
Leptospira fainei Detected in Testicles and Epididymis of Wild Boar (Sus scrofa)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples Collection
2.2. Leptospira spp. Isolation
2.3. Multiplex Real-Time PCR and Genotyping of Leptospira spp.
2.4. Statistical Analysis
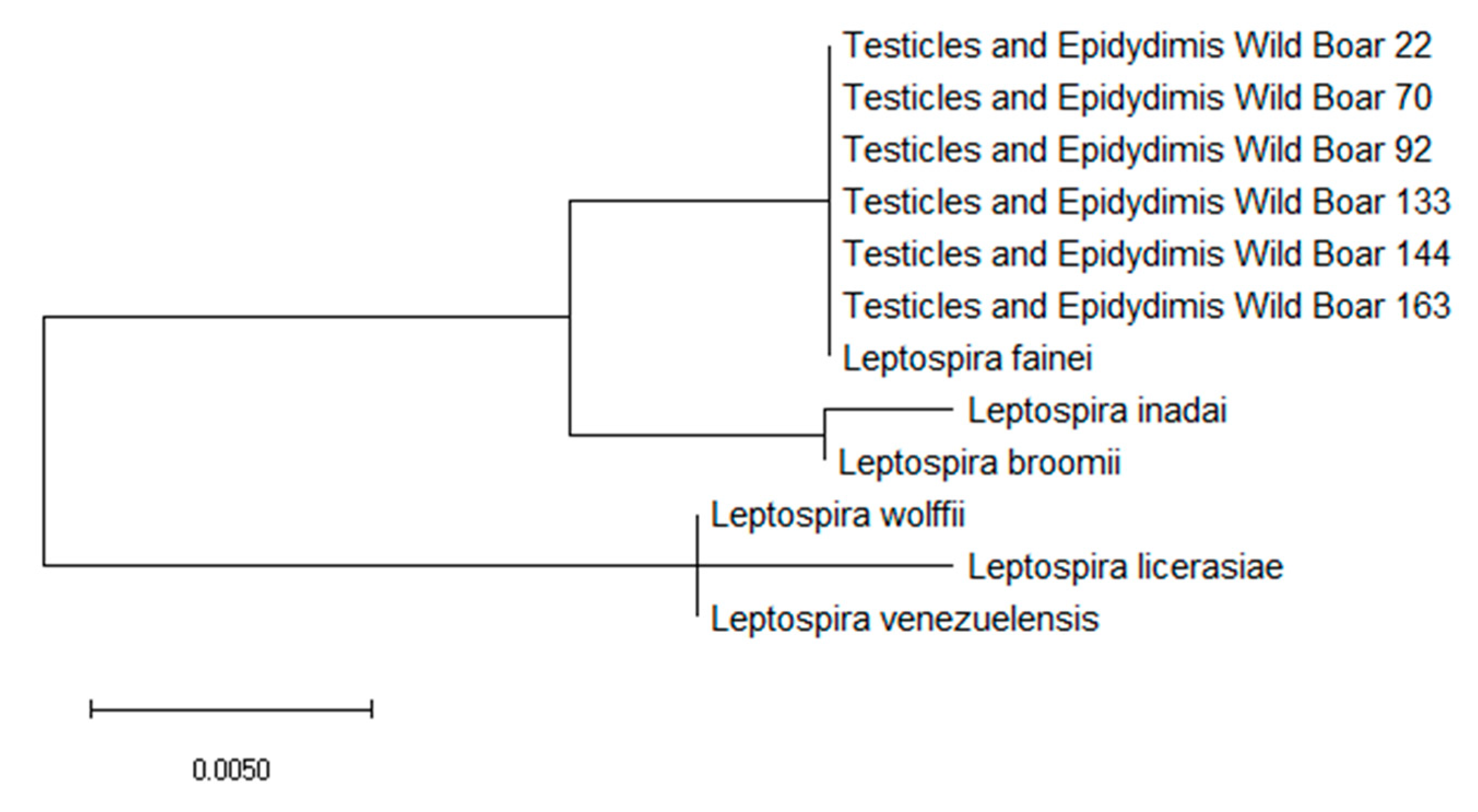
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Levett, P.N. Leptospirosis. Clin. Microbiol. Rev. 2001, 14, 296–326. [Google Scholar] [CrossRef] [Green Version]
- Adler, B. Leptospira and Leptospirosis. In Current Topics in Microbiology and Immunology; Springer: Cham, Switzerland, 2015; ISBN 978-3-662-45059-8. [Google Scholar]
- Thibeaux, R.; Girault, D.; Bierque, E.; Soupé-Gilbert, M.-E.; Rettinger, A.; Douyère, A.; Meyer, M.; Iraola, G.; Picardeau, M.; Goarant, C. Biodiversity of Environmental Leptospira: Improving Identification and Revisiting the Diagnosis. Front. Microbiol. 2018, 9, 816. [Google Scholar] [CrossRef] [Green Version]
- Barragan, V.; Chiriboga, J.; Miller, E.; Olivas, S.; Birdsell, D.; Hepp, C.; Hornstra, H.; Schupp, J.M.; Morales, M.; Gonzalez, M.; et al. High Leptospira Diversity in Animals and Humans Complicates the Search for Common Reservoirs of Human Disease in Rural Ecuador. PLoS Negl. Trop. Dis. 2016, 10, e0004990. [Google Scholar] [CrossRef]
- Balamurugan, V.; Gangadhar, N.L.; Mohandoss, N.; Thirumalesh, S.R.A.; Dhar, M.; Shome, R.; Krishnamoorthy, P.; Prabhudas, K.; Rahman, H. Characterization of leptospira isolates from animals and humans: Phylogenetic analysis identifies the prevalence of intermediate species in India. Springerplus 2013, 2, 362. [Google Scholar] [CrossRef] [Green Version]
- Cerqueira, G.M.; Picardeau, M. A century of Leptospira strain typing. Infect. Genet. Evol. 2009, 9, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.T.; Schiettekatte, O.; Goarant, C.; Neela, V.K.; Bernet, E.; Thibeaux, R.; Ismail, N.; Mohd Khalid, M.K.N.; Amran, F.; Masuzawa, T.; et al. Revisiting the taxonomy and evolution of pathogenicity of the genus Leptospira through the prism of genomics. PLoS Negl. Trop. Dis. 2019, 13, e0007270. [Google Scholar] [CrossRef] [Green Version]
- Bourhy, P.; Vray, M.; Picardeau, M. Evaluation of an in-house ELISA using the intermediate species Leptospira fainei for diagnosis of leptospirosis. J. Med. Microbiol. 2013, 62, 822–827. [Google Scholar] [CrossRef]
- Perolat, P.; Chappel, R.J.; Adler, B.; Baranton, G.; Bulach, D.M.; Billinghurst, M.L.; Letocart, M.; Merien, F.; Serrano, M.S. Leptospira fainei sp. nov., isolated from pigs in Australia. Int. J. Syst. Bacteriol. 1998, 48, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Chappel, R.J.; Khalik, D.A.; Adler, B.; Bulach, D.M.; Faine, S.; Perolat, P.; Vallance, V. Serological titres to Leptospira fainei serovar hurstbridge in human sera in Australia. Epidemiol. Infect. 1998, 121, 473–475. [Google Scholar] [CrossRef] [PubMed]
- Yersin, C.; Bovet, P.; Mérien, F.; Wong, T.; Panowsky, J.; Perolat, P. Human leptospirosis in the Seychelles (Indian ocean): A population- based study. Am. J. Trop. Med. Hyg. 1998, 59, 933–940. [Google Scholar] [CrossRef] [Green Version]
- Petersen, A.M.; Boye, K.; Blom, J.; Schlichting, P.; Krogfelt, K.A. First isolation of Leptospira fainei serovar Hurstbridge from two human patients with Weil’s syndrome. J. Med. Microbiol. 2001, 50, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Arzouni, J.P.; Parola, P.; La Scola, B.; Postic, D.; Brouqui, P.; Raoult, D. Human infection caused by Leptospira fainei. Emerg. Infect. Dis. 2002, 8, 865–868. [Google Scholar] [CrossRef]
- Matthias, M.A.; Ricaldi, J.N.; Cespedes, M.; Diaz, M.M.; Galloway, R.L.; Saito, M.; Steigerwalt, A.G.; Patra, K.P.; Ore, C.V.; Gotuzzo, E.; et al. Human Leptospirosis Caused by a New, Antigenically Unique Leptospira Associated with a Rattus Species Reservoir in the Peruvian Amazon. PLoS Negl. Trop. Dis. 2008, 2, e213. [Google Scholar] [CrossRef] [Green Version]
- Chiriboga, J.; Barragan, V.; Arroyo, G.; Sosa, A.; Birdsell, D.N.; España, K.; Mora, A.; Espín, E.; Mejía, M.E.; Morales, M.; et al. High Prevalence of Intermediate Leptospira spp. DNA in Febrile Humans from Urban and Rural Ecuador. Emerg. Infect. Dis. 2015, 21, 2141–2147. [Google Scholar] [CrossRef] [Green Version]
- Ganoza, C.A.; Matthias, M.A.; Saito, M.; Cespedes, M.; Gotuzzo, E.; Vinetz, J.M. Asymptomatic Renal Colonization of Humans in the Peruvian Amazon by Leptospira. PLoS Negl. Trop. Dis. 2010, 4, e612. [Google Scholar] [CrossRef] [PubMed]
- Matthias, M.A.; Mónica Díaz, M.; Campos, K.J.; Calderon, M.; Willig, M.R.; Pacheco, V.; Gotuzzo, E.; Gilman, R.H.; Vinetz, J.M. Diversity of Bat-Associated Leptospira In The Peruvian Amazon Inferred By Bayesian Phylogenetic Analysis Of 16s Ribosomal Dna Sequences. Am. J. Trop. Med. Hyg. 2005, 73, 964–974. [Google Scholar] [CrossRef]
- Cilia, G.; Bertelloni, F.; Mignone, W.; Spina, S.; Berio, E.; Razzuoli, E.; Vencia, W.; Franco, V.; Cecchi, F.; Bogi, S.; et al. Molecular detection of Leptospira spp. in wild boar (Sus scrofa) hunted in Liguria region (Italy). Comp. Immunol. Microbiol. Infect. Dis. 2020, 68, 101410. [Google Scholar] [CrossRef]
- Cilia, G.; Bertelloni, F.; Angelini, M.; Cerri, D.; Fratini, F. Leptospira Survey in Wild Boar (Sus scrofa) Hunted in Tuscany, Central Italy. Pathogens 2020, 9, 377. [Google Scholar] [CrossRef]
- Massei, G.; Kindberg, J.; Licoppe, A.; Gačić, D.; Šprem, N.; Kamler, J.; Baubet, E.; Hohmann, U.; Monaco, A.; Ozoliņš, J.; et al. Wild boar populations up, numbers of hunters down? A review of trends and implications for Europe. Pest Manag. Sci. 2015, 71, 492–500. [Google Scholar] [CrossRef]
- Santilli, F.; Varuzza, P. Factors affecting wild boar (Sus scrofa) abundance in southern Tuscany. Hystrix, Ital. J. Mammal. 2013, 24, 169–173. [Google Scholar]
- Castillo-Contreras, R.; Carvalho, J.; Serrano, E.; Mentaberre, G.; Fernández-Aguilar, X.; Colom, A.; González-Crespo, C.; Lavín, S.; López-Olvera, J.R. Urban wild boars prefer fragmented areas with food resources near natural corridors. Sci. Total Environ. 2018, 615, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Bertelloni, F.; Mazzei, M.; Cilia, G.; Forzan, M.; Felicioli, A.; Sagona, S.; Bandecchi, P.; Turchi, B.; Cerri, D.; Fratini, F. Serological Survey on Bacterial and Viral Pathogens in Wild Boars Hunted in Tuscany. Ecohealth 2020, 17, 85–93. [Google Scholar] [CrossRef]
- Bertelloni, F.; Cilia, G.; Turchi, B.; Pinzauti, P.; Cerri, D.; Fratini, F. Epidemiology of leptospirosis in North-Central Italy: Fifteen years of serological data (2002–2016). Comp. Immunol. Microbiol. Infect. Dis. 2019, 65, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Montagnaro, S.; Sasso, S.; De Martino, L.; Longo, M.; Iovane, V.; Ghiurmino, G.; Pisanelli, G.; Nava, D.; Baldi, L.; Pagnini, U. Prevalence of Antibodies to Selected Viral and Bacterial Pathogens in Wild Boar (Sus scrofa) in Campania Region, Italy. J. Wildl. Dis. 2010, 46, 316–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- San-Miguel Ayanz, J.M.; Garcia-Peña, F.J.; García-Lunar, P.; Ortega-Mora, L.M.; Ruano, M.J.; Álvarez-García, G.; Collantes-Fernández, E. Seroprevalence of Leptospirosis, Brucellosis, and Q Fever in a Wild Red Deer (Cervus elaphus) Population Kept in a Fenced Reserve in Absence of Contact with Livestock. Vector-Borne Zoonotic Dis. 2017, 17, 692–697. [Google Scholar] [CrossRef] [PubMed]
- Boqvist, S.; Bergström, K.; Magnusson, U. Prevalence of antibody to six Leptospira Servovars in Swedish wild boars. J. Wildl. Dis. 2012, 48, 492–496. [Google Scholar] [CrossRef] [Green Version]
- Vicente, J.; León-Vizcaíno, L.; Gortázar, C.; Cubero, M.J.; González, M.; Martín-Atance, P. Antibodies to selected viral and bacterial pathogens in European wild boars from southcentral Spain. J. Wildl. Dis. 2002, 38, 649–652. [Google Scholar] [CrossRef] [Green Version]
- Żmudzki, J.; Jabłoński, A.; Nowak, A.; Zębek, S.; Arent, Z.; Bocian, Ł.; Pejsak, Z. First overall report of Leptospira infections in wild boars in Poland. Acta Vet. Scand. 2016, 58, 3. [Google Scholar] [CrossRef] [Green Version]
- Vale-Goncalves, H.M.; Cabral, J.A.; Faria, M.C.; Nunes-Pereira, M.; Faria, A.S.; Veloso, O.; Vieira, M.L.; Paiva-Cardoso, M.N. Prevalence of Leptospira antibodies in wild boars ( Sus scrofa ) from Northern Portugal: Risk factor analysis. Epidemiol. Infect. 2015, 143, 2126–2130. [Google Scholar] [CrossRef] [Green Version]
- Ellis, W.A. Animal Leptospirosis. Curr. Top. Microbiol. Immunol. 2015, 387, 99–137. [Google Scholar]
- Ellis, W.A.; McParland, P.J.; Bryson, D.G.; Cassells, J.A. Boars as carriers of leptospires of the Australis serogroup on farms with an abortion problem. Vet. Rec. 1986, 118, 563. [Google Scholar] [CrossRef] [PubMed]
- Ellis, W.A.; Songer, J.G.; Montgomery, J.; Cassells, J.A. Prevalence of Leptospira interrogans serovar hardjo in the genital and urinary tracts of non-pregnant cattle. Vet. Rec. 1986, 118, 11–13. [Google Scholar] [CrossRef]
- Ellis, W.A.; McParland, P.J.; Bryson, D.G.; Cassells, J.A. Prevalence of Leptospira infection in aborted pigs in Northern Ireland. Vet. Rec. 1986, 118, 63–65. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, A.P.; Lilenbaum, W. Genital bovine leptospirosis: A new look for an old disease. Theriogenology 2020, 141, 41–47. [Google Scholar] [CrossRef]
- Dhaliwal, G.S.; Murray, R.D.; Dobson, H.; Montgomery, J.; Ellis, W.A.; Baker, J.R. Presence of antigen and antibodies in serum and genital discharges of heifers after experimental intrauterine inoculation with Leptospira interrogans serovar hardjo. Res. Vet. Sci. 1996, 60, 157–162. [Google Scholar] [CrossRef]
- Strutzberg-Minder, K.; Tschentscher, A.; Beyerbach, M.; Homuth, M.; Kreienbrock, L. Passive surveillance of Leptospira infection in swine in Germany. Porc. Heal. Manag. 2018, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Ellis, W.A.; McParland, P.J.; Bryson, D.G.; Thiermann, A.B.; Montgomery, J. Isolation of leptospires from the genital tract and kidneys of aborted sows. Vet. Rec. 1986, 118, 294–295. [Google Scholar] [CrossRef] [PubMed]
- Brihuega, B.; Loffler, S.G.; Samartino, L.; Romero, G.; Auteri, C.; Martinez, M. First Isolation of Leptospira borgpetersenii from Fetuses of Wild Boars (Sus scrofa). Electron. J. Biol. 2017, 13, 63–66. [Google Scholar]
- Cilia, G.; Bertelloni, F.; Piredda, I.; Ponti, M.N.; Turchi, B.; Cantinle, C.; Parisi, F.; Pinzauti, P.; Armani, A.; Palmas, B.; et al. Presence of pathogenic Leptospira spp. in the reproductive system and fetuses of wild boars (Sus scrofa) in Italy. PLoS Negl. Trop. Dis. 2020, 14, e0008982. [Google Scholar] [CrossRef]
- Sáez-Royuela, C.; Gomariz, R.P.; Luis Tellería, J. Age Determination of European Wild Boar. Wildl. Soc. Bull. 1989, 17, 326–329. [Google Scholar]
- Bedir, O.; Kilic, A.; Atabek, E.; Kuskucu, A.M.; Turhan, V.; Basustaoglu, A.C. Simultaneous detection and differentiation of pathogenic and nonpathogenic Leptospira spp. by multiplex real-time PCR (TaqMan) assay. Polish J. Microbiol. 2010, 59, 167–173. [Google Scholar] [CrossRef]
- Ahmed, N.; Devi, S.M.; de los Á Valverde, M.; Vijayachari, P.; Machang’u, R.S.; Ellis, W.A.; Hartskeerl, R.A. Multilocus sequence typing method for identification and genotypic classification of pathogenic Leptospira species. Ann. Clin. Microbiol. Antimicrob. 2006, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Givens, M.D. Review: Risks of disease transmission through semen in cattle. Animal 2018, 12, s165–s171. [Google Scholar] [CrossRef] [Green Version]
- Bielanski, A.; Surujballi, O.; Thomas, E.G.; Tanaka, E. Sanitary status of oocytes and embryos collected from heifers experimentally exposed to Leptospira borgpetersenii serovar hardjobovis. Anim. Reprod. Sci. 1998, 54, 65–73. [Google Scholar] [CrossRef]
- Lilenbaum, W.; Varges, R.; Brandão, F.Z.; Cortez, A.; de Souza, S.O.; Brandão, P.E.; Richtzenhain, L.J.; Vasconcellos, S.A. Detection of Leptospira spp. in semen and vaginal fluids of goats and sheep by polymerase chain reaction. Theriogenology 2008, 69, 837–842. [Google Scholar] [CrossRef]
- Bolin, C.A.; Cassells, J.A. Isolation of Leptospira interrogans serovars bratislava and hardjo from swine at slaughter. J. Vet. Diagn. Investig. 1992, 4, 87–89. [Google Scholar] [CrossRef]
- Barlozzari, G.; Franco, A.; Macrì, G.; Lorenzetti, S.; Maggiori, F.; Dottarelli, S.; Maurelli, M.; Di Giannatale, E.; Tittarelli, M.; Battisti, A.; et al. First report of Brucella suis biovar 2 in a semi free-range pig farm, Italy. Vet. Ital. 2015, 51, 151–154. [Google Scholar]
- Carnevali, L.; Pedrotti, L.; Riga, F.; Toso, S. Banca Dati Ungulati: Status, distribuzione, consistenza, gestione e prelievo venatorio delle popolazioni di Ungulati in Italia. Rapporto 2001–2005. Biol. e Conserv. della Fauna 2009, 117, 1–168. [Google Scholar]


{kind=link}
| Samples | Province | Age Class | Leptospira Species |
|---|---|---|---|
| Wild Boar 22 | Livorno | Young | Leptospira fainei |
| Wild Boar 70 | Grosseto | Adult | Leptospira fainei |
| Wild Boar 92 | Pisa | Young | Leptospira fainei |
| Wild Boar 133 | Grosseto | Adult | Leptospira fainei |
| Wild Boar 144 | Pisa | Young | Leptospira fainei |
| Wild Boar 163 | Grosseto | Adult | Leptospira fainei |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cilia, G.; Bertelloni, F.; Cerri, D.; Fratini, F. Leptospira fainei Detected in Testicles and Epididymis of Wild Boar (Sus scrofa). Biology 2021, 10, 193. https://doi.org/10.3390/biology10030193
Cilia G, Bertelloni F, Cerri D, Fratini F. Leptospira fainei Detected in Testicles and Epididymis of Wild Boar (Sus scrofa). Biology. 2021; 10(3):193. https://doi.org/10.3390/biology10030193
Chicago/Turabian StyleCilia, Giovanni, Fabrizio Bertelloni, Domenico Cerri, and Filippo Fratini. 2021. "Leptospira fainei Detected in Testicles and Epididymis of Wild Boar (Sus scrofa)" Biology 10, no. 3: 193. https://doi.org/10.3390/biology10030193