
Energy-Saving LED Light Affects the Efficiency of the Photosynthetic Apparatus and Carbohydrate Content in Gerbera jamesonii Bolus ex Hook. f. Axillary Shoots Multiplied In Vitro




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Data Collection
2.3. Statistical Analysis
3. Results
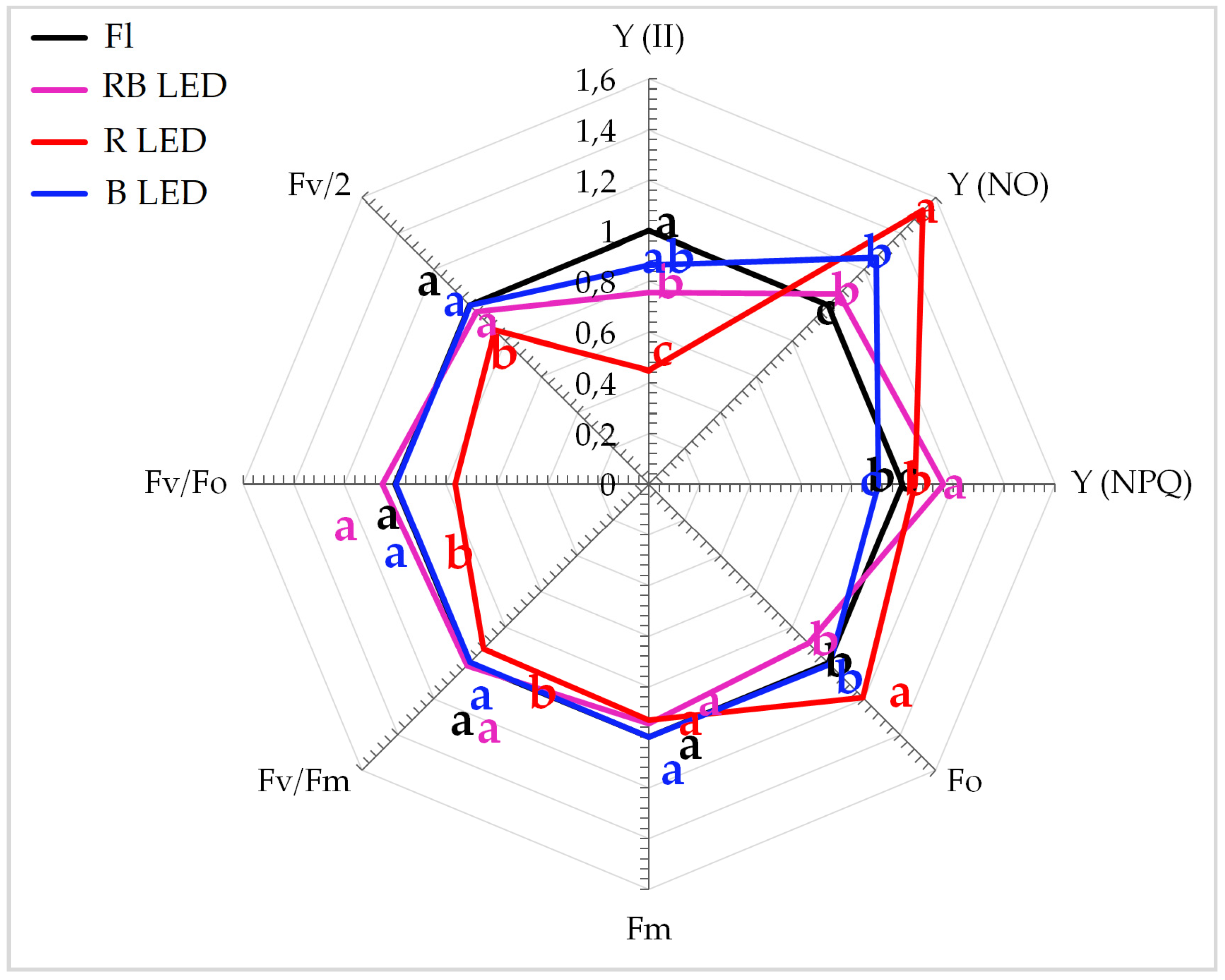
3.1. Photosynthetic Apparatus
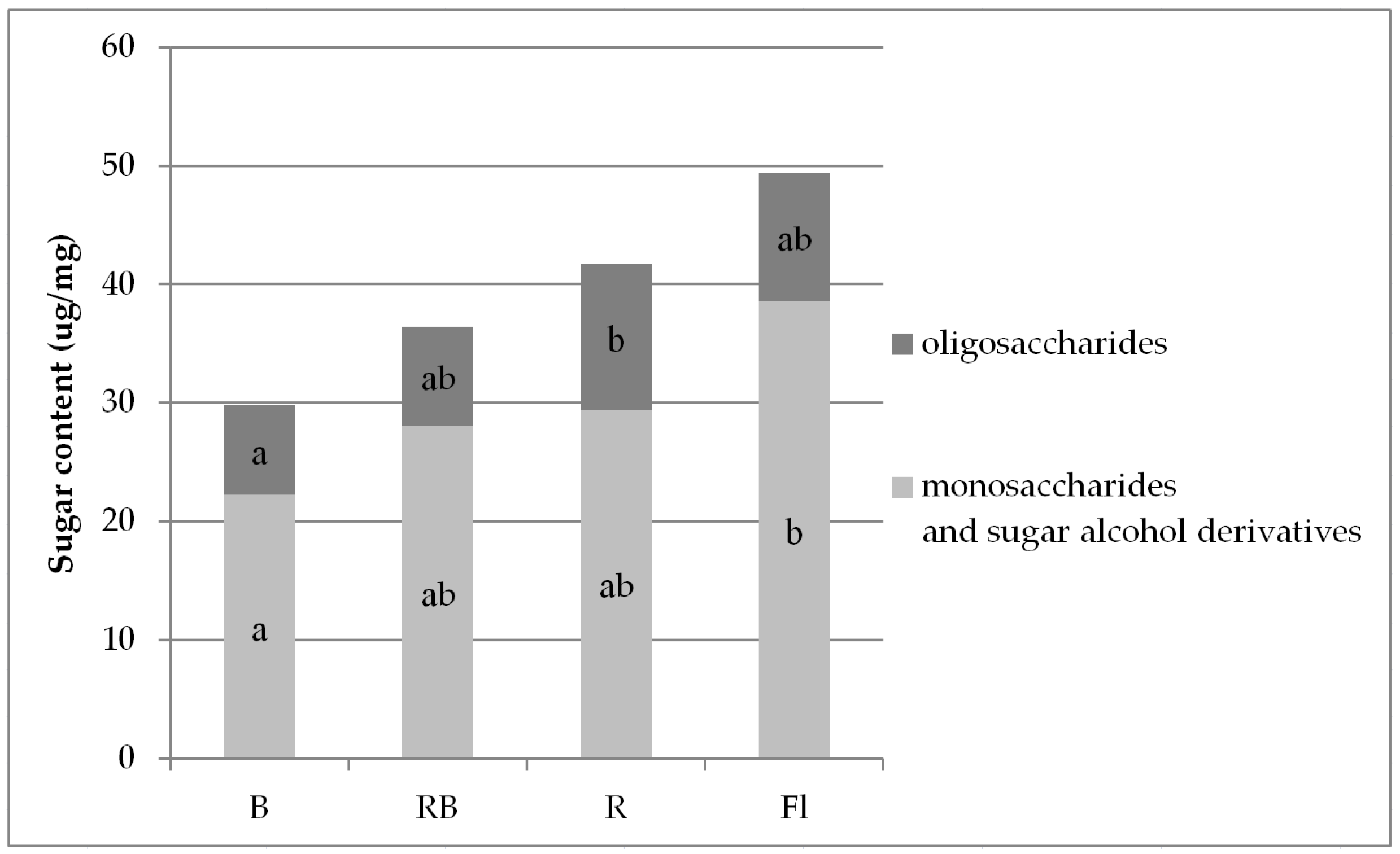
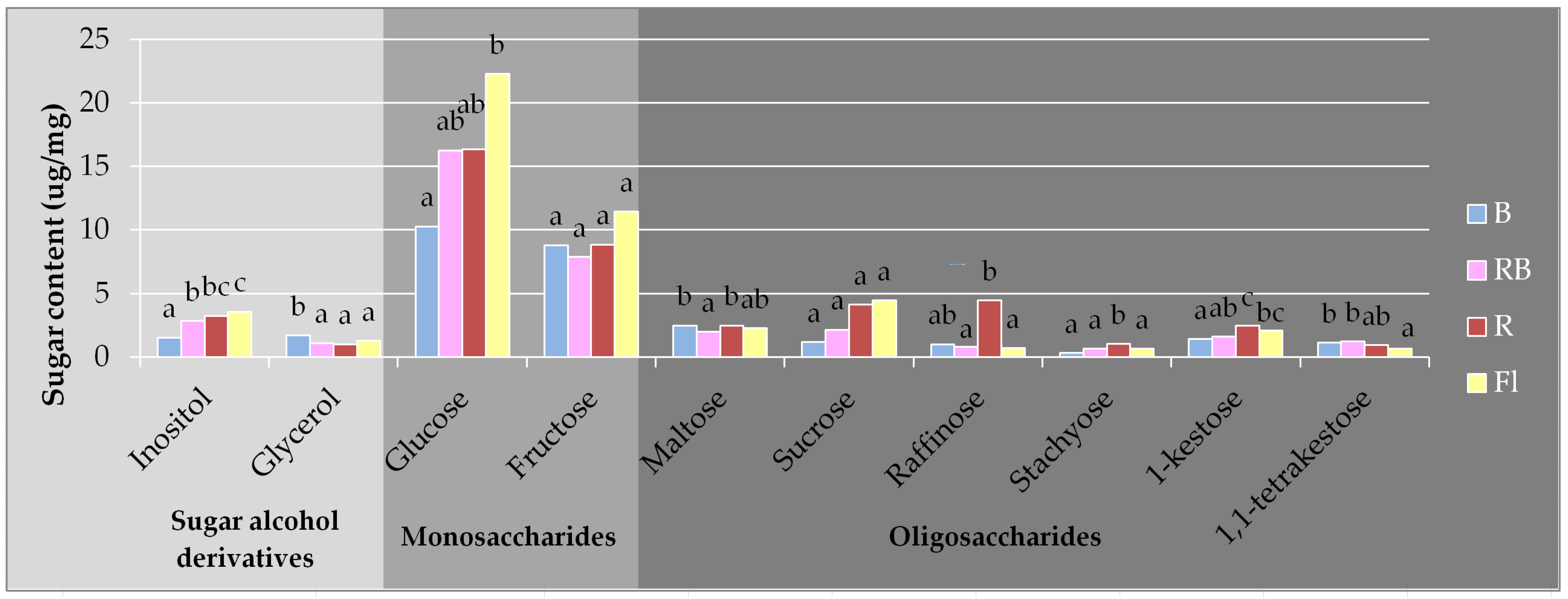
3.2. Carbohydrates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reinten, E.Y.; Coetzee, J.H.; van Wyk, B.-E. The potential of South African indigenous plants for the international cut flower trade. S. Afr. J. Bot. 2011, 77, 934–946. [Google Scholar] [CrossRef]
- Sudhagar, S.; Com, M.; Phil, M. Production and marketing of cut flower (rose and gerbera) in Hosur Taluk. Int. J. Bus. Manag. Invent. 2013, 2, 15–25. [Google Scholar]
- Jain, S.M.; Ochatt, S.J. (Eds.) Protocols for In Vitro Propagation of Ornamental Plants; Humana Press: Totowa, NJ, USA, 2010; p. 400. [Google Scholar] [CrossRef]
- Gupta, S.D.; Jatothu, B. Fundamentals and applications of light-emitting diodes (LEDs) in in vitro plant growth and morphogenesis. Plant Biotechnol. Rep. 2013, 7, 211–220. [Google Scholar] [CrossRef]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: A review. J. Sci. Food Agric. 2014, 95, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Viršilė, A.; Olle, M.; Duchovskis, P. LED Lighting in Horticulture. In Light Emitting Diodes for Agriculture; Gupta, S., Ed.; Springer Nature: Singapore, 2017; pp. 113–147. [Google Scholar] [CrossRef]
- George, E.F.; Hall, M.A.; De Klerk, G.-J. Effects of the physical environment. In Plant Propagation by Tissue Culture 3rd Edition. Volume 1. The Background; George, E.F., Davies, W., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 423–464. ISBN 9781402050046. [Google Scholar]
- Ouzounis, T.; Giday, H.; Kjaer, K.H.; Ottosen, C.-O. LED or HPS in ornamentals? A case study in roses and campanulas. Eur. J. Hortic. Sci. 2018, 83, 166–172. [Google Scholar] [CrossRef]
- Cioć, M.; Pawłowska, B. Leaf response to different light spectrum compositions during micropropagation of Gerbera axillary shoots. Agronomy 2020, 10, 1832. [Google Scholar] [CrossRef]
- Miler, N.; Kulus, D.; Woźny, A.; Rymarz, D.; Hajzer, M.; Wierzbowski, K.; Nelke, R.; Szeffs, L. Application of wide-spectrum light-emitting diodes in micropropagation of popular ornamental plant species: A study on plant quality and cost reduction. Vitr. Cell. Dev. Biol.—Plant 2019, 55, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Pawłowska, B.; Cioć, M.; Prokopiuk, B. How LED light rooting in vitro affected Gerbera acclimatization efficiency. Acta Hortic. 2018, 583–590. [Google Scholar] [CrossRef]
- Pawłowska, B.; Żupnik, M.; Szewczyk-Taranek, B.; Cioć, M. Impact of LED light sources on morphogenesis and levels of photosynthetic pigments in Gerbera jamesonii grown in vitro. Hortic. Environ. Biotechnol. 2018, 59, 115–123. [Google Scholar] [CrossRef]
- Gök, K.M.; Şan, B.; Bayhan, A.K. Micropropagation of gerbera (Gerbera jamesonii Bolus) under different color of light-emitting diodes. J. Nat. Appl. Sci. 2016, 20, 468–474. [Google Scholar] [CrossRef] [Green Version]
- Tarkowski, Ł.P.; Van de Poel, B.; Höfte, M.; Van den Ende, W. Sweet immunity: Inulin boosts resistance of lettuce (Lactuca sativa) against grey mold (Botrytis cinerea) in an ethylene-dependent manner. Int. J. Mol. Sci. 2019, 20, 1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labudda, M.; Tokarz, K.; Tokarz, B.; Muszyńska, E.; Gietler, M.; Górecka, M.; Różańska, E.; Rybarczyk-Płońska, A.; Fidler, J.; Prabucka, B.; et al. Reactive oxygen species metabolism and photosynthetic performance in leaves of Hordeum vulgare plants co-infested with Heterodera filipjevi and Aceria tosichella. Plant Cell Rep. 2020, 39, 1719–1741. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 2016, 7, 250. [Google Scholar] [CrossRef] [Green Version]
- Formela-Luboińska, M.; Chadzinikolau, T.; Drzewiecka, K.; Jeleń, H.; Bocianowski, J.; Kęsy, J.; Labudda, M.; Jeandet, P.; Morkunas, I. The role of sugars in the regulation of the level of endogenous signaling molecules during defense response of yellow lupine to Fusarium oxysporum. Int. J. Mol. Sci. 2020, 21, 4133. [Google Scholar] [CrossRef] [PubMed]
- Eveland, A.L.; Jackson, D.P. Sugars, signalling, and plant development. J. Exp. Bot. 2012, 63, 3367–3377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evich, M.; Spring-Connell, A.M.; Germann, M.W. Impact of modified ribose sugars on nucleic acid conformation and function. Heterocycl. Commun. 2017, 23, 155–165. [Google Scholar] [CrossRef]
- Alén, R. Carbohydrate Chemistry: Fundamentals and Applications; World Scientific Publishing Company: Singapore, 2018. [Google Scholar]
- León, P.; Sheen, J. Sugar and hormone connections. Trends Plant Sci. 2003, 8, 110–116. [Google Scholar] [CrossRef]
- Gibson, S.I. Sugar and phytohormone response pathways: Navigating a signalling network. J. Exp. Bot. 2004, 55, 253–264. [Google Scholar] [CrossRef]
- Francis, D.; Halford, N.G. Nutrient sensing in plant meristems. Plant Mol. Biol. 2006, 60, 981–993. [Google Scholar] [CrossRef]
- Eckstein, A.; Zieba, P.; Gabryś, H. Sugar and light effects on the condition of the photosynthetic apparatus of Arabidopsis thaliana cultured in vitro. J. Plant Growth Regul. 2012, 31, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Desjardisn, Y. Photosynthesis in vitro—on the factor regulationg CO2 assimilation in micropropagation systems. Acta Hortic. 1995, 393, 45–62. [Google Scholar] [CrossRef]
- Sáez, P.L.; Bravo, L.A.; Latsague, M.I.; Sánchez, M.E.; Ríos, D.G. Increased light intensity during in vitro culture improves water loss control and photosynthetic performance of Castanea sativa grown in ventilated vessels. Sci. Hortic. 2012, 138, 7–16. [Google Scholar] [CrossRef]
- Hdider, C.; Desjardins, Y. Effects of sucrose on photosynthesis and phosphoenolpyruvate carboxylase activity of in vitro cultured strawberry plantlets. Plant Cell. Tissue Organ Cult. 1994, 36, 27–33. [Google Scholar] [CrossRef]
- Van Huylenbroeck, J.M.; Debergh, P.C. Impact of sugar concentration in vitro on photosynthesis and carbon metabolism during ex vitro acclimatization of Spathiphyllum plantlets. Physiol. Plant. 1996, 96, 298–304. [Google Scholar] [CrossRef]
- Rybczyński, J.J.; Borkowska, B.; Fiuk, A.; Gawrońska, H.; Śliwińska, E.; Mikuła, A. Effect of sucrose concentration on photosynthetic activity of in vitro cultures Gentiana kurroo (Royle) germlings. Acta Physiol. Plant. 2007, 29, 445–453. [Google Scholar] [CrossRef]
- Xiao, Y.; Niu, G.; Kozai, T. Development and application of photoautotrophic micropropagation plant system. Plant Cell. Tissue Organ Cult. 2011, 105, 149–158. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, D.; Ma, Z.; Li, X.; Xiao, Y. Growth and photosynthetic capability of Momordica grosvenori plantlets grown photoautotrophically in response to light intensity. HortScience 2009, 44, 757–763. [Google Scholar] [CrossRef]
- Xue, X.; Wang, Q.; Qu, Y.; Wu, H.; Dong, F.; Cao, H.; Wang, H.L.; Xiao, J.; Shen, Y.; Wan, Y. Development of the photosynthetic apparatus of Cunninghamia lanceolata in light and darkness. New Phytol. 2017, 213, 300–313. [Google Scholar] [CrossRef] [PubMed]
- Tokarz, K.M.; Wesołowski, W.; Tokarz, B.; Makowski, W.; Wysocka, A.; Jędrzejczyk, R.J.; Chrabaszcz, K.; Malek, K.; Kostecka-Gugała, A. Stem photosynthesis—A key element of grass pea (Lathyrus sativus L.) acclimatisation to salinity. Int. J. Mol. Sci. 2021, 22, 685. [Google Scholar] [CrossRef]
- Zheng, J.; Hu, M.-J.; Guo, Y.-P. Regulation of photosynthesis by light quality and its mechanism in plants. Ying Yong Sheng Tai Xue Bao = J. Appl. Ecol. 2008, 19, 1619–1624. [Google Scholar]
- Gururani, M.A.; Venkatesh, J.; Tran, L.S.P. Regulation of photosynthesis during abiotic stress-induced photoinhibition. Mol. Plant 2015, 8, 1304–1320. [Google Scholar] [CrossRef] [Green Version]
- Faraloni, C.; Cutino, I.; Petruccelli, R.; Leva, A.R.; Lazzeri, S.; Torzillo, G. Chlorophyll fluorescence technique as a rapid tool for in vitro screening of olive cultivars (Olea europaea L.) tolerant to drought stress. Environ. Exp. Bot. 2011, 73, 49–56. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Roeber, V.M.; Bajaj, I.; Rohde, M.; Schmülling, T.; Cortleven, A. Light acts as a stressor and influences abiotic and biotic stress responses in plants. Plant Cell Environ. 2021, 44, 645–664. [Google Scholar] [CrossRef]
- Sheen, J. Feedback control of gene expression. Photosynth. Res. 1994, 39, 427–438. [Google Scholar] [CrossRef]
- Koch, K.E. Carbohydrate-modulated gene expression in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 509–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, L.C.; Osório, M.L.; Chaves, M.M.; Amâncio, S. Chlorophyll fluorescence as an indicator of photosynthetic functioning of in vitro grapevine and chestnut plantlets under ex vitro acclimatization. Plant Cell. Tissue Organ Cult. 2001, 67, 271–280. [Google Scholar] [CrossRef]
- Woo, N.S.; Badger, M.R.; Pogson, B.J. A rapid, non-invasive procedure for quantitative assessment of drought survival using chlorophyll fluorescence. Plant Methods 2008, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Piwowarczyk, B.; Tokarz, K.; Muszyńska, E.; Makowski, W.; Jędrzejczyk, R.; Gajewski, Z.; Hanus-Fajerska, E. The acclimatization strategies of kidney vetch (Anthyllis vulneraria L.) to Pb toxicity. Environ. Sci. Pollut. Res. 2018, 25, 19739–19752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tabacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Bolhar-Nordenkampf, H.R.; Long, S.P.; Baker, N.R.; Oquist, G.; Schreiber, U.; Lechner, E.G. Chlorophyll fluorescence as a probe of the photosynthetic competence of leaves in the field: A review of current instrumentation. Funct. Ecol. 1989, 3, 497–514. [Google Scholar] [CrossRef]
- Jiang, H.X.; Chen, L.S.; Zheng, J.G.; Han, S.; Tang, N.; Smith, B.R. Aluminum-induced effects on Photosystem II photochemistry in Citrus leaves assessed by the chlorophyll a fluorescence transient. Tree Physiol. 2008, 28, 1863–1871. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, H.M.; Govindjee; Bosa, K.; Kościelniak, J.; Zuk-Gołaszewska, K. Effects of salt stress on photosystem II efficiency and CO2 assimilation of two Syrian barley landraces. Environ. Exp. Bot. 2011, 73, 64–72. [Google Scholar] [CrossRef]
- Tokarz, K.M.; Makowski, W.; Tokarz, B.; Hanula, M.; Sitek, E.; Muszyńska, E.; Jędrzejczyk, R.; Banasiuk, R.; Chajec, Ł.; Mazur, S. Can ceylon leadwort (Plumbago zeylanica L.) acclimate to lead toxicity?—Studies of photosynthetic apparatus efficiency. Int. J. Mol. Sci. 2020, 21, 1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klughammer, C.; Schreiber, U. Complementary PS II quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the Saturation Pulse method. PAM Appl. Notes 2008, 1, 27–35. [Google Scholar]
- Janeczko, A.; Oklešťková, J.; Siwek, A.; Dziurka, M.; Pociecha, E.; Kocurek, M.; Novák, O. Endogenous progesterone and its cellular binding sites in wheat exposed to drought stress. J. Steroid Biochem. Mol. Biol. 2013, 138, 384–394. [Google Scholar] [CrossRef]
- Hura, T.; Dziurka, M.; Hura, K.; Ostrowska, A.; Dziurka, K. Different allocation of carbohydrates and phenolics in dehydrated leaves of triticale. J. Plant Physiol. 2016, 202, 1–9. [Google Scholar] [CrossRef]
- Mikuła, A.; Tomaszewicz, W.; Dziurka, M.; Kaźmierczak, A.; Grzyb, M.; Sobczak, M.; Zdańkowski, P.; Rybczyński, J. The origin of the Cyathea delgadii Sternb. somatic embryos is determined by the developmental state of donor tissue and mutual balance of selected metabolites. Cells 2021, 10, 1388. [Google Scholar] [CrossRef]
- Tokarz, K.; Makowski, W.; Banasiuk, R.; Krolicka, A.; Piwowarczyk, B. Response of Dionaea muscipula J. Ellis to light stress in in vitro: Physiological study. Plant Cell. Tissue Organ Cult. 2018, 134, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.Y.; Wang, L.T.; Ma, J.H.; Ma, E.D.; Li, J.Y.; Gong, M. Effects of light quality on growth and development, photosynthetic characteristics and content of carbohydrates in tobacco (Nicotiana tabacum L.) plants. Photosynthetica 2017, 55, 467–477. [Google Scholar] [CrossRef]
- Tennessen, D.J.; Singsaas, E.L.; Sharkey, T.D. Light-emitting diodes as a light source for photosynthesis research. Photosynth. Res. 1994, 39, 85–92. [Google Scholar] [CrossRef]
- Fuentes, G.; Talavera, C.; Oropeza, C.; Desjardins, Y.; Santamaria, J.M. Exogenous sucrose can decrease in vitro photosynthesis but improve field survival and growth of coconut (Cocos nucifera L.) in vitro plantlets. Vitr. Cell. Dev. Biol.—Plant 2005, 41, 69–76. [Google Scholar] [CrossRef]
- Smith, A.M.; Stitt, M. Coordination of carbon supply and plant growth. Plant Cell Environ. 2007, 30, 1126–1149. [Google Scholar] [CrossRef]
- Ohto, M.A.; Onai, K.; Furukawa, Y.; Aoki, E.; Araki, T.; Nakamura, K. Effects of sugar on vegetative development and floral transition in Arabidopsis. Plant Physiol. 2001, 127, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, H.F.; Ma, Y.; Jin, Y.; Kong, G.; Lin, J.M. Microwave assisted extraction-solid phase extraction for high-efficient and rapid analysis of monosaccharides in plants. Talanta 2014, 129, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, L.D.R.; Paim, D.C.; Schwambach, J.; Fett-Neto, A.G. Carbohydrates as regulatory factors on the rooting of Eucalyptus saligna Smith and Eucalyptus globulus Labill. Plant Growth Regul. 2005, 45, 63–73. [Google Scholar] [CrossRef]
- Borisjuk, L.; Walenta, S.; Weber, H.; Mueller-Klieser, W.; Wobus, U. High-resolution histographical mapping of glucose concentrations in developing cotyledons of Vicia faba in relation to mitotic activity and storage processes: Glucose as a possible developmental trigger. Plant J. 1998, 15, 583–591. [Google Scholar] [CrossRef]
- Bach, A.; Kapczyńska, A.; Dziurka, K.; Dziurka, M. The importance of applied light quality on the process of shoot organogenesis and production of phenolics and carbohydrates in Lachenalia sp. cultures in vitro. S. Afr. J. Bot. 2018, 114, 14–19. [Google Scholar] [CrossRef]
- Cho, Y.H.; Yoo, S.D. Signaling role of fructose mediated by FINS1/FBP in Arabidopsis thaliana. PLoS Genet. 2011, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Perera, I.Y.; Hung, C.Y.; Brady, S.; Muday, G.K.; Boss, W.F. A universal role for inositol 1,4,5-trisphosphate-mediated signaling in plant gravitropism. Plant Physiol. 2006, 140, 746–760. [Google Scholar] [CrossRef] [Green Version]
- Venugopal, S.C.; Chanda, B.; Vaillancourt, L.; Kachroo, A.; Kachroo, P. The common metabolite glycerol-3-phosphate is a novel regulator of plant defense signaling. Plant Signal. Behav. 2009, 4, 746–749. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Xu, Z.; Tang, C. Effect of light-emitting diodes on growth and morphogenesis of upland cotton (Gossypium hirsutum L.) plantlets in vitro. Plant Cell. Tissue Organ Cult. 2010, 103, 155–163. [Google Scholar] [CrossRef]
- Wang, H.; Gu, M.; Cui, J.; Shi, K.; Zhou, Y.; Yu, J. Effects of light quality on CO2 assimilation, chlorophyll-fluorescence quenching, expression of Calvin cycle genes and carbohydrate accumulation in Cucumis sativus. J. Photochem. Photobiol. B Biol. 2009, 96, 30–37. [Google Scholar] [CrossRef]
- Heo, J.W.; Shin, K.S.; Kim, S.K.; Paek, K.Y. Light quality affects in vitro growth of grape ‘Teleki 5BB’. J. Plant Biol. 2006, 49, 276–280. [Google Scholar] [CrossRef]
- Qiu, D.; Vuong, T.; Valliyodan, B.; Shi, H.; Guo, B.; Shannon, J.G.; Nguyen, H.T. Identification and characterization of a stachyose synthase gene controlling reduced stachyose content in soybean. Theor. Appl. Genet. 2015, 128, 2167–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsayed, A.I.; Rafudeen, M.S.; Golldack, D. Physiological aspects of raffinose family oligosaccharides in plants: Protection against abiotic stress. Plant Biol. 2014, 16, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bentsink, L.; Alonso-Blanco, C.; Vreugdenhil, D.; Tesnier, K.; Groot, S.P.C.; Koornneef, M. Genetic analysis of seed-soluble oligosaccharides in relation to seed storability of Arabidopsis. Plant Physiol. 2000, 124, 1595–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karner, U.; Peterbauer, T.; Raboy, V.; Jones, D.A.; Hedley, C.L.; Richter, A. myo-Inositol and sucrose concentrations affect the accumulation of raffinose family oligosaccharides in seeds. J. Exp. Bot. 2004, 55, 1981–1987. [Google Scholar] [CrossRef]
- Van den Ende, W. Multifunctional fructans and raffinose family oligosaccharides. Front. Plant Sci. 2013, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Walter, A.; Spies, H.; Terjung, S.; Küsters, R.; Kirchgeßner, N.; Schurr, U. Spatio-temporal dynamics of expansion growth in roots: Automatic quantification of diurnal course and temperature response by digital image sequence processing. J. Exp. Bot. 2002, 53, 689–698. [Google Scholar] [CrossRef] [Green Version]
- Reddy, G.V.; Heisler, M.G.; Ehrhardt, D.W.; Meyerowitz, E.M. Real-time lineage analysis reveals oriented cell divisions associated with morphogenesis at the shoot apex of Arabidopsis thaliana. Development 2004, 131, 4225–4237. [Google Scholar] [CrossRef] [Green Version]
- Walter, A.; Schurr, U. Dynamics of leaf and root growth: Endogenous control versus environmental impact. Ann. Bot. 2005, 95, 891–900. [Google Scholar] [CrossRef]
- Nozue, K.; Maloof, J.N. Diurnal regulation of plant growth. Plant Cell Environ. 2006, 29, 396–408. [Google Scholar] [CrossRef]
- Gordon, A.J.; Ryle, G.J.A.; Powell, C.E.; Mitchell, D. Export, mobilization, and respiration of assimilates in uniculm barley during light and darkness. J. Exp. Bot. 1980, 31, 461–473. [Google Scholar] [CrossRef]
- Cross, J.M.; von Korff, M.; Altmann, T.; Bartzenko, L.; Sulpice, R.; Gibon, Y.; Palacios, N.; Stitt, M. Natural variation in carbon-nitrogen interactions: Changes of metabolite levels and enzyme activities across 24 Arabidopsis thaliana accessions. Plant Physiol. 2006, 142, 1574–1588. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Tang, C.; Xu, Z. The effects of different light qualities on rapeseed (Brassica napus L.) plantlet growth and morphogenesis in vitro. Sci. Hortic. 2013, 150, 117–124. [Google Scholar] [CrossRef]
- Altenbach, D.; Nüesch, E.; Ritsema, T.; Boller, T.; Wiemken, A. Mutational analysis of the active center of plant fructosyltransferases: Festuca 1-SST and barley 6-SFT. FEBS Lett. 2005, 579, 4647–4653. [Google Scholar] [CrossRef] [Green Version]
- Chia, T.; Thorneycroft, D.; Chapple, A.; Messerli, G.; Chen, J.; Zeeman, S.C.; Smith, S.M.; Smith, A.M. A cytosolic glucosyltransferase is required for conversion of starch to sucrose in Arabidopsis leaves at night. Plant J. 2004, 37, 853–863. [Google Scholar] [CrossRef]
- Lu, Y.; Steichen, J.M.; Yao, J.; Sharkey, T.D. The role of cytosolic α-glucan phosphorylase in maltose metabolism and the comparison of amylomaltase in Arabidopsis and Escherichia coli. Plant Physiol. 2006, 142, 878–889. [Google Scholar] [CrossRef] [Green Version]
- Batty, N.; Dunwell, J. Effect of maltose on the response of potato anthers in culture. Plant Cell. Tissue Organ Cult. 1989, 18, 221–226. [Google Scholar] [CrossRef]
- Raquin, C. Utilization of different sugars as carbon source for in vitro anther culture of Petunia. Z. Pflanzenphysiol. 1983, 111, 453–457. [Google Scholar] [CrossRef]
- Strickland, S.G.; Nichol, J.W.; McCall, C.M.; Stuart, D.A. Effect of carbohydrate source on alfalfa somatic embryogenesis. Plant Sci. 1987, 48, 113–121. [Google Scholar] [CrossRef]
- Ming, N.J.; Mostafiz, S.B.; Johon, N.S.; Zulkifli, N.S.A.; Wagiran, A. Combination of plant growth regulators, maltose, and partial desiccation treatment enhance somatic embryogenesis in selected malaysian rice cultivar. Plants 2019, 8, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuraida, A.R.; Naziah, B.; Zamri, Z.; Sreeramanan, S. Efficient plant regeneration of Malaysian indica rice MR 219 and 232 via somatic embryogenesis system. Acta Physiol. Plant 2011, 33, 1913–1921. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Quality | Glucose | Fructose |
|---|---|---|
| B 1 | 39.83 ± 7.29 a 2 | 20.20 ± 3.65 a |
| RB | 56.88 ± 11.84 ab | 22.39 ± 2.99 a |
| R | 56.84 ± 5.46 ab | 24.01 ± 6.30 a |
| Fl | 74.95 ± 21.78 b | 32.93 ± 15.89 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cioć, M.; Tokarz, K.; Dziurka, M.; Pawłowska, B. Energy-Saving LED Light Affects the Efficiency of the Photosynthetic Apparatus and Carbohydrate Content in Gerbera jamesonii Bolus ex Hook. f. Axillary Shoots Multiplied In Vitro. Biology 2021, 10, 1035. https://doi.org/10.3390/biology10101035
Cioć M, Tokarz K, Dziurka M, Pawłowska B. Energy-Saving LED Light Affects the Efficiency of the Photosynthetic Apparatus and Carbohydrate Content in Gerbera jamesonii Bolus ex Hook. f. Axillary Shoots Multiplied In Vitro. Biology. 2021; 10(10):1035. https://doi.org/10.3390/biology10101035
Chicago/Turabian StyleCioć, Monika, Krzysztof Tokarz, Michał Dziurka, and Bożena Pawłowska. 2021. "Energy-Saving LED Light Affects the Efficiency of the Photosynthetic Apparatus and Carbohydrate Content in Gerbera jamesonii Bolus ex Hook. f. Axillary Shoots Multiplied In Vitro" Biology 10, no. 10: 1035. https://doi.org/10.3390/biology10101035