
Genetic Diversity in Invasive Populations of Lupinus polyphyllus Lindl. and Heracleum sosnowskyi Manden.

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Isolation
2.3. DNA Amplification
2.4. Gel Electrophoresis of the Amplification Products
2.5. Quantitative Estimates of Genetic Diversity
3. Results
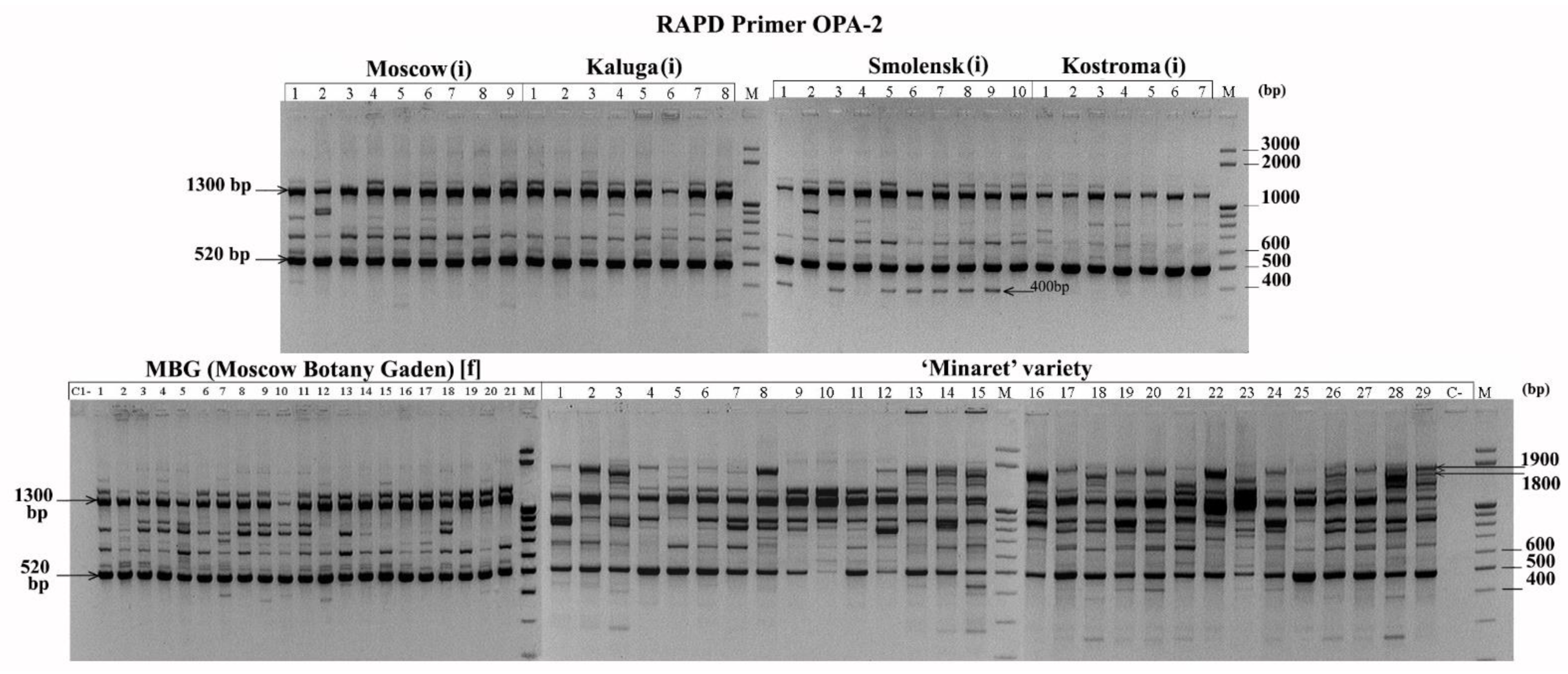
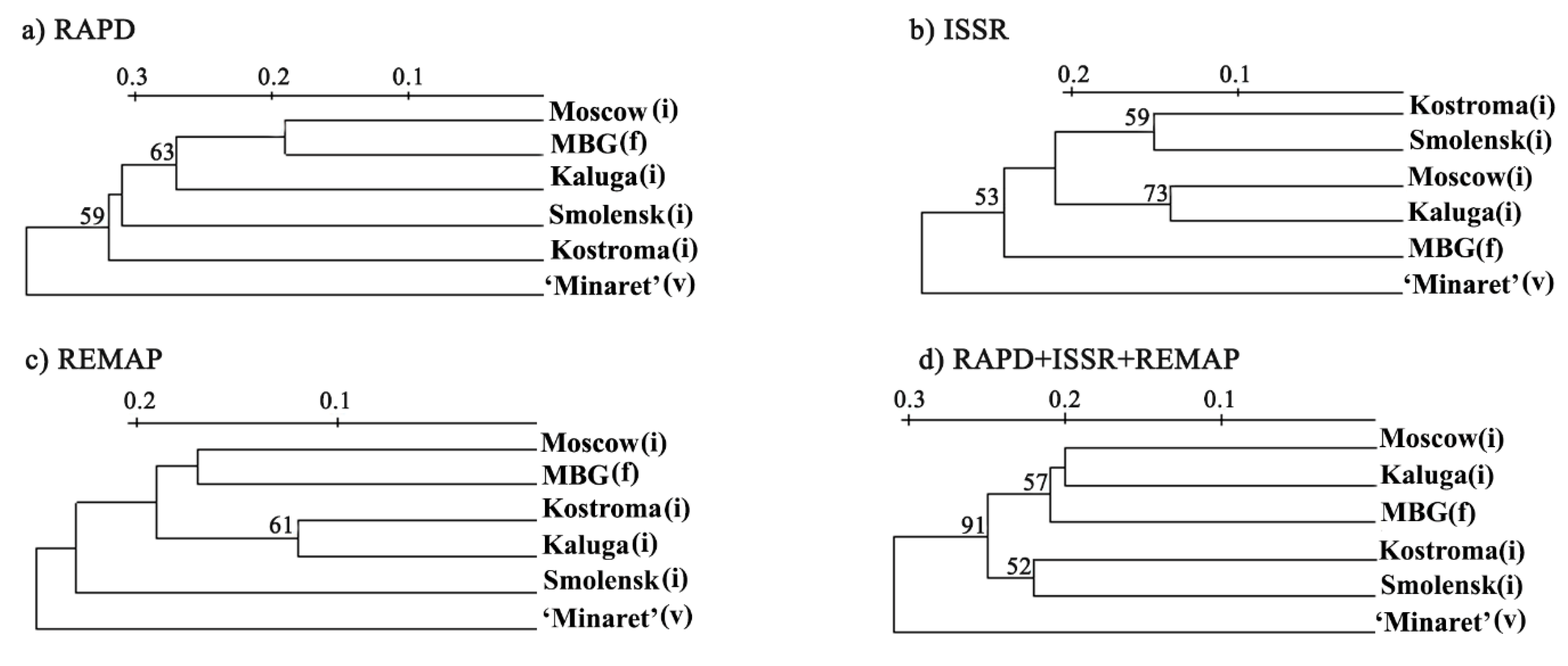
3.1. Molecular Analyses of L. polyphyllus
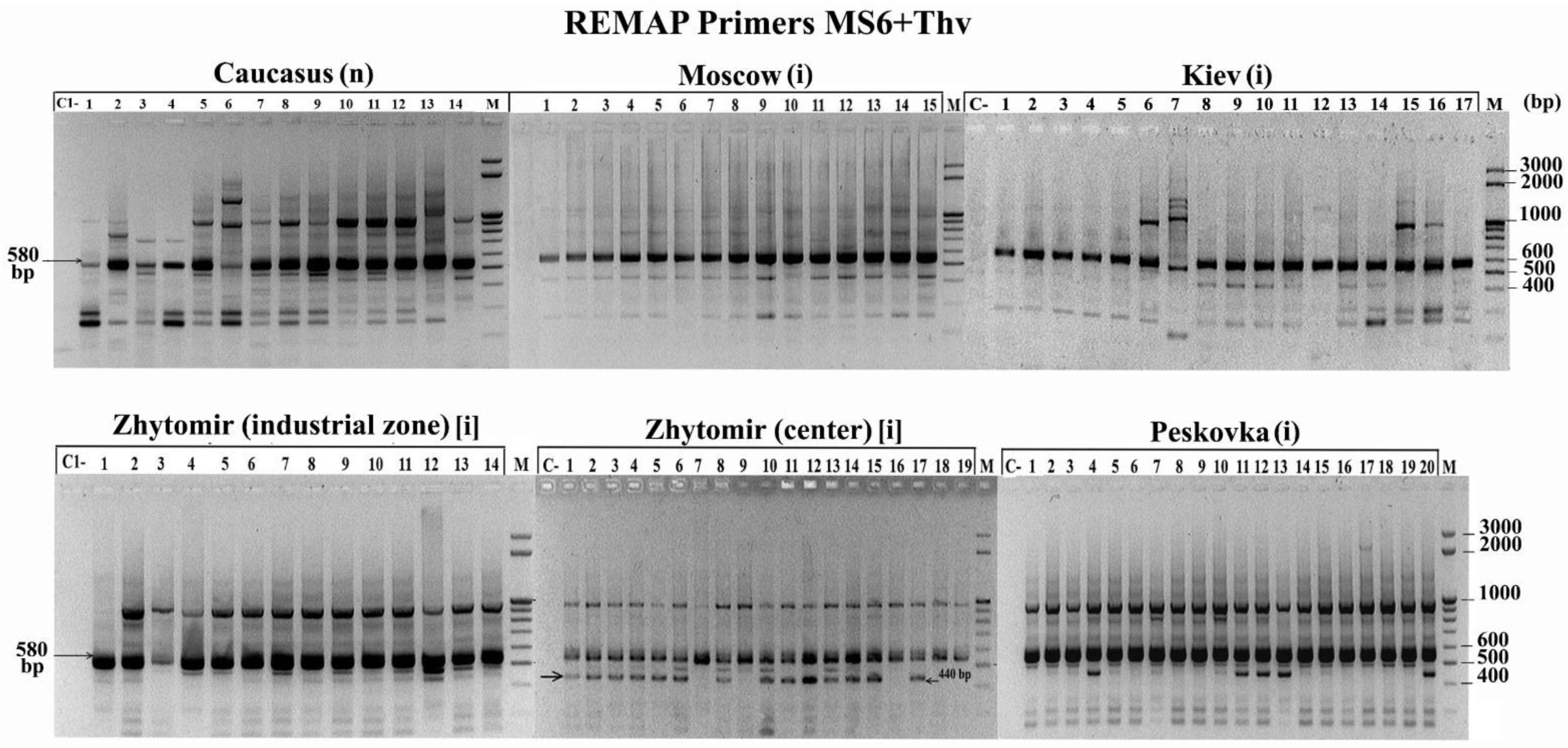
3.2. Molecular Analyses of H. sosnowskyi
4. Discussion
4.1. Discussion of the Genetic Diversity within Populations of Lupin
4.2. Discussion of the Genetic Diversity within Populations of Hogweed
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olmstead, R.G. Review: Are invasive plants an inevitable consequence of evolution? Am. J. Bot. 2006, 93, 1236–1239. [Google Scholar] [CrossRef]
- Capizzi, D.; Genovesi, P.; Monaco, A.; Scalera, R.; Carnevali, L. Impact of alien mammals on human health. In Invasive Species and Human Health; Mazza, G., Tricarico, E., Eds.; CABI: New York, NY, USA, 2018. [Google Scholar] [CrossRef]
- Adriaens, T.; Branquart, E.; Gosse, D.; Reniers, J.; Vanderhoeven, S. Feasibility of Eradication and Spread Limitation for Species of Union Concern Sensu the EU IAS Regulation (EU 1143/2014) in Belgium; Report Prepared in Support of Implementing the IAS Regulation in Belgium; INBO: Brussel, Belgium, 2019. [Google Scholar] [CrossRef]
- Bertolino, S.; Vimercati, G.; Paoloni, D.; Martinoli, A.; Wauters, L.A.; Genovesi, P.; La Morgia, V. Restricted access to private properties limits management of invasive alien species: A literature review and case studies. J Environ. Manag. 2021, 297, 113318. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://ec.europa.eu/environment/nature/invasivealien/index_en.htm (accessed on 12 August 2021).
- Vinogradova, Y.K.; Aistova, E.V.; Antonova, L.A.; Chernyagina, O.A.; Chubar, E.A.; Darman, G.F.; Devyatova, E.A.; Khoreva, M.G.; Kotenko, O.V.; Marchuk, E.A.; et al. Invasive plants in flora of the Russian far east: The checklist and comments. Bot. Pacifica 2020, 9, 103–129. [Google Scholar] [CrossRef]
- Scoggan, H.J. The Flora of Canada. Pt 3; National Museums of Canada: Ottawa, ON, Canada, 1978; p. 1115. [Google Scholar] [CrossRef]
- Schmid, R.; Preston, C.D.; Pearman, D.A.; Dines, T.D.; Arnold, H.R.; Croft, J.M. New Atlas of the British and Irish Flora: An Atlas of the Vascular Plants of Britain, Ireland, the Isle of Man and the Channel Islands; Taxon: Oxford, UK, 2003; Volume 52, p. 884. [Google Scholar] [CrossRef]
- Andrews, M.; James, E.K.; Sprent, J.I.; Boddey, R.M.; Fábio, E.; Bueno dos Reis, F., Jr. Nitrogen fixation in legumes and actinorhizal plants in natural ecosystems: Values obtained using 15N natural abundance. Plant Ecol. Divers. 2011, 4, 131–140. [Google Scholar] [CrossRef]
- Fremstad, E. NOBANIS—Invasive Alien Species Fact Sheet—Lupinus polyphyllus. In Online Database of the European Network on Invasive Alien Species—NOBANIS; 2010; Available online: https://www.nobanis.org/globalassets/speciesinfo/l/lupinus-polyphyllus/lupinus-polyphyllus.pdf (accessed on 12 August 2021).
- Prjanishnikov, D.N. Ob Udobrenii Polej i Sevooborotah; Moscow Publisher of Agriculture economy RSFSR: Moscow, Russia, 1962; p. 422. [Google Scholar]
- Kurlovich, B.S.; Repyev, S.I. The gene bank and breeding of grain legumes; Institute of Plant Industry: Moscow, Russia, 1995; 438p. [Google Scholar]
- Majsurjan, N.A.; Atabekova, A.I. Ljupin; Moskva Kolos: Moscow, Russia, 1974; p. 463. [Google Scholar]
- Vinogradova, J.K.; Majorov, S.R.; Horun, L.V. Chernaja kniga flory Srednej Rossii; Moskva GEOS: Moscow, Russia, 2010; p. 512. [Google Scholar]
- Pimenov, M.G.; Ostroumova, T.A. Zontichnye (Umbelliferae) Rossii; Tovarishhestvo nauchnyh izdanij KMK: Moskva, Russia, 2012; p. 477. [Google Scholar]
- Mandenova, I.P. Kavkazskie vidy roda Heracleum; Akademia Nauk Gruzinskoy SSR: Tbilisi, Georgia, 1950; p. 104. [Google Scholar]
- Satsyperova, I.F. Borshheviki Flory SSSR—Novye Kormovye Rastenija: Perspektivy Ispol’zovanija v Narodnom Hozjajstve; Nauka: Leningrad, Russia, 1984; p. 223. [Google Scholar]
- Pesnya, D.S.; Serov, D.A.; Vakorin, S.A.; Prokhorova, I.M. Research of the Toxic, Mitosis Modifying and Mutagen Effect of Heracleum Sosnowskyi. Yarosl. Pedagog. Bull. Sci. Mag. 2011, 3, 93–98. [Google Scholar]
- Ivanov, M.F.; Kolosov, A.A. Bor’ba s borshhevikom Sosnovskogo v Novgorodskoj oblasti. Zashhita Karantin Rastenij 2012, 9, 26–27. [Google Scholar]
- Nielsen, C.; Ravn, H.P.; Nentwig, W.; Wade, M. The Giant Hogweed Best Practice Manual. Guidelines for the Management and Control of an Invasive Weed in Europe; Forest & Landscape Denmark: Hoersholm, Denmark, 2005; p. 44. [Google Scholar]
- Pyšek, P.; Cock, M.J.W.; Nentwig, W.; Ravn, H.P. Ecology and Management of Giant Hogweed: (Heracleum mantegazzianum); CABI: Cambridge, MA, USA, 2007; pp. 1–324. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The Population Biology of Invasive Species. Annu. Rev. Ecol. Syst. 2003, 32, 305–332. [Google Scholar] [CrossRef] [Green Version]
- Ellstrand, N.C.; Schierenbeck, K.A. Hybridization as a stimulus for the evolution of invasiveness in plants? Proc. Natl. Acad. Sci. USA 2000, 97, 7043–7050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossdorf, O.; Auge, H.; Lafuma, L.; Rogers, W.E.; Siemann, E.; Prati, D. Phenotypic and genetic differentiation between native and introduced plant populations. Oecologia 2005, 144, 1–11. [Google Scholar] [CrossRef]
- Novak, S.; Mack, R.N. Genetic bottlenecks in alien plant species: Influence of mating systems and introduction dynamics. In Species Invasions: Insights into Ecology, Evolution, and Biogeography; Sax, D.F., Stachowicz, J.J., Gaines, S.D., Eds.; Sinauer Associates: Sunderland, MA, USA, 2005; pp. 201–228. [Google Scholar]
- Williams, J.G.K.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef] [Green Version]
- Kantety, R.V.; Zeng, X.; Bennetzen, J.L.; Zehr, B.E. Assessment of genetic diversity in dent and popcorn (Zea mays L.) inbred lines using inter-simple sequence repeat (ISSR) amplification. Mol. Breed. 1995, 1, 365–373. [Google Scholar] [CrossRef]
- Kalendar, R.; Grob, T.; Regina, M.; Suoniemi, A.; Schulman, A.H. IRAP and REMAP: Two new retrotransposon-based DNA fingerprinting techniques. Theor. Appl. Genet. 1999, 98, 704–711. [Google Scholar] [CrossRef]
- Pyshek, P.; Richardson, D.M.; Rejmanek, M.; Webster, G.L.; Williamson, M.; Kirschuer, J. Alien plants in checklists and floras: Towards better communication between taxonomists and ecologists. Taxon 2004, 53, 131–143. [Google Scholar] [CrossRef]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version II. Plant Mol. Biol. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokal, R.; Michener, C. A statistical method for evaluating systematic relationships. Univ. Kansas Bull. 1958, 38, 1409–1438. [Google Scholar]
- Van De Peer, Y.; De Wachter, R. Treecon for windows: A software package for the construction and drawing of evolutionary trees for the microsoft windows environment. Bioinformatics 1994, 10, 569–570. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783. [Google Scholar] [CrossRef] [PubMed]
- Vysniauskiene, R.; Ranftlientl, V.; Zvingila, D. Genetic diversity of invasive alien species Lupinus polyphyllus populations in Lithuania. Zemdirbyste 2011, 98, 383–390. [Google Scholar]
- Li, S.-L.; Vasemägi, A.; Ramula, S. Genetic variation and population structure of the garden escaper Lupinus polyphyllus in Finland. Plant Syst. Evol. 2016, 302, 399–407. [Google Scholar] [CrossRef]
- Qiu, J.; van Santen, E.; Tuzun, S. Optimization of DNA amplification fingerprinting techniques to study genetic relationships of white lupin germplasm. Plant Breed. 1995, 114, 525–529. [Google Scholar] [CrossRef]
- Wolko, B.; Świȩcicki, W.K.; Kruszka, K.; Irzykowska, L. Isozyme and RAPD markers for the identification of pea, field bean and lupin cultivars. J. Appl. Genet. 2000, 41, 151–165. [Google Scholar]
- Gilbert, J.E.; Lewis, R.V.; Wilkinson, M.J.; Caligari, P.D.S. Developing an appropriate strategy to assess genetic variability in plant germplasm collections. Theor. Appl. Genet. 1999, 98, 1125–1131. [Google Scholar] [CrossRef]
- Yorgancilar, M.; Babaoglu, M.; Hakki, E.E.; Atalay, E. Determination of the relationship among Old World Lupin (Lupinus sp.) species using RAPD and ISSR markers. Afr. J. Biotechnol. 2009, 8, 3524–3530. [Google Scholar]
- Artyukhova, A.V. Passportization of Lupin varieties by ISSR-PCR and RAPD-PCR methods for biotechnological research. Ph.D. Thesis, Bryansk State Acad. I.G. Petrovsky University, Ufa, Russia, 2011. [Google Scholar]
- Clements, J.; Galek, R.; Kozak, B.; Michalczyk, D.J.; Piotrowicz-Cieślak, A.I.; Sawicka-Sienkiewicz, E.; Stawiński, S.; Zalewski, D. Diversity of selected Lupinus angustifolius L. genotypes at the phenotypic and DNA level with respect to microscopic seed coat structure and thickness. PLoS One 2014, 9, e102874. [Google Scholar] [CrossRef] [PubMed]
- Guilengue, N.; Alves, S.; Talhinhas, P.; Neves–Martins, J. Genetic and genomic diversity in a Tarwi (Lupinus mutabilis Sweet) germplasm collection and adaptability to mediterranean climate conditions. Agronomy 2020, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Khapilina, O.N.; Daniyarov, A.Z.; Amenov, A.A.; Novakovskaya, A.P.; Turzhanova, A.S.; Tagimanova, D.S.; Filippova, N.I.; Kalendar, R.N. Analysis of genetic diversity in legumes germplasm using retrotransposon based molecular markers. Biotechnol. Theory Pract. 2017. [Google Scholar] [CrossRef]
- Oumer, A.; Petros, Y.; Tesfaye, K.; Abel, T.; Endashaw, B. Inter simple sequence repeat (ISSR) analysis of Ethiopian white lupine (Lupinus albus L.). Afr. J. Biotechnol. 2015, 14, 1552–1563. [Google Scholar] [CrossRef] [Green Version]
- Mahfouze, S.A.; Mahfouze, H.A.; Mubarak, D.M.F.; Esmail, R.M. Evaluation of six imported accessions of Lupinus albus for nutritional and molecular characterizations under Egyptian conditions. Jordan J. Biol. Sci. 2018, 11, 47–56. [Google Scholar]
- AlRawashdeh, I.M.; AlRawashdeh, N.Q. Evaluating the genetic relatedness within Lupinus pilosus L. species based on RAPD analysis. Jordan J. Biol. Sci. 2015, 8, 61–64. [Google Scholar] [CrossRef]
- Sbabou, L.; Brhada, F.; Alami, I.T.; Maltouf, A.F. Genetic diversity of moroccan Lupinus germplasm investigated using ISSR and AFLP Markers. Int. J. Agric. Biol. 2010, 12, 26–32. [Google Scholar]
- Talhinhas, P.; Neves-Martins, J.; Leitão, J. AFLP, ISSR and RAPD markers reveal high levels of genetic diversity among Lupinus spp. Plant Breed. 2003, 122, 507–510. [Google Scholar] [CrossRef]
- Tkacheva, E.V. Biological features of species Leguminosae Juss. with different invasive levels. Ph.D. Thesis, Main Botanical Garden of the Russian Academy of Sciences, Moscow, Russia, 2011. [Google Scholar]
- Ramula, S.; Kalske, A. Introduced plants of Lupinus polyphyllus are larger but flower less frequently than conspecifics from the native range: Results of the first year. Ecol. Evol. 2020, 10, 13742–13751. [Google Scholar] [CrossRef]
- Blossey, B.; Notzold, R. Evolution of increased competitive ability in invasive nonindigenous plants: A Hypothesis. J. Ecol. 1995, 83, 887–889. [Google Scholar] [CrossRef]
- Jahodova, S.; Trybush, S.; Pyshek, P.; Wade, M.; Karp, A. Invasive species of Heracleum in Europe: An insight into genetic relationships and invasion history. Divers. Distrib. 2007, 13, 99–114. [Google Scholar] [CrossRef]
- Solov’ova, A.I.; Dolgikh, Y.I.; Osipova, E.S.; Stepanova, A.Y.; Yavorskaya, O.G. Reveling of HERACLEUM SOSNOWSKYI polymorphism by means RAPD, ISSR, REMAP. In Proceedings of the Biologija Rastenij i Biotehnologija, Ukraine, Belaja cerkov’, October 2011; Institut Pishhevoj Biotehnologii i Genomiki, Nacional’noj Akademii Nauk Ukrainy: Belaja cerkov’, Ukraine, 2011. [Google Scholar]
- Kolmakov, P.Y.; Vysotsky, Y.I.; Bavtuno, A.V.; Kisova, A.S. Jekstrakcija DNK i vyjavlenie polimorfizma Heracleum sp. s pomoshh’ju RAPD-diagnostiki. In Proceedings of the Science—Education, Production, Economy; VGU imeni P.M.Masherova: Vitebsk, Belarus, 2017; pp. 72–73. Available online: https://rep.vsu.by/handle/123456789/10351 (accessed on 24 August 2021).
- Stry`gina, K.V.; Shipilina, L.Y.; Chuxina, I. Pervy`e Shagi v Izuchenii Genoma Borshhevika. 2020. Available online: https://sochisirius.ru/uploads/files2/4-abt.pdf (accessed on 12 August 2021).
- Niinikoski, P.; Korpelainen, H. Population genetics of the invasive giant hogweed (Heracleum sp.) in a northern European region. Plant Ecol. 2015, 216, 1155–1162. [Google Scholar] [CrossRef]
- Gacek, P.; Góralski, G.; Joachimiak, A.J. Chromosome numbers and polyploidy in Polish angiosperms. Acta Biol. Crac. Ser. Bot. 2011, 53, 37–49. [Google Scholar] [CrossRef]
- Weimarck, G.; Stewart, F.; Grace, J. Morphometric and chromatographic variation and male meiosis in the hybrid Heracleum mantegazzianum X H. sphondylium (Apiaceae) and its parents. Hereditas 1979, 91, 117–127. [Google Scholar] [CrossRef]
- Walker, N.F.; Hulme, P.E.; Hoelzel, A.R. Population genetics of an invasive species, Heracleum mantegazzianum: Implications for the role of life history, demographics and independent introductions. Mol. Ecol. 2003, 12, 1747–1756. [Google Scholar] [CrossRef]
- Henry, P.; Le Lay, G.; Goudet, J.; Guisan, A.; Jahodova, Š.; Besnard, G. Reduced genetic diversity, increased isolation and multiple introductions of invasive giant hogweed in the western Swiss Alps. Mol. Ecol. 2009, 18, 2819–2831. [Google Scholar] [CrossRef]
- Rijal, D.P.; Alm, T.; Jahodová, Š.; Stenøien, H.K.; Alsos, I.G. Reconstructing the invasion history of Heracleum persicum (Apiaceae) into Europe. Mol. Ecol. 2015, 24, 5522–5543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, S.C.H.; Kohn, J.R. Genetic and evolutionary consequences of small population size in plants: Implications for conservation. In Genetics and Conservation of Rare Plants; Falk, D.A., Holsinger, K.E., Eds.; CABI: New York, NY, USA, 1991; p. 283. [Google Scholar]
- Ozerova, N.A.; Krivosheina, M.G. Patterns of secondary range formation for Heracleum sosnowskyi and H. mantegazzianum on the territory of Russia. Russ. J. Biol. Invasions 2018, 9, 155–162. [Google Scholar] [CrossRef]
- Ebel’, A.L.; Strelnikova, T.O.; Kuprijanov, A.N.; Verkhozina, A.V.; Mikhailova, S.I.; Prokopyev, A.S.; Strelnikova, T.O.; Sheremetova, S.A.; Khrustaleva, I.A. Chernaja Kniga flory Sibiri; Vinogradova, J.K., Kuprijanov, A.N., Eds.; Novosibirsk: Geo, Russia, 2016; p. 439. [Google Scholar]
- Kabuce, N.; Priede, A. NOBANIS—Invasive Alien Species Fact Sheet—Heracleum sosnowskyi. In Online Database of the European Network on Invasive Alien Species—NOBANIS; 2010; Available online: https://gd.eppo.int/taxon/HERSO/distribution/LT (accessed on 12 August 2021).
- Estoup, A.; Ravigné, V.; Hufbauer, R.; Vitalis, R.; Gautier, M.; Facon, B. Is TIs There a Genetic Paradox of Biological Invasion?here a Genetic Paradox of Biological Invasion? Annu. Rev. Ecol. Evol. Syst. 2016, 47, 51–72. [Google Scholar] [CrossRef]
- Rijal, D.P.; Alm, T.; Nilsen, L.; Alsos, I.G. Giant invasive Heracleum persicum: Friend or foe of plant diversity? Ecol. Evol. 2017, 7, 4936–4950. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collection Site | Population Type | Number of Samples | Colour of Flowers | Latitude | Longitude |
|---|---|---|---|---|---|
| MBG (Moscow Botany Garden) territory, Russia | Feral, running wild for about five years | 21 | blue, pink, white | 55°83′52.9″ | 37°60′92.0″ |
| Moscow region, Russia | Invasive, grows for decades | 9 | blue, pink, white | 55°38′64.6″ | 39°18′24.1″ |
| Kaluga region, Russia | 8 | blue | 54°50′91.3″ | 36°21′24.6″ | |
| Kostroma region, Russia | 7 | blue | 57°81′45.7″ | 40°98′37.7″ | |
| Smolensk region, Russia | 10 | blue, pink | 54°72′77.5″ | 32°99′65.3″ | |
| Plants grown from the seeds in the greenhouse | ‘Minaret’ variety | 29 | blue, pink, white |
| Collection Site | Population Type | Number of Samples | Colour of Flowers | Latitude | Longitude |
|---|---|---|---|---|---|
| Moscow, Russia | Invasive, grows for decades | 15 | white | 55°57′41.7″ | 37°55′56.2″ |
| Peskovka, Ukraine | 20 | white | 50°70′69.1″ | 29°58′15.3″ | |
| Kiev, Ukraine | 17 | white | 50°46′12.2″ | 30°55′63.1″ | |
| Zhitomir (industrial), Ukraine | 14 | white | 50°24′14.3″ | 28°74′12.8″ | |
| Zhitomir (center), Ukraine | 19 | white | 50°25′29.0″ | 28°65′14.9″ | |
| Caucasus, Russia | native | 14 | white | 43°77′66.0″ | 43°28′72.5″ |
| Collection Site | Kostroma (i) | Smolensk (i) | Moscow (i) | Kaluga (i) | MBG (f) | ‘Minaret’ Variety |
|---|---|---|---|---|---|---|
| Kostroma (i) | 0.000 | |||||
| Smolensk (i) | 0.216 | 0.000 | ||||
| Moscow (i) | 0.209 | 0.260 | 0.000 | |||
| Kaluga (i) | 0.231 | 0.258 | 0.196 | 0.000 | ||
| MBG (f) | 0.269 | 0.284 | 0.198 | 0.230 | 0.000 | |
| ‘Minaret’ Variety | 0.302 | 0.315 | 0.318 | 0.341 | 0.276 | 0.000 |
| Collection Site | RAPD | ISSR | REMAP | RAPD+ISSR+REMAP | ||||
|---|---|---|---|---|---|---|---|---|
| GD | RATIO | GD | RATIO | GD | RATIO | GD | RATIO | |
| Invasive population | 0.322 | 0.253 | 0.328 | 0.294 | ||||
| Feral MBG Population | 0.234 | 1.4 | 0.238 | 1.1 | 0.290 | 1.1 | 0.248 | 1.2 |
| ‘Minaret’ Variety | 0.205 | 1.6 | 0.171 | 1.5 | 0.220 | 1.5 | 0.194 | 1.5 |
| Collection site | Caucasus (n) | Moscow (i) | Kiev (i) | Zhitomir (i) | Pescovka (i) |
|---|---|---|---|---|---|
| Caucasus (n) | 0.000 | ||||
| Moscow (i) | 0.614 | 0.000 | |||
| Kiev (i) | 0.611 | 0.512 | 0.000 | ||
| Zhitomir (i) | 0.591 | 0.530 | 0.493 | 0.000 | |
| Pescovka (i) | 0.569 | 0.562 | 0.466 | 0.438 | 0.000 |
| Populations | RAPD+ISSR+REMAP | |||
|---|---|---|---|---|
| Min GD | Max GD | Mean GD | Difference of GD between Native and Invasive Populations | |
| Moscow (i) | 0.217 | 0.606 | 0.398 | 0.104 |
| Peskovka (i) | 0.224 | 0.678 | 0.418 | 0.084 |
| Kiev (i) | 0.265 | 0.622 | 0.441 | 0.061 |
| Zhitomir (industrial) (i) | 0.216 | 0.640 | 0.446 | 0.056 |
| Zhitomir (center) (i) | 0.248 | 0.629 | 0.457 | 0.045 |
| The average Value of Genetic Diversity within Invasive Populations | 0.234 | 0.635 | 0.432 | 0.070 |
| Caucasus (n) | 0.244 | 0.739 | 0.502 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osipova, E.S.; Stepanova, A.Y.; Tereshonok, D.V.; Gladkov, E.A.; Vysotskaya, O.N. Genetic Diversity in Invasive Populations of Lupinus polyphyllus Lindl. and Heracleum sosnowskyi Manden. Biology 2021, 10, 1094. https://doi.org/10.3390/biology10111094
Osipova ES, Stepanova AY, Tereshonok DV, Gladkov EA, Vysotskaya ON. Genetic Diversity in Invasive Populations of Lupinus polyphyllus Lindl. and Heracleum sosnowskyi Manden. Biology. 2021; 10(11):1094. https://doi.org/10.3390/biology10111094
Chicago/Turabian StyleOsipova, Ekaterina Sergeevna, Anna Yurievna Stepanova, Dmitry Viktorovich Tereshonok, Evgeny Aleksandrovich Gladkov, and Olga Nikolaevna Vysotskaya. 2021. "Genetic Diversity in Invasive Populations of Lupinus polyphyllus Lindl. and Heracleum sosnowskyi Manden." Biology 10, no. 11: 1094. https://doi.org/10.3390/biology10111094