheny-Adams, T.; Langer, S.; Bancroft, I. Genetic Architecture of Glucosinolate Variation in Brassica Napus. J. Plant Physiol. 2019, 240. [Google Scholar] [CrossRef] [PubMed]
- Pires, J.C.; Zhao, J.; Schranz, M.E.; Leon, E.J.; Quijada, P.A.; Lukens, L.N.; Osborn, T.C. Flowering Time Divergence and Genomic Rearrangements in Resynthesized Brassica Polyploids (Brassicaceae). Biol. J. Linn. Soc. 2004, 82, 675–688. [Google Scholar] [CrossRef]
- Zhao, J.; Udall, J.A.; Quijada, P.A.; Grau, C.R.; Meng, J.; Osborn, T.C. Quantitative Trait Loci for Resistance to Sclerotinia Sclerotiorum and Its Association with a Homeologous Non-Reciprocal Transposition in Brassica Napus L. Theor. Appl. Genet. 2006, 112, 509–516. [Google Scholar] [CrossRef]
- Stein, A.; Coriton, O.; Rousseau-Gueutin, M.; Samans, B.; Schiessl, S.V.; Obermeier, C.; Parkin, I.A.P.; Chèvre, A.-M.; Snowdon, R.J. Mapping of Homoeologous Chromosome Exchanges Influencing Quantitative Trait Variation in Brassica Napus. Plant Biotechnol. J. 2017, 15, 1478–1489. [Google Scholar] [CrossRef] [Green Version]
- Diederichsen, E.; Sacristan, M.D. Disease Response of Resynthesized Brassica Napus L. Lines Carrying Different Combinations of Resistance to Plasmodiophora Brassicae Wor. Plant Breed. 1996, 115, 5–10. [Google Scholar] [CrossRef]
- Diederichsen, E.; Beckmann, J.; Schondelmeier, J.; Dreyer, F. Genetics of Clubroot Resistance in Brassica Napus “Mendel”. Acta Hortic. 2006, 706, 307–311. [Google Scholar] [CrossRef]
- Qian, W.; Chen, X.; Fu, D.; Zou, J.; Meng, J. Intersubgenomic Heterosis in Seed Yield Potential Observed in a New Type of Brassica Napus Introgressed with Partial Brassica Rapa Genome. Theor. Appl. Genet. Theor. Angew. Genet. 2005, 110. [Google Scholar] [CrossRef] [PubMed]
- Attri, R.; Rahman, H. Introgression of Allelic Diversity from Genetically Distinct Variants of Brassica Rapa into Brassica Napus Canola and Inheritance of the B. Rapa Alleles. Crop Pasture Sci. 2017, 69, 94–106. [Google Scholar] [CrossRef]
- Pan, Q.; Zhu, B.; Zhang, D.; Tong, C.; Ge, X.; Liu, S.; Li, Z. Gene Expression Changes During the Allo-/Deallopolyploidization Process of Brassica Napus. Front. Genet. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Mao, L.; Qiu, J.; Wang, M.; Jia, L.; Wu, D.; He, Z.; Chen, M.; Shen, Y.; Shen, E.; et al. Genome-Wide Selection Footprints and Deleterious Variations in Young Asian Allotetraploid Rapeseed. Plant Biotechnol. J. 2019, 17, 1998–2010. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, A.; Liang, F.; Yao, X.; Wang, Y.; Liu, X.; Zhang, Y.; Dalelhan, J.; Zhang, B.; Qin, M.; et al. Screening of Clubroot-Resistant Varieties and Transfer of Clubroot Resistance Genes to Brassica Napus Using Distant Hybridization. Breed. Sci. 2018, 68, 258–267. [Google Scholar] [CrossRef] [Green Version]
- Balesdent, M.; Fudal, I.; Ollivier, B.; Bally, P.; Grandaubert, J.; Eber, F.; Chèvre, A.; Leflon, M.; Rouxel, T. The Dispensable Chromosome of Leptosphaeria Maculans Shelters an Effector Gene Conferring Avirulence towards Brassica Rapa. New Phytol. 2013, 198, 887–898. [Google Scholar] [CrossRef]
- Delourme, R.; Laperche, A.; Bouchet, A.-S.; Jubault, M.; Paillard, S.; Manzanares-Dauleux, M.-J.; Nesi, N. Genes and quantitative trait loci mapping for major agronomic traits in Brassica napus L. In The Brassica napus Genome; Liu, S., Snowdon, R., Chalhoub, B., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 41–85. ISBN 978-3-319-43692-0. [Google Scholar]
- Song, J.-M.; Guan, Z.; Hu, J.; Guo, C.; Yang, Z.; Wang, S.; Liu, D.; Wang, B.; Lu, S.; Zhou, R.; et al. Eight High-Quality Genomes Reveal Pan-Genome Architecture and Ecotype Differentiation of Brassica Napus. Nat. Plants 2020, 6, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Downey, R.K.; Klassen, A.J.; Stringam, G.R. Rapeseed and mustard. In Hybridization of Crop Plants; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1980; pp. 495–509. ISBN 978-0-89118-566-6. [Google Scholar]
- Nikzad, A.; Kebede, B.; Pinzon, J.; Bhavikkumar, J.; Wang, X.; Yang, R.-C.; Rahman, H. Potential of the C Genome of the Different Variants of Brassica Oleracea for Heterosis in Spring B. Napus Canola. Front. Plant Sci. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Chiang, M.S.; Chiang, B.Y.; Grant, W.F. Transfer of Resistance to Race 2 of Plasmodiophora Brassicae from Brassica Napus to Cabbage (B. Oleracea Var. Capitata). I. Interspecific Hybridization between B. Napus and B. Oleracea Var. Capitata. Euphytica 1977, 26, 319–336. [Google Scholar] [CrossRef]
- Yu, H.; Li, Z.; Ren, W.; Han, F.; Yang, L.; Zhuang, M.; Lv, H.; Liu, Y.; Fang, Z.; Zhang, Y. Creation of Fertility-Restored Materials for Ogura CMS in Brassica Oleracea by Introducing Rfo Gene from Brassica Napus via an Allotriploid Strategy. Theor. Appl. Genet. 2020, 133, 2825–2837. [Google Scholar] [CrossRef] [PubMed]
- Nagaharu, U. Genome Analysis in Brassica with Special Reference to the Experimental Formation of B. Napus and Peculiar Mode of Fertilization. Jpn. J. Bot. 1935, 7, 389–452. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boideau, F.; Pelé, A.; Tanguy, C.; Trotoux, G.; Eber, F.; Maillet, L.; Gilet, M.; Lodé-Taburel, M.; Huteau, V.; Morice, J.; et al. A Modified Meiotic Recombination in Brassica napus Largely Improves Its Breeding Efficiency. Biology 2021, 10, 771. https://doi.org/10.3390/biology10080771
Boideau F, Pelé A, Tanguy C, Trotoux G, Eber F, Maillet L, Gilet M, Lodé-Taburel M, Huteau V, Morice J, et al. A Modified Meiotic Recombination in Brassica napus Largely Improves Its Breeding Efficiency. Biology. 2021; 10(8):771. https://doi.org/10.3390/biology10080771
Chicago/Turabian StyleBoideau, Franz, Alexandre Pelé, Coleen Tanguy, Gwenn Trotoux, Frédérique Eber, Loeiz Maillet, Marie Gilet, Maryse Lodé-Taburel, Virginie Huteau, Jérôme Morice, and et al. 2021. "A Modified Meiotic Recombination in Brassica napus Largely Improves Its Breeding Efficiency" Biology 10, no. 8: 771. https://doi.org/10.3390/biology10080771