

Screening for Antimicrobial Resistance and Genes of Exotoxins in Pseudomonas aeruginosa Isolates from Infected Dogs and Cats in Poland

Abstract
:1. Introduction
2. Materials and Methods
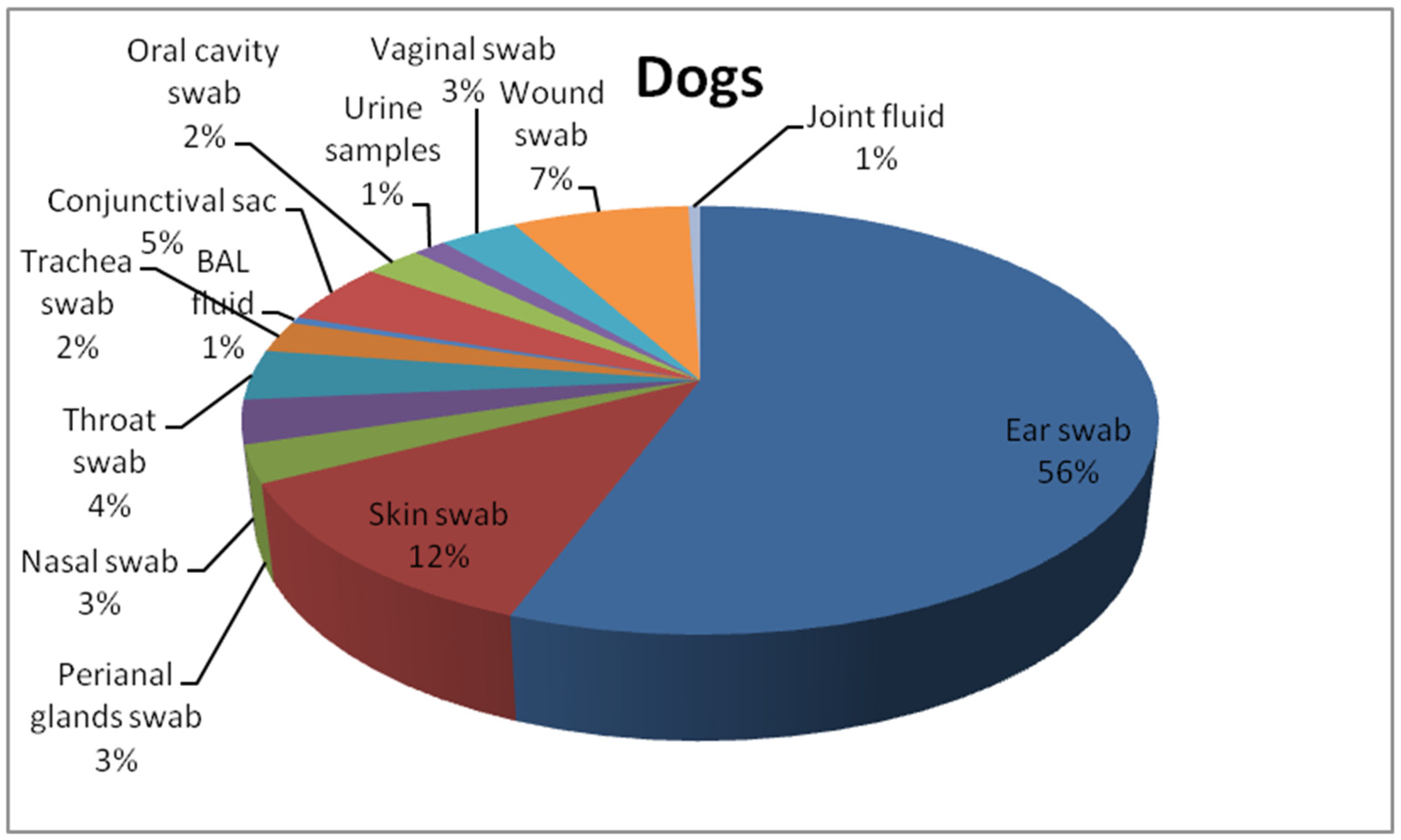
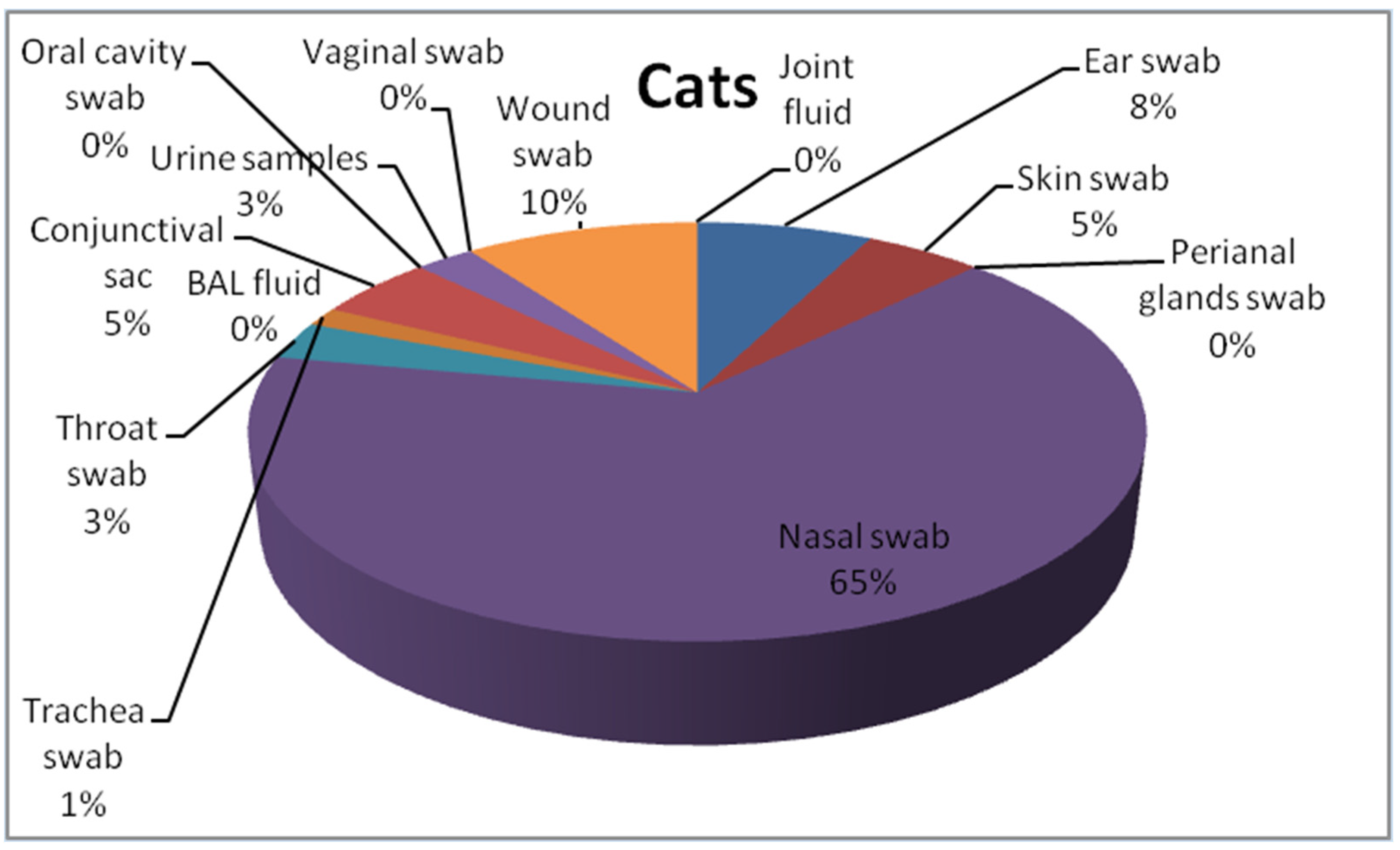
2.1. Isolates
2.2. Antibiotic Resistance
2.3. Detection of Virulence Genes
2.4. Statistical Analysis
3. Results
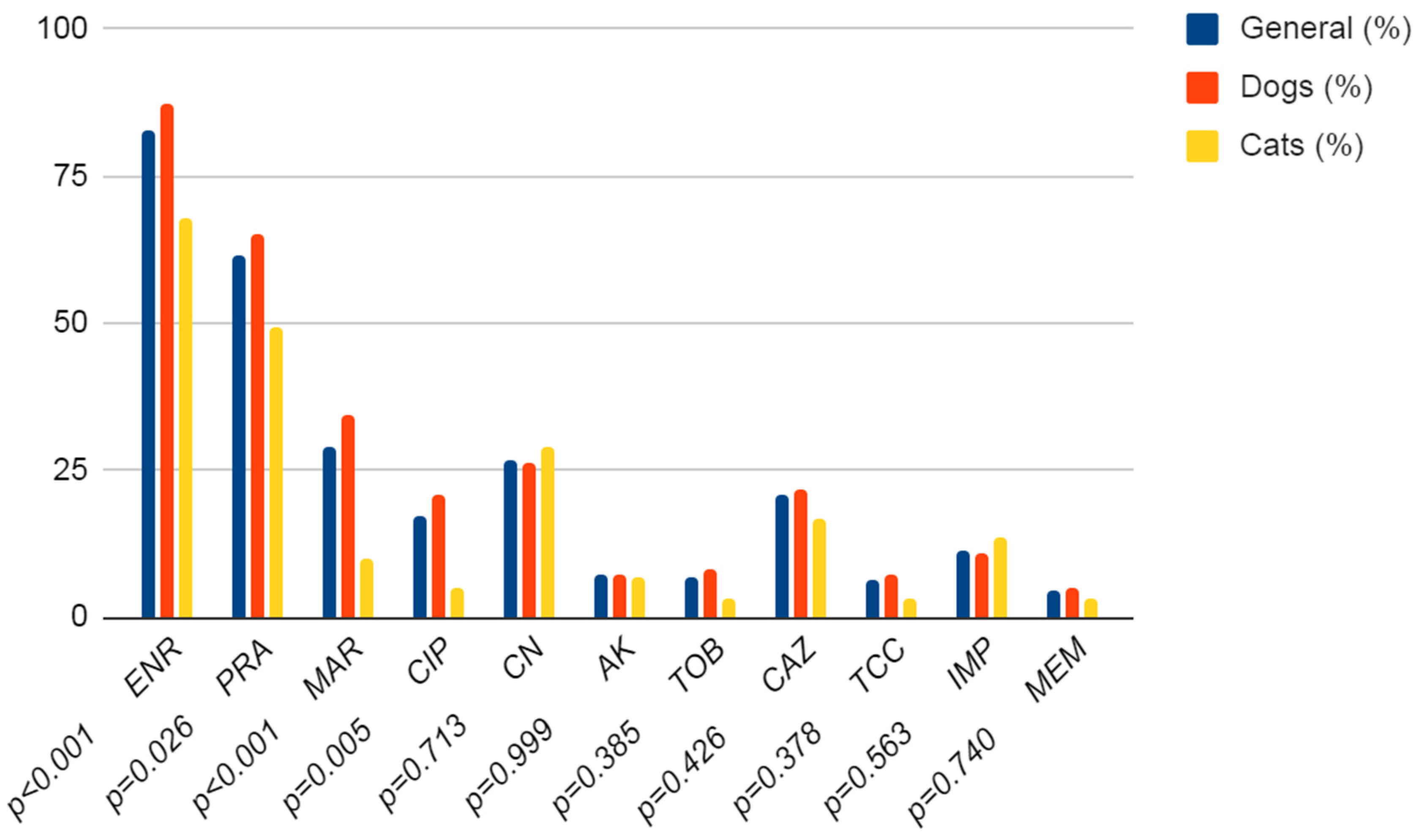
3.1. Antibiotic Resistance
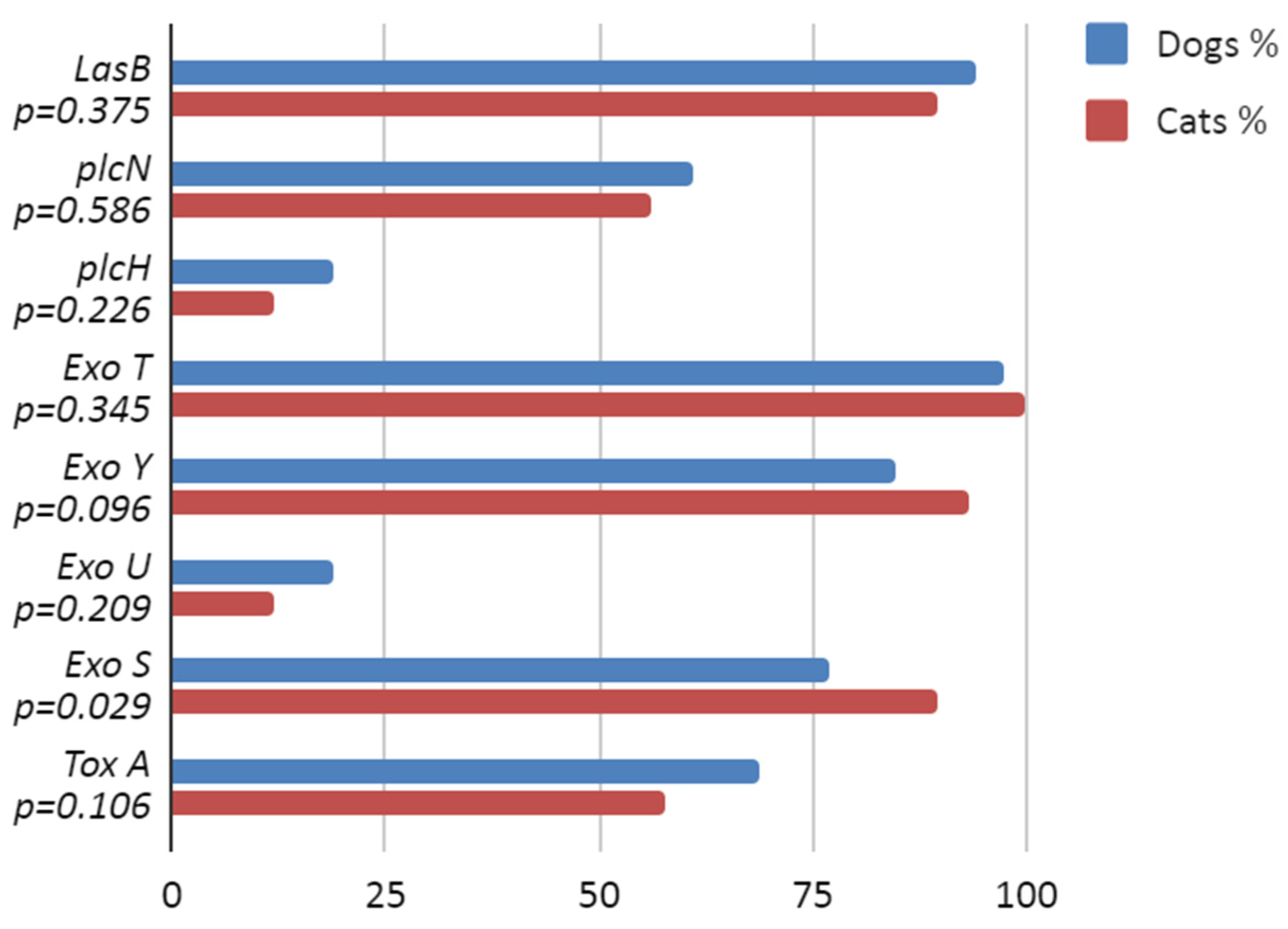
3.2. Virulence Factor Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wolska, K.; Kot, B.; Piechota, M.; Frankowska, A. Resistance of Pseudomonas aeruginosa to antibiotics. PHMD 2013, 67, 1300–1311. [Google Scholar] [CrossRef]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef]
- González-Alsina, A.; Mateu-Borrás, M.; Doménech-Sánchez, A.; Albertí, S. Pseudomonas aeruginosa and the Complement System: A Review of the Evasion Strategies. Microorganisms 2023, 11, 664. [Google Scholar] [CrossRef]
- Wood, S.J.; Kuzel, T.M.; Shafikhani, S.H. Pseudomonas aeruginosa: Infections, Animal Modeling, and Therapeutics. Cells 2023, 12, 199. [Google Scholar] [CrossRef]
- Litwin, A.; Rojek, S.; Gozdzik, W.; Duszynska, W. Pseudomonas aeruginosa device associated—Healthcare associated infections and its multidrug resistance at intensive care unit of University Hospital: Polish, 8.5-year, prospective, single-centre study. BMC Infect. Dis. 2021, 21, 180. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Point Prevalence Survey of Healthcare-Associated Infections and Antimicrobial Use in European Long-Term. Care Facilities 2016–2017; ECDC: Stockholm, Sweden, 2023. [Google Scholar]
- Rello, J.; Ausina, V.; Ricart, M.; Puzo, C.; Quintana, E.; Net, A.; Prats, G. Risk factors for infection by Pseudomonas aeruginosa in patients with ventilator-associated pneumonia. Intensive Care Med. 1994, 20, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Bassetti, M.; Vena, A.; Croxatto, A.; Righi, E.; Guery, B. How to manage Pseudomonas aeruginosa infections. Drugs Context. 2018, 7, 212527. [Google Scholar] [CrossRef] [PubMed]
- González-Díaz, A.; Medina-Polo, J.; Abad-López, P.; González-Padilla, D.A.; García-Rojo, E.; Santos-Pérez de la Blanca, R.; Hernández-Arroyo, M.; Tejido-Sánchez, Á. The challenge of Pseudomonas aeruginosa related infections in a Urology Ward: Epidemiology, risk factors and patterns of antibiotic resistance. Arch. Esp. Urol. 2020, 73, 299–306. [Google Scholar]
- Robinson, V.H.; Paterson, S.; Bennett, C.; Steen, S.I. Biofilm production of Pseudomonas spp. isolates from canine otitis in three different enrichment broths. Vet. Dermatol. 2019, 30, 218-e67. [Google Scholar] [CrossRef]
- Bajwa, J. Canine otitis externa—Treatment and complications. Can. Vet. J. 2019, 60, 97–99. [Google Scholar]
- De Sousa, T.; Garcês, A.; Silva, A.; Lopes, R.; Alegria, N.; Hébraud, M.; Igrejas, G.; Poeta, P. The Impact of the Virulence of Pseudomonas aeruginosa Isolated from Dogs. Vet. Sci. 2023, 10, 343. [Google Scholar] [CrossRef]
- Gentilini, F.; Turba, M.E.; Pasquali, F.; Mion, D.; Romagnoli, N.; Zambon, E.; Terni, D.; Peirano, G.; Pitout, J.D.D.; Parisi, A.; et al. Hospitalized Pets as a Source of Carbapenem-Resistance. Front. Microbiol. 2018, 6, 2872. [Google Scholar] [CrossRef]
- Sharma, D.; Pakravanm, N.; Pritchard, J.C.; Hartmann, F.A.; Young, K.M. Mucoid Pseudomonas aeruginosa infection in a cat with severe chronic rhinosinusitis. Vet. Clin. Pathol. 2019, 48, 300–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lister, P.D.; Wolter, D.J.; Hanson, N.D. Antibacterial-resistant Pseudomonas aeruginosa: Clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin. Microbiol. Rev. 2009, 22, 582–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Briyne, N.; Atkinson, J.; Pokludová, L.; Borriello, S.P. Antibiotics used most commonly to treat animals in Europe. Vet. Rec. 2014, 175, 325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langendonk, R.F.; Neill, D.R.; Fothergill, J.L. The building blocks of antimicrobial resistance in Pseudomonas aeruginosa: Implications for current resistance-breaking therapies. Front. Cell Infect. 2021, 11, 665759. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency (EMA). Categorisation of Antibiotics in the European Union; European Medicines Agency: London, UK, 2019. [Google Scholar]
- Velazquez-Meza, M.E.; Galarde-López, M.; Carrillo-Quiróz, B.; Alpuche-Aranda, C.M. Antimicrobial resistance: One Health approach. Vet. World. 2022, 15, 743–749. [Google Scholar] [CrossRef]
- Wi, Y.M.; Choi, J.Y.; Lee, J.Y.; Kang, C.I.; Chung, D.R.; Peck, K.R.; Song, J.H.; Ko, K.S. Antimicrobial effects of β-lactams on imipenem-resistant ceftazidime-susceptible Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2017, 61, e00054-17. [Google Scholar] [CrossRef] [Green Version]
- Çopur Çiçek, A.; Ertürk, A.; Ejder, N.; Rakici, E.; Kostakoğlu, U.; Esen Yıldız, İ.; Özyurt, S.; Sönmez, E. Screening of Antimicrobial Resistance Genes and Epidemiological Features in Hospital and Community-Associated Carbapenem-Resistant Pseudomonas aeruginosa Infections. Infect. Drug Resist. 2021, 14, 1517–1526. [Google Scholar] [CrossRef]
- World Organisation for Animal Health (OIE). OIE List of Antimicrobial Agents of Veterinary Importance. In Proceedings of the FAO2/OIE/WHO3 Expert Workshop on Non-Human Antimicrobial Usage and Antimicrobial Resistance, Geneva, Switzerland, 1–5 December 2003; Volume Xxviii, pp. 1–10. [Google Scholar]
- Ellappan, K.; Belgode Narasimha, H.; Kumar, S. Coexistence of multidrug resistance mechanisms and virulence genes in carbapenem-resistant Pseudomonas aeruginosa strains from a tertiary care hospital in South India. J. Glob. Antimicrob. Resist. 2018, 12, 37–43. [Google Scholar] [CrossRef]
- Fadhil, L.; Al-Marzoqi, A.H.; Zahraa, M.A.; Shalan, A.A. Molecular and phenotypic study of virulence genes in a pathogenic strain of Pseudomonas aeruginosa isolated from various clinical origins by PCR: Profiles of genes and toxins. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 590–598. [Google Scholar]
- Yang, J.J.; Tsuei, K.C.; Shen, E.P. The role of Type III secretion system in the pathogenesis of Pseudomonas aeruginosa microbial keratitis. Tzu. Chi. Med. J. 2022, 34, 8–14. [Google Scholar] [PubMed]
- Bradbury, R.S.; Roddam, L.F.; Merritt, A.; Reid, D.W.; Champion, A.C. Virulence gene distribution in clinical, nosocomial and environmental isolates of Pseudomonas aeruginosa. J. Med. Microbiol. 2010, 59, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Płókarz, D.; Czopowicz, M.; Bierowiec, K.; Rypuła, K. Virulence Genes as Markers for Pseudomonas aeruginosa Biofilm Formation in Dogs and Cats. Animals 2022, 12, 422. [Google Scholar] [CrossRef] [PubMed]
- James, S. Performance Standards of Antimicrobial Susceptibility Testing; Twenty-Fourth International Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022. [Google Scholar]
- Wolska, K.; Szweda, P. Genetic features of clinical Pseudomonas aeruginosa strains. Pol. J. Microbiol. 2009, 58, 255–260. [Google Scholar]
- Mokhtari, A.; Amini, K. Genotyping of Pseudomonas Aeruginosa Strains As A Multidrug Resistant (MDR) Bacterium And Evaluating The Prevalence of Esbls and Some Virulence Factors Encoding Genes By PFGE and ERIC-PCR Methods. Iran. J. Pharm. Res. 2019, 18, 1580–1594. [Google Scholar]
- Rémy, A.; Bonnin, C.E.; Agnès, B.; Jousset, T.N.; Laurent, D. Comparison of disk diffusion, MIC test strip and broth microdilution methods for cefiderocol susceptibility testing on carbapenem-resistant enterobacterales. Clin. Microbiol. Inf. 2022, 28, e1–e1156. [Google Scholar]
- Penna, B.; Thomé, S.; Martins, R.; Martins, G.; Lilenbaum, W. In Vitro antimicrobial resistance of Pseudomonas aeruginosa isolated from canine otitis externa in Rio de Janeiro. Braz. J. Microbiol. 2011, 42, 1434–1436. [Google Scholar] [CrossRef] [Green Version]
- Eliasi, U.L.; Sebola, D.; Oguttu, J.W.; Qekwana, D.N. Antimicrobial resistance patterns of Pseudomonas aeruginosa isolated from canine clinical cases at a veterinary academic hospital in South Africa. J. S. Afr. Vet. Assoc. 2020, 91, e1–e6. [Google Scholar] [CrossRef]
- Djuikoue, C.I.; Djouela Djoulako, P.D.; Same Njanjo, H.V.; Kiyang, C.P.; Djantou Biankeu, F.L.; Guegang, C.; Tchouotou, A.S.D.; Wouambo, R.K.; Thumamo Pokam, B.D.; Apalata, T.; et al. Phenotypic Characterization and Prevalence of Carbapenemase-Producing Pseudomonas aeruginosa Isolates in Six Health Facilities in Cameroon. BioMed 2023, 3, 77–88. [Google Scholar] [CrossRef]
- Yukawa, S.; Tsuyuki, Y.; Sato, T.; Fukuda, A.; Usui, M.; Tamura, Y. Antimicrobial Resistance of Pseudomonas aeruginosa Isolated from Dogs and Cats in Primary Veterinary Hospitals in Japan. Jpn. J. Infect. Dis. 2017, 70, 461–463. [Google Scholar] [CrossRef] [Green Version]
- Pottier, M.; Castagnet, S.; Gravey, F.; Leduc, G.; Sévin, C.; Petry, S.; Giard, J.-C.; Le Hello, S.; Léon, A. Antimicrobial Resistance and Genetic Diversity of Pseudomonas aeruginosa Strains Isolated from Equine and Other Veterinary Samples. Pathogens 2023, 12, 64. [Google Scholar] [CrossRef] [PubMed]
- Mekić, S.; Matanović, K.; Šeol, B. Antimicrobial susceptibility of Pseudomonas aeruginosa isolates from dogs with otitis externa. Vet. Rec. 2011, 169, 125. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, W.; Izumi, K.; Yoshida, S.; Takizawa, S.; Sakaguchi, K.; Iyori, K.; Minoshima, K.I.; Takano, S.; Kitagawa, M.; Nagano, Y.; et al. Antimicrobial Resistance and Type III Secretion System Virulotypes of Pseudomonas aeruginosa Isolates from Dogs and Cats in Primary Veterinary Hospitals in Japan: Identification of the International High-Risk Clone Sequence Type 235. Microbiol. Spectr. 2021, 9, e0040821. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.C.; Medeiros, C.M.O.; Silva, I.N.G.; Monteiro, A.J.; Leite, C.A.L.; Carvalho, C.B.M. Susceptibilidade a antimicrobianos de bactérias isoladas de otite externa em cães. Arq. Bras. Med. Veterinária Zootec. 2005, 57, 405–408. [Google Scholar] [CrossRef]
- Dégi, J.; Moțco, O.A.; Dégi, D.M.; Suici, T.; Mareș, M.; Imre, K.; Cristina, R.T. Antibiotic Susceptibility Profile of Pseudomonas aeruginosa Canine Isolates from a Multicentric Study in Romania. Antibiotics. 2021, 10, 846. [Google Scholar] [CrossRef]
- Callens, M.; Scornavacca, C.; Bedhomme, S. Evolutionary responses to codon usage of horizontally transferred genes in Pseudomonas aeruginosa: Gene retention, amelioration and compensatory evolution. Microb. Genom. 2021, 7, 000587. [Google Scholar] [CrossRef]
- Brzozowski, M.; Krukowska, Ż.; Galant, K.; Jursa-Kulesza, J.; Kosik-Bogacka, D. Genotypic characterisation and antimicrobial resistance of Pseudomonas aeruginosa strains isolated from patients of different hospitals and medical centres in Poland. BMC Infect. Dis. 2020, 20, 693. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Surveillance of Antimicrobial Resistance in Europe; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2017. [Google Scholar]
- Elmouaden, C.; Laglaoui, A.; Ennanei, L.; Bakkali, M.; Abid, M. Virulence genes and antibiotic resistance of Pseudomonas aeruginosa isolated from patients in the Northwestern of Morocco. J. Infect. Dev. Ctries. 2019, 13, 892–898. [Google Scholar] [CrossRef]
- Feltman, H.; Schulert, G.; Khan, S.; Jain, M.; Peterson, L.; Hauser, A.R. Prevalence of type III secretion genes in clinical and environmental isolates of Pseudomonas aeruginosa. Microbiology 2001, 147, 2659–2669. [Google Scholar] [CrossRef] [Green Version]
- Horna, G.; Amaro, C.; Palacios, A.; Guerra, H.; Ruiz, J. High frequency of the exoU+/exoS+ genotype associated with multidrug-resistant “high-risk clones” of Pseudomonas aeruginosa clinical isolates from Peruvian hospitals. Sci. Rep. 2019, 9, 10874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattab, J.; Mosca, F.; Di Francesco, C.E.; Aste, G.; Marruchella, G.; Guardiani, P.; Tiscar, P.G. Occurrence, antimicrobial susceptibility, and pathogenic factors of Pseudomonas aeruginosa in canine clinical samples. Vet. World 2021, 14, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Aslataş, Ö.; Tűrkyilmaz, S.; KESKİN, O.; Gűllű Yűcetepe, A.; Bűykatal, K. Molecular Characterization of Pseudomonas aeruginosa Isolated From Clinical Bovine Mastitis Cases. Kafkas. Univ. Vet. Fak. Derg. 2022, 28, 747–759. [Google Scholar]
- Amirmozafari, N.; Fallah, J.; Habibi, A. Association of the Exotoxin A and Exoenzyme S with Antimicrobial Resistance in Pseudomonas Aeruginosa Strains. Arch. Iran. Med. 2016, 19, 353–358. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gen | Primers | Annealing Temperature | Product Size | References |
|---|---|---|---|---|
| Las B | F:5′-GGAATGAACGAAGCGTTCTCCGAC-3′ R:5′-TTGGCGTCGACGAACACCTCG-3′ | 55 °C a | 284 | Wolska et al., 2009 [29] |
| plcN | F:5′-TCCGTTATCGCAACCAGCCCTACG-3′ R:5′-TCGCTGTCGAGCAGGTCGAAC-3′ | 55 °C a | 481 | Wolska et al., 2009 [29] |
| plcH | F:5′-GCACGTGGTCATCCTGATGC-3′ R:5′-TCCGTAGGCGTCGACGTAC-3′ | 55 °C a | 608 | Wolska et al., 2009 [29] |
| ToxA | F:5′-CTGCGCGGGTCTATGTGCC-3′ R:5′-GATGCTGGACGGGTCGAG-3′ | 55 °C a | 270 | Wolska et al., 2009 [29] |
| ExoS | F:5′-CGTCGTGTTCAAGCAGATGGTGCTG-3′ R:5′-CCGAACCGCTTCACCAGGC-3′ | 55 °C a | 444 | Wolska et al., 2009 [29] |
| ExoU | F:5′-CCGTTGTGGTGCCGTTGAAG-3′ R:5′-CCAGATGTTCACCGACTCGC-3′ | 58 °C b | 134 | Mokhtaria and Amini 2019 [29] |
| ExoY | F:5′-CGGATTCTATGGCAGGGAGG-3′ R:5′-GCCCTTGATGCACTCGACCA-3′ | 58 °C b | 289 | Mokhtaria and Amini 2019 [30] |
| ExoT | F:5′-AATCGCCGTCCAACTGCATGCG-3′ R: 5′-TGTTCGCCGAGGTACTGCTC-3′ | 58 °C b | 152 | Mokhtaria and Amini 2019 [30] |
| Antibiotic | External Auricular Canal (n = 118) | Skin and Appendages (n = 31) | Respiratory System and Oral Cavity (n = 36) | Uro-Genital System (n = 10) | Wounds (n = 16) | p | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| n | % (95% PU) | n | % (95% PU) | n | % (95% PU) | n | % (95% PU) | n | % (95% PU) | ||
| ENR | 103 | 87.3 (80.1–92.1) | 24 | 77.4 (60.2–88.6) | 31 | 86.1 (713–93.9) | 10 | 100 (72.2–100) | 16 | 100 (8.6–100) | 0.052 |
| MAR | 50 | 42.4 (33.8–51.4) | 6 | 19.4 (9.2–36.3) | 7 | 19.4 (9.8–35.0) | 2 | 20.0 (5.7–51.0) | 7 | 43.8 (23.1–66.8) | 0.016 * |
| CIP | 31 | 26.3 (19.2–34.9) | 1 | 3.2 (0.6–16.2) | 7 | 19.4 (9.8–35.0) | 1 | 10.0 (1.8–40.4) | 4 | 25.0 (10.2–49.5) | 0.023 * |
| PRA | 80 | 67.8 (58.9–75.6) | 16 | 51.6 (34.8–68.0) | 25 | 69.4 (53.1–82.0) | 4 | 40.0 (16.8–68.7) | 12 | 75.0 (50.5–89.8) | 0.172 |
| CAZ | 32 | 27.1 (19.9–35.8) | 2 | 6.5 (1.8–20.7) | 6 | 16.7 (7.9–31.9) | 2 | 20.0 (5.7–51.0) | 4 | 25.0 (10.2–49.5) | 0.089 |
| AK | 11 | 9.3 (5.3–15.9) | 1 | 3.2 (0.6–16.2) | 2 | 5.6 (1.5–18.1) | 1 | 10.0 (1.8–40.4) | 1 | 6.3 (1.1–28.3) | 0.757 |
| CN | 40 | 33.9 (26.0–42.8) | 3 | 9.7 (3.3–24.9) | 7 | 19.4 (9.8–350) | 1 | 10.0 (1.8–40.4) | 5 | 31.3 (14.2–55.6) | 0.022 * |
| TOB | 11 | 9.3 (5.3–15.9) | 2 | 6.5 (1.8–20.7) | 1 | 2.8 (0.5–14.2) | 0 | 0 (0–27.8) | 3 | 18.8 (6.6–43.0) | 0.225 |
| TTC | 8 | 6.8 (3.5–12.8) | 0 | 0 (0–11.0) | 1 | 2.8 (0.5–14.2) | 2 | 20.0 (5.7–51.0) | 4 | 25.0 (10.2–49.5) | 0.014 * |
| IMP | 11 | 9.3 (5.3–15.9) | 2 | 6.5 (1.8–20.7) | 4 | 11.1 (4.4–25.3) | 4 | 40.0 (16.8–68.7) | 2 | 12.5 (3.5–36.0) | 0.150 |
| MEM | 7 | 5.9 (2.9–11.7) | 0 | 0 (0–11.0) | 0 | 0 (0–9.6) | 2 | 20.0 (5.7–51.0) | 1 | 6.3 (1.1–28.3) | 0.042 * |
| Antibiotic | Respiratory System (n = 49) | Others (n = 10) | |||
|---|---|---|---|---|---|
| n | % (95% PU) | n | % (95% PU) | p | |
| ENR | 32 | 65.3 (51.3–77.1) | 8 | 80.0 (49.0–94.3) | 0.476 |
| MAR | 5 | 10.2 (4.4–21.8) | 1 | 10.0 (1.8–40.4) | 0.999 |
| CIP | 2 | 4.1 (1.1–13.7) | 1 | 10.0 (1.8–40.4) | 0.433 |
| PRA | 23 | 46.9 (33.7–60.6) | 6 | 60.0 (31.3–83.2) | 0.452 |
| CAZ | 8 | 16.3 (8.5–29.0) | 2 | 20.0 (5.7–51.0) | 0.673 |
| AK | 3 | 6.1 (2.1–16.5) | 1 | 10.0 (1.8–40.4) | 0.535 |
| CN | 13 | 26.5 (16.2–40.3) | 4 | 40.0 (16.8–68.7) | 0.453 |
| TOB | 0 | 0 (0–7.3) | 2 | 20.0 (5.7–51.0) | 0.026 * |
| TTC | 2 | 4.1 (1.1–13.7) | 0 | 0 (0–27.8) | 0.999 |
| IMP | 6 | 12.2 (5.7–24.2) | 2 | 20.0 (5.7–51.0) | 0.613 |
| MEM | 1 | 2.0 (0.4–10.7) | 1 | 10.0 (1.8–40.4) | 0.313 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Płókarz, D.; Bierowiec, K.; Rypuła, K. Screening for Antimicrobial Resistance and Genes of Exotoxins in Pseudomonas aeruginosa Isolates from Infected Dogs and Cats in Poland. Antibiotics 2023, 12, 1226. https://doi.org/10.3390/antibiotics12071226
Płókarz D, Bierowiec K, Rypuła K. Screening for Antimicrobial Resistance and Genes of Exotoxins in Pseudomonas aeruginosa Isolates from Infected Dogs and Cats in Poland. Antibiotics. 2023; 12(7):1226. https://doi.org/10.3390/antibiotics12071226
Chicago/Turabian StylePłókarz, Daria, Karolina Bierowiec, and Krzysztof Rypuła. 2023. "Screening for Antimicrobial Resistance and Genes of Exotoxins in Pseudomonas aeruginosa Isolates from Infected Dogs and Cats in Poland" Antibiotics 12, no. 7: 1226. https://doi.org/10.3390/antibiotics12071226