
Molecular Analysis of Carbapenem and Aminoglycoside Resistance Genes in Carbapenem-Resistant Pseudomonas aeruginosa Clinical Strains: A Challenge for Tertiary Care Hospitals


Abstract
:1. Introduction
2. Results
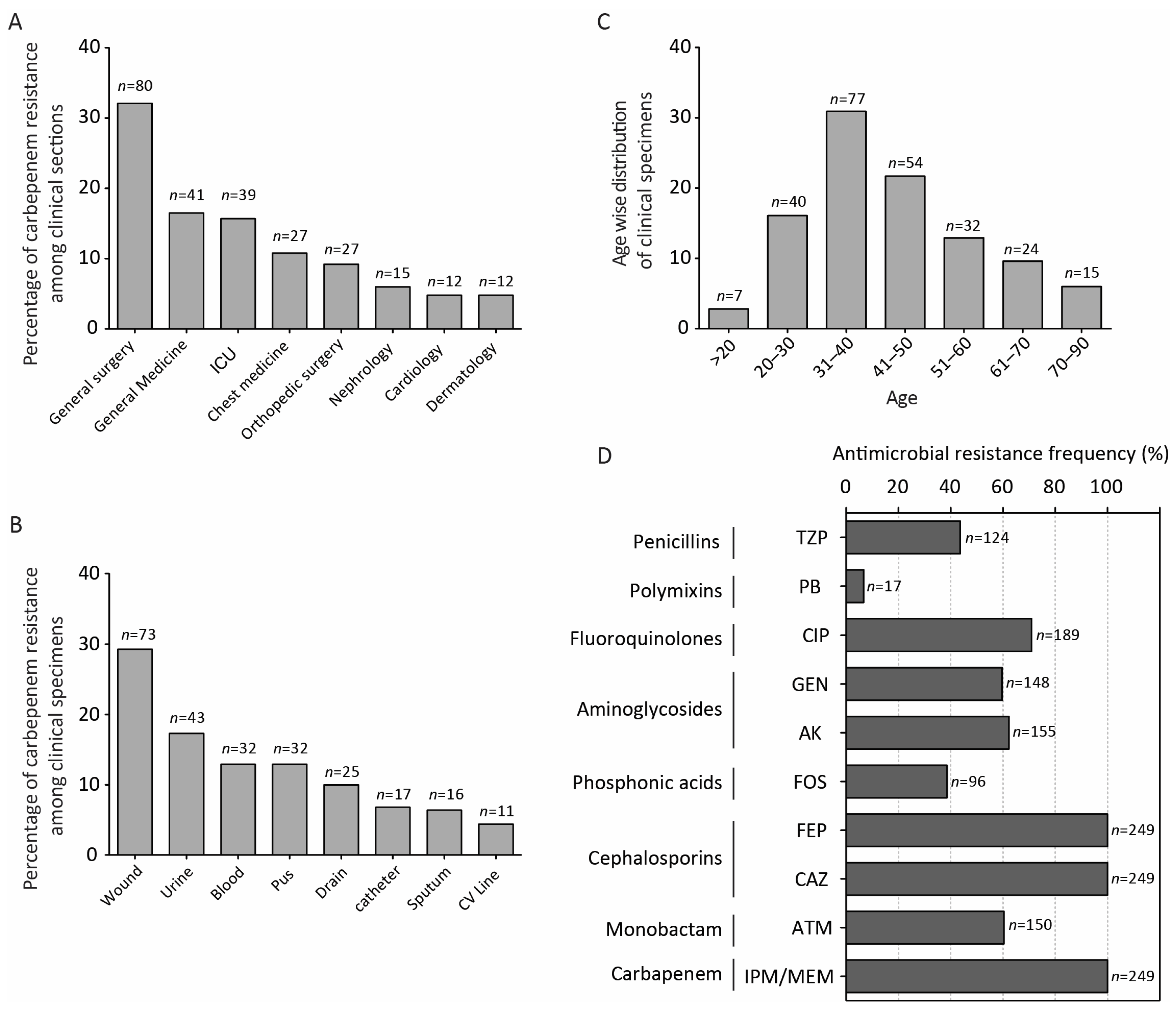
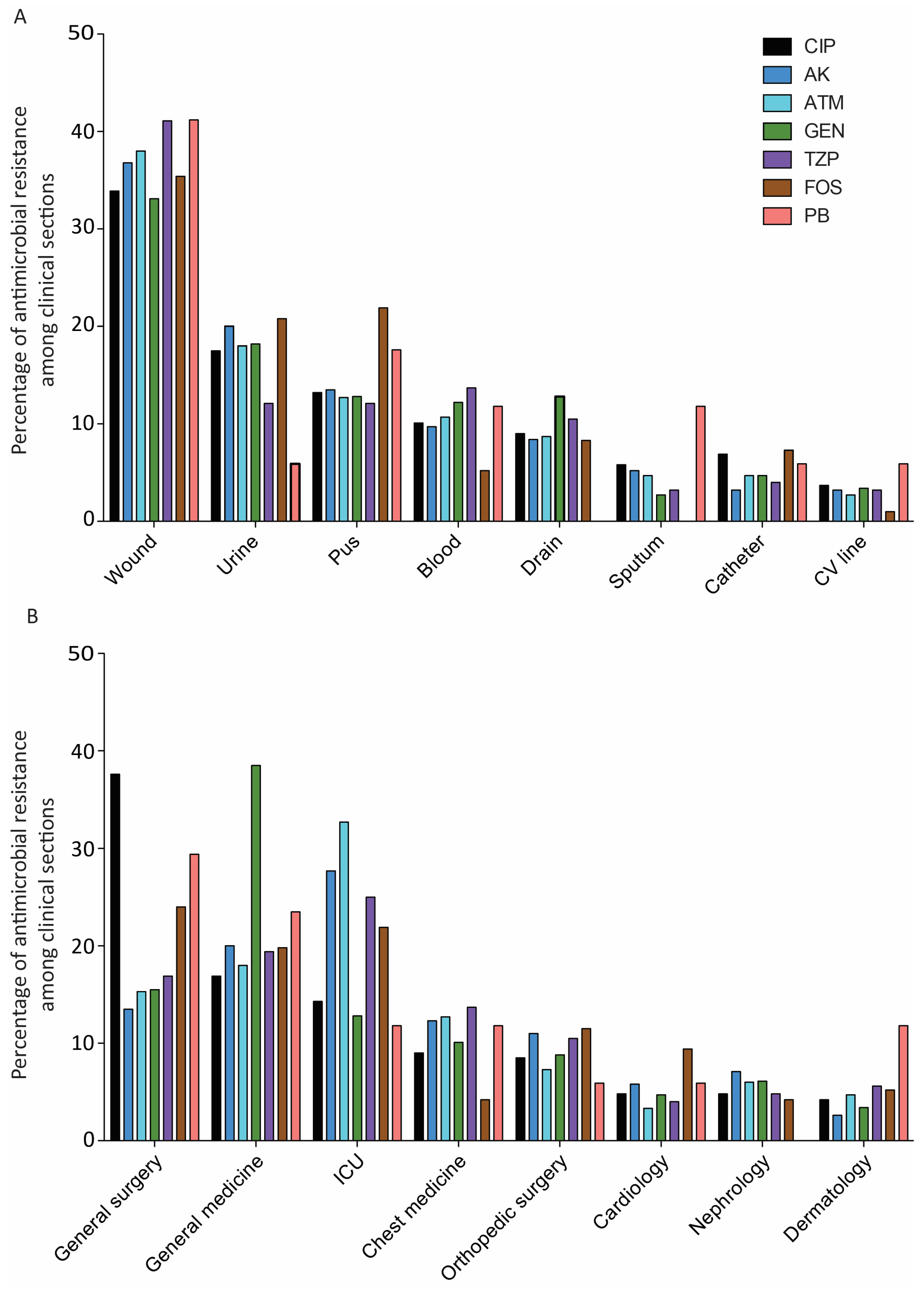
2.1. Characteristics of Bacterial Strains and Antimicrobial Resistance Profile
2.2. Genomic Analysis of Antimicrobial Resistance Genes
2.3. Genetic Variability Profiling and Replicon Typing Analysis
2.4. Antimicrobial Efficacy of Silver Nanoparticles
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sampling and Identification of Clinical Strains
5.2. Antimicrobial Susceptibility Profile Analysis
5.3. Molecular Identification of P. aeruginosa and Antibiotic Resistance Genes Detection
5.4. Determination of blaNDM and blaKPC Alleles
5.5. Multilocus Sequence Typing and Plasmid Typing
5.6. Evaluation of Antimicrobial Activity of AgNPs
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barbier, F.; Andremont, A.; Wolff, M.; Bouadma, L. Hospital-acquired pneumonia and ventilator-associated pneumonia: Recent advances in epidemiology and management. Curr. Opin. Pulm. Med. 2013, 19, 216–228. [Google Scholar] [CrossRef]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Pottier, M.; Gravey, F.; Castagnet, S.; Auzou, M.; Langlois, B.; Guérin, F.; Giard, J.-C.; Léon, A.; Le Hello, S. A 10-year microbiological study of Pseudomonas aeruginosa strains revealed the circulation of populations resistant to both carbapenems and quaternary ammonium compounds. Sci. Rep. 2023, 13, 2639. [Google Scholar] [CrossRef]
- Yasuda, N.; Fujita, T.; Fujioka, T.; Tagawa, M.; Kohira, N.; Torimaru, K.; Shiota, S.; Kumagai, T.; Morita, D.; Ogawa, W.; et al. Effects of the order of exposure to antimicrobials on the incidence of multidrug-resistant Pseudomonas aeruginosa. Sci. Rep. 2023, 13, 8826. [Google Scholar] [CrossRef]
- Cortes-Lara, S.; del Barrio-Tofiño, E.; López-Causapé, C.; Oliver, A.; Martínez-Martínez, L.; Bou, G.; Zamorano, L.; Sánchez-Diener, I.; Galán, F.; Gracia, I. Predicting Pseudomonas aeruginosa susceptibility phenotypes from whole genome sequence resistome analysis. Clin. Microbiol. Infect. 2021, 27, 1631–1637. [Google Scholar] [CrossRef]
- Hwang, W.; Yoon, S.S. Virulence characteristics and an action mode of antibiotic resistance in multidrug-resistant Pseudomonas aeruginosa. Sci. Rep. 2019, 9, 487. [Google Scholar] [CrossRef]
- Ramanathan, B.; Jindal, H.M.; Le, C.F.; Gudimella, R.; Anwar, A.; Razali, R.; Poole-Johnson, J.; Manikam, R.; Sekaran, S.D. Next generation sequencing reveals the antibiotic resistant variants in the genome of Pseudomonas aeruginosa. PLoS ONE 2017, 12, e0182524. [Google Scholar] [CrossRef]
- Treepong, P.; Kos, V.; Guyeux, C.; Blanc, D.; Bertrand, X.; Valot, B.; Hocquet, D. Global emergence of the widespread Pseudomonas aeruginosa ST235 clone. Clin. Microbiol. Infect. 2018, 24, 258–266. [Google Scholar] [CrossRef]
- Lodise, T.P.; Bassetti, M.; Ferrer, R.; Naas, T.; Niki, Y.; Paterson, D.L.; Zeitlinger, M.; Echols, R. All-cause mortality rates in adults with carbapenem-resistant Gram-negative bacterial infections: A comprehensive review of pathogen-focused, prospective, randomized, interventional clinical studies. Expert Rev. Anti-Infect. Ther. 2022, 20, 707–719. [Google Scholar] [CrossRef]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Rossolini, G.M.; Bochenska, M.; Fumagalli, L.; Dowzicky, M. Trends of major antimicrobial resistance phenotypes in enterobacterales and gram-negative non-fermenters from ATLAS and EARS-net surveillance systems: Italian vs. European and global data, 2008–2018. Diagn. Microbiol. Infect. Dis. 2021, 101, 115512. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, D.; Chen, K.; Xie, M.; Guo, J.; Chan, E.W.C.; Xie, L.; Wang, J.; Chen, E.; Chen, S.; et al. Epidemiological and genetic characteristics of clinical carbapenem-resistant Pseudomonas aeruginosa strains in Guangdong Province, China. Microbiol. Spectr. 2023, 11, e04261-22. [Google Scholar] [CrossRef]
- Abdeta, A.; Negeri, A.A.; Beyene, D.; Adamu, E.; Fekede, E.; Fentaw, S.; Tesfaye, M.; Wakoya, G.K. Prevalence and trends of carbapenem-resistant Pseudomonas aeruginosa and Acinetobacter species isolated from clinical specimens at the Ethiopian public health institute, Addis Ababa, Ethiopia: A Retrospective Analysis. Infect. Drug Resist. 2023, 16, 1381–1390. [Google Scholar] [CrossRef]
- Lee, Y.-L.; Ko, W.-C.; Hsueh, P.-R. Geographic patterns of carbapenem-resistant Pseudomonas aeruginosa in the Asia-Pacific Region: Results from the Antimicrobial Testing Leadership and Surveillance (ATLAS) program, 2015–2019. Antimicrob. Agents Chemother. 2022, 66, e02000-21. [Google Scholar] [CrossRef]
- Ssekatawa, K.; Byarugaba, D.K.; Wampande, E.; Ejobi, F. A systematic review: The current status of carbapenem resistance in East Africa. BMC Res. Notes 2018, 11, 629. [Google Scholar] [CrossRef]
- Kunz Coyne, A.J.; El Ghali, A.; Holger, D.; Rebold, N.; Rybak, M.J. Therapeutic strategies for emerging multidrug-resistant Pseudomonas aeruginosa. Infect. Dis. Ther. 2022, 11, 661–682. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Reyes, J.; Komarow, L.; Chen, L.; Ge, L.; Hanson, B.M.; Cober, E.; Herc, E.; Alenazi, T.; Kaye, K.S.; Garcia-Diaz, J.; et al. Global epidemiology and clinical outcomes of carbapenem-resistant Pseudomonas aeruginosa and associated carbapenemases (POP): A prospective cohort study. Lancet Microbe 2023, 4, e159–e170. [Google Scholar] [CrossRef]
- Van Duin, D.; Arias, C.A.; Komarow, L.; Chen, L.; Hanson, B.M.; Weston, G.; Cober, E.; Garner, O.B.; Jacob, J.T.; Satlin, M.J.; et al. Molecular and clinical epidemiology of carbapenem-resistant Enterobacterales in the USA (CRACKLE-2): A prospective cohort study. Lancet Infect. Dis. 2020, 20, 731–741. [Google Scholar] [CrossRef]
- Castanheira, M.; Deshpande, L.M.; Costello, A.; Davies, T.A.; Jones, R.N. Epidemiology and carbapenem resistance mechanisms of carbapenem-non-susceptible Pseudomonas aeruginosa collected during 2009–11 in 14 European and Mediterranean countries. J. Antimicrob. Chemother. 2014, 69, 1804–1814. [Google Scholar] [CrossRef]
- Gondal, A.J.; Choudhry, N.; Bukhari, H.; Rizvi, Z.; Jahan, S.; Yasmin, N. Estimation, evaluation and characterization of carbapenem resistanceburden from a tertiary care hospital, Pakistan. Antibiotics 2023, 12, 525. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Liu, C.; Wang, Q.; Zeng, Y.; Sun, Q.; Shu, L.; Lu, J.; Cai, J.; Wang, S.; Zhang, R.; et al. Emergence and expansion of a carbapenem-resistant Pseudomonas aeruginosa clone are associated with plasmid-borne blaKPC-2 and virulence-related genes. Msystems 2021, 6, e00154-21. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, R.; Inoue-Tsuda, M.; Matsui, H.; Ito, T.; Hanaki, H. Classification of the metallo β-lactamase subtype produced by the carbapenem-resistant Pseudomonas aeruginosa isolates in Japan. J. Infect. Chemother. 2022, 28, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Ngoi, S.T.; Chong, C.W.; Ponnampalavanar, S.S.L.S.; Tang, S.N.; Idris, N.; Abdul Jabar, K.; Gregory, M.J.; Husain, T.; Teh, C.S.J. Genetic mechanisms and correlated risk factors of antimicrobial-resistant ESKAPEE pathogens isolated in a tertiary hospital in Malaysia. Antimicrob. Resist. Infect. Control 2021, 10, 70. [Google Scholar] [CrossRef]
- Park, Y.; Koo, S.H. Epidemiology, molecular characteristics, and virulence factors of carbapenem-resistant Pseudomonas aeruginosa isolated from patients with urinary tract infections. Infect. Drug Resist. 2022, 15, 141–151. [Google Scholar] [CrossRef]
- Ramadan, H.K.-A.; Mahmoud, M.A.; Aburahma, M.Z.; Elkhawaga, A.A.; El-Mokhtar, M.A.; Sayed, I.M.; Hosni, A.; Hassany, S.M.; Medhat, M.A. Predictors of severity and co-infection resistance profile in COVID-19 patients: First report from upper Egypt. Infect. Drug Resist. 2020, 13, 3409–3422. [Google Scholar] [CrossRef]
- Tartari, D.C.; Zamparette, C.P.; Martini, G.; Christakis, S.; Costa, L.H.; de Oliveira Silveira, A.C.; Sincero, T.C.M. Genomic analysis of an extensively drug-resistant Pseudomonas aeruginosa ST312 harbouring IncU plasmid-mediated blaKPC-2 isolated from ascitic fluid. J. Glob. Antimicrob. Resist. 2021, 25, 151–153. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Wang, D.; Zhu, Y.; Li, J.; Jiang, Y.; Wu, W.; Li, X.; Zhou, H. Emergence of a KPC-90 variant that confers resistance to ceftazidime-avibactam in an ST463 carbapenem-resistant Pseudomonas aeruginosa strain. Microbiol. Spectr. 2022, 10, e01869-21. [Google Scholar] [CrossRef] [PubMed]
- Wendel, A.F.; Malecki, M.; Mattner, F.; Xanthopoulou, K.; Wille, J.; Seifert, H.; Higgins, P.G. Genomic-based transmission analysis of carbapenem-resistant Pseudomonas aeruginosa at a tertiary care centre in Cologne (Germany) from 2015 to 2020. JAC-Antimicrob. Resist. 2022, 4, dlac057. [Google Scholar] [CrossRef]
- Giani, T.; Arena, F.; Pollini, S.; Di Pilato, V.; D’Andrea, M.M.; Henrici De Angelis, L.; Bassetti, M.; Rossolini, G.M. Italian nationwide survey on Pseudomonas aeruginosa from invasive infections: Activity of ceftolozane/tazobactam and comparators, and molecular epidemiology of carbapenemase producers. J. Antimicrob. Chemother. 2018, 73, 664–671. [Google Scholar] [CrossRef]
- Ratjen, F.; Brockhaus, F.; Angyalosi, G. Aminoglycoside therapy against Pseudomonas aeruginosa in cystic fibrosis: A review. J. Cyst. Fibros. 2009, 8, 361–369. [Google Scholar] [CrossRef]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside modifying enzymes. Drug Resist. Updat. 2010, 13, 151–171. [Google Scholar] [CrossRef]
- Atassi, G.; Medernach, R.; Scheetz, M.; Nozick, S.; Rhodes, N.J.; Murphy-Belcaster, M.; Murphy, K.R.; Alisoltani, A.; Ozer, E.A.; Hauser, A.R. Genomics of aminoglycoside resistance in Pseudomonas aeruginosa bloodstream infections at a United States Academic Hospital. Microbiol. Spectr. 2023, 11, e05087-22. [Google Scholar] [CrossRef]
- Panahi, T.; Asadpour, L.; Ranji, N. Distribution of aminoglycoside resistance genes in clinical isolates of Pseudomonas aeruginosa in north of Iran. Gene Rep. 2020, 21, 100929. [Google Scholar] [CrossRef]
- Doi, Y.; Wachino, J.-I.; Arakawa, Y. Aminoglycoside resistance: The emergence of acquired 16S ribosomal RNA methyltransferases. Infect. Dis. Clin. 2016, 30, 523–537. [Google Scholar] [CrossRef]
- Taylor, E.; Jauneikaite, E.; Sriskandan, S.; Woodford, N.; Hopkins, K.L. Detection and characterisation of 16S rRNA methyltransferase-producing Pseudomonas aeruginosa from the UK and Republic of Ireland from 2003–2015. Int. J. Antimicrob. Agents 2022, 59, 106550. [Google Scholar] [CrossRef] [PubMed]
- Kocsis, B.; Gulyás, D.; Szabó, D. Diversity and distribution of resistance markers in Pseudomonas aeruginosa international high-risk clones. Microorganisms 2021, 9, 359. [Google Scholar] [CrossRef] [PubMed]
- del Barrio-Tofiño, E.; López-Causapé, C.; Oliver, A. Pseudomonas aeruginosa epidemic high-risk clones and their association with horizontally-acquired β-lactamases: 2020 update. Int. J. Antimicrob. Agents 2020, 56, 106196. [Google Scholar] [CrossRef]
- Aguilar-Rodea, P.; Estrada-Javier, E.L.; Jiménez-Rojas, V.; Gomez-Ramirez, U.; Nolasco-Romero, C.G.; Rodea, G.E.; Rodríguez-Espino, B.A.; Mendoza-Elizalde, S.; Arellano, C.; López-Marcelino, B.; et al. New variants of Pseudomonas aeruginosa high-risk clone ST233 associated with an outbreak in a Mexican Paediatric Hospital. Microorganisms 2022, 10, 1533. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.-J.; Jeong, S.H. Mobile carbapenemase genes in Pseudomonas aeruginosa. Front. Microbiol. 2021, 12, 614058. [Google Scholar] [CrossRef] [PubMed]
- Bogaerts, P.; Bauraing, C.; Deplano, A.; Glupczynski, Y. Emergence and dissemination of BEL-1-producing Pseudomonas aeruginosa isolates in Belgium. Antimicrob. Agents Chemother. 2007, 51, 1584. [Google Scholar] [CrossRef]
- Libisch, B.; Poirel, L.; Lepsanovic, Z.; Mirovic, V.; Balogh, B.; Pászti, J.; Hunyadi, Z.; Dobák, A.; Füzi, M.; Nordmann, P. Identification of PER-1 extended-spectrum β-lactamase producing Pseudomonas aeruginosa clinical isolates of the international clonal complex CC11 from Hungary and Serbia. FEMS Microbiol. Immunol. 2008, 54, 330–338. [Google Scholar] [CrossRef]
- Viedma, E.; Juan, C.; Acosta, J.; Zamorano, L.; Otero, J.R.; Sanz, F.; Chaves, F.; Oliver, A. Nosocomial spread of colistin-only-sensitive sequence type 235 Pseudomonas aeruginosa isolates producing the extended-spectrum β-lactamases GES-1 and GES-5 in Spain. Antimicrob. Agents Chemother. 2009, 53, 4930–4933. [Google Scholar] [CrossRef] [PubMed]
- Samuelsen, Ø.; Toleman, M.A.; Sundsfjord, A.; Rydberg, J.; Leegaard, T.M.; Walder, M.; Lia, A.; Ranheim, T.E.; Rajendra, Y.; Hermansen, N.O.; et al. Molecular epidemiology of metallo-β-lactamase-producing Pseudomonas aeruginosa isolates from Norway and Sweden shows import of international clones and local clonal expansion. Antimicrob. Agents Chemother. 2010, 54, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Kos, V.N.; Déraspe, M.; McLaughlin, R.E.; Whiteaker, J.D.; Roy, P.H.; Alm, R.A.; Corbeil, J.; Gardner, H. The resistome of Pseudomonas aeruginosa in relationship to phenotypic susceptibility. Antimicrob. Agents Chemother. 2015, 59, 427–436. [Google Scholar] [CrossRef]
- Pfennigwerth, N.; Geis, G.; Gatermann, S.G.; Kaase, M. Description of IMP-31, a novel metallo-β-lactamase found in an ST235 Pseudomonas aeruginosa strain in Western Germany. J. Antimicrob. Chemother. 2015, 70, 1973–1980. [Google Scholar] [CrossRef]
- Zamorano, L.; Mulet, X.; Garau, M.; Gallegos, C.; Juan, C.; Pérez, J.L.; Oliver, A. VIM-47, a new variant of the autochthonous metallo-β-lactamase VIM-13 from the Balearic Islands in Spain. Antimicrob. Agents Chemother. 2016, 60, 3251. [Google Scholar] [CrossRef] [PubMed]
- de Paula-Petroli, S.B.; Campana, E.H.; Bocchi, M.; Bordinhão, T.; Picão, R.C.; Yamada-Ogatta, S.F.; Carrara-Marroni, F.E. Early detection of a hypervirulent KPC-2-producing Pseudomonas aeruginosa ST235 in Brazil. J. Glob. Antimicrob. Resist. 2018, 12, 153–154. [Google Scholar] [CrossRef]
- Tada, T.; Nhung, P.H.; Miyoshi-Akiyama, T.; Shimada, K.; Tsuchiya, M.; Phuong, D.M.; Anh, N.Q.; Ohmagari, N.; Kirikae, T. Multidrug-resistant sequence type 235 Pseudomonas aeruginosa clinical isolates producing IMP-26 with increased carbapenem-hydrolyzing activities in Vietnam. Antimicrob. Agents Chemother. 2016, 60, 6853–6858. [Google Scholar] [CrossRef]
- Correa, A.; Montealegre, M.C.; Mojica, M.F.; Maya, J.J.; Rojas, L.J.; De La Cadena, E.P.; Ruiz, S.J.; Recalde, M.; Rosso, F.; Quinn, J.P.; et al. First report of a Pseudomonas aeruginosa isolate coharboring KPC and VIM carbapenemases. Antimicrob. Agents Chemother. 2012, 56, 5422. [Google Scholar] [CrossRef]
- Turton, J.F.; Wright, L.; Underwood, A.; Witney, A.A.; Chan, Y.-T.; Al-Shahib, A.; Arnold, C.; Doumith, M.; Patel, B.; Planche, T.D.; et al. High-resolution analysis by whole-genome sequencing of an international lineage (sequence type 111) of Pseudomonas aeruginosa associated with metallo-carbapenemases in the United Kingdom. J. Clin. Microbiol. 2015, 53, 2622–2631. [Google Scholar] [CrossRef]
- Wendel, A.F.; Kolbe-Busch, S.; Ressina, S.; Schulze-Röbbecke, R.; Kindgen-Milles, D.; Lorenz, C.; Pfeffer, K.; MacKenzie, C.R. Detection and termination of an extended low-frequency hospital outbreak of GIM-1–producing Pseudomonas aeruginosa ST111 in Germany. Am. J. Infect. Control 2015, 43, 635–639. [Google Scholar] [CrossRef]
- Guzvinec, M.; Izdebski, R.; Butic, I.; Jelic, M.; Abram, M.; Koscak, I.; Baraniak, A.; Hryniewicz, W.; Gniadkowski, M.; Tambic Andrasevic, A. Sequence types 235, 111, and 132 predominate among multidrug-resistant Pseudomonas aeruginosa clinical isolates in Croatia. Antimicrob. Agents Chemother. 2014, 58, 6277–6283. [Google Scholar] [CrossRef]
- Vatcheva-Dobrevska, R.; Mulet, X.; Ivanov, I.; Zamorano, L.; Dobreva, E.; Velinov, T.; Kantardjiev, T.; Oliver, A. Molecular epidemiology and multidrug resistance mechanisms of Pseudomonas aeruginosa isolates from Bulgarian hospitals. Microb. Drug Resist. 2013, 19, 355–361. [Google Scholar] [CrossRef]
- Cottarel, G.; Wierzbowski, J. Combination drugs, an emerging option for antibacterial therapy. Trends Biotechnol. 2007, 25, 547–555. [Google Scholar] [CrossRef]
- Vilchèze, C.; Jacobs, W.R., Jr. The combination of sulfamethoxazole, trimethoprim, and isoniazid or rifampin is bactericidal and prevents the emergence of drug resistance in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2012, 56, 5142–5148. [Google Scholar] [CrossRef]
- Abeer Mohammed, A.; Abd Elhamid, M.M.; Khalil, M.K.M.; Ali, A.S.; Abbas, R.N. The potential activity of biosynthesized silver nanoparticles of Pseudomonas aeruginosa as an antibacterial agent against multidrug-resistant isolates from intensive care unit and anticancer agent. Environ. Sci. Eur. 2022, 34, 109. [Google Scholar] [CrossRef]
- Wypij, M.; Czarnecka, J.; Świecimska, M.; Dahm, H.; Rai, M.; Golinska, P. Synthesis, characterization and evaluation of antimicrobial and cytotoxic activities of biogenic silver nanoparticles synthesized from Streptomyces xinghaiensis OF1 strain. World J. Microbiol. Biotechnol. 2018, 34, 23. [Google Scholar] [CrossRef] [PubMed]
- Campo-Beleño, C.; Villamizar-Gallardo, R.; López-Jácome, L.; González, E.; Muñoz-Carranza, S.; Franco, B.; Morales-Espinosa, R.; Coria-Jimenez, R.; Franco-Cendejas, R.; Hernández-Durán, M.; et al. Biologically synthesized silver nanoparticles as potent antibacterial effective against multidrug-resistant Pseudomonas aeruginosa. Lett. Appl. Microbiol. 2022, 75, 680–688. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.; Zhang, Y.; Pan, X.; Zhu, F.; Jiang, C.; Liu, Q.; Cheng, Z.; Dai, G.; Wu, G.; Wang, L.; et al. Antibacterial activity and mechanism of silver nanoparticles against multidrug-resistant Pseudomonas aeruginosa. Int. J. Nanomed. 2019, 14, 1469–1487. [Google Scholar] [CrossRef] [PubMed]
- Orlov, I.A.; Sankova, T.P.; Babich, P.S.; Sosnin, I.M.; Ilyechova, E.Y.; Kirilenko, D.A.; Brunkov, P.N.; Ataev, G.L.; Romanov, A.E.; Puchkova, L.V. New silver nanoparticles induce apoptosis-like process in E. coli and interfere with mammalian copper metabolism. Int. J. Nanomed. 2016, 11, 6561–6574. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, E.; MacFadden, D.R.; Lee, R.S.; Lipsitch, M. Outpatient prescribing of four major antibiotic classes and prevalence of antimicrobial resistance in US adults. bioRxiv 2018. [Google Scholar] [CrossRef]
- Sanz-García, F.; Hernando-Amado, S.; Martínez, J.L. Mutational evolution of Pseudomonas aeruginosa resistance to ribosome-targeting antibiotics. Front. Genet. 2018, 9, 451. [Google Scholar] [CrossRef] [PubMed]
- Dou, Y.; Huan, J.; Guo, F.; Zhou, Z.; Shi, Y. Pseudomonas aeruginosa prevalence, antibiotic resistance and antimicrobial use in Chinese burn wards from 2007 to 2014. J. Int. Med. Res. 2017, 45, 1124–1137. [Google Scholar] [CrossRef] [PubMed]
- Farhan, S.M.; Ibrahim, R.A.; Mahran, K.M.; Hetta, H.F.; Abd El-Baky, R.M. Antimicrobial resistance pattern and molecular genetic distribution of metallo-β-lactamases producing Pseudomonas aeruginosa isolated from hospitals in Minia, Egypt. Infect. Drug Resist. 2019, 12, 2125–2133. [Google Scholar] [CrossRef]
- Hosu, M.C.; Vasaikar, S.D.; Okuthe, G.E.; Apalata, T. Detection of extended spectrum beta-lactamase genes in Pseudomonas aeruginosa isolated from patients in rural Eastern Cape Province, South Africa. Sci. Rep. 2021, 11, 7110. [Google Scholar] [CrossRef]
- Nathwani, D.; Raman, G.; Sulham, K.; Gavaghan, M.; Menon, V. Clinical and economic consequences of hospital-acquired resistant and multidrug-resistant Pseudomonas aeruginosa infections: A systematic review and meta-analysis. Antimicrob. Resist. Infect. Control 2014, 3, 32. [Google Scholar] [CrossRef]
- Nørgaard, S.M.; Jensen, C.S.; Aalestrup, J.; Vandenbroucke-Grauls, C.M.; de Boer, M.G.; Pedersen, A.B. Choice of therapeutic interventions and outcomes for the treatment of infections caused by multidrug-resistant gram-negative pathogens: A systematic review. Antimicrob. Resist. Infect. Control 2019, 8, 170. [Google Scholar] [CrossRef]
- Farooq, L.; Memon, Z.; Ismail, M.O.; Sadiq, S. Frequency and antibiogram of multi-drug resistant Pseudomonas aeruginosa in a Tertiary Care Hospital of Pakistan. Pak. J. Med. Sci. 2019, 35, 1622–1626. [Google Scholar] [CrossRef]
- Khan, F.; Khan, A.; Kazmi, S.U. Prevalence and susceptibility pattern of multi drug resistant clinical isolates of Pseudomonas aeruginosa in Karachi. Pak. J. Med. Sci. 2014, 30, 951–954. [Google Scholar] [CrossRef] [PubMed]
- Saleem, S.; Bokhari, H. Resistance profile of genetically distinct clinical Pseudomonas aeruginosa isolates from public hospitals in central Pakistan. J. Infect. Public Health 2020, 13, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.R.; Azam, S.; Ahmad, S.; Ali, Q.; Liaqat, Z.; Rehman, N.; Khan, I.; Alharbi, M.; Alshammari, A. Molecular Characterization and epidemiology of antibiotic resistance genes of β-lactamase producing bacterial pathogens causing septicemia from tertiary care hospitals. Antibiotics 2023, 12, 617. [Google Scholar] [CrossRef] [PubMed]
- El-Far, A.; Samir, S.; El-Gebaly, E.; Omar, M.; Dahroug, H.; El-Shenawy, A.; Soliman, N.S.; Gamal, D. High rates of aminoglycoside methyltransferases associated with metallo-beta-lactamases in multidrug-resistant and extensively drug-resistant Pseudomonas aeruginosa clinical isolates from a tertiary care hospital in Egypt. Infect. Drug Resist. 2021, 14, 4849–4858. [Google Scholar] [CrossRef] [PubMed]
- Çekin, Z.K.; Dabos, L.; Malkoçoğlu, G.; Fortineau, N.; Bayraktar, B.; Iorga, B.I.; Naas, T.; Aktaş, E. Carbapenemase-producing Pseudomonas aeruginosa isolates from Turkey: First report of P. aeruginosa high-risk clones with VIM-5–and IMP-7–type carbapenemases in a tertiary hospital. Diagn. Microbiol. Infect. Dis. 2021, 99, 115174. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.; Chen, X.; Dong, S.; Mai, H.; Lopes, B.S.; Liu, S.; Wen, F. Resistance genomics and molecular epidemiology of high-risk clones of ESBL-producing Pseudomonas aeruginosa in young children. Front. Cell. Infect. Microbiol. 2023, 13, 1168096. [Google Scholar] [CrossRef] [PubMed]
- Olalekan, A.; Bader, B.K.; Iwalokun, B.; Wolf, S.; Lalremruata, A.; Dike, A.; Mannie-Udoh, M.; Lo Presti, L.; Liese, J.; Guther, J.; et al. High incidence of carbapenemase-producing Pseudomonas aeruginosa clinical isolates from Lagos, Nigeria. JAC Antimicrob. Resist. 2023, 5, dlad038. [Google Scholar] [CrossRef]
- Nasrin, S.; Hegerle, N.; Sen, S.; Nkeze, J.; Sen, S.; Permala-Booth, J.; Choi, M.; Sinclair, J.; Tapia, M.D.; Johnson, J.K.; et al. Distribution of serotypes and antibiotic resistance of invasive Pseudomonas aeruginosa in a multi-country collection. BMC Microbiol. 2022, 22, 13. [Google Scholar] [CrossRef]
- Shortridge, D.; Gales, A.C.; Streit, J.M.; Huband, M.D.; Tsakris, A.; Jones, R.N. Geographic and temporal patterns of antimicrobial resistance in Pseudomonas aeruginosa over 20 years from the SENTRY antimicrobial surveillance program, 1997–2016. Open Forum Infect. Dis. 2019, 15, S63–S68. [Google Scholar] [CrossRef]
- Ali, F.; Mirza, I.A.; Waseem, H.; Zafar, H. Antibiotic sensitivity pattern of carbapenem-resistant Pseudomonas aeruginosa. Pak. Armed Forces Med. J. 2023, 73, 33–37. [Google Scholar] [CrossRef]
- Hu, Y.-Y.; Cao, J.-M.; Yang, Q.; Chen, S.; Lv, H.-Y.; Zhou, H.-W.; Wu, Z.; Zhang, R. Risk factors for carbapenem-resistant Pseudomonas aeruginosa, Zhejiang Province, China. Emerg. Infect. Dis. 2019, 25, 1861. [Google Scholar] [CrossRef]
- Uc-Cachón, A.H.; Gracida-Osorno, C.; Luna-Chi, I.G.; Jiménez-Guillermo, J.G.; Molina-Salinas, G.M. High prevalence of antimicrobial resistance among gram-negative isolated bacilli in intensive care units at a tertiary-care hospital in Yucatán Mexico. Medicina 2019, 55, 588. [Google Scholar] [CrossRef] [PubMed]
- Djuikoue, C.I.; Djouela Djoulako, P.D.; Same Njanjo, H.V.; Kiyang, C.P.; Djantou Biankeu, F.L.; Guegang, C.; Tchouotou, A.S.D.; Wouambo, R.K.; Thumamo Pokam, B.D.; Apalata, T.; et al. Phenotypic Characterization and Prevalence of Carbapenemase-producing Pseudomonas aeruginosa isolates in six health facilities in Cameroon. BioMed 2023, 3, 77–88. [Google Scholar] [CrossRef]
- Rostami, S.; Sheikh, A.F.; Shoja, S.; Farahani, A.; Tabatabaiefar, M.A.; Jolodar, A.; Sheikhi, R. Investigating of four main carbapenem-resistance mechanisms in high-level carbapenem resistant Pseudomonas aeruginosa isolated from burn patients. J. Chin. Med. Assoc. 2018, 81, 127–132. [Google Scholar] [CrossRef]
- Huang, W.; Wei, X.; Xu, G.; Zhang, X.; Wang, X. Carbapenem-resistant Pseudomonas aeruginosa infections in critically ill children: Prevalence, risk factors, and impact on outcome in a large tertiary pediatric hospital of China. Front. Public Health 2023, 11, 1088262. [Google Scholar] [CrossRef] [PubMed]
- Din, M.; Awan, M.A.; ur Rahman, S.; Ali, M.; Aslam But, M. Co-existence of blaIMP, blaNDM-1, and blaSHV, genes of Pseudomonas aeruginosa isolated from Quetta: Antimicrobial resistance and clinical significance. Pak. J. Med. Sci. 2023, 39, 1507. [Google Scholar]
- Kumar, S.; Kothari, A.; Omar, B.J.; Kiran, K. Detection of extended-spectrum beta-lactamase (ESBL) production by disc diffusion method among Pseudomonas species from various clinical samples. J. Family Med. Prim. Care 2020, 9, 683–693. [Google Scholar] [CrossRef]
- Villedieu, A.; Papesh, E.; Weinberg, S.E.; Teare, L.; Radhakrishnan, J.; Elamin, W.F. Seasonal variation of Pseudomonas aeruginosa in culture positive otitis externa in South East England. Epidemiol. Infect. 2018, 146, 1811–1812. [Google Scholar] [CrossRef]
- Sannathimmappa, M.B.; Nambiar, V.; Aravindakshan, R. A 5-Year Retrospective Study of Antibiotic Resistance Pattern of Pseudomonas aeruginosa Isolated from Various Clinical Samples at a Tertiary Care Hospital. Med. J. Dr. D. Y. Patil Vidyapeeth 2023, 16, 687–692. [Google Scholar] [CrossRef]
- Wang, M.-G.; Liu, Z.-Y.; Liao, X.-P.; Sun, R.-Y.; Li, R.-B.; Liu, Y.; Fang, L.-X.; Sun, J.; Liu, Y.-H.; Zhang, R.-M. Retrospective data insight into the global distribution of carbapenemase-producing Pseudomonas aeruginosa. Antibiotics 2021, 10, 548. [Google Scholar] [CrossRef]
- Simner, P.J.; Opene, B.N.; Chambers, K.K.; Naumann, M.E.; Carroll, K.C.; Tamma, P.D. Carbapenemase detection among carbapenem-resistant glucose nonfermenting Gram-negative bacilli. J. Clin. Microbiol. 2017, 55, 2858–2864. [Google Scholar] [CrossRef]
- Tenover, F.C.; Nicolau, D.P.; Gill, C.M. Carbapenemase-producing Pseudomonas aeruginosa–an emerging challenge. Emerg. Microbes Infect. 2022, 11, 811–814. [Google Scholar] [CrossRef] [PubMed]
- Awan, M.; Rasheed, F.; Saeed, M.; Irum, S.; Ashraf, F.; Imran, A.A. Dissemination and detection of carbapenemases producing Gram-negative rods. Pak. Armed Forces Med. J. 2019, 69, 9–14. [Google Scholar]
- Hadjadj, L.; Syed, M.A.; Abbasi, S.A.; Rolain, J.-M.; Jamil, B. Diversity of carbapenem resistance mechanisms in clinical Gram-negative bacteria in Pakistan. Microb. Drug Resist. 2021, 27, 760–767. [Google Scholar] [CrossRef]
- Ahmed, N.; Ali, Z.; Riaz, M.; Zeshan, B.; Wattoo, J.I.; Aslam, M.N. Evaluation of antibiotic resistance and virulence genes among clinical isolates of Pseudomonas aeruginosa from cancer patients. Asian Pac. J. Cancer Prev. 2020, 21, 1333–1338. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Devkota, M.D.; Pokhrel, B.M.; Banjara, M.R. Detection of blaNDM−1, mcr-1 and MexB in multidrug resistant Pseudomonas aeruginosa isolated from clinical specimens in a tertiary care hospital of Nepal. BMC Microbiol. 2023, 23, 153. [Google Scholar] [CrossRef]
- Fortunato, G.; Vaz-Moreira, I.; Gajic, I.; Manaia, C.M. Insight into phylogenomic bias of blaVIM-2 or blaNDM-1 dissemination amongst carbapenem-resistant Pseudomonas aeruginosa. Int. J. Antimicrob. Agents 2023, 61, 106788. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kim, Y.A.; Junglim, K.; Jeong, S.H.; Shin, J.H.; Shin, K.S.; Shin, J.H.; Kim, Y.R.; Kim, H.S.; Uh, Y.; et al. Emergence of NDM-1–producing Pseudomonas aeruginosa sequence type 773 clone: Shift of carbapenemase molecular epidemiology and spread of 16S rRNA methylase genes in Korea. Ann. Lab. Med. 2023, 43, 196–199. [Google Scholar] [CrossRef] [PubMed]
- Al-Alaq, F.T.; Mahmood, S.S.; AL-Khafaji, N.S.; Al-Dahmoshi, H.O.; Memariani, M. Investigation of blaIMP-1, blaVIM-1, blaOXA-48 and blaNDM-1 carbapenemase encoding genes among MBL-producing Pseudomonas aeruginosa. J. Appl. Nat. Sci. 2022, 14, 740–745. [Google Scholar] [CrossRef]
- Hong, J.S.; Song, W.; Park, M.-J.; Jeong, S.; Lee, N.; Jeong, S.H. Molecular characterization of the first emerged NDM-1-producing Pseudomonas aeruginosa isolates in South Korea. Microb. Drug Resist. 2021, 27, 1063–1070. [Google Scholar] [CrossRef]
- Schäfer, E.; Malecki, M.; Tellez-Castillo, C.J.; Pfennigwerth, N.; Marlinghaus, L.; Higgins, P.G.; Mattner, F.; Wendel, A.F. Molecular surveillance of carbapenemase-producing Pseudomonas aeruginosa at three medical centres in Cologne, Germany. Antimicrob. Resist. Infect. Control 2019, 8, 208. [Google Scholar] [CrossRef]
- Merradi, M.; Kassah-Laouar, A.; Ayachi, A.; Heleili, N.; Menasria, T.; Hocquet, D.; Cholley, P.; Sauget, M. Occurrence of VIM-4 metallo-β-lactamase-producing Pseudomonas aeruginosa in an Algerian hospital. J. Infect. Dev. Ctries. 2019, 13, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Forero-Hurtado, D.; Corredor-Rozo, Z.L.; Ruiz-Castellanos, J.S.; Márquez-Ortiz, R.A.; Abril, D.; Vanegas, N.; Lafaurie, G.I.; Chambrone, L.; Escobar-Pérez, J. Worldwide dissemination of blaKPC gene by novel mobilization platforms in Pseudomonas aeruginosa: A systematic review. Antibiotics 2023, 12, 658. [Google Scholar] [CrossRef] [PubMed]
- Rada, A.M.; De La Cadena, E.; Agudelo, C.A.; Pallares, C.; Restrepo, E.; Correa, A.; Villegas, M.V.; Capataz, C. Genetic diversity of multidrug-resistant Pseudomonas aeruginosa isolates carrying blaVIM–2 and blaKPC–2 genes that spread on different genetic environment in Colombia. Front. Microbiol. 2021, 12, 663020. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Zhang, Y.; Zhang, P.; Wang, J.; Yuan, Q.; Shi, W.; Zhang, S.; Feng, H.; Chen, Y.; Yu, M.; et al. Bloodstream infections caused by Klebsiella pneumoniae carbapenemase producing P. aeruginosa sequence type 463, associated with high mortality rates in China: A retrospective cohort study. Front. Cell. Infect. Microbiol. 2021, 11, 756782. [Google Scholar] [CrossRef]
- Shi, L.; Liang, Q.; Feng, J.; Zhan, Z.; Zhao, Y.; Yang, W.; Yang, H.; Chen, Y.; Huang, M.; Tong, Y.; et al. Coexistence of two novel resistance plasmids, blaKPC-2-carrying p14057A and tetA (A)-carrying p14057B, in Pseudomonas aeruginosa. Virulence 2018, 9, 306–311. [Google Scholar] [CrossRef]
- Hagemann, J.B.; Pfennigwerth, N.; Gatermann, S.G.; von Baum, H.; Essig, A. KPC-2 carbapenemase-producing Pseudomonas aeruginosa reaching Germany. J. Antimicrob. Chemother. 2018, 73, 1812–1814. [Google Scholar] [CrossRef]
- Vanegas, J.M.; Cienfuegos, A.V.; Ocampo, A.M.; López, L.; del Corral, H.; Roncancio, G.; Sierra, P.; Echeverri-Toro, L.; Ospina, S.; Maldonado, N.; et al. Similar frequencies of Pseudomonas aeruginosa isolates producing KPC and VIM carbapenemases in diverse genetic clones at tertiary-care hospitals in Medellín, Colombia. J. Clin. Microbiol. 2014, 52, 3978–3986. [Google Scholar] [CrossRef]
- Villegas, M.V.; Lolans, K.; Correa, A.; Kattan, J.N.; Lopez, J.A.; Quinn, J.P. First identification of Pseudomonas aeruginosa isolates producing a KPC-type carbapenem-hydrolyzing β-lactamase. Antimicrob. Agents Chemother. 2007, 51, 1553–1555. [Google Scholar] [CrossRef]
- Irum, S.; Naz, K.; Ullah, N.; Mustafa, Z.; Ali, A.; Arslan, M.; Khalid, K.; Andleeb, S. Antimicrobial resistance and genomic characterization of six new sequence types in multidrug-resistant Pseudomonas aeruginosa clinical isolates from Pakistan. Antibiotics 2021, 10, 1386. [Google Scholar] [CrossRef]
- Rather, P.; Munayyer, H.; Mann, P.; Hare, R.; Miller, G.; Shaw, K. Genetic analysis of bacterial acetyltransferases: Identification of amino acids determining the specificities of the aminoglycoside 6′-N-acetyltransferase Ib and IIa proteins. J. Bacteriol. 1992, 174, 3196–3203. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, N.; Wang, M.; Luo, M.; Peng, Y.; Li, Z.; Xu, J.; Ou, M.; Kan, B.; Li, X.; et al. The prevalence and distribution of aminoglycoside resistance genes. Biosaf. Health 2023, 5, 14–20. [Google Scholar] [CrossRef]
- Raheem, H.Q.; Hussein, E.F. Incidence of aminoglycoside resistance genes in Pseudomonas aeruginosa isolated from burns and wounds infections. Ann. Romanian Soc. Cell Biol. 2022, 26, 274–281. [Google Scholar]
- Ahmadian, L.; Norouzi Bazgir, Z.; Ahanjan, M.; Valadan, R.; Goli, H.R. Role of aminoglycoside-modifying enzymes (AMEs) in resistance to aminoglycosides among clinical isolates of Pseudomonas aeruginosa in the north of Iran. BioMed Res. Int. 2021, 2021, 7077344. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, B.; Rodulfo, H.; Carreno, N.; Guzman, M.; Salazar, E.; Donato, M.D. Aminoglycoside resistance genes in Pseudomonas aeruginosa isolates from Cumana, Venezuela. Rev. Inst. Med. Trop. Sao Paulo 2016, 58, 13. [Google Scholar] [CrossRef] [PubMed]
- Wachino, J.-I.; Arakawa, Y. Exogenously acquired 16S rRNA methyltransferases found in aminoglycoside-resistant pathogenic Gram-negative bacteria: An update. Drug Resist. Updat. 2012, 15, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Prasad, K.N.; Pathak, A.; Pati, B.K.; Singh, A.; Ovejero, C.M.; Ahmad, S.; Gonzalez-Zorn, B. RmtC and RmtF 16S rRNA methyltransferase in NDM-1–producing Pseudomonas aeruginosa. Emerg. Infect. Dis. 2015, 21, 2059. [Google Scholar] [CrossRef] [PubMed]
- Castanheira, M.; Fritsche, T.R.; Sader, H.l.S.; Jones, R.N. RmtD 16S RNA methylase in epidemiologically unrelated SPM-1-producing Pseudomonas aeruginosa isolates from Brazil. Antimicrob. Agents Chemother. 2008, 52, 1587. [Google Scholar] [CrossRef]
- Doi, Y.; Adams, J.M.; Yamane, K.; Paterson, D.L. Identification of 16S rRNA methylase-producing Acinetobacter baumannii clinical strains in North America. Antimicrob. Agents Chemother. 2007, 51, 4209–4210. [Google Scholar] [CrossRef]
- Samuelsen, Ø.; Toleman, M.; Hasseltvedt, V.; Fuursted, K.; Leegaard, T.; Walsh, T.R.; Sundsfjord, A.; Giske, C. Molecular characterization of VIM-producing Klebsiella pneumoniae from Scandinavia reveals genetic relatedness with international clonal complexes encoding transferable multidrug resistance. Clin. Microbiol. Infect. 2011, 17, 1811–1816. [Google Scholar] [CrossRef]
- Jiang, Y.; Yu, D.; Wei, Z.; Shen, P.; Zhou, Z.; Yu, Y. Complete nucleotide sequence of Klebsiella pneumoniae multidrug resistance plasmid pKP048, carrying blaKPC-2, blaDHA-1, qnrB4, and armA. Antimicrob. Agents Chemother. 2010, 54, 3967–3969. [Google Scholar] [CrossRef]
- Sekizuka, T.; Matsui, M.; Yamane, K.; Takeuchi, F.; Ohnishi, M.; Hishinuma, A.; Arakawa, Y.; Kuroda, M. Complete sequencing of the blaNDM-1-positive IncA/C plasmid from Escherichia coli ST38 isolate suggests a possible origin from plant pathogens. PLoS ONE 2011, 6, e25334. [Google Scholar] [CrossRef]
- Ho, P.L.; Lo, W.U.; Yeung, M.K.; Lin, C.H.; Chow, K.H.; Ang, I.; Tong, A.H.Y.; Bao, J.Y.-J.; Lok, S.; Lo, J.Y.C. Complete sequencing of pNDM-HK encoding NDM-1 carbapenemase from a multidrug-resistant Escherichia coli strain isolated in Hong Kong. PLoS ONE 2011, 6, e17989. [Google Scholar] [CrossRef]
- Carattoli, A.; Villa, L.; Poirel, L.; Bonnin, R.A.; Nordmann, P. Evolution of IncA/C blaCMY-2-carrying plasmids by acquisition of the blaNDM-1 carbapenemase gene. Antimicrob. Agents Chemother. 2012, 56, 783–786. [Google Scholar] [CrossRef]
- Mc Gann, P.; Hang, J.; Clifford, R.J.; Yang, Y.; Kwak, Y.I.; Kuschner, R.A.; Lesho, E.P.; Waterman, P.E. Complete sequence of a novel 178-kilobase plasmid carrying blaNDM-1 in a Providencia stuartii strain isolated in Afghanistan. Antimicrob. Agents Chemother. 2012, 56, 1673–1679. [Google Scholar] [CrossRef]
- Xu, C.; Wang, D.; Zhang, X.; Liu, H.; Zhu, G.; Wang, T.; Cheng, Z.; Wu, W.; Bai, F.; Jin, Y. Mechanisms for rapid evolution of carbapenem resistance in a clinical isolate of Pseudomonas aeruginosa. Front. Microbiol. 2020, 11, 1390. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Outbreak of Extensively Drug-Resistant Pseudomonas aeruginosa Associated with Artificial Tears; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2023. [Google Scholar]
- Recio, R.; Sánchez-Diener, I.; Viedma, E.; Meléndez-Carmona, M.Á.; Villa, J.; Orellana, M.Á.; Mancheño, M.; Juan, C.; Zamorano, L.; Lora-Tamayo, J.; et al. Pathogenic characteristics of Pseudomonas aeruginosa bacteraemia isolates in a high-endemicity setting for ST175 and ST235 high-risk clones. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 671–678. [Google Scholar] [CrossRef]
- Papagiannitsis, C.C.; Verra, A.; Galani, V.; Xitsas, S.; Bitar, I.; Hrabak, J.; Petinaki, E. Unravelling the features of success of VIM-producing ST111 and ST235 Pseudomonas aeruginosa in a Greek hospital. Microorganisms 2020, 8, 1884. [Google Scholar] [CrossRef]
- Loconsole, D.; Accogli, M.; Monaco, M.; Del Grosso, M.; De Robertis, A.L.; Morea, A.; Capozzi, L.; Del Sambro, L.; Simone, A.; De Letteriis, V.; et al. First detection of autochthonous extensively drug-resistant NDM-1 Pseudomonas aeruginosa ST235 from a patient with bloodstream infection in Italy, October 2019. Antimicrob. Resist. Infect. Control 2020, 9, 73. [Google Scholar] [CrossRef] [PubMed]
- Abril, D.; Marquez-Ortiz, R.A.; Castro-Cardozo, B.; Moncayo-Ortiz, J.I.; Olarte Escobar, N.M.; Corredor Rozo, Z.L.; Reyes, N.; Tovar, C.; Sánchez, H.F.; Castellanos, J.; et al. Genome plasticity favours double chromosomal Tn4401b-blaKPC-2 transposon insertion in the Pseudomonas aeruginosa ST235 clone. BMC Microbiol. 2019, 19, 45. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Bae, I.K.; Jeong, S.H.; Kim, S.H.; Song, J.H.; Choi, J.Y.; Yoon, S.S.; Thamlikitkul, V.; Hsueh, P.-R.; Yasin, R.M.; et al. Dissemination of metallo-β-lactamase-producing Pseudomonas aeruginosa of sequence type 235 in Asian countries. J. Antimicrob. Chemother. 2013, 68, 2820–2824. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.H.; Kwon, K.C.; Sung, J.Y.; Koo, S.H. Prevalence and genetic analysis of multidrug-resistant Pseudomonas aeruginosa ST235 isolated from a hospital in Korea, 2008–2012. Ann. Clin. Lab. Sci. 2013, 43, 414–419. [Google Scholar]
- Dos Santos, P.A.S.; Rodrigues, Y.C.; Marcon, D.J.; Lobato, A.R.F.; Cazuza, T.B.; Gouveia, M.I.M.; Silva, M.J.A.; Souza, A.B.; Lima, L.N.G.C.; Quaresma, A.J.P.G.; et al. Endemic high-risk clone ST277 is related to the spread of SPM-1-producing Pseudomonas aeruginosa during the COVID-19 pandemic period in Northern Brazil. Microorganisms 2023, 11, 2069. [Google Scholar] [CrossRef] [PubMed]
- Silveira, M.C.; Rocha-de-Souza, C.M.; Albano, R.M.; de Oliveira Santos, I.C.; Carvalho-Assef, A.P.D.A. Exploring the success of Brazilian endemic clone Pseudomonas aeruginosa ST277 and its association with the CRISPR-Cas system type I-C. BMC Genom. 2020, 21, 255. [Google Scholar] [CrossRef] [PubMed]
- Martins, W.M.; Narciso, A.C.; Cayo, R.; Santos, S.V.; Fehlberg, L.C.; Ramos, P.L.; da Cruz, J.B.; Gales, A.C. SPM-1-producing Pseudomonas aeruginosa ST277 clone recovered from microbiota of migratory birds. Diagn. Microbiol. Infect. Dis. 2018, 90, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Turano, H.; Gomes, F.; Medeiros, M.; Oliveira, S.; Fontes, L.C.; Sato, M.I.; Lincopan, N. Presence of high-risk clones of OXA-23-producing Acinetobacter baumannii (ST79) and SPM-1-producing Pseudomonas aeruginosa (ST277) in environmental water samples in Brazil. Diagn. Microbiol. Infect. Dis. 2016, 86, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, K.L.; Meunier, D.; Findlay, J.; Mustafa, N.; Parsons, H.; Pike, R.; Wright, L.; Woodford, N. SPM-1 metallo-β-lactamase-producing Pseudomonas aeruginosa ST277 in the UK. J. Med. Microbiol. 2016, 65, 696–697. [Google Scholar] [CrossRef]
- Mano, Y.; Saga, T.; Ishii, Y.; Yoshizumi, A.; Bonomo, R.A.; Yamaguchi, K.; Tateda, K. Molecular analysis of the integrons of metallo-β-lactamase-producing Pseudomonas aeruginosa isolates collected by nationwide surveillance programs across Japan. BMC Microbiol. 2015, 15, 41. [Google Scholar] [CrossRef] [PubMed]
- Cejas, D.; Elena, A.; González-Espinosa, F.E.; Pallecchi, L.; Vay, C.; Rossolini, G.M.; Gutkind, G.; Di Pilato, V.; Radice, M. Characterisation of blaKPC-2–harbouring plasmids recovered from Pseudomonas aeruginosa ST654 and ST235 high-risk clones. J. Glob. Antimicrob. Resist. 2022, 29, 310–312. [Google Scholar] [CrossRef] [PubMed]
- Galimand, M.; Sabtcheva, S.; Courvalin, P.; Lambert, T. Worldwide disseminated armA aminoglycoside resistance methylase gene is borne by composite transposon Tn 1548. Antimicrob. Agents Chemother. 2005, 49, 2949–2953. [Google Scholar] [CrossRef]
- Gonzalez-Zorn, B.; Catalan, A.; Escudero, J.A.; Domínguez, L.; Teshager, T.; Porrero, C.; Moreno, M.A. Genetic basis for dissemination of armA. J. Antimicrob. Chemother. 2005, 56, 583–585. [Google Scholar] [CrossRef]
- Du, X.-D.; Li, D.-X.; Hu, G.-Z.; Wang, Y.; Shang, Y.-H.; Wu, C.-M.; Liu, H.-B.; Li, X.-S. Tn 1548-associated armA is co-located with qnrB2, aac (6′)-Ib-cr and blaCTX-M-3 on an IncFII plasmid in a Salmonella enterica subsp. enterica serovar Paratyphi B strain isolated from chickens in China. J. Antimicrob. Chemother. 2012, 67, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Yamane, K.; Wachino, J.-I.; Suzuki, S.; Kimura, K.; Shibata, N.; Kato, H.; Shibayama, K.; Konda, T.; Arakawa, Y. New plasmid-mediated fluoroquinolone efflux pump, QepA, found in an Escherichia coli clinical isolate. Antimicrob. Agents Chemother. 2007, 51, 3354–3360. [Google Scholar] [CrossRef] [PubMed]
- Périchon, B.; Bogaerts, P.; Lambert, T.; Frangeul, L.; Courvalin, P.; Galimand, M. Sequence of conjugative plasmid pIP1206 mediating resistance to aminoglycosides by 16S rRNA methylation and to hydrophilic fluoroquinolones by efflux. Antimicrob. Agents Chemother. 2008, 52, 2581–2592. [Google Scholar] [CrossRef]
- Poirel, L.; Bonnin, R.A.; Nordmann, P. Analysis of the resistome of a multidrug-resistant NDM-1-producing Escherichia coli strain by high-throughput genome sequencing. Antimicrob. Agents Chemother. 2011, 55, 4224–4229. [Google Scholar] [CrossRef]
- Wang, L.; Hu, C.; Shao, L. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2017, 12, 1227–1249. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, M.; Anju, C.; Biswas, L.; Kumar, V.A.; Mohan, C.G.; Biswas, R. Antibiotic resistance in Pseudomonas aeruginosa and alternative therapeutic options. Int. J. Med. Microbiol. 2016, 306, 48–58. [Google Scholar] [CrossRef]
- Salomoni, R.; Léo, P.; Montemor, A.; Rinaldi, B.; Rodrigues, M. Antibacterial effect of silver nanoparticles in Pseudomonas aeruginosa. Nanotechnol. Sci. Appl. 2017, 10, 115–121. [Google Scholar] [CrossRef]
- Brown, A.N.; Smith, K.; Samuels, T.A.; Lu, J.; Obare, S.O.; Scott, M.E. Nanoparticles functionalized with ampicillin destroy multiple-antibiotic-resistant isolates of Pseudomonas aeruginosa and Enterobacter aerogenes and methicillin-resistant Staphylococcus aureus. Appl. Environ. Microbiol. 2012, 78, 2768–2774. [Google Scholar] [CrossRef]
- de Lacerda Coriolano, D.; de Souza, J.B.; Bueno, E.V.; Medeiros, S.M.d.F.R.d.S.; Cavalcanti, I.D.L.; Cavalcanti, I.M.F. Antibacterial and antibiofilm potential of silver nanoparticles against antibiotic-sensitive and multidrug-resistant Pseudomonas aeruginosa strains. Braz. J. Microbiol. 2021, 52, 267–278. [Google Scholar] [CrossRef]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.; Giske, C.; Harbarth, S.; Hindler, J.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Pierce, V.M.; Simner, P.J.; Lonsway, D.R.; Roe-Carpenter, D.E.; Johnson, J.K.; Brasso, W.B.; Bobenchik, A.M.; Lockett, Z.C.; Charnot-Katsikas, A.; Ferraro, M.J.; et al. Modified carbapenem inactivation method for phenotypic detection of carbapenemase production among Enterobacteriaceae. J. Clin. Microbiol. 2017, 55, 2321–2333. [Google Scholar] [CrossRef] [PubMed]
- Gondal, A.J.; Choudhry, N.; Bukhari, H.; Rizvi, Z.; Yasmin, N. Characterization of genomic diversity among carbapenem-resistant Escherichia coli clinical isolates and antibacterial efficacy of silver nanoparticles from Pakistan. Microorganisms 2022, 10, 2283. [Google Scholar] [CrossRef] [PubMed]
- Spilker, T.; Coenye, T.; Vandamme, P.; LiPuma, J.J. PCR-based assay for differentiation of Pseudomonas aeruginosa from other Pseudomonas species recovered from cystic fibrosis patients. J. Clin. Microbiol. 2004, 42, 2074–2079. [Google Scholar] [CrossRef] [PubMed]
- Curran, B.; Jonas, D.; Grundmann, H.; Pitt, T.; Dowson, C.G. Development of a multilocus sequence typing scheme for the opportunistic pathogen Pseudomonas aeruginosa. J. Clin. Microbiol. 2004, 42, 5644–5649. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Carloni, E.; Andreoni, F.; Omiccioli, E.; Villa, L.; Magnani, M.; Carattoli, A. Comparative analysis of the standard PCR-based replicon typing (PBRT) with the commercial PBRT-KIT. Plasmid 2017, 90, 10–14. [Google Scholar] [CrossRef]
- Alvarado, A.; Garcillán-Barcia, M.P.; de la Cruz, F. A degenerate primer MOB typing (DPMT) method to classify gamma-proteobacterial plasmids in clinical and environmental settings. PLoS ONE 2012, 7, e40438. [Google Scholar] [CrossRef]
- Wan, G.; Ruan, L.; Yin, Y.; Yang, T.; Ge, M.; Cheng, X. Effects of silver nanoparticles in combination with antibiotics on the resistant bacteria Acinetobacter baumannii. Int. J. Nanomed. 2016, 11, 3789–3800. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Genomic Characterization, n (%) | ||
|---|---|---|
| Carbapenemase-Producing Strains (n = 201) | Non-Carbapenemase-Producing Strains (n = 48) | |
| Carbapenemases (n = 179) | ||
| blaOXA-48 | 67 (37.43) | - |
| blaNDM-1 | 59 (32.96) | - |
| blaIMP | 13 (7.26) | - |
| blaVIM | 9 (5.03) | - |
| blaKPC-2 | 2 (1.12) | - |
| blaNDM-1/blaOXA48 | 25 (13.97) | - |
| blaOXA-48/blaVIM | 3 (1.68) | - |
| blaVIM/blaIMP | 1 (0.56) | - |
| ESBLs (n = 249) | ||
| blaSHV | 63 (31.34) | 14 (29.17) |
| blaCTX-M | 49 (24.38) | 17 (35.42) |
| blaTEM | 27 (13.43) | 8 (16.67) |
| blaSHV/blaCTX-M | 29 (14.43) | 4 (8.33) |
| blaCTX-M/blaTEM | 19 (9.45) | 3 (6.25) |
| blaSHV/blaTEM | 8 (3.98) | 1 (2.08) |
| blaSHV/blaCTX-M/blaTEM | 6 (2.99) | 1 (2.08) |
| Aminoglycoside resistance genes (n = 109) | ||
| aac(6′)-lb | 11 (12.79) | 3 (13.04) |
| aac(3)-lla | 8 (9.30) | 5 (21.74) |
| rmtB | 17 (19.77) | 6 (26.09) |
| armA | 13 (15.12) | 1 (4.35) |
| rmtC | 9 (10.47) | 3 (13.04) |
| rmtD | 5 (5.81) | - |
| rmtF | 4 (4.65) | 3 (13.04) |
| rmtB/aac(3)-lla | 9 (10.47) | - |
| rmtB/aac(6′)-lla | 7 (8.14) | 1 (4.35) |
| rmtB/armA | 3 (3.49) | 1 (4.35) |
| Strain ID | Carbapenemases | AME/RMT Genes | ESBLs | Sequence Typing | Plasmid Typing | |
|---|---|---|---|---|---|---|
| PBRT | MOB | |||||
| PA-01 | blaOXA-48 | rmtD | blaSHV | 277 | IncFII, IncFI, IncA/C, IncL/M | F12, P131, H121 |
| PA-03 | blaNDM-1 | aac(6′)-lb | blaCTX-M | 313 | IncFII, IncA/C, IncN | F11, F12, H121 |
| PA-04 | blaNDM-1 | armA | blaCTX-M | 234 | IncFII, IncA/C, IncN, IncL/M | F12, P131, H121 |
| PA-08 | blaOXA-48 | rmtB/aac(3)-lla | blaSHV/blaCTX-M | 235 | IncFII, IncA/C, IncL, IncN | F11, F12, H121 |
| PA-09 | blaNDM-1 | armA | blaCTX-M | 234 | IncFII, IncA/C, IncN | F11, F12, H121 |
| PA-10 | blaNDM-1 | rmtB | blaSHV/blaTEM | 235 | IncFII, IncA/C, IncL/M | F12, P131, H121 |
| PA-11 | blaNDM-1 | aac(6′)-lb | blaSHV/blaCTX-M | 357 | IncFII, IncA/C, IncN, IncFIIK | F11, F12, H121 |
| PA-14 | blaNDM-1 | aac(6′)-lb | blaSHV/blaCTX-M | 235 | IncFII, IncA/C, IncN | F11, F12, H121 |
| PA-15 | blaNDM-1/blaOXA-48 | rmtB | blaCTX-M/blaTEM | 235 | IncFII, IncA/C, IncL/M, IncN | F11, F12, H121 |
| PA-23 | blaOXA-48 | rmtB/aac(3)-lla | blaSHV/blaCTX-M | 235 | IncFII, IncA/C, IncL/M | F12, P131, H121 |
| PA-25 | blaOXA-48 | rmtB | blaSHV | 277 | IncFII, IncA/C, IncL/M | F12, P131, H121 |
| PA-26 | blaNDM-1 | rmtB/aac(3)-lla | blaSHV/blaCTX-M | 313 | IncFII, IncA/C, IncN | F11, H121 |
| PA-32 | blaNDM-1/blaOXA-48 | armA | blaSHV | 313 | IncFII, IncA/C, IncL/M, IncN | F11, P131, H121 |
| PA-33 | blaNDM-1 | rmtB | blaCTX-M | 277 | IncFII, IncA/C, IncN | F11, H121 |
| PA-34 | blaOXA-48 | rmtB | blaTEM | 235 | IncFII, IncA/C, IncL/M | F12, P131, H121 |
| PA-35 | blaNDM-1 | rmtD | blaSHV | 235 | IncFII, IncA/C, IncN, IncL/M | F12, P131, H121 |
| PA-36 | blaVIM | rmtB | blaCTX-M | 235 | IncFII, IncA/C, IncN | F11, F12, H121 |
| PA-39 | blaNDM-1 | rmtD | blaSHV | 170 | IncFII, IncA/C, IncN | F11, F12, H121 |
| PA-44 | blaOXA-48 | rmtB/armA | blaTEM | 170 | IncFII, IncA/C, IncL/M | F12, P131, H121 |
| PA-45 | blaNDM-1 | rmtB | blaSHV | 235 | IncFII, IncA/C, IncFIIK | F12, H121 |
| PA-49 | blaOXA-48 | aac(6′)-lb | blaSHV | 235 | IncFI, IncA/C, IncL/M | F12, P131, H121 |
| PA-51 | blaOXA-48 | rmtF | blaSHV | 1455 | IncFII, IncA/C, IncN | F11, F12, H121 |
| PA-59 | blaOXA-48 | aac(6′)-lb | blaCTX-M | 170 | IncFII, IncA/C, IncL/M | F12, H121, P131 |
| PA-62 | blaIMP | rmtB | blaTEM | 235 | IncFI, IncA/C, IncN, IncX | F11, H121, P3 |
| PA-64 | blaOXA-48 | rmtB/aac(3)-lla | blaSHV/blaCTX-M/blaTEM | 170 | IncFI, IncA/C, IncL/M | F12, H121, P131 |
| PA-67 | blaVIM | rmtB | blaSHV | 234 | IncFII, IncA/C, IncN | F12, H121 |
| PA-68 | blaNDM-1 | armA | blaCTX-M/blaTEM | 1455 | IncFI, IncA/C, IncN | F11, F12, H121 |
| PA-71 | blaKPC-2 | armA | blaSHV | 235 | IncFI, IncA/C, IncL/M, IncP4 | F12, P131 |
| PA-72 | blaNDM-1 | rmtB | blaCTX-M | 664 | IncFII, IncA/C, IncX | F12, H121, P3 |
| PA-73 | blaNDM-1/blaOXA-48 | rmtB | blaSHV | 235 | IncFII, IncA/C | F12, H121 |
| PA-77 | blaOXA-48 | rmtB/aac(3)-lla | blaSHV/blaCTX-M | 235 | IncFII, IncA/C, IncL/M, IncX | F12, P131, H121, P3 |
| PA-78 | blaNDM-1/blaOXA-48 | aac(3)-lla | blaCTX-M/blaTEM | 170 | IncFI, IncA/C, IncN, IncFIIK | F11, F12, H121 |
| PA-80 | blaOXA-48 | rmtB/aac(6′)-lla | blaSHV/blaCTX-M | 235 | IncFI, IncA/C, IncN, IncL/M | F11, F12, P131, H121 |
| PA-81 | blaVIM | rmtB/aac(6′)-lla | blaSHV | 277 | IncFII, IncA/C, IncN, | F11, F12, H121 |
| PA-82 | blaOXA-48 | rmtF | blaCTX-M | 1963 | IncFII, IncA/C, IncL/M, IncN | F11, F12, H121, P131 |
| PA-85 | blaNDM-1 | armA | blaSHV | 277 | IncFI, IncA/C, IncN, IncX | F11, F12, H121, P3 |
| PA-93 | blaNDM-1 | aac(3)-lla | blaTEM | 1963 | IncFII, IncA/C, IncN | F11, F12, H121 |
| PA-95 | blaNDM-1/blaOXA-48 | armA | blaCTX-M | 348 | IncFII, IncA/C, IncL/M, IncHI1 | F12, P131, H121, H11 |
| PA-96 | blaNDM-1/blaOXA-48 | aac(6′)-lb | blaSHV/blaCTX-M | 235 | IncFII, IncA/C, IncL/M | F12, P131, H121 |
| PA-99 | blaIMP | rmtB | blaTEM | 234 | IncFII, IncFIIK, IncA/C, IncN | F11, F12, H121 |
| PA-104 | blaNDM-1 | rmtB | blaSHV | 1455 | IncN, IncFII, IncFIIK, IncA/C | F11, F12, H121 |
| PA-105 | blaNDM-1 | rmtD | blaSHV/blaCTX-M | 170 | IncN, IncFII, IncA/C | F11, F12, H121 |
| PA-108 | blaNDM-1 | aac(3)-lla | blaSHV | 277 | IncFII, IncN, IncA/C, IncFIIK | F11, F12, H121 |
| PA-113 | blaNDM-1 | aac(3)-lla | blaSHV/blaCTX-M | 277 | IncFII, IncN, IncA/C | F11, F12, H121 |
| PA-116 | blaVIM | rmtB | blaCTX-M | 277 | IncN, IncFII, IncA/C | F12, F11, H121 |
| PA-120 | blaNDM-1 | rmtB | blaSHV | 381 | IncFII, IncA/C, IncX | F12, H121, P3 |
| PA-122 | blaOXA-48 | rmtB/aac(3)-lla | blaSHV | 1963 | IncFII, IncA/C, IncL/M | F12, P131, H121 |
| PA-127 | blaVIM | armA | blaSHV | 170 | IncFII, IncA/C | F12, H121 |
| PA-131 | blaNDM-1/blaOXA-48 | rmtB | blaSHV/blaCTX-M | 170 | IncFII, IncA/C, IncL/M, IncN | F11, F12, P131, H121 |
| PA-138 | blaIMP | rmtB/aac(3)-lla | blaSHV/blaCTX-M | 235 | IncFII, IncA/C, IncL/M | F12, P131, H121 |
| PA-139 | blaNDM-1 | armA | blaSHV | 235 | IncFII, IncA/C, IncFIIK | F12, H121 |
| PA-140 | blaNDM-1 | armA | blaTEM | 235 | IncFII, IncA/C | F12, H121 |
| PA-141 | blaOXA-48 | rmtB/aac(6′)-lla | blaCTX-M/blaTEM | 1455 | IncFII, IncA/C | F12, H121 |
| PA-152 | blaNDM-1 | rmtB/aac(3)-lla | blaCTX-M | 235 | IncN, IncFII, IncA/C, IncL/M | F11, F12, P131, H121 |
| PA-158 | blaNDM-1 | rmtB/aac(6′)-lla | blaSHV/blaCTX-M | 235 | IncN, IncFII, IncA/C, IncX | F11, F12, H121, P3 |
| PA-162 | blaNDM-1/blaOXA-48 | aac(6′)-lb | blaSHV/blaTEM | 175 | IncFII, IncA/C, IncL/M, IncX | F12, P131, H121, P3 |
| PA-163 | blaVIM | aac(6′)-lb | blaTEM | 235 | IncFII, IncA/C | F12, H121 |
| PA-164 | blaOXA-48 | aac(3)-lla | blaSHV | 175 | IncFII, IncA/C, IncL/M | F12, P131, H121 |
| PA-165 | blaOXA-48 | aac(6′)-lb | blaSHV/blaTEM | 175 | IncFII, IncA/C, IncN | F11, F12, H121 |
| PA-171 | blaOXA-48 | aac(6′)-lb | blaSHV | 234 | IncFII, IncA/C, IncL/M | F12, P131, H121 |
| PA-175 | blaNDM-1 | rmtB/aac(6′)-lla | blaSHV/blaCTX-M/blaTEM | 234 | IncFII, IncA/C, IncN | F11, F12, H121 |
| PA-178 | blaVIM | aac(3)-lla | blaSHV/blaCTX-M | 234 | IncFII, IncA/C, IncFIIK | F12, H121 |
| PA-183 | blaNDM-1 | rmtB/armA | blaSHV/blaCTX-M/blaTEM | 277 | IncFII, IncFIIK, IncA/C | F12, H121 |
| PA-184 | blaNDM-1 | rmtC | blaSHV/blaCTX-M | 277 | IncFII, IncA/C, IncN | F11, F12, H121 |
| PA-185 | blaVIM | rmtC | blaSHV/blaTEM | 277 | IncFII, IncA/C, IncN, IncR | F12, H121 |
| PA-189 | blaNDM-1 | rmtD | blaSHV/blaCTX-M | 235 | IncFII, IncA/C, IncN | F11, F12, H121 |
| PA-194 | blaIMP | rmtC | blaSHV/blaTEM | 381 | IncFII, IncA/C, IncL/M | F12, P131, H121 |
| PA-198 | blaVIM | rmtB/aac(6′)-lla | blaSHV/blaCTX-M/blaTEM | 277 | IncFII, IncA/C, IncFIIK, IncX | F12, H121, P3 |
| PA-202 | blaNDM-1 | rmtC | blaSHV/blaCTX-M | 235 | IncFII, IncA/C, IncN, IncR | F12, H121 |
| PA-209 | blaNDM-1/blaOXA-48 | rmtB/aac(6′)-lla | blaSHV/blaTEM | 235 | IncFII, IncA/C, IncL/M, IncN | F11, P131, H121 |
| PA-211 | blaNDM-1 | rmtB/armA | blaSHV/blaCTX-M | 235 | IncFII, IncA/C, IncN, IncFIIK | F11, F12, H121 |
| PA-218 | blaOXA-48 | rmtC | blaSHV/blaCTX-M | 234 | IncFII, IncA/C, IncL/M | F12, P131, H121 |
| PA-227 | blaOXA-48 | rmtB | blaCTX-M | 175 | IncFII, IncA/C, IncL/M | F12, P131, H121 |
| PA-231 | blaOXA-48 | rmtB/aac(3)-lla | blaSHV/blaTEM | 381 | IncFI, IncA/C, IncL/M, IncX | F12, P131, H121 |
| PA-243 | blaNDM-1/blaOXA-48 | aac(3)-lla | blaSHV/blaCTX-M | 381 | IncFII, IncA/C, IncL/M, IncN | F11, F12, P131, H121 |
| PA-244 | blaNDM-1/blaOXA-48 | rmtB | blaSHV/blaCTX-M | 381 | IncFII, IncA/C, IncL/M, IncN | F11, F12, P131, H121 |
| PA-249 | blaIMP | aac(6′)-lb | blaCTX-M | 207 | IncFII, IncA/C, IncN | F11, F12, H121 |
| For MEM | For AK | |
|---|---|---|
| ∑FIC Interpretation | Strains ID | Strains ID |
| Synergism | PA-01, PA-04, PA-09, PA-10, PA-14, PA-15, PA-23, PA-25, PA-26, PA-32, PA-33, PA-34, PA-39, PA-44, PA-45, PA-51, PA-59, PA-62, PA-64, PA-71, PA-72, PA-77, PA-78, PA-81, PA-82, PA-93, PA-95, PA-96, PA-104, PA-105, PA-108, PA-116, PA-120, PA-122, PA-131, PA-139, PA-138, PA-140, PA-141, PA-152, PA-158, PA-163, PA-165, PA-171, PA-175, PA-178, PA-183, PA-184, PA-185, PA-189, PA-202, PA-209, PA-211, PA-218, PA-227, PA-231, PA-243, PA-244, PA-249 | PA-01, PA-04, PA-09, PA-10, PA-14, PA-15, PA-23, PA-25, PA-26, PA-32, PA-34, PA-33, PA-35, PA-39, PA-45, PA-51, PA-59, PA-62, PA-64, PA-67, PA-71, PA-72, PA-73, PA-77, PA-78, PA-81, PA-82, PA-93, PA-95, PA-96, PA-99, PA-104, PA-105, PA-108, PA-113, PA-116, PA-120, PA-122, PA-131, PA-138, PA-139, PA-140, PA-141, PA-152, PA-158, PA-163, PA-165, PA-171, PA-175, PA-178, PA-183, PA-184, PA-185, PA-189, PA-194PA-198, PA-202, PA-209, PA-211, PA-218, PA-243, PA-244, PA-227, PA-231, PA-249 |
| Indifferent | PA-03, PA-11, PA-35, PA-49, PA-68, PA-73, PA-99, PA-162, PA-164 | PA-03, PA-11, PA-49, PA-68, PA-162, PA-164 |
| Antagonism | PA-36, PA-67, PA-85, PA-113, PA-127, PA-194, PA-198 | PA-36, PA-44, PA-85, PA-127 |
| Primer Sequences (5′–3′) | Tm (°C) | Product Size (bp) | |
|---|---|---|---|
| Molecular Identification of P. aeruginosa [154] | |||
| PA-SS-F | F: ggg gga tct tcg gac ctc a R: tcc tta gag tgc cca ccc g | 56 | 956 |
| Carbapenemase Resistance Genes [153] | |||
| blaKPC-2 | F: gct aca cct agc tcc acc ttc R: aca gtg gtt ggt aat cca tgc | 55 | 989 |
| blaNDM-1 | F: ggg cag tcg ctt cca acg gt R: gta gtg ctc agt gtc ggc at | 53 | 476 |
| blaVIM | F: gat ggt gtt tgg tcg cat a R: cga atg cgc agc acc ag | 52 | 390 |
| blaOXA-48 | F: gcg tgg tta agg atg aac ac R: cat caa gtt caa ccc aac cg | 52 | 438 |
| blaIMP | F: gga ata gag tgg ctt aay tct c R: ggt tta aya aaa caa cca cc | 52 | 232 |
| ESBLs [153] | |||
| blaSHV | F: ctt tat cgg ccc tca ctc aa R: agg tgc tca tca tgg gaa ag | 55 | 237 |
| blaTEM | F: cgc cgc ata cac tat tct cag aat ga R: acg ctc acc ggc tcc aga ttt at | 55 | 445 |
| blaCTX-M | F: atg tgc agy acc agt aar gtk atg gc R: tgg gtr aar tar gts acc aga ayc agc gg | 55 | 593 |
| Aminoglycoside Resistance Genes [73] | |||
| aac(6′)-lb | F: ttg cga tgc tct atg agt ggc ta R: ctc gaa tgc ctg gcg tgt tt | 59 | 482 |
| aac(3)-lla | F: ggc aat aac gga ggc gct tca aaa R: ttc cag gca tcg gca tct cat acg | 60 | 563 |
| rmtB | F: gct ttc tgc ggg cga tgt aa R: atg caa tgc cgc gct cgt at | 59 | 173 |
| rmtC | F: gct gcc ctt tgt att gtc R: aga tgt tgg gtt aag tcc c | 55 | 711 |
| armA | F: att ctg cct atc cta att gg R: acc tat act tta tcg tcg tc | 53 | 315 |
| rmtD | F: cgg cac gcg att ggg aag c R: cgg aaa cga tgc gac gat | 58 | 401 |
| rmtF | F: gcg ata cag aaa acc gaa gg R: acc agt cgg cat agt gct tt | 60 | 589 |
| MLST PCR amplification [155] | |||
| acsA | F: acc tgg tgt acg cct cgc tga c R: gac ata gat gcc ctg ccc ctt gat | 55 | 842 |
| aroE | F: tggggctatgactggaaacc R: taa ccc ggt ttt gtg att cct aca | 55 | 825 |
| guaA | F: cgg cct cga cgt gtg gat ga R: gaa cgc ctg gct ggt ctt gtg gta | 55 | 940 |
| mutL | F: cca gat cgc cgc cgg tga ggt g R: cag ggt gcc ata gag gaa gtc | 55 | 940 |
| muoD | F: acc gcc acc cgt act g R: tct cgc cca tct tga cca | 55 | 1042 |
| ppsA | F: ggt cgc tcg gtc aag gta gtg g R: ggg ttc tct tct tcc ggc tcg tag | 55 | 989 |
| trpE | F: gcg gcc cag ggt cgt gag R: ccc ggc gct tgt tga tgg tt | 55 | 811 |
| MLST PCR Sequencing [155] | |||
| acsA | F: gcc aca cct aca tcg tct at R: gtg gac aac ctc ggc aac ct | 390 | |
| aroE | F: atg tca ccg tgc cgt tca ag R: tga agg cag tcg gtt cct tg | 495 | |
| guaA | F: agg tcg gtt cct cca agg tc R: tca agt cgc acc aca acg tc | 372 | |
| mutL | F: aga aga ccg agt tcg acc at R: atg act tcc tct atg gca cc | 441 | |
| muoD | F: acg gcg aga acg agg act ac R: ttc acc ttc acc gac cgc ca | 366 | |
| ppsA | F: ggt gac gac ggc aag ctg ta R: tcc tgt gcc gaa ggc gat ac | 369 | |
| trpE | F: ttc aac ttc ggc gac ttc ca R: ggt gtc cat gtt gcc gtt cc | 441 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gondal, A.J.; Choudhry, N.; Niaz, A.; Yasmin, N. Molecular Analysis of Carbapenem and Aminoglycoside Resistance Genes in Carbapenem-Resistant Pseudomonas aeruginosa Clinical Strains: A Challenge for Tertiary Care Hospitals. Antibiotics 2024, 13, 191. https://doi.org/10.3390/antibiotics13020191
Gondal AJ, Choudhry N, Niaz A, Yasmin N. Molecular Analysis of Carbapenem and Aminoglycoside Resistance Genes in Carbapenem-Resistant Pseudomonas aeruginosa Clinical Strains: A Challenge for Tertiary Care Hospitals. Antibiotics. 2024; 13(2):191. https://doi.org/10.3390/antibiotics13020191
Chicago/Turabian StyleGondal, Aamir Jamal, Nakhshab Choudhry, Ammara Niaz, and Nighat Yasmin. 2024. "Molecular Analysis of Carbapenem and Aminoglycoside Resistance Genes in Carbapenem-Resistant Pseudomonas aeruginosa Clinical Strains: A Challenge for Tertiary Care Hospitals" Antibiotics 13, no. 2: 191. https://doi.org/10.3390/antibiotics13020191