
Response of Annual Herbaceous Plant Leaching and Decomposition to Periodic Submergence in Mega-Reservoirs: Changes in Litter Nutrients and Soil Properties for Restoration



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
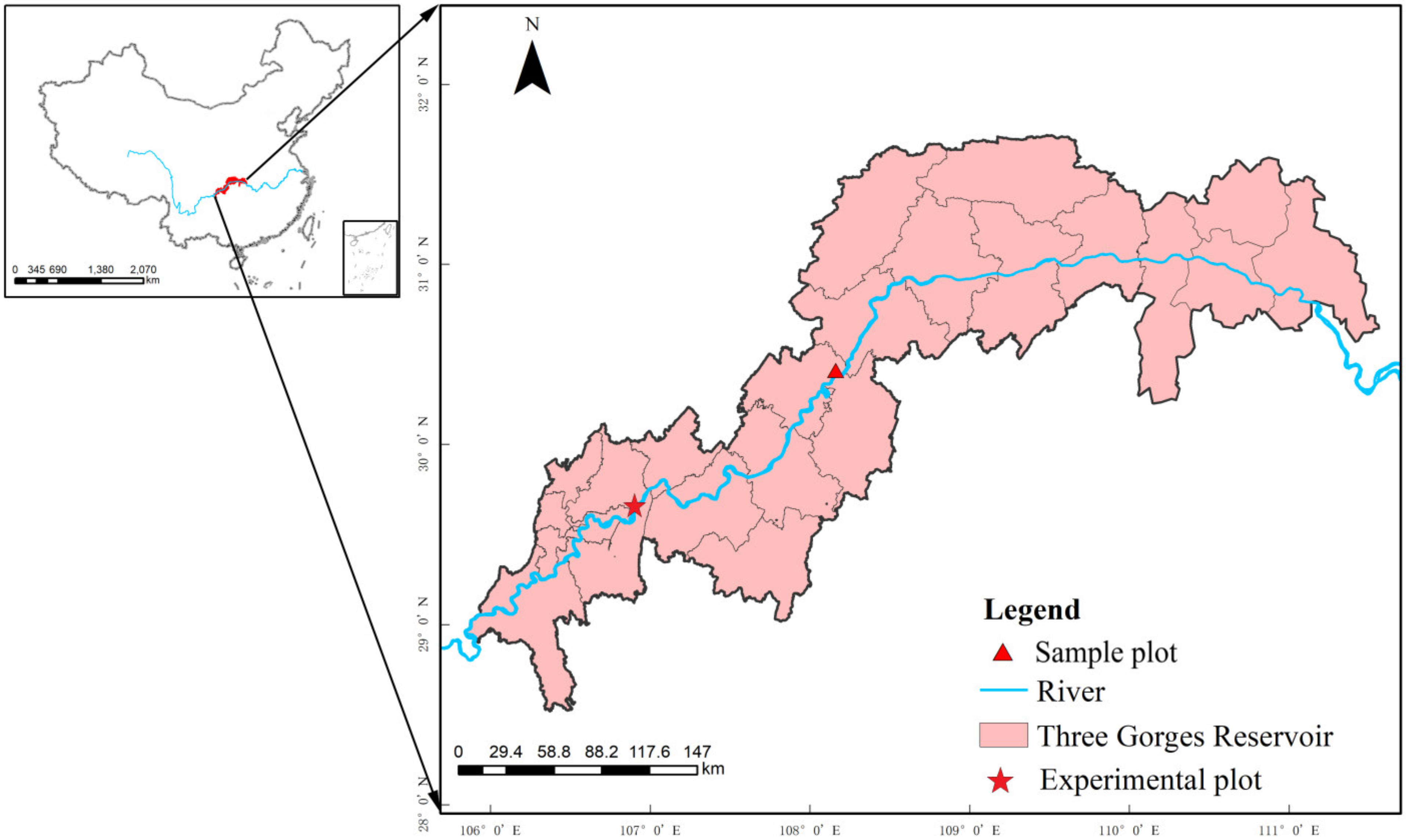
2.1. Study Site and Experimental Materials
2.2. Experimental Design
2.3. Treatments and Measuring Methods
2.4. Statistical Processing and Analysis
3. Results
3.1. Dynamic Characteristics of Mass and Nutrient Elements in the Litter
3.2. Change in Carbon and Nutrient Content in the Soil
3.3. Change in Nutrient Contents in the Overlying Water
3.4. Relationship between Nutrient Change and Soil Properties in Litter Decomposition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, L.; Chen, Y.; Zhou, Y.; Xu, Z.; Tan, B.; You, C.; Zhang, L.; Li, H.; Zheng, H.; Guo, L.; et al. Environmental Conditions and Litter Nutrients are Key Determinants of Soluble C, N, and P Release during Litter Mixture Decomposition. Soil Tillage Res. 2021, 209, 104928. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, S.; Shu, X.; Yang, Y.; Li, Y.; Chen, J.; Pan, Y.; Sun, S. Effects of Norfloxacin on Decomposition and Nutrient Release in Leaves of the Submerged Macrophyte Vallisneria natans (Lour.) Hara. Environ. Pollut. 2021, 274, 116557. [Google Scholar] [CrossRef] [PubMed]
- Simone, S.L.; Scott, D.T.; Mark, O.G.; Klement, T. Leaf-Decomposition Heterogeneity across a Riverine Floodplain Mosaic. Aquat. Sci. 2008, 70, 337–346. [Google Scholar]
- Zhang, P.; Scheu, S.; Li, B.; Lin, G.; Zhao, J.; Wu, J. Litter C Transformations of Invasive Spartina alterniflora Affected by Litter Type and Soil Source. Biol. Fertil. Soils 2020, 56, 369–379. [Google Scholar] [CrossRef]
- Sun, H.; Wang, Q.; Liu, N.; Li, L.; Zhang, C.; Liu, Z.; Zhang, Y. Effects of Different Leaf Litters on the Physicochemical Properties and Soil Microbial Communities in Panax Ginseng-Growing Soil. Acta Ecol. Sin. 2018, 38, 3603–3615. [Google Scholar]
- Qiao, L.; Schaefer, D.; Zou, X. Variations in Net Litter Nutrient Input Associated with Tree Species Influence on Soil Nutrient Contents in a Subtropical Evergreen Broad-Leaved Forest. Chin. Sci. Bull. 2014, 59, 46–53. [Google Scholar] [CrossRef]
- Wu, H.; Lü, X.; Yang, Q. Factors Affecting Litter Decomposition of Wetland Herbaceous Macrophytes. Chin. J. Ecol. 2006, 25, 1405–1411. [Google Scholar]
- Webster, J.R.; Benfield, E.F. Vascular Plant Breakdown in Freshwater Ecosystems. Ann. Rev. Ecol. Syst. 1986, 17, 567–594. [Google Scholar] [CrossRef]
- Xie, Y.; Xie, Y.-H.; Xiao, H. Differential Responses of Litter Decomposition to Climate Between Wetland and Upland Ecosystems in China. Plant Soil 2019, 440, 1–9. [Google Scholar] [CrossRef]
- Enríquez, S.; Duarte, C.M.; Sand-Jensen, K. Patterns in Decomposition Rates among Photosynthetic Organisms: The Importance of Detritus C:N:P Content. Oecologia 1993, 94, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.L.; Martins, P.; Ricardo, F.; Rodrigues, A.M.; Quintino, V. In situ Experimental Decomposition Studies in Estuaries: A Comparison of Phragmites australis and Fucus vesiculosus. Estuar. Coast. Shelf Sci. 2011, 92, 573–580. [Google Scholar]
- Nicolas, B.; Alexandre, C.; Jacinthe, R.; Steven, W.K.; David, R. Microsite Conditions Influence Leaf Litter Decomposition in Sugar Maple Bioclimatic Domain of Quebec. Biogeochemistry 2019, 145, 107–126. [Google Scholar]
- William, F.D.; Ramesh, R.K. Litter Decomposition and Nutrient Dynamics in a Phosphorus Enriched Everglades Marsh. Biogeochemistry 2005, 75, 217–240. [Google Scholar]
- Xu, C.; Zhao, W.; Yu, X. Decomposition of Wetland Plant Residue and its Influencing Factors: A review. Chin. J. Ecol. 2020, 39, 3865–3872. [Google Scholar]
- Chen, Z.; Wang, C.; Chen, X.; Yuan, Z.; Song, H.; Li, C. Heterogeneous Leaves of Predominant Trees Species Enhance Decomposition and Nutrient Release in the Riparian Zone of the Three Gorges Reservoir. Sci. Rep. 2020, 10, 17382. [Google Scholar] [CrossRef] [PubMed]
- Seema, S.; Pirzada, J.S.; Ra, A.; Syed, S.S.; Zeba, F. Decomposition of Avicennia marina (Forsk.) Vierh. Foliage under Field and Laboratory Conditions in the Backwaters of Karachi, Pakistan. Bangladesh J. Bot. 2015, 44, 1–7. [Google Scholar]
- Li, S.; Deng, Y.; Shi, F.; Hu, M.; Pang, B.; Wang, Y.; Li, K.; Chen, M.; Peng, W.; Qu, X.; et al. Research Progress on Water-level-fluctuation Zones of Reservoirs: A review. Wetl. Sci. 2019, 17, 689–696. [Google Scholar]
- Du Laing, G.; Bontinck, A.; Samson, R.; Vandecasteele, B.; Vanthuyne, D.; Meers, E.; Lesage, E.; Tack, F.; Verloo, M. Effect of Decomposing Litter on the Mobility and Availability of Metals in the Soil of a Recently Created Floodplain. Geoderma 2008, 147, 34–46. [Google Scholar] [CrossRef]
- Osim, E.; Natalie, H.; Grace, I.; Chuxia, L. Effects of Heavy Metals on Organic Matter Decomposition in Inundated Soils: Microcosm Experiment and Field Examination. Sci. Total Environ. 2020, 724, 138223. [Google Scholar]
- Niu, A.; Zhou, T.; Yang, X.; Gao, Y.; Xu, S.; Lin, C. Evaluating Litter Yield and Decomposition for Re-Vegetated Mangroves in a Subtropical Mudflat. Appl. Sci. 2019, 9, 3340. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, A.; Zheng, J.; Charles, W.; Zhang, S.; Yuan, Z.; Chen, Z.; Dong, Z.; He, X.; Li, C. Assessing Riparian Zone Changes under the Influence of Stress Factors in Higher-Order Streams and Tributaries: Implications for the Management of Massive Dams and Reservoirs. Sci. Total Environ. 2021, 776, 146011. [Google Scholar]
- Zheng, J.; Arif, M.; Zhang, S.; Yuan, Z.; Zhang, L.; Dong, Z.; Tan, X.; Charles, W.; Li, C. The Convergence of Species Composition along the Drawdown Zone of the Three Gorges Dam Reservoir, China: Implications for restoration. Environ. Sci. Pollut. Res. 2021, 31, 42609–42621. [Google Scholar] [CrossRef] [PubMed]
- Arif, M.; Zhang, S.; Jie, Z.; Charles, W.; Sanelisiwe Mzondi, P.; Changxiao, L. Evaluating the Effects of Pressure Indicators on Riparian Zone Health Conditions in the Three Gorges Dam Reservoir, China. Forests 2020, 11, 214–234. [Google Scholar] [CrossRef] [Green Version]
- Arif, M.; Tahir, M.; Jie, Z.; Changxiao, L. Impacts of Riparian Width and Stream Channel Width on Ecological Networks in Main Waterways and Tributaries. Sci. Total Environ. 2021, 792, 148457. [Google Scholar] [CrossRef]
- Chen, Z.; Arif, M.; Wang, C.; Chen, X.; Li, C. Effects of Hydrological Regime on Foliar Decomposition and Nutrient Release in the Riparian Zone of the Three Gorges Reservoir, China. Front. Plant Sci. 2021, 12, 14. [Google Scholar]
- Wang, B.; Yan, D.; Wen, A.; Chen, J. Influencing Factors of Sediment Deposition and their Spatial Variability in riparian zone of the Three Gorges Reservoir, China. J. Mt. Sci. 2016, 13, 1387–1396. [Google Scholar] [CrossRef]
- Zhou, Z.; Lu, Y.; Zhong, R.; Bi, X.; Yuan, X.; Guo, Y. Research Progress on Soil Environment of Water-level Fluctuation Zone Affected by Dam Operation. J. China Hydrol. 2019, 39, 15–19. [Google Scholar]
- Jing, L.; Stephen, J.F.; Stuart, E.B.; Michele, A.B. Macrophyte Beds in a Subtropical Reservoir Shifted from a Nutrient Sink to a Source after Drying then Rewetting. Freshw. Biol. 2017, 62, 854–867. [Google Scholar]
- Chen, G.P.; Cheng, S.S.; Cong, M.Y.; Liu, J.; Gao, X.; Wang, H.; Shi, F. Effects of Litter Decomposition on Soil Nutrients in Three Broad-Leaved Forests. Chin. J. Ecol. 2014, 33, 874–879. [Google Scholar]
- Tang, X.; Wu, M.; Li, R. Phosphorus Distribution and Bioavailability Dynamics in the Mainstream Water and Surface Sediment of the Three Gorges Reservoir between 2003 and 2010. Water Res. 2018, 145, 321–331. [Google Scholar] [CrossRef]
- Wei, Y.; Zhang, M.; Cui, L.; Pan, X.; Liu, W.; Li, W.; Lei, Y. Winter Decomposition of Emergent Macrophytes Affects Water Quality under Ice in a Temperate Shallow Lake. Water 2020, 12, 2640. [Google Scholar] [CrossRef]
- Qin, Q.; Wang, H.; Li, X.; Xie, Y.; Lei, X.; Zheng, Y.; Yang, D. Ecological Function of Semi-Decomposition Litter in Natural Spruce-Fir Mixed Forest of Changbai Mountains. For. Res. 2019, 32, 147–152. [Google Scholar]
- Wu, E.H.; Liu, Q.; Wang, X.D.; Huang, Y.C. Litter Decomposition and Dynamics of Soil Nutrients in Casuarina equisetifolia Plantation on the Coast of Hainan Island. Hubei Agric. Sci. 2013, 52, 60–64. [Google Scholar]
- Robert, L.E.; Kerri, M.T. Red Alder Leaf Decomposition and Nutrient Release in Alder and Conifer Riparian Patches in Western Washington, USA. For. Ecol. Manag. 2010, 259, 2375–2381. [Google Scholar]
- Cao, C.; Liu, S.-Q.; Ma, Z.-B.; Lin, Y.; Su, Q.; Chen, H.; Wang, J.-J.W. Dynamics of Multiple Elements in Fast Decomposing Vegetable Residues. Sci. Total Environ. 2018, 616–617, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Mark, O.G.; Eric, C.; Mike, D. A Perspective on Leaf Litter Breakdown in Streams. Oikos 1999, 85, 377–384. [Google Scholar]
- Zawislanski, P.T.; Chau, S.; Mountford, H.; Wong, H.C.; Sears, T.C. Accumulation of Selenium and Trace Metals on Plant Litter in a Tidal Marsh. Estuar. Coast. Shelf Sci. 2001, 52, 589–603. [Google Scholar] [CrossRef]
- Cory, C.C.; Jason, C.N.; Alan, R.T.; Eran, H. Composition, Dynamics, and Fate of Leached Dissolved Organic Matter in Terrestrial Ecosystems: Results from a Decomposition Experiment. Ecosystems 2004, 7, 275–285. [Google Scholar]
- Li, C.; Wang, X.; Sun, B. Characteristics of Nutrient Release and its Affecting Factors during Plant Residue Decomposition under Different Climate and Soil Conditions. Acta Ecol. Sin. 2017, 5, 1206–1217. [Google Scholar]
- Lutz, H.J.; Chandler, R.F. Forest Soils. Q. Rev. Biol. 1946, 3, 249. [Google Scholar] [CrossRef]
- Yong, X. The Decomposition of Triarrhena lutarioriparia and Carex cineraseens Litter and Dynamic Change of Microbial Community in the Wetland of Nanjishan. Master’s Thesis, Nanchang University, Nanchang, China, 2012. [Google Scholar]
- Chao, L.; Liu, Y.; Freschet, G.T.; Zhang, W.; Yu, X.; Zheng, W.; Guan, X.; Yang, Q.; Chen, L.; Dijkstra, F.A.; et al. Litter Carbon and Nutrient Chemistry Control the Magnitude of Soil Priming Effect. Funct. Ecol. 2019, 33, 876–888. [Google Scholar] [CrossRef]
- Ryosuke, N.; Hirofumi, K.; Kiyosada, K.; Wataru, N.; Masataka, O.; Naoki, O. Diverse Recalcitrant Substrates Slow Down Decomposition of Leaf Litter from Trees in a Serpentine Ecosystem. Plant Soil 2019, 442, 247–255. [Google Scholar]
- John, M.B.; Robert, W.P.; Michael, H.B. Decay Rates, Nitrogen Fluxes, and Decomposer Communities of Single- and Mixed-Species Foliar Litter. Ecology 1990, 71, 1976–1985. [Google Scholar]
- Wang, C.; Dong, X.T.; Du, R.-P.; Zhang, Z.D.; Huang, X.R. Changes of Nutrient Release and Enzyme Activity during the Decomposition of Mixed Leaf Litter of Larix principis-rupprechtii and Broadleaved Tree Species. Chin. J. Appl. Ecol. 2021, 32, 1709–1716. [Google Scholar]
- Wang, J.; Huang, J.-H. Comparison of Major Nutrient Release Patterns in Leaf Litter Decomposition in Warm Temperate Zone of China. Acta Phytoecol. Sin. 2001, 25, 375–380. [Google Scholar]
- Ondřej, K.; František, N.; Richard, H.; Miroslav, V. Saprotrophic Fungi Transform Organic Phosphorus from Spruce Needle Litter. Soil Biol. Biochem. 2006, 38, 3372–3379. [Google Scholar]
- Yan, H.; Gu, X.; Shen, H. Microbial Decomposition of Forest Litter: A Review. Chin. J. Ecol. 2010, 29, 1827–1835. [Google Scholar]
- Ge, X.G.; Xiao, W.F.; Zeng, L.X.; Huang, Z.L.; Fu, T.; Feng, X.H. Relationships between Litter Substrate Quality and Soil Nutrients in Different-Aged Pinus massoniana stands. Acta Ecol. Sin. 2012, 32, 852–862. [Google Scholar]
- Liang, J.; Jiang, T.; Lu, S.; Wei, S.Q.; Wang, D.Y.; Chen, X.S.; Wang, Q.L. Spectral Characteristics of Dissolved Organic Matter (Dom) Releases from Soils of Typical Water-Level Fluctuation Zones of Three Gorges Reservoir Areas: UV-vis Spectrum. Environ. Sci. 2016, 37, 2496–2505. [Google Scholar]
- Yan, X. Accumulation and Transportation of Heavy Metals During the Growth and Decomposition of Spartina alterniflora and Phragmites australis. Master’s Thesis, East China Normal University, Shanghai, China, 2018. [Google Scholar]
- Sharon, S.V.; Robert, K.N.; Stephen, M.K. Emergent Plant Decomposition and Sedimentation: Response to Sediments Varying in Texture, Phosphorus Content and Frequency of Deposition. Environ. Exp. Bot. 1998, 40, 43–58. [Google Scholar]
- Cornut, J.; Elger, A.; Lambrigot, D.; Marmonier, P.; Chauvet, E. Early Stages of Leaf Decomposition are Mediated by Aquatic Fungi in the Hyporheic Zone of Woodland Streams. Freshw. Biol. 2010, 55, 2541–2556. [Google Scholar] [CrossRef]
- Zheng, W.; Lei, H.; Bo, Z. Characteristics of Greenhouse Gases Emission under Inundation from Dominant Herbaceous Plants in the Water-Level Fluctuation Zone of the Three Gorges Reservoir. Resour. Environ. Yangtze Basin 2020, 29, 1965–1974. [Google Scholar]
- Yavitt, J.B.; Kryczka, A.K.; Huber, M.E.; Pipes, G.T.; Rodriguez, A.M. Inferring Methane Production by Decomposing Tree, Shrub, and Grass Leaf Litter in Bog and Rich Fen Peatlands. Front. Environ. Sci. 2019, 7, 182. [Google Scholar] [CrossRef]
- William, K.C.; Johannes, H.C.C.; Kathryn, A.; Ellen, D.; Valerie, T.E.; Oscar, G.; Sarah, E.H.; Bart, H.; Hiroko, K.; Natalia, P.; et al. Plant Species Traits are the Predominant Control on Litter Decomposition Rates Within Biomes Worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar]
- Nuket, A.; Beth, M. Leaf Litter Decomposition along the Porsuk River, Eskisehir, Turkey. Can. J. Bot. 1997, 75, 1394–1397. [Google Scholar]
- Sang, C.; Zheng, Y.; Zhou, Q.; Li, D.; Liang, G.; Gao, Y. Effects of Water Impoundment and water-Level Manipulation on the Bioaccumulation Pattern, Trophic Transfer and Health Risk of Heavy Metals in the Food Web of Three Gorges Reservoir (China). Chemosphere 2019, 232, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.L.; Zhou, R.Y.; Xia, C.; Xu, J.L. Effects of Oxidation-Reduction Potential and Microorganism on the Release of Phosphorus from Sediments. Environ. Chem. 2014, 33, 930–936. [Google Scholar]
- Li, Q.; Liu, Y.; Du, Y.; Cui, Z.; Shi, L.; Wang, L.; Li, H. The Behavior of Heavy Metals in Tidal Flat Sediments during Fresh Water Leaching. Chemosphere 2011, 82, 834–838. [Google Scholar] [CrossRef]
- Cao, X.; Han, R.; Zhang, T.; Wang, G.; Wei, H.; Ma, Y.; Ji, F.; Ma, J. Decomposition of Aquatic Plants during Winter and its Influence on Water Quality. J. Agro-Environ. Sci. 2015, 34, 361–369. [Google Scholar]
- McLatchey, G.P.; Reddy, K.R. Regulation of Organic Matter Decomposition and Nutrient Release in a Wetland Soil. J. Environ. Qual. 1998, 27, 1268–1274. [Google Scholar] [CrossRef]
- Mäggi, H.; Mark, O.G. Contribution of Stream detrivores, Fungi, and Bacteria to Leaf Breakdown Based on Biomass Estimates. Ecology 2002, 83, 1026–1038. [Google Scholar]
- Zheng, J.; Arif, M.; Zhang, S.; Yuan, Z.; Zhang, L.; Li, J.; Li, C. Dam Inundation Simplifies the Plant Community Composition. Sci. Total Environ. 2021, 801, 149827. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | g C kg−1 | g N kg−1 | g P kg−1 | g K kg−1 | C/N | C/P |
|---|---|---|---|---|---|---|
| Echinochloa crusgali | 384.17 ± 2.16 | 12.20 ± 0.53 | 3.56 ± 0.21 | 16.73 ± 1.05 | 31.53 ± 1.51 | 108.24 ± 6.77 |
| Bidens tripartita | 428.70 ± 0.87 | 12.07 ± 0.35 | 3.87 ± 0.14 | 14.5 ± 1.06 | 35.55 ± 0.97 | 110.91 ± 3.97 |
| Species | Time | Buried Depth | ORP/mv | T/°C | pH |
|---|---|---|---|---|---|
| E. crusgali | 30 d | 0 cm | 416.33 ± 1.15 a | 13.23 ± 0.12 b | 8.37 ± 0.06 a |
| 5 cm | −34.67 ± 11.59 b | 14.20 ± 0.36 a | 7.79 ± 0.18 b | ||
| 10 cm | −52.33 ± 23.46 b | 14.17 ± 0.12 a | 7.73 ± 0.03 b | ||
| 90 d | 0 cm | 468.67 ± 0.58 a | 7.03 ± 0.06 a | 8.97 ± 0.05 a | |
| 5 cm | −48.33 ± 24.01 b | 7.03 ± 0.12 a | 7.5 ± 0.06 b | ||
| 10 cm | −71.67 ± 39.88 b | 7.23 ± 0.42 a | 7.53 ± 0.05 b | ||
| 180 d | 0 cm | 413.33 ± 2.52 a | 20.03 ± 0.12 b | 8.08 ± 0.07 a | |
| 5 cm | 141.33 ± 48.95 b | 20.60 ± 0.30 a | 6.79 ± 0.15 b | ||
| 10 cm | 124.33 ± 12.58 b | 20.33 ± 0.12 ab | 6.47 ± 0.39 b | ||
| B. tripartite | 30 d | 0 cm | 407.67 ± 2.31 a | 13.30 ± 0.10 b | 8.36 ± 0.21 a |
| 5 cm | −78.67 ± 30.89 b | 14.13 ± 0.06 a | 7.75 ± 0.10 b | ||
| 10 cm | −85.33 ± 45.39 b | 14.30 ± 0.10 a | 7.87 ± 0.07 b | ||
| 90 d | 0 cm | 465.00 ± 0.00 a | 7.03 ± 0.06 a | 8.94 ± 0.12 a | |
| 5 cm | −32.00 ± 10.58 b | 7.00 ± 0.10 a | 7.42 ± 0.06 b | ||
| 10 cm | −52.67 ± 22.28 b | 7.03 ± 0.15 a | 7.44 ± 0.10 b | ||
| 180 d | 0 cm | 408.00 ± 2.00 a | 20.17 ± 0.15 b | 8.15 ± 0.06 a | |
| 5 cm | 113.00 ± 13.08 b | 21.47 ± 0.29 a | 6.06 ± 0.39 b | ||
| 10 cm | 164.33 ± 35.85 b | 20.97 ± 0.31 a | 6.41 ± 0.47 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, X.; Xie, T.; Arif, M.; Ding, D.; Li, J.; Yuan, Z.; Li, C. Response of Annual Herbaceous Plant Leaching and Decomposition to Periodic Submergence in Mega-Reservoirs: Changes in Litter Nutrients and Soil Properties for Restoration. Biology 2021, 10, 1141. https://doi.org/10.3390/biology10111141
Hu X, Xie T, Arif M, Ding D, Li J, Yuan Z, Li C. Response of Annual Herbaceous Plant Leaching and Decomposition to Periodic Submergence in Mega-Reservoirs: Changes in Litter Nutrients and Soil Properties for Restoration. Biology. 2021; 10(11):1141. https://doi.org/10.3390/biology10111141
Chicago/Turabian StyleHu, Xin, Tingting Xie, Muhammad Arif, Dongdong Ding, Jiajia Li, Zhongxun Yuan, and Changxiao Li. 2021. "Response of Annual Herbaceous Plant Leaching and Decomposition to Periodic Submergence in Mega-Reservoirs: Changes in Litter Nutrients and Soil Properties for Restoration" Biology 10, no. 11: 1141. https://doi.org/10.3390/biology10111141