Effects on Intestinal Mucosal Morphology, Productive Parameters and Microbiota Composition after Supplementation with Fermented Defatted Alperujo (FDA) in Laying Hens

 , ,
, ,
Abstract
:1. Introduction
2. Results
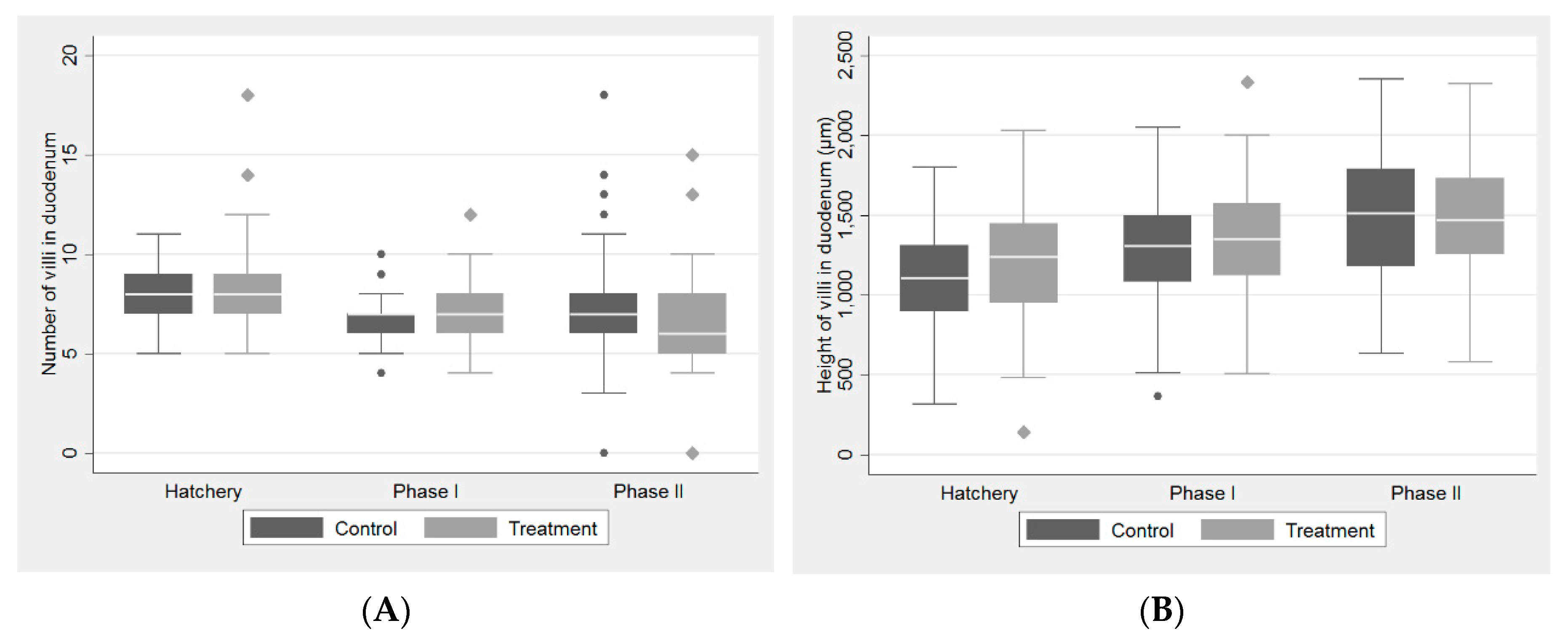
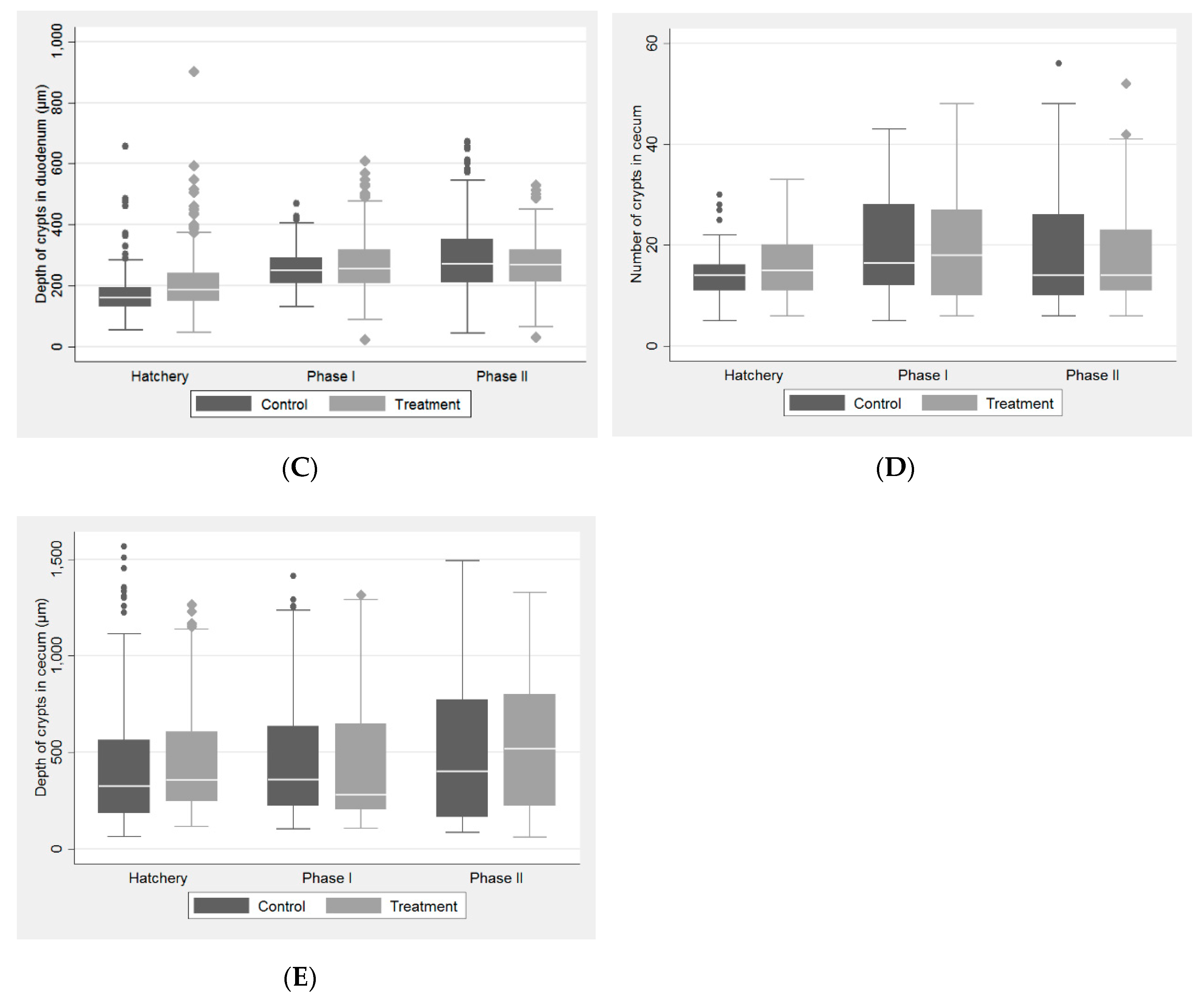
2.1. Histological Study
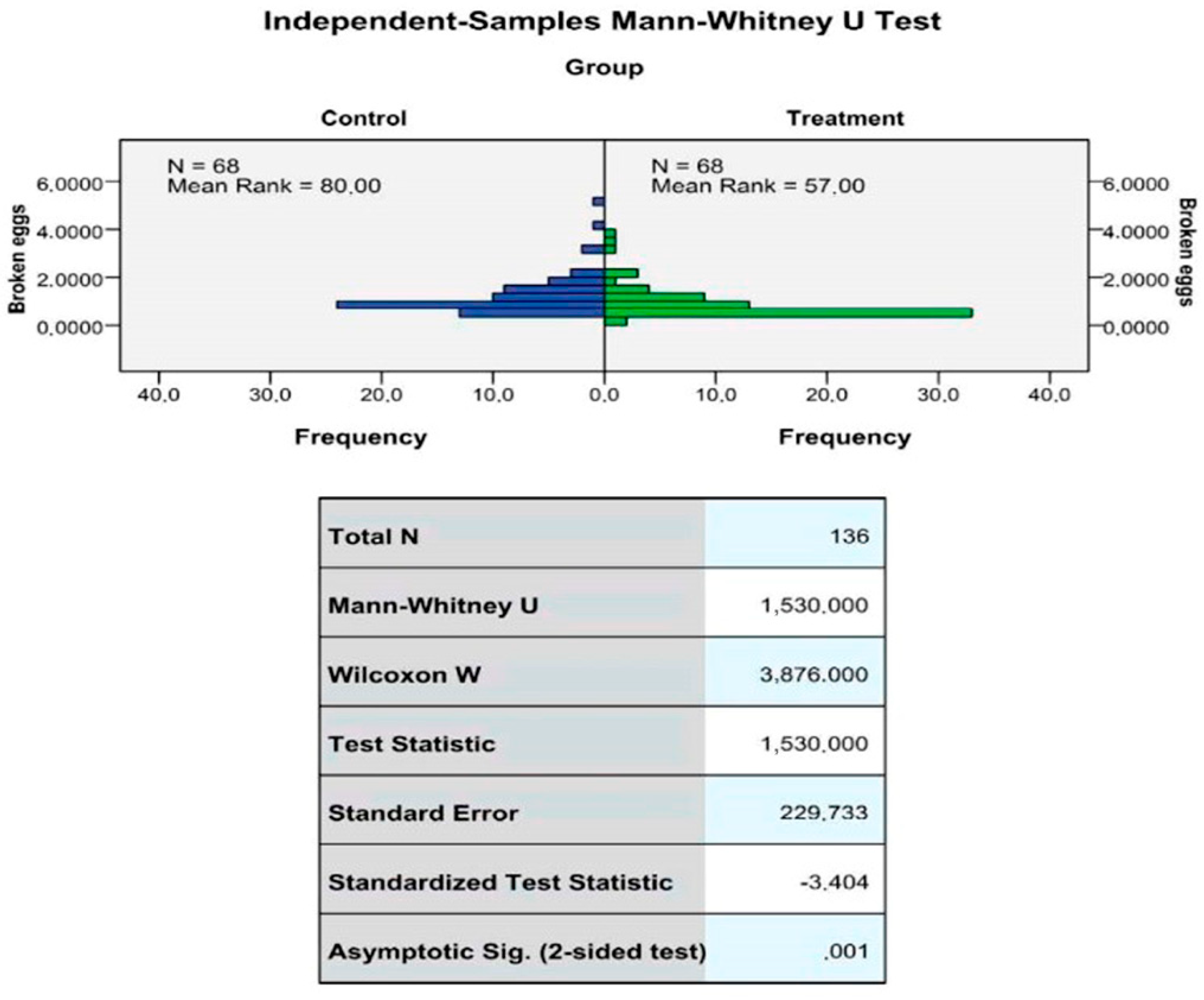
2.2. Production Performance
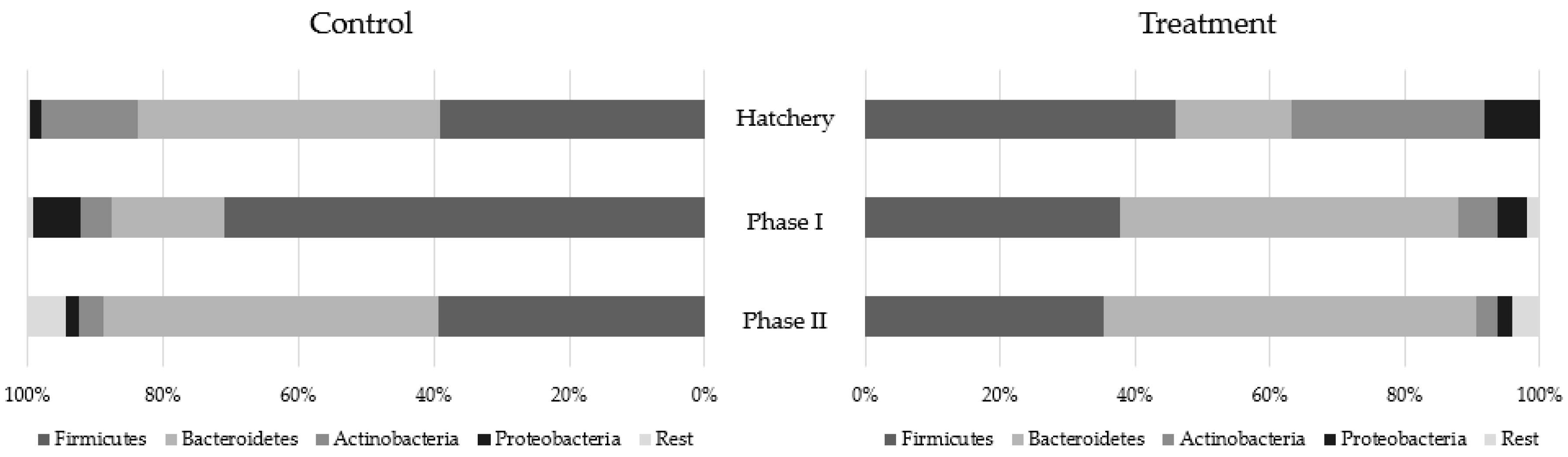
2.3. Metagenomics
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Animals and Rearing Conditions
4.3. Experimental Diets
4.4. Production Performance
4.5. Samplings and Necropsy
4.6. Histology
4.7. Metagenomics
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- International Olive Council. Available online: http://www.internationaloliveoil.org/ (accessed on 9 October 2019).
- European Commission. Available online: https://ec.europa.eu/ (accessed on 9 October 2019).
- Ministerio de Agricultura, Pesca y Alimentación. Available online: https://www.mapa.gob.es/en/ (accessed on 9 October 2019).
- Banias, G.; Achillas, C.; Vlachokostas, C.; Moussiopoulos, N. Environmental impacts in the life cycle of olive oil: A literature review. J. Sci. Food Agric. 2017, 97, 1686–1697. [Google Scholar] [CrossRef] [PubMed]
- Kapellakis, I.E.; Tsagarakis, K.P.; Crowther, J.C. Olive oil history, production and by-product management. Rev. Environ. Sci. Biotechnol. 2008, 7, 1–26. [Google Scholar] [CrossRef]
- Alburquerque, J.A.; Gonzálvez, J.; García, D.; Cegarra, J. Agrochemical characterisation of “alperujo”, a solid by-product of the two-phase centrifugation method for olive oil extraction. Bioresour. Technol. 2004, 91, 195–200. [Google Scholar] [CrossRef]
- Duborija-Kovacevic, N.; Shavrina, K. New findings on the pharmacodynamic actions of olive oil: Our contribution to better evidence about its remedial properties. Prog. Nutr. 2018, 20, 30–38. [Google Scholar]
- Leouifoudi, I.; Harnafi, H.; Zyad, A. Olive mill waste extracts: Polyphenols content, antioxidant, and antimicrobial activities. Adv. Pharmacol. Sci. 2015, 2015, 714138. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Alli, I.; Ereifej, K.; Alhamad, M.; Al-Tawaha, A.R.; Rababah, T. Optimisation, characterisation and quantification of phenolic compounds in olive cake. Food Chem. 2010, 123, 117–122. [Google Scholar] [CrossRef]
- Aissa, I.; Kharrat, N.; Aloui, F.; Sellami, M.; Bouaziz, M.; Gargouri, Y. Valorization of antioxidants extracted from olive oil mill wastewater. Biotechnol. Appl. Biochem. 2017, 64, 579–589. [Google Scholar] [CrossRef]
- Sabino, M.; Cappelli, K.; Capomaccio, S.; Pascucci, L.; Biasato, I.; Verini-Supplizi, A.; Valiani, A.; Trabalza-Marinucci, M. Dietary supplementation with olive mill wastewaters induces modifications on chicken jejunum epithelial cell transcriptome and modulates jejunum morphology. BMC Genom. 2018, 19, 576. [Google Scholar] [CrossRef]
- Roila, R.; Ranucci, D.; Valiani, A.; Galarini, R.; Servili, M.; Branciari, R. Antimicrobial and anti-biofilm activity of olive oil by-products against Campylobacter spp. isolated from chicken meat. Acta Sci. Pol. Technol. Aliment. 2019, 18, 43–52. [Google Scholar]
- Berbel, J.; Posadillo, A. Review and analysis of alternatives for the valorisation of agro-industrial olive oil by-products. Sustainability 2018, 10, 237. [Google Scholar] [CrossRef]
- Al-Harthi, M.A. The efficacy of using olive cake as a by-product in broiler feeding with and without yeast. Ital. J. Anim. Sci. 2016, 15, 512–520. [Google Scholar] [CrossRef]
- Herrero-Encinas, J.; Blanch, M.; Pastor, J.J.; Mereu, A.; Ipharraguerre, I.R.; Menoyo, D. Effects of a bioactive olive pomace extract from Olea europaea on growth performance, gut function, and intestinal microbiota in broiler chickens. Poult. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Shimao, R.; Muroi, H.; Furukawa, K.; Toyomizu, M.; Kikusato, M. Effects of low-dose oleuropein diet supplementation on the oxidative status of skeletal muscles and plasma hormonal concentration of growing broiler chickens. Br. Poult. Sci. 2019. [Google Scholar] [CrossRef]
- Gerasopoulos, K.; Stagos, D.; Kokkas, S.; Petrotos, K.; Kantas, D.; Goulas, P.; Kouretas, D. Feed supplemented with byproducts from olive oil mill wastewater processing increases antioxidant capacity in broiler chickens. Food Chem. Toxicol. 2015, 82, 42–49. [Google Scholar] [CrossRef]
- Debbou-Iouknane, N.; Nerín, C.; Amrane, M.; Ghemghar, M.; Madani, K.; Ayad, A. In vitro anticoccidial activity of olive pulp (Olea europaea L. var. chemlal) extract against Eimeria oocysts in broiler chickens. Acta Parasitol. 2019. [Google Scholar] [CrossRef]
- Iannaccone, M.; Ianni, A.; Ramazzotti, S.; Grotta, L.; Marone, E.; Cichelli, A.; Martino, G. Whole blood transcriptome analysis reveals positive effects of dried olive pomace-supplemented diet on inflammation and cholesterol in laying hens. Animals 2019, 9, 427. [Google Scholar] [CrossRef]
- Afsari, M.; Mohebbifar, A.; Torki, M. Effects of dietary inclusion of olive pulp supplemented with probiotics on productive performance, egg quality and blood parameters of laying hens. Annu. Res. Rev. Biol. 2014, 4, 198–211. [Google Scholar] [CrossRef]
- Zarei, M.; Ehsani, M.; Torki, M. Productive performance of laying hens fed wheat-based diets included olive pulp with or without a commercial enzyme product. Afr. J. Biotechnol. 2011, 10, 4303–4312. [Google Scholar]
- Zangeneh, S.; Torki, M. Effects of b-mannanase supplementing of olive pulp-included diet on performance of laying hens, egg quality characteristics, humoral and cellular immune response and blood parameters. Glob. Vet. 2011, 7, 391–398. [Google Scholar]
- Ghasemi, R.; Torki, M.; Ghasemi, H.A.; Zarei, M. Single or combined effects of date pits and olive pulps on productive traits, egg quality, serum lipids and leucocytes profiles of laying hens. J. Appl. Anim. Res. 2013, 42, 103–109. [Google Scholar] [CrossRef]
- Al-Harthi, M.A.; Attia, Y.A. Effect of citric acid on the utilization of olive cake diets for laying hens. Ital. J. Anim. Sci. 2015, 14, 3966. [Google Scholar] [CrossRef]
- Yegani, M.; Korver, D.R. Factors affecting intestinal health in poultry. Poult. Sci. 2008, 87, 2052–2063. [Google Scholar] [CrossRef]
- Graziani, C.; Talocco, C.; de Sire, R.; Petito, V.; Lopetuso, L.R.; Gervasoni, J.; Persichilli, S.; Franceschi, F.; Ojetti, V.; Gasbarrini, A.; et al. Intestinal permeability in physiological and pathological conditions: Major determinants and assessment modalities. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 795–810. [Google Scholar] [PubMed]
- Rubio, L.A. Possibilities of early life programming in broiler chickens via intestinal microbiota modulation. Poult. Sci. 2019, 98, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Al-Harthi, M.A. The effect of olive cake, with or without enzymes supplementation, on growth performance, carcass characteristics, lymphoid organs and lipid metabolism of broiler chickens. Braz. J. Poult. Sci. 2017, 83–90. [Google Scholar] [CrossRef]
- Ding, X.M.; Li, D.D.; Bai, S.P.; Wang, J.P.; Zeng, Q.F.; Su, Z.W.; Xuan, Y.; Zhang, K.Y. Effect of dietary xylooligosaccharides on intestinal characteristics, gut microbiota, cecal short-chain fatty acids, and plasma immune parameters of laying hens. Poult. Sci. 2018, 97, 874–881. [Google Scholar] [CrossRef]
- Rattanawut, J.; Pimpa, O.; Yamauchi, K.E. Effects of dietary bamboo vinegar supplementation on performance, eggshell quality, ileal microflora composition, and intestinal villus morphology of laying hens in the late phase of production. Anim. Sci. J. 2018, 89, 1572–1580. [Google Scholar] [CrossRef]
- Wang, H.; Liang, S.; Li, X.; Yang, X.; Long, F.; Yang, X. Effects of encapsulated essential oils and organic acids on laying performance, egg quality, intestinal morphology, barrier function, and microflora count of hens during the early laying period. Poult. Sci. 2019. [Google Scholar] [CrossRef]
- Li, L.; Li, H.; Zhou, W.; Feng, J.; Zou, X. Effects of zinc methionine supplementation on laying performance, zinc status, intestinal morphology, and expressions of zinc transporters’ mRNA in laying hens. J. Sci. Food Agric. 2019, 99, 6582–6588. [Google Scholar] [CrossRef]
- Lokaewmanee, K.; Yamauchi, K.; Okuda, N. Effects of dietary red pepper on egg yolk colour and histological intestinal morphology in laying hens. J. Anim. Physiol. Anim. Nutr. 2013, 97, 986–995. [Google Scholar] [CrossRef]
- Zhu, L.P.; Wang, J.P.; Ding, X.M.; Bai, S.P.; Zeng, Q.F.; Su, Z.W.; Xuan, Y.; Applegate, T.J.; Zhang, K.Y. The effects of varieties and levels of rapeseed expeller cake on egg production performance, egg quality, nutrient digestibility, and duodenum morphology in laying hens. Poult. Sci. 2019, 98, 4942–4953. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.H.; Wang, J.P.; Zhang, K.Y.; Ding, X.M.; Bai, S.P.; Zeng, Q.F.; Xuan, Y.; Su, Z.W. Effect of vanadium and tea polyphenols on intestinal morphology, microflora and short-chain fatty acid profile of laying hens. Biol. Trace Elem. Res. 2016, 174, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Nain, S.; Renema, R.A.; Zuidhof, M.J.; Korver, D.R. Effect of metabolic efficiency and intestinal morphology on variability in n-3 polyunsaturated fatty acid enrichment of eggs. Poult. Sci. 2012, 91, 888–898. [Google Scholar] [CrossRef]
- Gong, H.Z.; Wu, M.; Lang, W.Y.; Yang, M.; Wang, J.H.; Wang, Y.Q.; Zhang, Y.; Zheng, X. Effects of laying breeder hens dietary β-carotene, curcumin, allicin, and sodium butyrate supplementation on the growth performance, immunity, and jejunum morphology of their offspring chicks. Poult. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, O.J.; Abdul-Aziz, T. Alimentary System. In Avian Histopathology, 4th ed.; Abdul-Aziz, T., Ed.; The American Association of Avian Pathologists: Jacksonville, FL, USA, 2016; pp. 271–280. [Google Scholar]
- Lilburn, M.S.; Loeffler, S. Early intestinal growth and development in poultry. Poult. Sci. 2015, 94, 1569–1576. [Google Scholar] [CrossRef]
- Pourabedin, M.; Zhao, X. Prebiotics and gut microbiota in chickens. FEMS Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef]
- Beck, M.M.; Hansen, K.K. Role of estrogen in avian osteoporosis. Poult. Sci. 2004, 83, 200–206. [Google Scholar] [CrossRef]
- Shalaei, M.; Hosseini, S.M.; Zergani, E. Effects of different supplements on eggshell quality, some characteristics of gastrointestinal tract and performance of laying hens. Vet. Res. Forum 2014, 5, 277–286. [Google Scholar]
- Boka, J.; Mahdavi, A.H.; Samie, A.H.; Jahanian, R. Effect of different levels of black cumin (Nigella sativa L.) on performance, intestinal Escherichia coli colonization and jejunal morphology in laying hens. J. Anim. Physiol. Anim. Nutr. 2014, 98, 373–383. [Google Scholar] [CrossRef]
- Videnska, P.; Sedlar, K.; Lukac, M.; Faldynova, M.; Gerzova, L.; Cejkova, D.; Sisak, F.; Rychlik, I. Succession and replacement of bacterial populations in the caecum of egg laying hens over their whole life. PLoS ONE 2014, 9, e115142. [Google Scholar] [CrossRef]
- Qi, Z.; Shi, S.; Tu, J.; Li, S. Comparative metagenomic sequencing analysis of cecum microbiotal diversity and function in broilers and layers. 3 Biotech 2019, 9, 316. [Google Scholar] [CrossRef] [PubMed]
- Videnska, P.; Rahman, M.M.; Faldynova, M.; Babak, V.; Matulova, M.E.; Prukner-Radovcic, E.; Krizek, I.; Smole-Mozina, S.; Kovac, J.; Szmolka, A.; et al. Characterization of egg laying hen and broiler fecal microbiota in poultry farms in Croatia, Czech Republic, Hungary and Slovenia. PLoS ONE 2014, 9, e110076. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Wang, Q.; Liu, S.; Sun, R.; Zhou, Y.; Li, Y. Age-related variations in intestinal microflora of free-range and caged. Front. Microbiol. 2017, 8, 1310. [Google Scholar] [CrossRef] [PubMed]
- Han, G.P.; Lee, K.C.; Kang, H.K.; Oh, H.N.; Sul, W.J.; Kil, D.Y. Analysis of excreta bacterial community after forced-molting in aged laying. Asian-Australas. J. Anim. Sci. 2019, 32, 1715–1724. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.W.; Jia, H.J.; Zhang, H.J.; Wang, J.; Lv, H.Y.; Wu, S.G.; Qi, G.H. Supplemental plant extracts from Flos lonicerae in combination with Baikal skullcap attenuate intestinal disruption and modulate gut microbiota in laying hens challenged by Salmonella pullorum. Front. Microbiol. 2019, 10, 1681. [Google Scholar] [CrossRef]
- Song, D.; Wang, Y.W.; Lu, Z.X.; Wang, W.W.; Miao, H.J.; Zhou, H.; Wang, L.; Li, A.K. Effects of dietary supplementation of microencapsulated Enterococcus fecalis and the extract of Camellia oleifera seed on laying performance, egg quality, serum biochemical parameters, and cecal microflora diversity in laying hens. Poult. Sci. 2019, 98, 2880–2887. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Histomorphometric Parameters | Control Group | FDA-Supplemented Group | p-Value 1 | ||
|---|---|---|---|---|---|
| Mean | [IQR] | Mean | [IQR] | ||
| Hatchery | |||||
| Duodenum | |||||
| Number of villi | 8.00 | [2.00] | 8.00 | [2.00] | 0.311 |
| Height of villi (µm) | 1109.95 | [410.00] | 1237.85 | [492.10] | <0.001 |
| Depth of crypts (µm) | 161.05 | [62.53] | 188.10 | [90.55] | <0.001 |
| Cecum | |||||
| Number of crypts | 14.00 | [5.00] | 15.00 | [9.00] | 0.028 |
| Depth of crypts (µm) | 325.65 | [374.20] | 358.15 | [356.60] | <0.001 |
| Production phase I | |||||
| Phase I | |||||
| Duodenum | |||||
| Number of villi | 7.00 | [1.00] | 7.00 | [2.00] | <0.001 |
| Height of villi (µm) | 1309.27 | [411.03] | 1353.32 | [449.94] | 0.157 |
| Depth of crypts (µm) | 249.66 | [81.83] | 255.15 | [107.59] | 0.135 |
| Cecum | |||||
| Number of crypts | 16.50 | [16.00] | 18.00 | [17.00] | 0.821 |
| Depth of crypts (µm) | 359.12 | [411.14] | 280.74 | [440.91] | 0.150 |
| Production phase II | |||||
| Phase II | |||||
| Duodenum | |||||
| Number of villi | 7.00 | [2.00] | 6.00 | [3.00] | 0.230 |
| Height of villi (µm) | 1511.99 | [605.57] | 1471.92 | [476.69] | 0.537 |
| Depth of crypts (µm) | 272.65 | [138.47] | 268.46 | [102.79] | 0.160 |
| Cecum | |||||
| Number of crypts | 14.00 | [16.00] | 14.00 | [12.00] | 0.883 |
| Depth of crypts (µm) | 402.68 | [606.61] | 518.50 | [576.58] | <0.001 |
| Productive Parameter | Control Group | Treatment Group | p-Value 1 |
|---|---|---|---|
| Mortality (%) | 0.15 | 0.16 | 0.299 |
| Laying (%) | 78.00 | 79.00 | 0.970 |
| Feed/hens (g) | 115.37 | 113.37 | 0.124 |
| Egg weight (g) | 62.58 | 62.59 | 0.720 |
| Egg mass (g/d) | 48.04 | 49.28 | 0.730 |
| Extra-large eggs (%) | 5.20 | 5.40 | 0.989 |
| Large eggs (%) | 44.25 | 43.94 | 0.467 |
| Medium eggs (%) | 39.04 | 40.39 | 0.917 |
| Small eggs (%) | 5.8 | 5.4 | 0.084 |
| Dirty eggs (%) | 2.3 | 2.1 | 0.424 |
| Broken eggs (%) | 3.47 | 2.83 | 0.001 |
| Total eggs (number) | 21,526,722 | 21,892,058 | 0.808 |
| Total eggs (Kg) | 1,342,571.52 | 1,363,916.15 | 0.931 |
| CI (Conversion Index) | 0.37 | 0.35 | - |
| Determination | Results |
|---|---|
| Moisture 103° (%w.w.) | 12.2 |
| Crude protein (Kjeldahl) (%w.w.) | 6.4 |
| Brute fat (%w.w.) | 3.0 |
| Ash content (%w.w.) | 7.7 |
| Lignin (%w.w.) | 23.3 |
| Acid detergent fiber (%w.w.) | 39.2 |
| Neutral detergent fiber (%w.w.) | 49.3 |
| Tannins (%w.w.) | 0.06 |
| Oleic acidity index (%w.w.) | 46.1 |
| Peroxide value (%w.w.) | 7.9 |
| Total polyphenols (meq/kg) | 0.89 |
| Crude fiber (%w.w.) | 27.7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rebollada-Merino, A.; Bárcena, C.; Ugarte-Ruiz, M.; Porras, N.; Mayoral-Alegre, F.J.; Tomé-Sánchez, I.; Domínguez, L.; Rodríguez-Bertos, A. Effects on Intestinal Mucosal Morphology, Productive Parameters and Microbiota Composition after Supplementation with Fermented Defatted Alperujo (FDA) in Laying Hens. Antibiotics 2019, 8, 215. https://doi.org/10.3390/antibiotics8040215
Rebollada-Merino A, Bárcena C, Ugarte-Ruiz M, Porras N, Mayoral-Alegre FJ, Tomé-Sánchez I, Domínguez L, Rodríguez-Bertos A. Effects on Intestinal Mucosal Morphology, Productive Parameters and Microbiota Composition after Supplementation with Fermented Defatted Alperujo (FDA) in Laying Hens. Antibiotics. 2019; 8(4):215. https://doi.org/10.3390/antibiotics8040215
Chicago/Turabian StyleRebollada-Merino, Agustín, Carmen Bárcena, María Ugarte-Ruiz, Néstor Porras, Francisco J. Mayoral-Alegre, Irene Tomé-Sánchez, Lucas Domínguez, and Antonio Rodríguez-Bertos. 2019. "Effects on Intestinal Mucosal Morphology, Productive Parameters and Microbiota Composition after Supplementation with Fermented Defatted Alperujo (FDA) in Laying Hens" Antibiotics 8, no. 4: 215. https://doi.org/10.3390/antibiotics8040215