
Population Ecology and Genetic Diversity of the Invasive Alien Species Procambarus clarkii in Lake Trasimeno (Italy)

 , , , , , , and
, , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
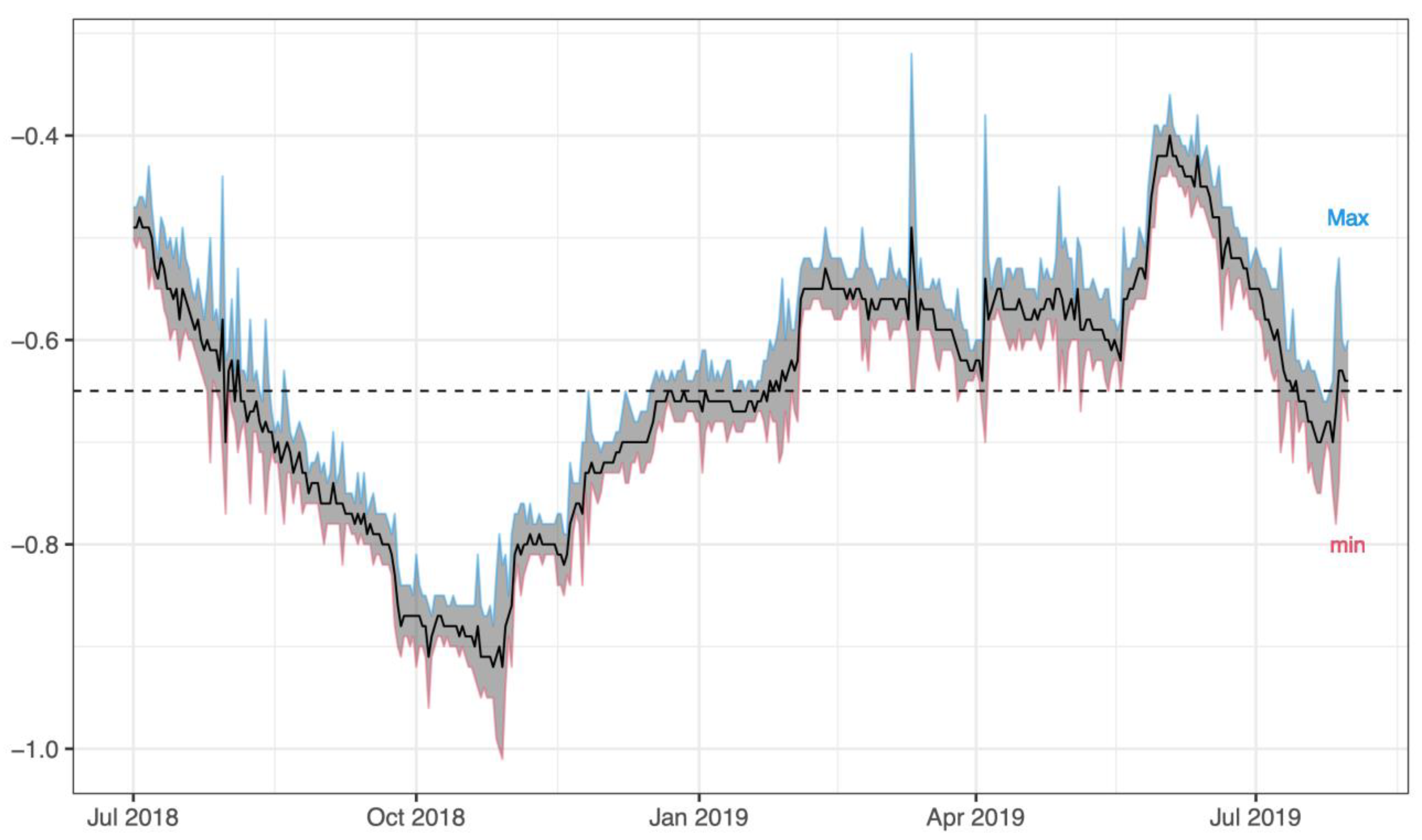
2.2. Chemical-Physical Parameters of Lake Trasimeno
2.3. Sampling of P. clarkii and Morphometric Data Collection
- total body length (from the tip of the rostrum to the posterior margin of the abdomen (TL)
- length of the cephalothorax or carapace (CL)
- total body weight (W)
- abdomen weight (AW)
- hepatopancreas weight (HIw)
- mature gonad weight (for females only)
2.4. Sex Ratio and Condition Indexes
2.5. Statistical Analysis
2.6. Genetic Analysis
3. Results
3.1. Chemical-Physical Parameters of Lake Trasimeno
3.2. Morphometric and Biological Analysis
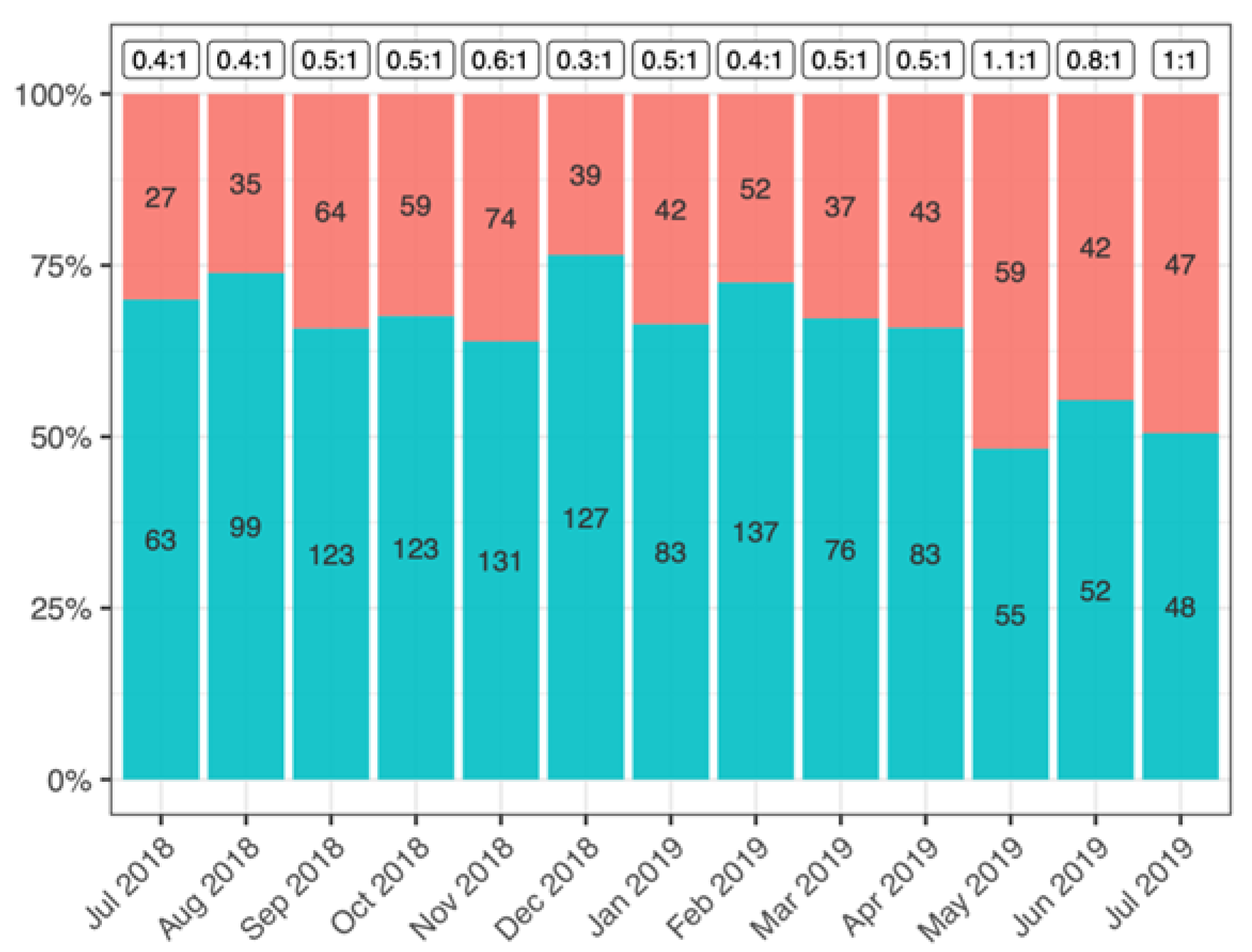
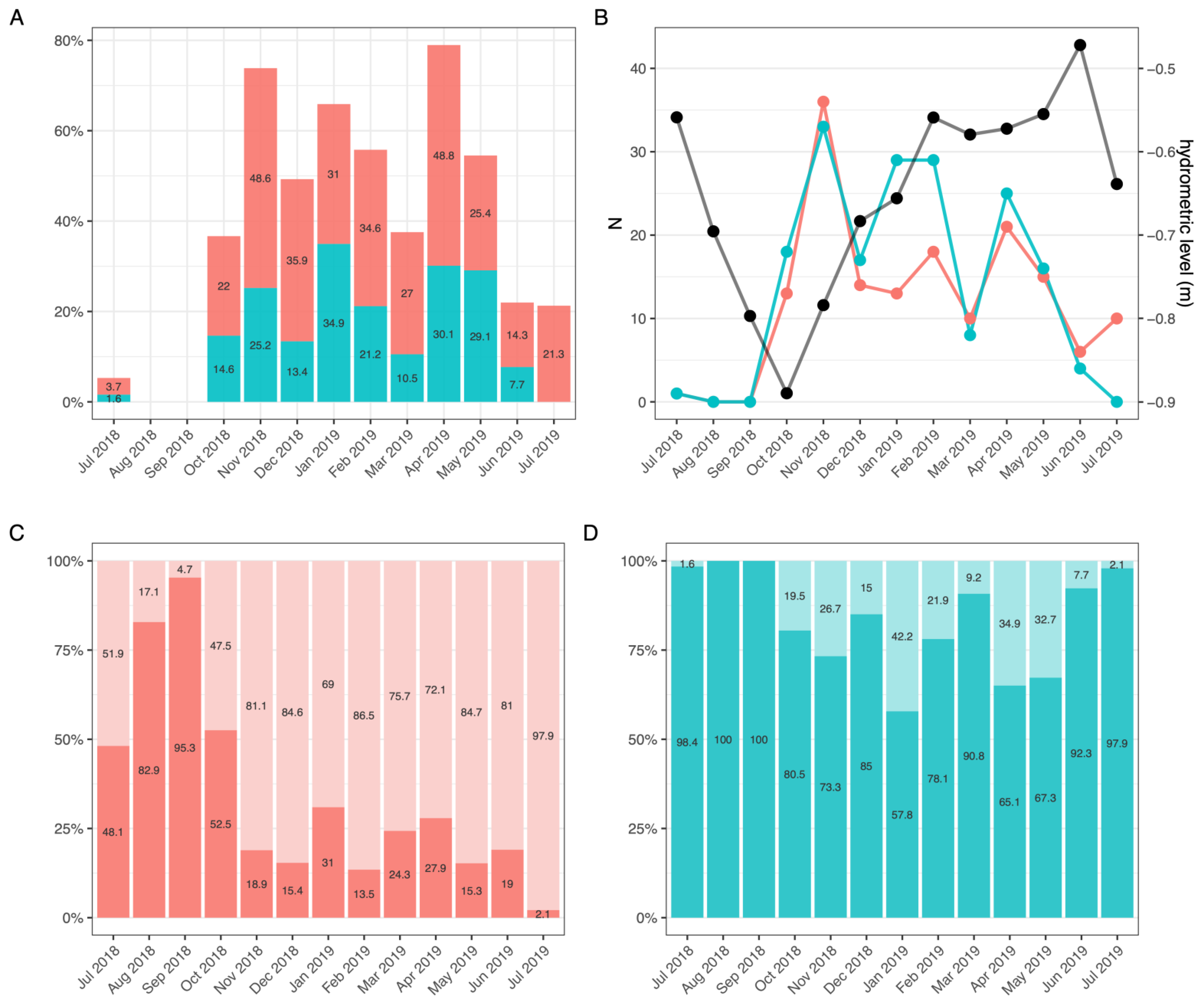
3.2.1. Sex Ratio and Population Size
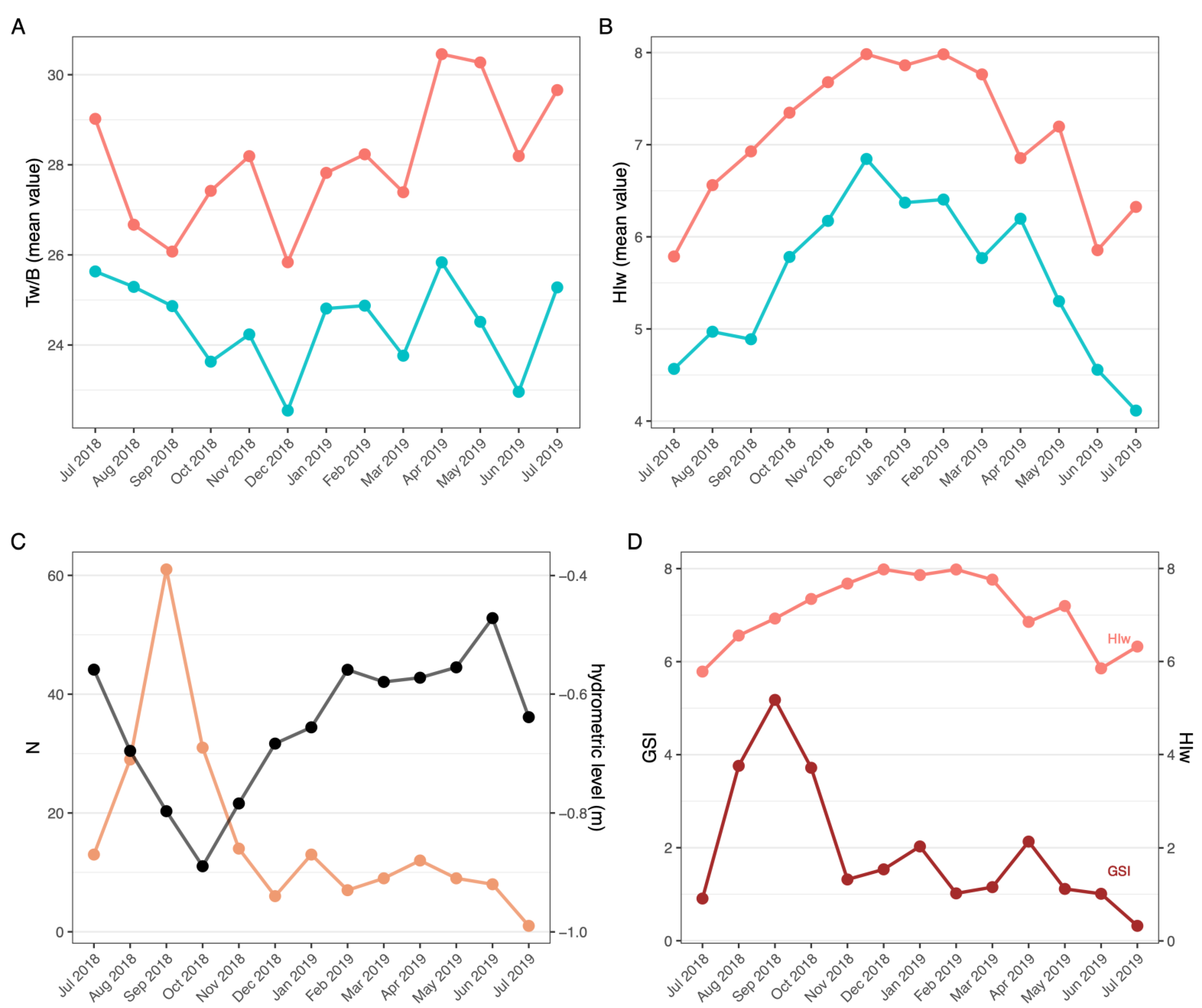
3.2.2. Molting and Reproductive Cycle
3.2.3. Condition Indexes
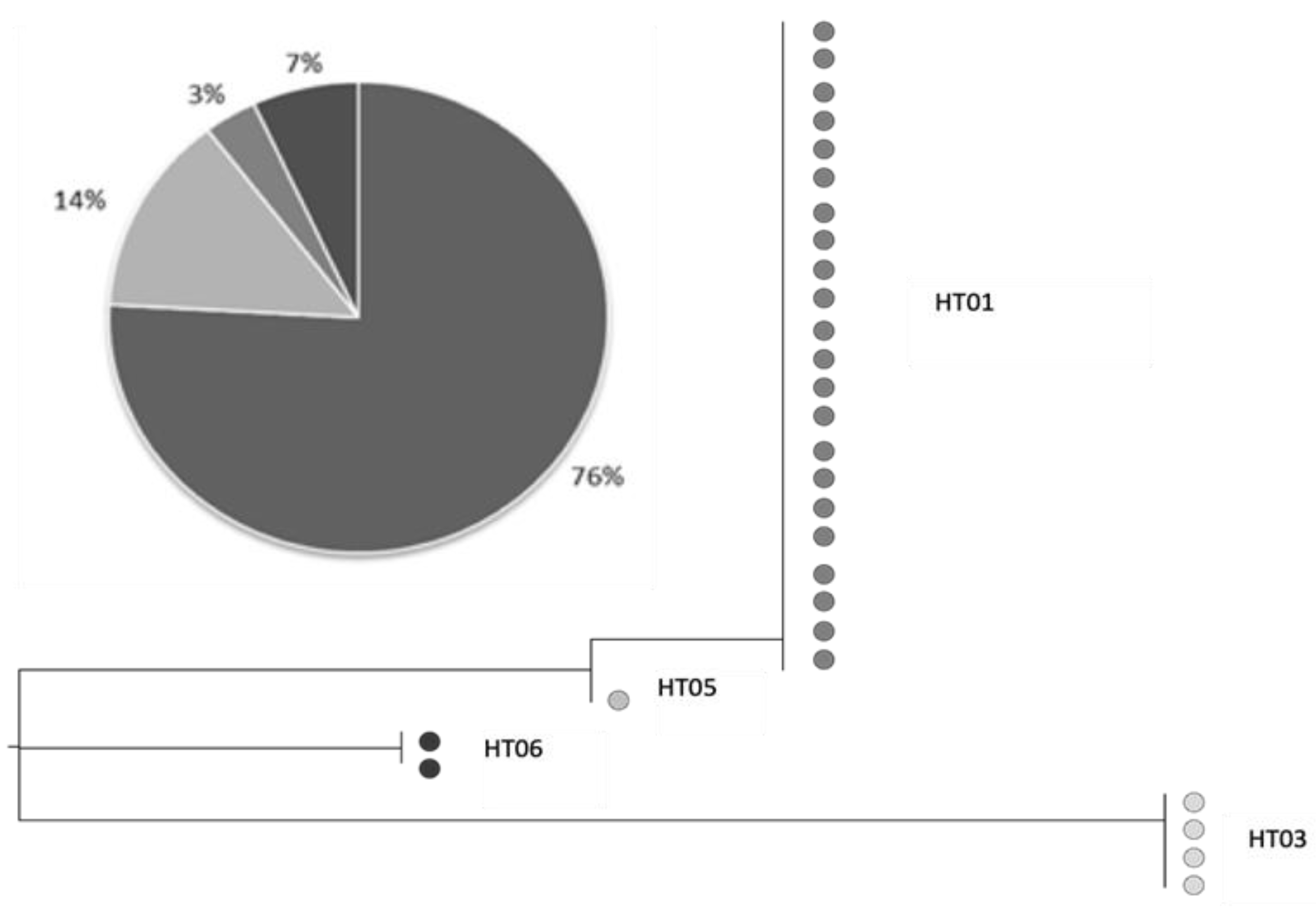
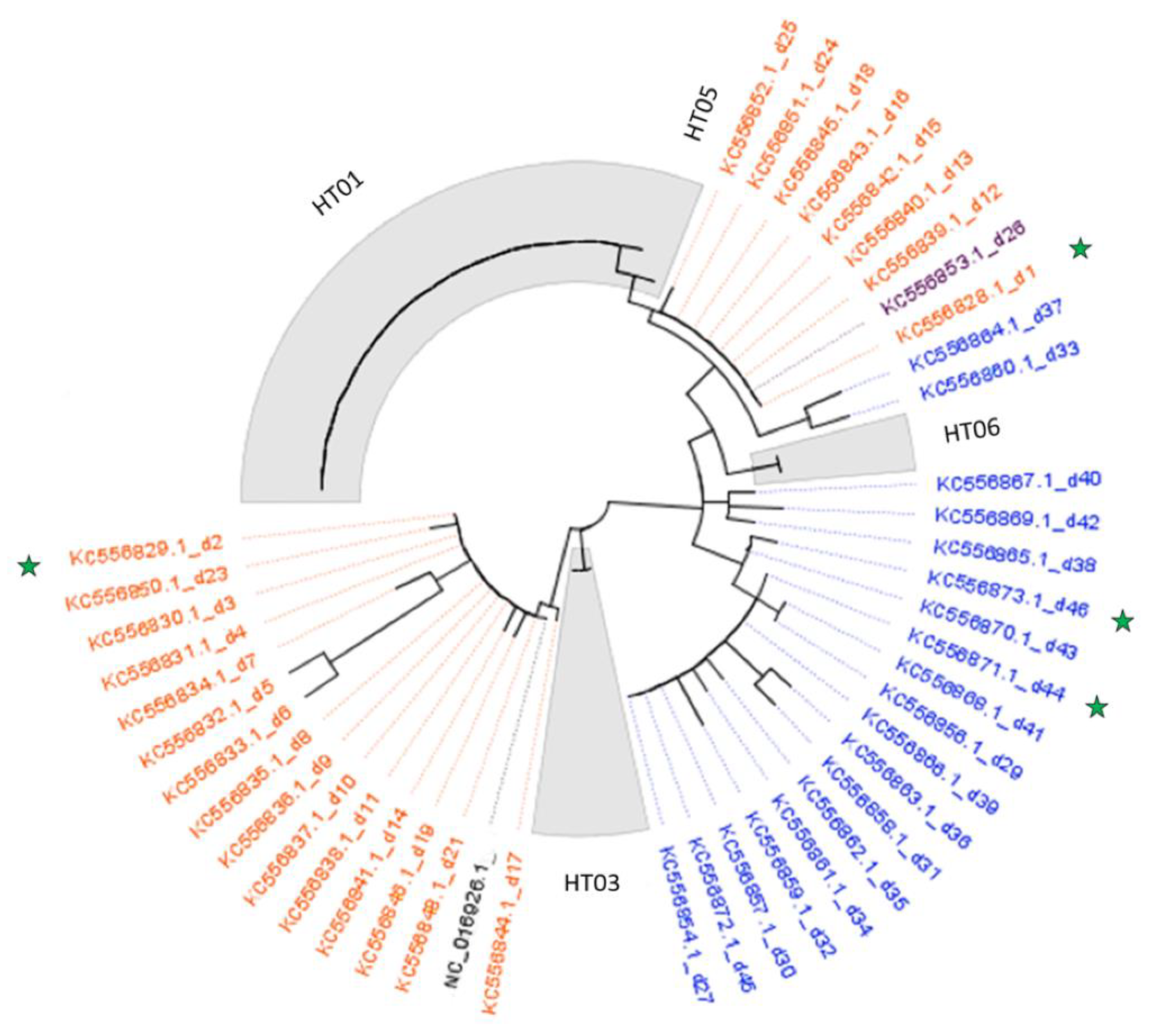
3.3. Genetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hobbs, H.H., Jr. Crayfish Distribution, Adaptive Radiation and Evolution. In Freshwater Crayfish: Biology, Management and Exploitation; Holdich, D.M., Lowery, R.S., Eds.; Croom Helm Ltd.: London, UK, 1988; pp. 52–82. [Google Scholar]
- Gherardi, F. Crayfish Invading Europe: The Case Study of Procambarus clarkii. Mar. Freshw. Behav. Physiol. 2006, 39, 175–191. [Google Scholar] [CrossRef]
- Claire, W.H.; Wroiten, J.W. First Record of the Crayfish, Procambarus clarkii, from Idaho, USA (Decapoda, Cambaridae). Crustaceana 1978, 35, 317–319. [Google Scholar]
- Dörr, A.J.M.; Scalici, M. Revisiting Reproduction and Population Structure and Dynamics of Procambarus clarkii Eight Years after Its Introduction into Lake Trasimeno (Central Italy). Knowl. Manag. Aquat. Ecosyst. 2013, 408, 10. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Q.; Zhang, M.-T.; Wu, Y.; Ding, H.; Lei, J.-C.; Zhu, S.-L.; Zhang, Z.-H.; Chen, L. Prediction of Potential Distribution of the Invasive Species Procambarus clarkii in China Based on Ecological Niche Models. J. Appl. Ecol. 2020, 31, 309–318. [Google Scholar]
- Piersanti, S.; Pallottini, M.; Salerno, G.; Goretti, E.; Elia, A.C.; Dörr, A.J.M.; Rebora, M. Resistance to Dehydration and Positive Hygrotaxis in the Invasive Red Swamp Crayfish Procambarus clarkii. Knowl. Manag. Aquat. Ecosyst. 2018, 419, 36. [Google Scholar] [CrossRef] [Green Version]
- Zanotto, F.P.; Wheatly, M.G. The Effect of Ambient PH on Electrolyte Regulation during the Postmoult Period in Freshwater Crayfish Procambarus clarkii. J. Exp. Biol. 1993, 178, 1–19. [Google Scholar] [CrossRef]
- Dörr, A.J.M.; Scalici, M.; Caldaroni, B.; Magara, G.; Scoparo, M.; Goretti, E.; Elia, A.C. Salinity Tolerance of the Invasive Red Swamp Crayfish Procambarus clarkii (Girard, 1852). Hydrobiologia 2020, 847, 2065–2081. [Google Scholar] [CrossRef]
- Scalici, M.; Chiesa, S.; Scuderi, S.; Celauro, D.; Gibertini, G. Population Structure and Dynamics of Procambarus clarkii (Girard, 1852) in a Mediterranean Brackish Wetland (Central Italy). Biol. Invasions 2010, 12, 1415–1425. [Google Scholar] [CrossRef]
- Elia, A.C.; Dörr, A.J.M.; Mastrangelo, C.; Prearo, M.; Abete, M.C. Glutathione and Antioxidant Enzymes in the Hepatopancreas of Crayfish Procambarus clarkii (Girard, 1852) of Lake Trasimeno (Italy). Bull. Fr. Pêche Piscic. 2006, 380–381, 1351–1361. [Google Scholar] [CrossRef]
- Kouba, A.; Buřič, M.; Kozák, P. Bioaccumulation and Effects of Heavy Metals in Crayfish: A Review. Water Air Soil Pollut. 2010, 211, 5–16. [Google Scholar] [CrossRef]
- Trombini, C.; Kazacova, J.; Montilla-López, A.; Fernández-Cisnal, R.; Hampel, M.; Fernández-Torres, R.; Bello-López, M.Á.; Abril, N.; Blasco, J. Assessment of Pharmaceutical Mixture (Ibuprofen, Ciprofloxacin and Flumequine) Effects to the Crayfish Procambarus clarkii: A Multilevel Analysis (Biochemical, Transcriptional and Proteomic Approaches). Environ. Res. 2021, 200, 111396. [Google Scholar] [CrossRef]
- Alcorlo, P.; Baltanás, A. The Trophic Ecology of the Red Swamp Crayfish (Procambarus clarkii) in Mediterranean Aquatic Ecosystems: A Stable Isotope Study. Limnetica 2013, 32, 121–138. [Google Scholar]
- Mancinelli, G.; Papadia, P.; Ludovisi, A.; Migoni, D.; Bardelli, R.; Fanizzi, F.P.; Vizzini, S. Beyond the Mean: A Comparison of Trace-and Macroelement Correlation Profiles of Two Lacustrine Populations of the Crayfish Procambarus clarkii. Sci. Total Environ. 2018, 624, 1455–1466. [Google Scholar] [CrossRef]
- Dörr, A.J.; Elia, A.C.; Rodolfi, M.; Garzoli, L.; Picco, A.M.; D’Amen, M.; Scalici, M. A Model of Co-Occurrence: Segregation and Aggregation Patterns in the Mycoflora of the Crayfish Procambarus clarkii in Lake Trasimeno (Central Italy). J. Limnol. 2012, 71, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Dörr, A.J.; Rodolfi, M.; Scalici, M.; Elia, A.C.; Garzoli, L.; Picco, A.M. Phoma Glomerata, a Potential New Threat to Italian Inland Waters. J. Nat. Conserv. 2011, 19, 370–373. [Google Scholar] [CrossRef]
- Souty-Grosset, C.; Anastacio, P.M.; Aquiloni, L.; Banha, F.; Choquer, J.; Chucholl, C.; Tricarico, E. The Red Swamp Crayfish Procambarus clarkii in Europe: Impacts on Aquatic Ecosystems and Human Well-Being. Limnologica 2016, 58, 78–93. [Google Scholar] [CrossRef]
- Veroli, M.; Martinoli, M.; Caprioli, R.; Angelici, C.; Pulcini, D.; Capoccioni, F. Population Structure and Dynamics of the Invasive Procambarus clarkii (Girard, 1852) in a Tiber River Ramsar Site, Central Italy. Int. J. Aquat. Biol. 2021, 9, 23–32. [Google Scholar]
- Delmastro, G.B. Sull’acclimatazione Del Gambero Della Louisiana Procambarus clarkii (Girard, 1852) Nelle Acque Dolci Italiane (Crustacea: Decapoda: Cambaridae). Pianura 1992, 4, 5–10. [Google Scholar]
- Dörr, A.J.M.; La Porta, G.; Pedicillo, G.; Lorenzoni, M. Biology of Procambarus clarkii (Girard, 1852) in Lake Trasimeno. Bull. Fr. Pêche Piscic. 2006, 380–381, 1155–1168. [Google Scholar] [CrossRef] [Green Version]
- Frondini, F.; Dragoni, W.; Morgantini, N.; Donnini, M.; Cardellini, C.; Caliro, S.; Melillo, M.; Chiodini, G. An Endorheic Lake in a Changing Climate: Geochemical Investigations at Lake Trasimeno (Italy). Water 2019, 11, 1319. [Google Scholar] [CrossRef] [Green Version]
- Ludovisi, A.; Gaino, E. Meteorological and Water Quality Changes in Lake Trasimeno (Umbria, Italy) during the Last Fifty Years. J. Limnol. 2010, 69, 174. [Google Scholar] [CrossRef] [Green Version]
- Coops, H.; Beklioglu, M.; Crisman, T.L. The Role of Water-Level Fluctuations in Shallow Lake Ecosystems–Workshop Conclusions. Hydrobiologia 2003, 506, 23–27. [Google Scholar] [CrossRef]
- Souty-Grosset, C.; Holdich, D.; Noel, P.; Reynolds, J.D.; Haffner, P. Atlas of Crayfish in Europe; Muséum National d’Histoire Naturelle: Paris, France, 2006; Volume 64, 187p. [Google Scholar]
- Alcorlo, P.; Geiger, W.; Otero, M. Reproductive Biology and Life Cycle of the Invasive Crayfish Procambarus clarkii (Crustacea: Decapoda) in Diverse Aquatic Habitats of South-Western Spain: Implications for Population Control. Fundam. Appl. Limnol. 2008, 173, 197. [Google Scholar] [CrossRef]
- Reynolds, J.D. Growth and Reproduction. In Biology of Freshwater Crayfish; Blackwell Science: Oxford, UK, 2002; pp. 152–191. [Google Scholar]
- Acevedo-Limón, L.; Oficialdegui, F.J.; Sánchez, M.I.; Clavero, M. Historical, Human, and Environmental Drivers of Genetic Diversity in the Red Swamp Crayfish (Procambarus clarkii) Invading the Iberian Peninsula. Freshw. Biol. 2020, 65, 1460–1474. [Google Scholar] [CrossRef]
- Barbaresi, S.; Gherardi, F.; Mengoni, A.; Souty-Grosset, C. Genetics and invasion biology in fresh waters: A pilot study of Procambarus clarkii in Europe. In Biological Invaders in Inland Waters: Profiles, Distribution, and Threats; Springer: Berlin/Heidelberg, Germany, 2007; pp. 381–400. [Google Scholar]
- Loureiro, T.G.; Anastácio, P.M.S.G.; Araujo, P.B.; Souty-Grosset, C.; Almerão, M.P. Red Swamp Crayfish: Biology, Ecology and Invasion-an Overview. Nauplius 2015, 23, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Oficialdegui, F.J.; Sánchez, M.I.; Clavero, M. One Century Away from Home: How the Red Swamp Crayfish Took over the World. Rev. Fish Biol. Fish. 2020, 30, 121–135. [Google Scholar] [CrossRef]
- Oficialdegui, F.J.; Clavero, M.; Sánchez, M.I.; Green, A.J.; Boyero, L.; Michot, T.C.; Klose, K.; Kawai, T.; Lejeusne, C. Unravelling the Global Invasion Routes of a Worldwide Invader, the Red Swamp Crayfish (Procambarus clarkii). Freshw. Biol. 2019, 64, 1382–1400. [Google Scholar] [CrossRef]
- Li, Y.; Wang, W.; Liu, X.; Luo, W.; Zhang, J.; Gul, Y. DNA Extraction from Crayfish Exoskeleton. Indian J. Exp. Biol. 2011, 49, 953–957. [Google Scholar]
- Yi, S.; Li, Y.; Shi, L.; Zhang, L.; Li, Q.; Chen, J. Characterization of Population Genetic Structure of Red Swamp Crayfish, Procambarus clarkii, in China. Sci. Rep. 2018, 8, 5586. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Tang, Z.; Huang, L.; Wang, D.; Lu, Z. Genetic Diversity of Procambarus clarkii Populations Based on Mitochondrial DNA and Microsatellite Markers in Different Areas of Guangxi, China. Mitochondrial DNA Part A 2020, 31, 48–56. [Google Scholar] [CrossRef]
- Quan, A.S.; Pease, K.M.; Breinholt, J.W.; Wayne, R.K. Origins of the Invasive Red Swamp Crayfish (Procambarus clarkii) in the Santa Monica Mountains. Aquat. Invasions 2014, 9, 211–219. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Wickham, H.; François, R.; Henry, L.; Müller, K. Dplyr: A Grammar of Data Manipulation. 2021. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 12 August 2021).
- Wickham, H. Tidyr: Tidy Messy Data. 2021. Available online: https://CRAN.R-project.org/package=tidyr (accessed on 6 July 2021).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Li, Y.; Guo, X.; Chen, L.; Bai, X.; Wei, X.; Zhou, X.; Huang, S.; Wang, W. Inferring Invasion History of Red Swamp Crayfish (Procambarus clarkii) in China from Mitochondrial Control Region and Nuclear Intron Sequences. Int. J. Mol. Sci. 2015, 16, 14623–14639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Park, M.-H.; Jung, J.-H.; Ahn, D.-H.; Sultana, T.; Kim, S.; Park, J.-K.; Choi, H.-G.; Min, G.-S. The Mitochondrial Genomes of Cambaroides Similis and Procambarus clarkii(Decapoda: Astacidea: Cambaridae): The Phylogenetic Implications for Reptantia. Zool. Scr. 2012, 41, 281–292. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A Software for Comprehensive Analysis of DNA Polymorphism Data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Huner, J.V. Procambarus. In Biology of Freshwater Crayfish; Blackwell Science: Oxford, UK, 2002; pp. 541–584. [Google Scholar]
- Oluoch, A.O. Breeding Biology of the Louisiana Red Swamp Crayfish Procambarus clarkii Girard in Lake Naivasha, Kenya. Hydrobiologia 1990, 208, 85–92. [Google Scholar] [CrossRef]
- Vogt, G. Abbreviation of Larval Development and Extension of Brood Care as Key Features of the Evolution of Freshwater Decapoda. Biol. Rev. 2013, 88, 81–116. [Google Scholar] [CrossRef]
- Elia, A.C.; Dörr, A.J.M.; Abete, M.C.; Prearo, M. Seasonal Variability of Detoxificant Response and Heavy Metal Accumulation in Tissues of Both Sexes in Tinca tinca (L.) from Lake Trasimeno. Rev. Fish Biol. Fish. 2010, 20, 425–434. [Google Scholar] [CrossRef]
- Donato, R.; Rollandin, M.; Favaro, L.; Ferrarese, A.; Pessani, D.; Ghia, D. Habitat Use and Population Structure of the Invasive Red Swamp Crayfish Procambarus clarkii (Girard, 1852) in a Protected Area in Northern Italy. Knowl. Manag. Aquat. Ecosyst. 2018, 419, 12. [Google Scholar] [CrossRef] [Green Version]
- Ligas, A. Population Dynamics of Procambarus clarkii (Girard, 1852) (Decapoda, Astacidea, Cambaridae) from Southern Tuscany (Italy). Crustaceana 2008, 81, 601–609. [Google Scholar] [CrossRef] [Green Version]
- Maccarrone, V.; Filiciotto, F.; Buffa, G.; Di Stefano, V.; Quinci, E.M.; de Vincenzi, G.; Mazzola, S.; Buscaino, G. An Invasive Species in a Protected Area of Southern Italy: The Structure, Dynamics and Spatial Distribution of the Crayfish Procambarus clarkii. Turk. J. Fish. Aquat. Sci. 2016, 16, 401–412. [Google Scholar] [CrossRef]
- Gherardi, F.; Tricarico, E.; Ilhéu, M. Movement Patterns of an Invasive Crayfish, Procambarus clarkii, in a Temporary Stream of Southern Portugal. Ethol. Ecol. Evol. 2002, 14, 183–197. [Google Scholar] [CrossRef]
- Gherardi, F.; Barbaresi, S.; Salvi, G. Spatial and Temporal Patterns in the Movement of Procambarus clarkii, an Invasive Crayfish. Aquat. Sci. 2000, 62, 179–193. [Google Scholar] [CrossRef]
- Gherardi, F.; Barbaresi, S. Invasive Crayfish: Activity Patterns of Procambarus clarkii in the Rice Fields of the Lower Guadalquivir (Spain). Arch. Für Hydrobiol. 2000, 150, 153–168. [Google Scholar] [CrossRef]
- Bravo, M.A.; Duarte, C.M.; Montes, C. Environmental Factors Controlling the Life History of Procambarus clarkii (Decapoda, Cambaridae) in a Temporary Marsh of the Doñana National Park (SW Spain). Int. Ver. Für Theor. Angew. Limnol. Verh. 1994, 25, 2450–2453. [Google Scholar] [CrossRef]
- Huner, J.V. Procambarus in North America and Elsewhere. In Freshwater Crayfish: Biology, Management and Exploitation; Holdich, D.M., Lowery, R.S., Eds.; Croom Helm Ltd.: London, UK, 1988; pp. 239–261. [Google Scholar]
- Penn, G.H. A Story of the Life History of the Louisiana Crayfish Procambarus clarkii (Girard). Ecology 1943, 24, 1–18. [Google Scholar] [CrossRef]
- Guerra, J.L.; Niño, A.E. Ecology of Red Swamp Crayfish (Procambarus clarkii, Girard) in the Central Meseta of Spain. Freshw. Crayfish 1995, 8, 179–200. [Google Scholar]
- Dalosto, M.M.; Palaoro, A.V.; Souty-Grosset, C.; de Siqueira Bueno, S.L.; Loureiro, T.G.; Almerão, M.P.; de Araujo, P.B.; Santos, S. One Step Ahead of the Enemy: Investigating Aggressive Interactions between Invasive and Native Crayfish before the Contact in Nature. Biol. Invasions 2015, 17, 3503–3515. [Google Scholar] [CrossRef]
- Hanshew, B.A.; Garcia, T.S. Invasion of the Shelter Snatchers: Behavioural Plasticity in Invasive Red Swamp Crayfish, Procambarus clarkii. Freshw. Biol. 2012, 57, 2285–2296. [Google Scholar] [CrossRef]
- Dörr, A.J.M.; Abete, M.C.; Prearo, M.; Pacini, N.; La Porta, G.; Natali, M.; Elia, A.C. Effects of Selenium Supplemented Diets on Growth and Condition Indexes in Juvenile Red Swamp Crayfish, Procambarus clarkii. Environ. Toxicol. Pharmacol. 2013, 36, 484–492. [Google Scholar] [CrossRef]
- Evans, L.H.; Jussila, J. Freshwater Crayfish Growth under Culture Conditions: Proposition for a Standard Reporting Approach. J. World Aquac. Soc. 1997, 28, 11–19. [Google Scholar] [CrossRef]
- Barbaresi, S.; Fani, R.; Gherardi, F.; Mengoni, A.; Souty-Grosset, C. Genetic Variability in European Populations of an Invasive American Crayfish: Preliminary Results. Biol. Invasions 2003, 5, 269–274. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Summer | Autumn | Winter | Spring |
|---|---|---|---|---|
| water temperature (°C) | 24.54 ± 4.34 | 10.52 ± 5.35 | 7.80 ± 2.94 | 21.17 ± 5.15 |
| water transparency (m) | 0.60 ± 0.10 | 0.52 ± 0.24 | 0.50 ± 0.17 | 0.83 ± 0.15 |
| pH | 8.54 ± 0.15 | 8.34 ± 0.06 | 8.42 ± 0.12 | 8.57 ± 0.19 |
| dissolved oxygen (mg/L) | 8.4 ± 1.02 | 10.42 ± 1.61 | 11.20 ± 1.02 | 8.13 ± 0.98 |
| oxygen saturation (%) | 97.12 ± 19.91 | 92.83 ± 3.58 | 94.00 ± 1.81 | 91.23 ± 3.83 |
| conductivity (µS/cm) | 1569.75 ± 116.80 | 1143.67 ± 192.55 | 1032.10 ± 89.06 | 1425.00 ± 167.82 |
| Total Length (mm) | Carapace Length (mm) | Total Weight (g) | Abdomen Weight (g) | Hepatopancreas Weight(g) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Month | Females | Males | Females | Males | Females | Males | Females | Males | Females | Males |
| July 2018 | 107.02 ± 14.60 | 90.65 ± 12.14 | 52.76 ± 9.42 | 48.45 ± 5.71 | 31.50 ± 13.32 | 26.05 ± 9.60 | 8.56 ± 2.51 | 6.46 ± 1.85 | 1.76 ± 0.66 | 1.18 ± 0.49 |
| August 2018 | 99.27 ± 11.39 | 88.34 ± 8.38 | 50.20 ± 7.04 | 43.80 ± 5.24 | 24.76 ± 10.44 | 19.37 ± 5.97 | 6.40 ± 2.10 | 4.80 ± 1.17 | 1.47 ± 0.55 | 0.95 ± 0.31 |
| September 2018 | 94.88 ± 9.15 | 87.55 ± 8.78 | 47.38 ± 4.51 | 43.31 ± 4.56 | 20.83 ± 6.71 | 17.60 ± 5.52 | 5.38 ± 1.58 | 4.31 ± 1.22 | 1.43 ± 0.50 | 0.84 ± 0.25 |
| October 2018 | 94.77 ± 11.50 | 92.88 ± 9.34 | 46.97 ± 5.46 | 46.25 ± 5.23 | 20.22 ± 7.75 | 21.50 ± 7.60 | 5.45 ± 2.00 | 4.92 ± 1.37 | 1.50 ± 0.76 | 1.20 ± 0.43 |
| November 2018 | 90.66 ± 12.31 | 91.49 ± 10.61 | 44.59 ± 6.38 | 45.76 ± 5.82 | 17.81 ± 8.33 | 20.56 ± 9.26 | 4.83 ± 1.88 | 4.72 ± 1.56 | 1.37 ± 0.87 | 1.25 ± 0.60 |
| December 2018 | 99.55 ± 13.22 | 94.99 ± 8.86 | 48.90 ± 6.70 | 47.50 ± 4.93 | 23.66 ± 10.39 | 23.63 ± 8.18 | 5.94 ± 2.24 | 5.13 ± 1.29 | 1.87 ± 1.01 | 1.60 ± 0.62 |
| January 2019 | 93.26 ± 10.81 | 91.47 ± 10.40 | 45.43 ± 5.85 | 45.31 ± 5.66 | 18.71 ± 7.77 | 20.07 ± 9.05 | 5.01 ± 1.70 | 4.75 ± 1.63 | 1.45 ± 0.66 | 1.24 ± 0.50 |
| February 2019 | 93.00 ± 10.25 | 92.29 ± 10.56 | 45.63 ± 5.47 | 45.92 ± 5.69 | 18.96 ± 7.98 | 20.75 ± 9.33 | 5.15 ± 1.67 | 4.82 ± 1.48 | 1.53 ± 0.87 | 1.31 ± 0.61 |
| March 2019 | 103.70 ± 13.71 | 99.08 ± 9.04 | 50.24 ± 7.10 | 48.62 ± 5.02 | 25.85 ± 11.91 | 24.54 ± 7.99 | 6.70 ± 2.42 | 5.67 ± 1.56 | 1.97 ± 0.95 | 1.40 ± 0.48 |
| April 2019 | 99.08 ± 12.15 | 97.98 ± 8.45 | 47.81 ± 6.49 | 47.92 ± 4.67 | 21.28 ± 9.35 | 22.69 ± 8.48 | 6.16 ± 2.17 | 5.57 ± 1.41 | 1.46 ± 0.76 | 1.34 ± 0.37 |
| May 2019 | 103.53 ± 12.93 | 103.03 ± 9.59 | 50.00 ± 6.57 | 50.70 ± 5.36 | 24.01 ± 9.62 | 27.97 ± 10.33 | 7.04 ± 2.52 | 6.41 ± 1.62 | 1.73 ± 0.78 | 1.43 ± 0.47 |
| June 2019 | 110.83 ± 10.40 | 106.92 ± 8.77 | 54.06 ± 5.31 | 53.02 ± 5.47 | 31.14 ± 10.40 | 33.13 ± 11.24 | 8.49 ± 2.36 | 7.24 ± 1.79 | 1.79 ± 0.63 | 1.47 ± 0.51 |
| July 2019 | 111.69 ± 9.86 | 102.64 ± 9.89 | 54.33 ± 5.61 | 50.22 ± 5.52 | 30.53 ± 9.45 | 27.79 ± 9.47 | 8.84 ± 2.24 | 6.79 ± 1.72 | 1.94 ± 0.71 | 1.13 ± 0.41 |
| Sample ID | Mutations (Range: nps 4717–5434) | Haplotype Code |
|---|---|---|
| TR01 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR02 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR03 | 4752 4771 4865 4880 5033 5056 5334 5335 5363 5370 5418 | HT05 |
| TR04 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR05 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR06 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR07 | 4752 4771 4986 5033 5056 5334 5335 5352 5363 5370 | HT06 |
| TR08 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR09 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR10 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR11 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR12 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR13 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR14 | 4752 4771 4986 5033 5056 5334 5335 5352 5363 5370 | HT06 |
| TR15 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR16 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR17 | 5254 5334 5335 5363 | HT03 |
| TR18 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR19 | 5254 5334 5335 5363 | HT03 |
| TR20 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR21 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR22 | 5254 5334 5335 5363 | HT03 |
| TR23 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR24 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR25 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR26 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR27 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR28 | 4752 4771 4865 4880 5033 5041 5056 5334 5335 5363 5370 5418 | HT01 |
| TR29 | 5254 5334 5335 5363 | HT03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dörr, A.J.M.; Scoparo, M.; Cardinali, I.; La Porta, G.; Caldaroni, B.; Magara, G.; Pallottini, M.; Selvaggi, R.; Cenci-Goga, B.; Goretti, E.; et al. Population Ecology and Genetic Diversity of the Invasive Alien Species Procambarus clarkii in Lake Trasimeno (Italy). Biology 2021, 10, 1059. https://doi.org/10.3390/biology10101059
Dörr AJM, Scoparo M, Cardinali I, La Porta G, Caldaroni B, Magara G, Pallottini M, Selvaggi R, Cenci-Goga B, Goretti E, et al. Population Ecology and Genetic Diversity of the Invasive Alien Species Procambarus clarkii in Lake Trasimeno (Italy). Biology. 2021; 10(10):1059. https://doi.org/10.3390/biology10101059
Chicago/Turabian StyleDörr, Ambrosius Josef Martin, Melissa Scoparo, Irene Cardinali, Gianandrea La Porta, Barbara Caldaroni, Gabriele Magara, Matteo Pallottini, Roberta Selvaggi, Beniamino Cenci-Goga, Enzo Goretti, and et al. 2021. "Population Ecology and Genetic Diversity of the Invasive Alien Species Procambarus clarkii in Lake Trasimeno (Italy)" Biology 10, no. 10: 1059. https://doi.org/10.3390/biology10101059