13-(2-Methylbenzyl) Berberine Is a More Potent Inhibitor of MexXY-Dependent Aminoglycoside Resistance than Berberine

Abstract
:1. Introduction
2. Results
2.1. Antibacterial Activity of Berberine Derivatives toward P. aeruginosa
2.2. Inhibition of Drug Resistance in P. aeruginosa Using Combined Berberine Derivatives
2.3. Interaction between 13-o-MBB and Aminoglycoside Drugs
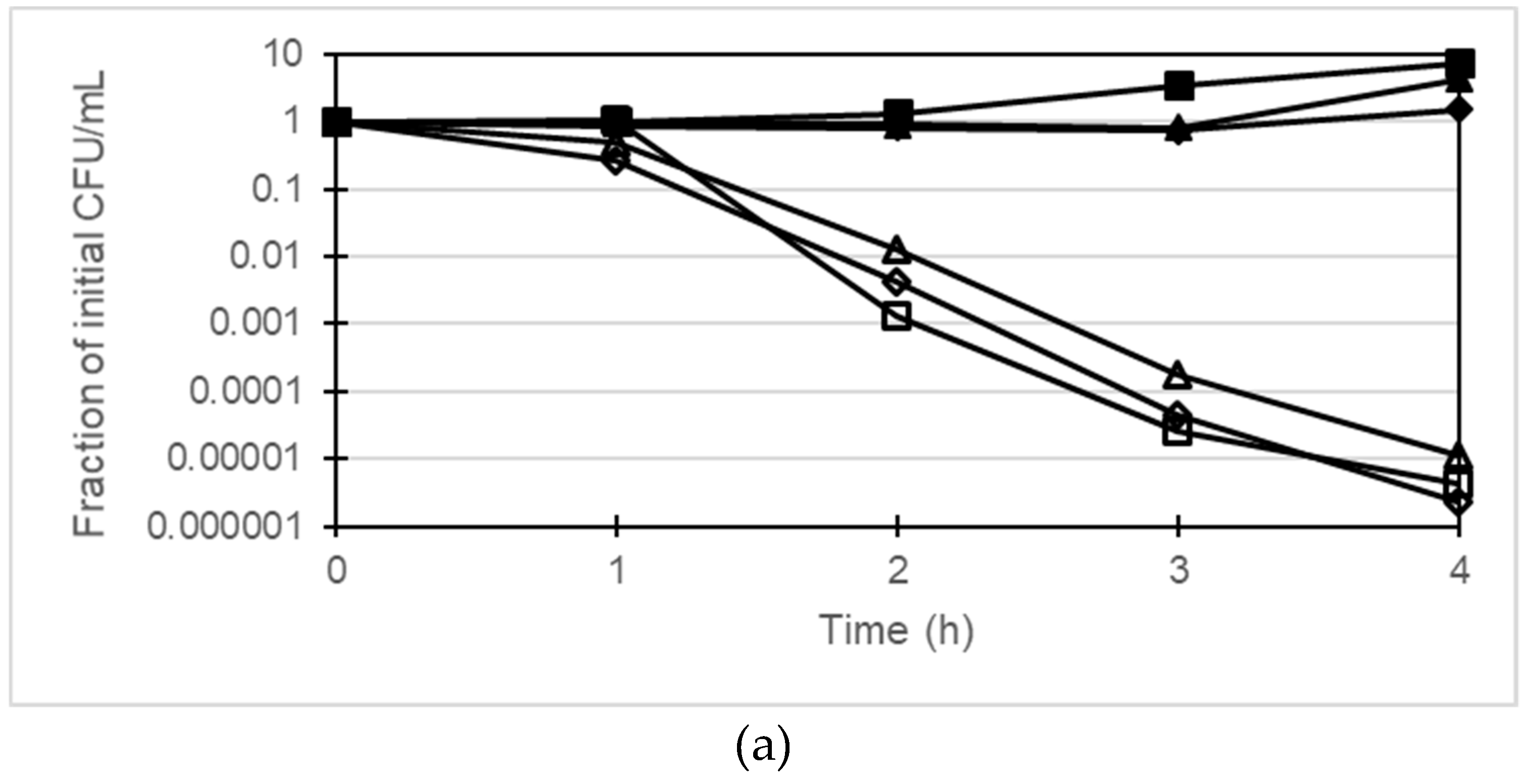
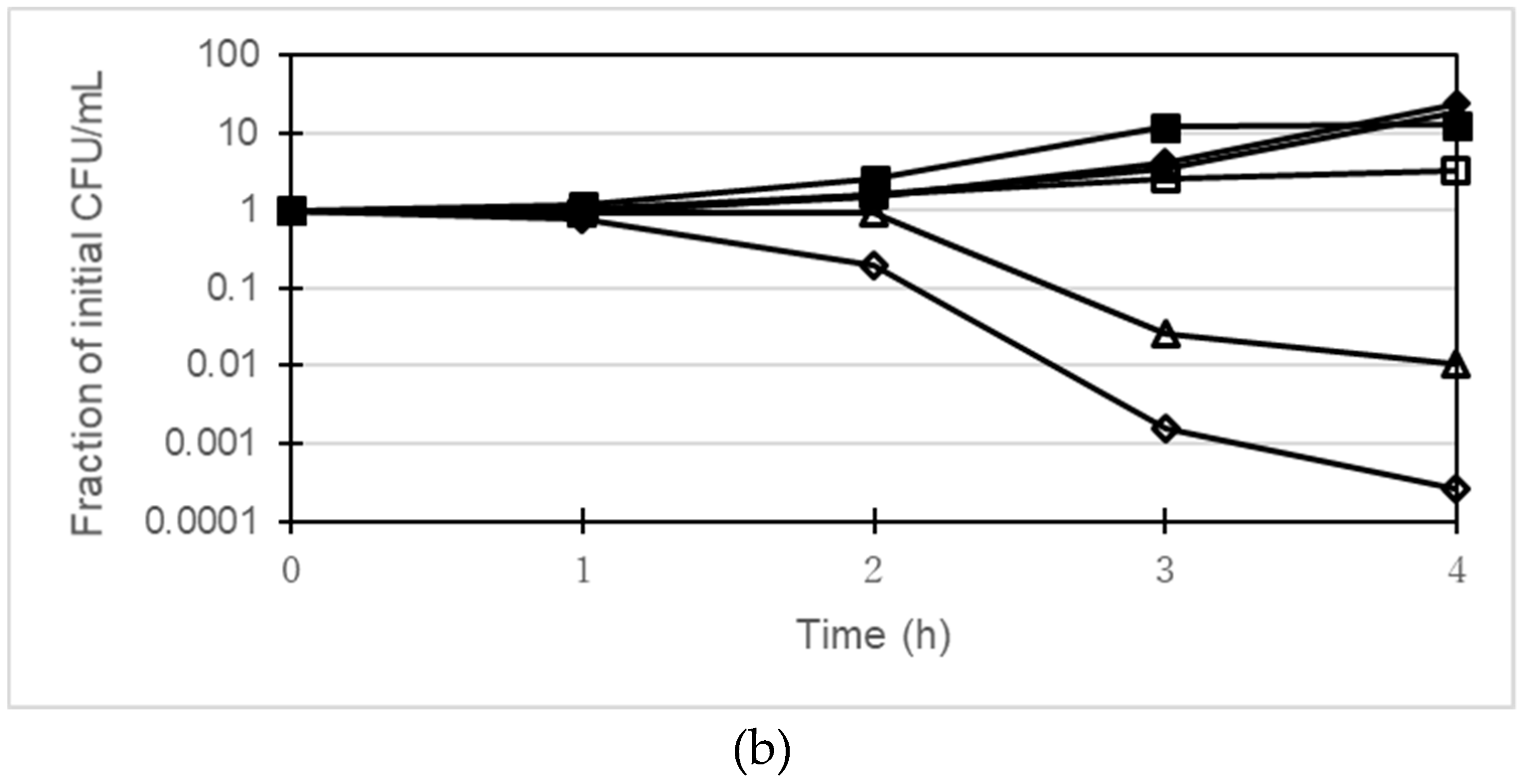
2.4. Time-Killing Assay
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Antibiotic Susceptibility Assay
4.3. Time-Killing Assay
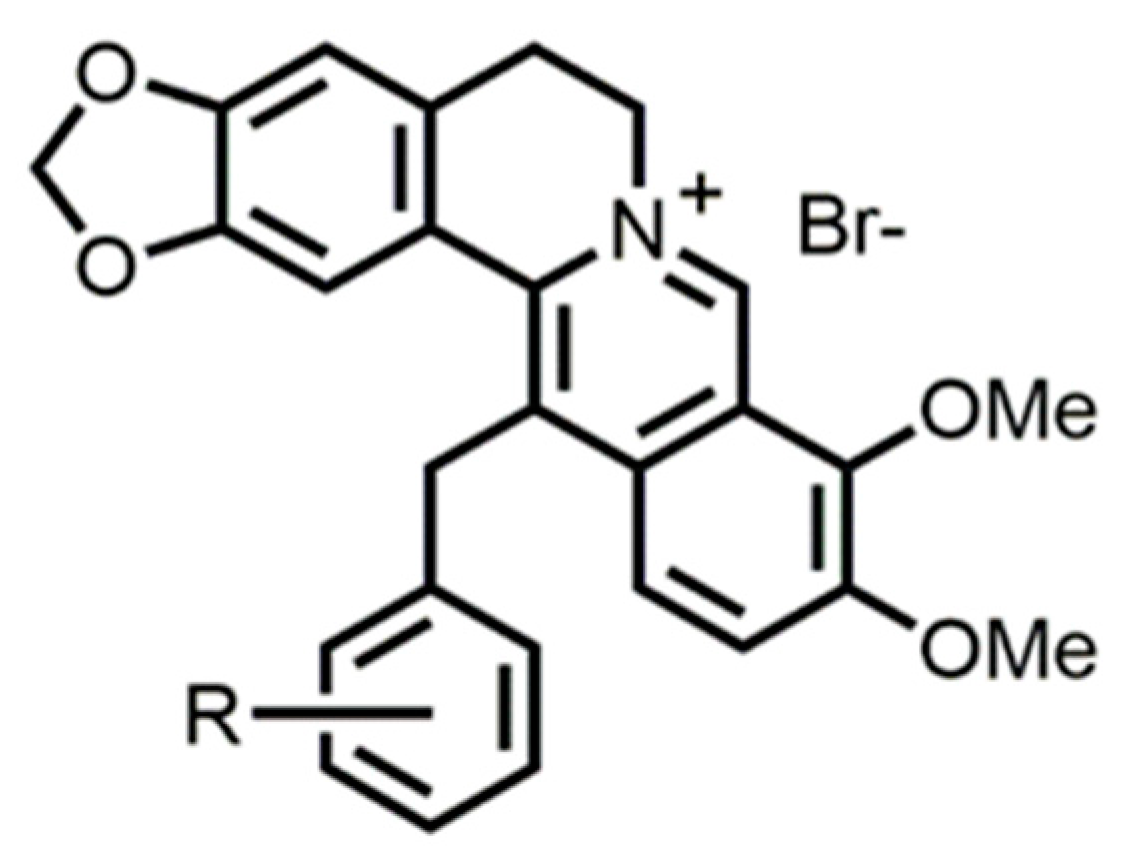
4.4. Synthesis
4.4.1. General Synthesis Information
4.4.2. 13-Benzylberberine Derivatives; General Procedure
4.4.3. Characterisation Data of 13-(2,6-Dichlorobenzyl)berberine Bromide (11)
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nair, C.G.; Chao, C.; Ryall, B.; Williams, H.D. Sub-lethal concentrations of antibiotics increase mutation frequency in the cystic fibrosis pathogen Pseudomonas aeruginosa. Lett. Appl. Microbiol. 2013, 56, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Dantas, R.C.; Ferreira, M.L.; Gontijo-Filho, P.P.; Ribas, R.M. Pseudomonas aeruginosa bacteraemia: Independent risk factors for mortality and impact of resistance on outcome. J. Med. Microbiol. 2014, 63, 1679–1687. [Google Scholar] [CrossRef] [PubMed]
- Poole, K. Pseudomonas aeruginosa: Resistance to the max. Front. Microbiol. 2011, 5, 65. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, K.; Yonehara, R.; Ishizaka-Ikeda, E.; Miyazaki, N.; Maeda, S.; Iwasaki, K.; Nakagawa, A.; Yamashita, E. Structures of the wild-type MexAB-OprM tripartite pump reveal its complex formation and drug efflux mechanism. Nat. Commun. 2019, 10, 1520. [Google Scholar] [CrossRef] [PubMed]
- Alcalde-Rico, M.; Olivares-Pacheco, J.; Alvarez-Ortega, C.; Cámara, M.; Martínez, J.L. Role of the Multidrug Resistance Efflux Pump MexCD-OprJ in the Pseudomonas aeruginosa Quorum Sensing Response. Front. Microbiol. 2018, 9, 2752. [Google Scholar] [CrossRef]
- Juarez, P.; Broutin, I.; Bordi, C.; Plésiat, P.; Llanes, C. Constitutive Activation of MexT by Amino Acid Substitutions Results in MexEF-OprN Overproduction in Clinical Isolates of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2018, 62, e02445-17. [Google Scholar] [CrossRef]
- Morita, Y.; Tomida, J.; Kawamura, Y. MexXY multidrug efflux system of Pseudomonas aeruginosa. Front. Microbiol. 2012, 28, 408. [Google Scholar] [CrossRef]
- Morita, Y.; Tomida, J.; Kawamura, Y. Primary mechanisms mediating aminoglycoside resistance in the multidrug-resistant Pseudomonas aeruginosa clinical isolate PA7. Microbiology 2012, 158, 1071–1083. [Google Scholar] [CrossRef]
- Nikaido, H.; Takatsuka, Y. Mechanisms of RND multidrug efflux pumps. Biochim. Biophys. Acta 2009, 1794, 769–781. [Google Scholar] [CrossRef] [Green Version]
- Blair, J.M.; Piddock, L.J. Structure, function and inhibition of RND efflux pumps in Gram-negative bacteria: An update. Curr. Opin. Microbiol. 2009, 12, 512–519. [Google Scholar] [CrossRef]
- Vogne, C.; Aires, J.R.; Bailly, C.; Hocquet, D.; Plésiat, P. Role of the multidrug efflux system MexXY in the emergence of moderate resistance to aminoglycosides among Pseudomonas aeruginosa isolates from patients with cystic fibrosis. Antimicrob. Agents Chemother. 2004, 48, 1676–1680. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Jalal, S.; Wretlind, B. Expression of the MexXY efflux pump in amikacin-resistant isolates of Pseudomonas aeruginosa. Clin. Microbiol. Infect. 2004, 10, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Novoa, J.M.; Quiros, Y.; Vicente, L.; Morales, A.I.; Lopez-Hernandez, F.J. New insights into the mechanism of aminoglycoside nephrotoxicity: An integrative point of view. Kidney Int. 2011, 79, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Jürgen, A.B.; Winfried, V.K. Antimicrobail Drug Efflux Pump Inhibitor. In Efflux-Mediated Antimicrobial Resistance in Bacteria, 2nd ed.; Xian-Zhi, L., Christpher, A.E., Helen, I.Z., Eds.; Springer International Publishing: Basel, Switzerland, 2016; pp. 755–795. [Google Scholar]
- Morita, Y.; Nakashima, K.; Nishino, K.; Kotani, K.; Tomida, J.; Inoue, M.; Kawamura, Y. Berberine Is a Novel Type Efflux Inhibitor Which Attenuates the MexXY-Mediated Aminoglycoside Resistance in Pseudomonas aeruginosa. Front. Microbiol. 2016, 7, 1223. [Google Scholar] [CrossRef] [PubMed]
- Laudadio, E.; Cedraro, N.; Mangiaterra, G.; Citterio, B.; Mobbili, G.; Minnelli, C.; Bizzaro, D.; Biavasco, F.; Galeazzi, R. Natural Alkaloid Berberine Activity against Pseudomonas aeruginosa MexXY-Mediated Aminoglycoside Resistance: In Silico and in Vitro Studies. J. Nat. Prod. 2019, 82, 1935–1944. [Google Scholar] [CrossRef] [PubMed]
- Imanshahidi, M.; Hosseinzadeh, H. Pharmacological and therapeutic effects of Berberis vulgaris and its active constituent, berberine. Phytother. Res. 2008, 22, 999–1012. [Google Scholar] [CrossRef]
- Tegos, G.; Stermitz, F.R.; Lomovskaya, O.; Lewis, K. Multidrug pump inhibitors uncover remarkable activity of plant antimicrobials. Antimicrob. Agents Chemother. 2002, 46, 3133–3141. [Google Scholar] [CrossRef]
- Jing, W.; Jie, T.; Li, F.Y.; Jia, L.; Jing, Y.L.; Fan, Y. Berberine Analogues: Progress towards Versatile Applications. Heterocycles 2015, 91, 2233–2270. [Google Scholar]
- Dolla, N.K.; Chen, C.; Larkins-Ford, J.; Rajamuthiah, R.; Jagadeesan, S.; Conery, A.L.; Ausubel, F.M.; Mylonakis, E.; Bremner, J.B.; Lewis, K.; et al. On the Mechanism of Berberine-INF55 (5-Nitro-2-phenylindole) Hybrid Antibacterials. Aust. J. Chem. 2015, 67, 1471–1480. [Google Scholar] [CrossRef]
- Liu, H.; Wang, L.; Li, Y.; Liu, J.; An, M.; Zhu, S.; Cao, Y.; Jiang, Z.; Zhao, M.; Cai, Z.; et al. Structural optimization of berberine as a synergist to restore antifungal activity of fluconazole against drug-resistant Candida albicans. ChemMedChem 2014, 9, 207–216. [Google Scholar] [CrossRef]
- Makvandi, P.; Gu, J.T.; Zare, E.N.; Ashtari, B.; Moeini, A.; Tay, F.R.; Niu, L.N. Polymeric and inorganic nanoscopical antimicrobial fillers in dentistry. Acta Biomater. 2019. [Google Scholar] [CrossRef] [PubMed]
- Bador, J.; Amoureux, L.; Blanc, E.; Neuwirth, C. Innate aminoglycoside resistance of Achromobacter xylosoxidans is due to AxyXY-OprZ, an RND-type multidrug efflux pump. Antimicrob. Agents Chemother. 2013, 57, 603–605. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Tomida, J.; Kawamura, Y. Responses of Pseudomonas aeruginosa to antimicrobials. Front. Microbiol. 2014, 8, 422. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.H.; Hughes, D.; Poole, K. MexY-promoted aminoglycoside resistance in Pseudomonas aeruginosa: Involvement of a putative proximal binding pocket in aminoglycoside recognition. MBio 2014, 5, e01068. [Google Scholar] [CrossRef] [PubMed]
- Su, F.; Wang, J. Berberine inhibits the MexXY-OprM efflux pump to reverse imipenem resistance in a clinical carbapenem-resistant Pseudomonas aeruginosa isolate in a planktonic state. Exp. Ther. Med. 2018, 15, 467–472. [Google Scholar] [CrossRef]
- Morita, Y.; Tomida, J.; Kawamura, Y. Resistance and Response to Anti-Pseudomonas Agents and Biocides, in Pseudomonas: New Aspects of Pseudomonas Biology, 2nd ed.; Ramos, J., Goldberg, J.B., Filloux, A., Eds.; Springer: New York, NY, USA, 2015; pp. 173–187. [Google Scholar]
- Morita, Y.; Sobel, M.L.; Poole, K. Antibiotic inducibility of the MexXY multidrug efflux system of Pseudomonas aeruginosa: Involvement of the antibiotic-inducible PA5471 gene product. J. Bacteriol. 2006, 188, 1847–1855. [Google Scholar] [CrossRef]
- Roy, P.H.; Tetu, S.G.; Larouche, A.; Elbourne, L.; Tremblay, S.; Ren, Q.; Robert, D.; Derek, H.; Ryan, S.; Kisha, W.; et al. Complete genome sequence of the multiresistant taxonomic outlier Pseudomonas aeruginosa PA7. PLoS ONE 2010, 5, e8842. [Google Scholar] [CrossRef]
- Sobel, M.L.; Mckay, G.A.; Poole, K. Contribution of the MexXY multidrug transporter to aminoglycoside resistance in Pseudomonas aeruginosa clinical isolates. Antimicrob. Agents Chemother. 2003, 47, 3202–3207. [Google Scholar] [CrossRef]
- Miyoshi-Akiyama, T.; Kuwahara, T.; Tada, T.; Kitao, T.; Kirikae, T. Complete genome sequence of highly multidrug-resistant Pseudomonas aeruginosa NCGM2.S1, a representative strain of a cluster endemic to Japan. J. Bacteriol. 2011, 193, 7010. [Google Scholar] [CrossRef]
- Yabuuchi, E.; Kawamura, Y.; Kosako, Y.; Ezaki, T. Emendation of genus Achromobacter and Achromobacter xylosoxidans (Yabuuchi and Yano) and proposal of Achromobacter ruhlandii (Packer and Vishniac) comb. nov., Achromobacter piechaudii (Kiredjian et al.) comb. nov., and Achromobacter xylosoxidans subsp. denitrificans (Ruger and Tan) comb. nov. Microbiol. Immunol. 1998, 42, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Yabuuchi, E.; Kosako, Y.; Oyaizu, H.; Yano, I.; Hotta, H.; Hashimoto, Y.; Ezaki, T.; Arakawa, M. Proposal of Burkholderia gen. nov. and transfer of seven species of the genus Pseudomonas homology group II to the new genus, with the type species Burkholderia cepacia (Palleroni and Holmes 1981) comb. nov. Microbiol. Immunol. 1992, 36, 1251–1275. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wang, L.; Liu, H.; Jiang, Z.; Li, Y.; Jiang, Y.; Zhu, S.; Li, P.; Xie, W.; Cai, Z.; et al. Preparation of Canadine Derivatives as Synergistic Antifungal Agents. 2012. Available online: URL https://worldwide.espacenet.com/publicationDetails/biblio?II=0&ND=3&adjacent=true&locale=en_EP&FT=D&date=20120725&CC=CN&NR=102603755A&KC=A (accessed on 25 July 2012).
- Mahapatra, A.; Maheswari, V.; Kalia, N.P.; Rajput, V.S.; Khan, I.A. Synthesis and antitubercular activity of berberine derivatives. Chem. Nat. Compd. 2014, 50, 321–325. [Google Scholar] [CrossRef]
- Wang, J.; Yang, T.; Chen, H.; Xu, Y.-N.; Yu, L.-F.; Liu, T.; Tang, J.; Yi, Z.; Yang, C.-G.; Xue, W.; et al. The synthesis and antistaphylococcal activity of 9,13-disubstituted berberine derivatives. Eur. J. Med. Chem. 2017, 127, 424–433. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Compound | -R | MIC of (µg/mL) | |
|---|---|---|---|
| PAGUg1927 | PAGUg1931 | ||
| GM2 | - | 1024 | 8 |
| Ber3 | - | >512 | >512 |
| 1 | -H | 256 | 128 |
| 2 | o-Br | 256 | 128 |
| 3 | p-Br | 128 | 64 |
| 4 | o-F | 512 | 256 |
| 5 | o-Cl | 256 | 128 |
| 6 | p-Cl | 256 | 128 |
| 7 | o-CH3 | 512 | 256 |
| 8 | m-CH3 | 256 | 128 |
| 9 | p-CH3 | 256 | 128 |
| 10 | o-NO2 | 512 | 256 |
| 11 | 2,6-Cl | 128 | 64 |
| Concomitant Compound | -R | GM MIC with Berberine Derivative (µg/mL) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PAGUg1927 | PAGUg1931 | ||||||||||||
| 256 * | 128 | 64 | 32 | 16 | 8 | 256 | 128 | 64 | 32 | 16 | 8 | ||
| Ber | − | 128 | 256 | 256 | 512 | - | − | 8 | 8 | 8 | 8 | − | − |
| 1 | -H | − | − | 32 | 64 | 128 | − | − | − | 8 | 8 | 8 | − |
| 2 | o−Br | − | − | 32 | 64 | 64 | − | − | − | 4 | 8 | 8 | − |
| 3 | p−Br | − | − | − | 128 | 256 | 512 | − | − | − | 4 | 8 | 8 |
| 4 | o−F | − | 32 | 64 | 128 | − | − | − | 4 | 8 | 8 | − | − |
| 5 | o−Cl | − | − | 32 | 64 | 128 | − | − | − | 4 | 8 | 8 | − |
| 6 | p−Cl | − | − | 256 | 256 | 256 | − | − | − | 4 | 8 | 8 | − |
| 7 | o−CH3 | − | 16 | 32 | 64 | − | − | − | 4 | 8 | 8 | − | − |
| 8 | m−CH3 | − | − | 256 | 256 | 512 | − | − | − | 4 | 8 | 8 | − |
| 9 | p−CH3 | − | − | 256 | 256 | 512 | − | − | − | 4 | 8 | 8 | − |
| 10 | o−NO2 | − | 128 | 128 | 256 | − | − | − | 8 | 8 | 8 | − | − |
| 11 | 2,6−Cl | − | − | − | 128 | 256 | 512 | − | − | − | 4 | 8 | 8 |
| Drug | MIC in the Presence of Berberine Derivative (µg/mL) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| PAGUg1927 | PAGUg1931 | |||||||||
| Ber | 13-o-MBB | 13−m−MBB | 13−p−MBB | − | Ber | 13-o-MBB | 13−m−MBB | 13−p−MBB | − | |
| AMK | 4 | 2 | 4 | 4 | 8 | 1 | 0.5 | 0.5 | 0.5 | 1 |
| TOB | 0.25 | 0.5 | 0.5 | 0.5 | 0.5 | 0.25 | 0.25 | 0.5 | 0.125 | 0.25 |
| KM | 128 | 64 | 256 | 256 | 512 | 64 | 32 | 32 | 64 | 64 |
| SPCM | 256 | 64 | 128 | 128 | 1024 | 64 | 8 | 8 | 32 | 64 |
| NLFX | 0.25 | 0.0625 | 1 | 1 | 1 | 0.015625 | 0.015625 | 0.015625 | 0.015625 | 0.015625 |
| EM | 128 | 64 | 512 | 256 | 512 | 16 | 16 | 16 | 8 | 16 |
| CBPC | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| EtBr | 512 | 128 | 256 | 256 | 256 | 64 | 8 | 8 | 16 | 64 |
| Tc | 2 | 1 | 8 | 8 | 16 | 0.125 | 0.125 | 0.125 | 0.125 | 0.25 |
| Cp | 4 | 2 | 8 | 8 | 8 | 2 | 2 | 2 | 1 | 2 |
| AZM | 64 | 64 | 256 | 256 | 512 | 8 | 8 | 8 | 4 | 8 |
| CEF | 1 | 0.25 | 0.125 | 2 | 8 | 0.0625 | 0.125 | 0.125 | 0.125 | 0.125 |
| Drug | MIC (µg/mL) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PAGU 1606 | PAGUg1659 | |||||||||||||
| - | Ber (256) 1 | Ber (128) | Ber (64) | 13-o-MBB (256) | 13-o-MBB (128) | 13-o-MBB (64) | - | Ber (256) | Ber (128) | Ber (64) | 13-o-MBB (256) | 13-o-MBB (128) | 13-o-MBB (64) | |
| AMK | 256 | 64 | 128 | 128 | 16 | 32 | 32 | 16 | 8 | 8 | 8 | 8 | 8 | 8 |
| TOB | 256 | 64 | 128 | 128 | 16 | 32 | 32 | 8 | 8 | 8 | 8 | 8 | 8 | 8 |
| KM | >2048 | 1024 | 1024 | 2048 | 256 | 512 | 1024 | 256 | 256 | 256 | 256 | 256 | 256 | 256 |
| GM | 64 | 4 | 8 | 16 | 2 | 4 | 4 | 0.5 | 0.25 | 0.25 | 0.5 | 0.25 | 0.25 | 0.5 |
| SPCM | >2048 | >2048 | >2048 | >2048 | 2048 | >2048 | >2048 | 2048 | 2048 | 2048 | 2048 | 1024 | 2048 | 2048 |
| NLFX | 256 | 256 | 256 | 256 | 256 | 256 | 256 | 64 | 256 | 256 | 256 | 128 | 128 | 128 |
| CPFX | 64 | 64 | 64 | 64 | 32 | 32 | 32 | 64 | 64 | 64 | 64 | 32 | 32 | 32 |
| EM | 256 | 128 | 256 | 256 | 128 | 256 | 256 | 128 | 256 | 256 | 256 | 256 | 256 | 256 |
| CBPC | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| EtBr | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 | >512 |
| Tc | 32 | 16 | 16 | 16 | 16 | 16 | 16 | 16 | 16 | 16 | 16 | 16 | 16 | 16 |
| Cp | 128 | 64 | 128 | 128 | 64 | 64 | 64 | 128 | 128 | 128 | 128 | 128 | 128 | 128 |
| AZM | 256 | 64 | 64 | 128 | 64 | 64 | 128 | 32 | 256 | 256 | 256 | 128 | 128 | 128 |
| CEF | 512 | 512 | 512 | 512 | 512 | 512 | 512 | 512 | 512 | 512 | 512 | 512 | 512 | 512 |
| Strain | MIC of Aminoglycoside (µg/mL) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AMK | GM | TOB | KM | SPEC | |||||||||||
| − | Ber | 13-o-MBB | − | Ber | 13-o-MBB | − | Ber | 13-o-MBB | − | Ber | 13-o-MBB | − | Ber | 13-o-MBB | |
| PAGU 0974 | 4 | 1 | 0.5 | 4 | 0.5 | 0.25 | 0.5 | 0.125 | 0.125 | 128 | 32 | 32 | 512 | 128 | 32 |
| PAGUg 0975 | 1 | 0.5 | 0.5 | 0.25 | 0.125 | 0.25 | 0.25 | 0.25 | 0.125 | 64 | 32 | 32 | 32 | 32 | 32 |
| PAGU 1498 | 32 | 8 | 1 | 1024 | 128 | 8 | 256 | 32 | 8 | >2048 | 512 | 256 | 512 | 128 | 32 |
| PAGUg1565 | 2 | 1 | 1 | 8 | 8 | 8 | 8 | 8 | 8 | 512 | 256 | 256 | 32 | 32 | 32 |
| PAGU 1569 | 256 | 64 | 32 | 256 | 32 | 8 | 16 | 8 | 4 | >2048 | >2048 | 1024 | 512 | 256 | 128 |
| PAGUg1627 | 32 | 32 | 32 | 8 | 8 | 8 | 8 | 8 | 4 | 1024 | 512 | 1024 | 128 | 128 | 128 |
| *PAGU 0013 | 128 | 32 | 4 | 128 | 32 | 4 | 64 | 8 | 1 | 64 | 8 | 2 | 1024 | 128 | 16 |
| PAGU 0002 | >2048 | 256 | 16 | >2048 | 32 | 4 | 512 | 16 | 4 | >2048 | 2048 | 256 | >2048 | 512 | 64 |
| Strain | MIC (µg/mL) for AMK in the Presence of: | MIC (µg/mL) for 13-o-MBB in the Presence of: | FIC | Mode of Interaction | ||
| − | 13-o-MBB | − | AMK | |||
| PAGUg1931 | 1 | 1 | 256 | 256 | 2.0 | Indifferent |
| PAGUg1927 | 8 | 2 | 512 | 128 | 0.5 | Synergy |
| PAGUg1659 | 16 | 8 | >512 | >512 | >1.5 | Indifferent |
| PAGU 1606 | 256 | 16 | >512 | 64 | <0.5 | Synergy |
| Strain | MIC (µg/mL) for AMK in the Presence of: | MIC (µg/mL) for Berberne in the Presence of: | FIC | Mode of Interaction | ||
| − | Berberine | − | AMK | |||
| PAGUg1931 | 1 | 1 | >512 | >512 | >2.0 | Indifferent |
| PAGUg1927 | 8 | 4 | >512 | 512 | <1.0 | Synergy or Addition |
| PAGUg1659 | 16 | 8 | >512 | >512 | 1.5 | Indifferent |
| PAGU 1606 | 256 | 64 | >512 | 512 | <0.75 | Synergy or Addition |
| Strain | MIC (µg/mL) for GM in the Presence of: | MIC (µg/mL) for 13-o-MBB in the Presence of: | FIC | Mode of Interaction | ||
| − | 13-o-MBB | − | GM | |||
| PAGUg1931 | 8 | 8 | 256 | 256 | 2.0 | Indifferent |
| PAGUg1927 | 1024 | 32 | 512 | 4 | 0.04 | Synergy |
| PAGUg1659 | 0.5 | 0.5 | >512 | >512 | >1.0 | Indifferent |
| PAGU 1606 | 64 | 2 | >512 | 8 | <0.5 | Synergy |
| Strain | MIC (µg/mL) for GM in the Presence of: | MIC(µg/mL) for Berberine in the Presence of: | FIC | Mode of Interaction | ||
| − | Berberine | − | GM | |||
| PAGUg1931 | 8 | 8 | >512 | >512 | >1.0 | Indifferent |
| PAGUg1927 | 1024 | 128 | >512 | 8 | <0.5 | Synergy |
| PAGUg1659 | 0.5 | 0.5 | >512 | >512 | >1.0 | Indifferent |
| PAGU 1606 | 64 | 8 | >512 | 256 | <0.5 | Synergy |
| Strain Name | Relevant Characteristics | Reference |
|---|---|---|
| Pseudomonas aeruginosa | ||
| PAGU 0974 | PAO1 (K. Poole Lab), wild type | [28] |
| PAGUg0975 | PAGU 0974ΔmexXY | [29] |
| PAGU 1498 | PA7 Non-respiratory clinical isolate | [8] |
| PAGUg1565 | PA7ΔmexXY-oprA | [8] |
| PAGU 1569 | K2162 Pan-aminoglycoside-resistant clinical isolate | [30] |
| PAGU 1606 | NCGM2. S1 Multidrug-resistant clinical isolate | [31] |
| PAGUg1627 | K2162ΔmexXY | [30] |
| PAGUg1659 | PAGU 1606ΔmexXY | [8] |
| PAGUg1927 | YM34 ΔmexZ, mexVW:: gfp-aacC1 | [15] |
| PAGUg1929 | YM34 ΔmexZ, mexVW | [15] |
| PAGUg1931 | PAGUg1927::ΔmexXY | [15] |
| PAGUg1933 | PAGUg1929::ΔmexXY | [15] |
| Others | ||
| PAGU 0002 | ATCC 27061 Achromobacter xylosoxidans subsp. xylosoxidans | [32] |
| PAGU 0013 | ATCC 25416 Burkholderia cepacia | [33] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotani, K.; Matsumura, M.; Morita, Y.; Tomida, J.; Kutsuna, R.; Nishino, K.; Yasuike, S.; Kawamura, Y. 13-(2-Methylbenzyl) Berberine Is a More Potent Inhibitor of MexXY-Dependent Aminoglycoside Resistance than Berberine. Antibiotics 2019, 8, 212. https://doi.org/10.3390/antibiotics8040212
Kotani K, Matsumura M, Morita Y, Tomida J, Kutsuna R, Nishino K, Yasuike S, Kawamura Y. 13-(2-Methylbenzyl) Berberine Is a More Potent Inhibitor of MexXY-Dependent Aminoglycoside Resistance than Berberine. Antibiotics. 2019; 8(4):212. https://doi.org/10.3390/antibiotics8040212
Chicago/Turabian StyleKotani, Kenta, Mio Matsumura, Yuji Morita, Junko Tomida, Ryo Kutsuna, Kunihiko Nishino, Shuji Yasuike, and Yoshiaki Kawamura. 2019. "13-(2-Methylbenzyl) Berberine Is a More Potent Inhibitor of MexXY-Dependent Aminoglycoside Resistance than Berberine" Antibiotics 8, no. 4: 212. https://doi.org/10.3390/antibiotics8040212