
Cracking the Code: Unveiling the Diversity of Carbapenem-Resistant Klebsiella pneumoniae Clones in the Arabian Peninsula through Genomic Surveillance
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Prevalence and Distribution of Carbapenem-Resistant K. pneumoniae Sequence Types (STs)
2.2. Resistome Characterization of Carbapenem-Resistant K. pneumoniae Isolates
2.3. Mobile Colistin Resistance Elements (mcr)
2.4. Clonal Clustering and Relatedness of the CRKP Isolates
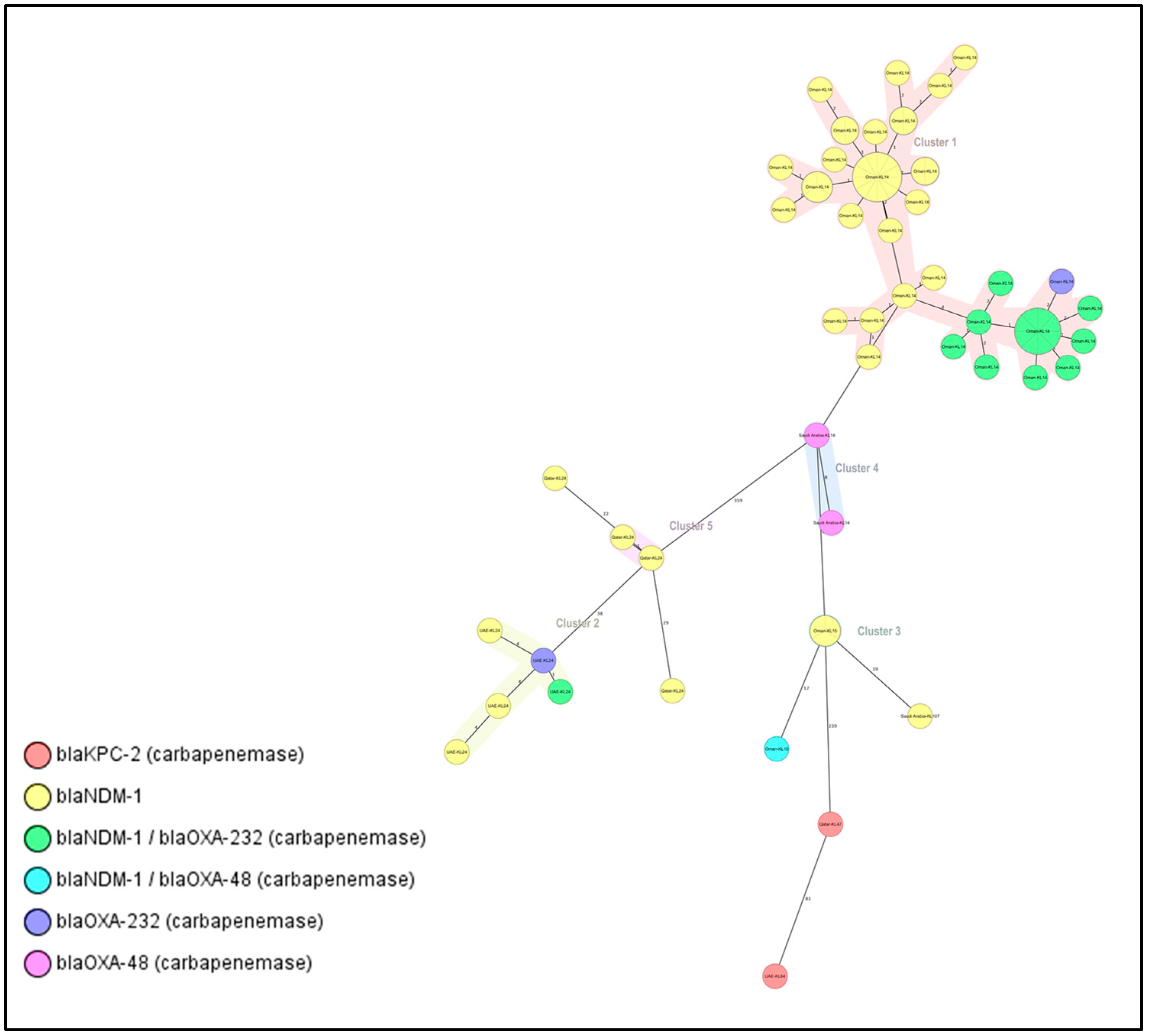
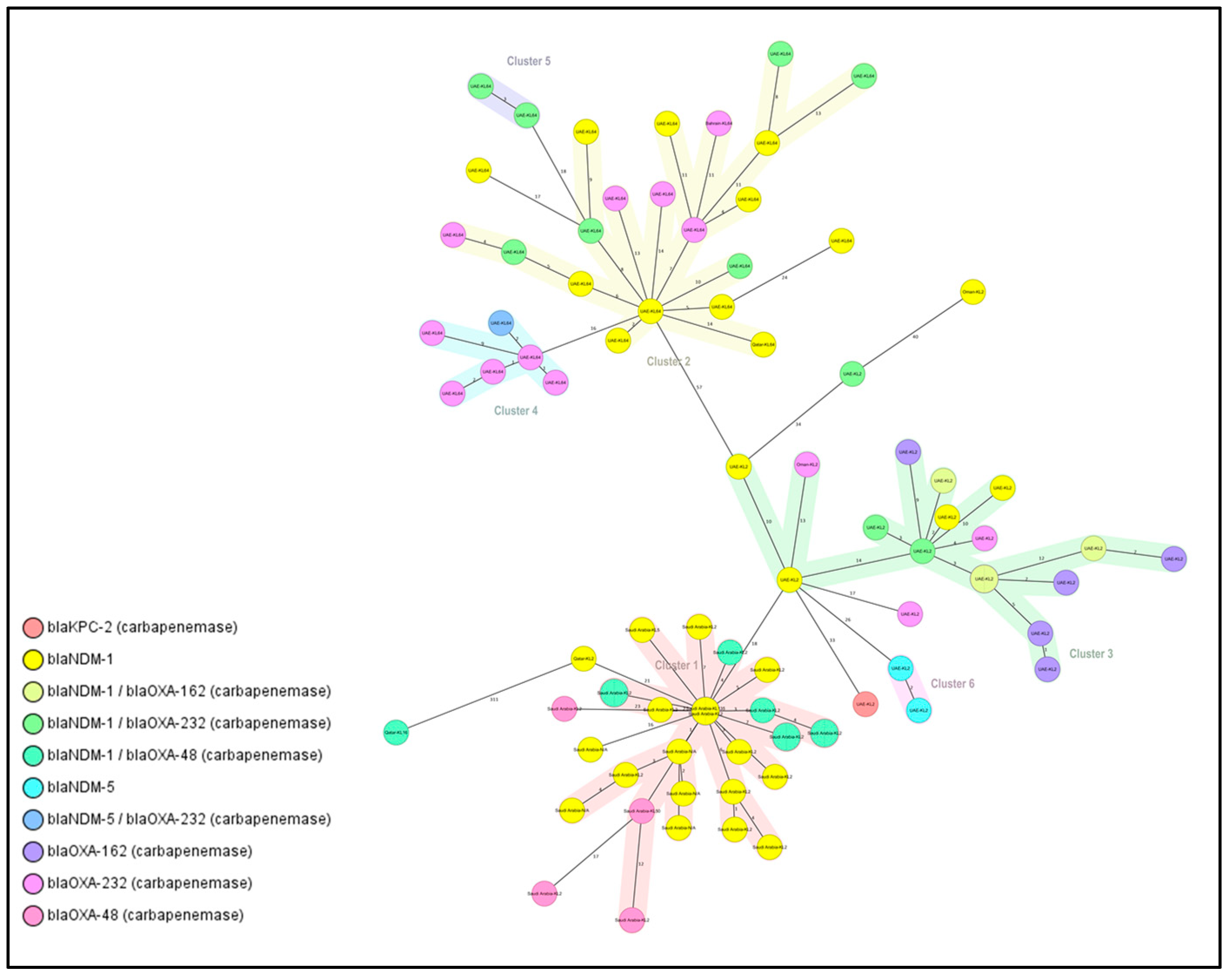
2.5. Clusters and Singletons Associated with ST-14 and ST-147
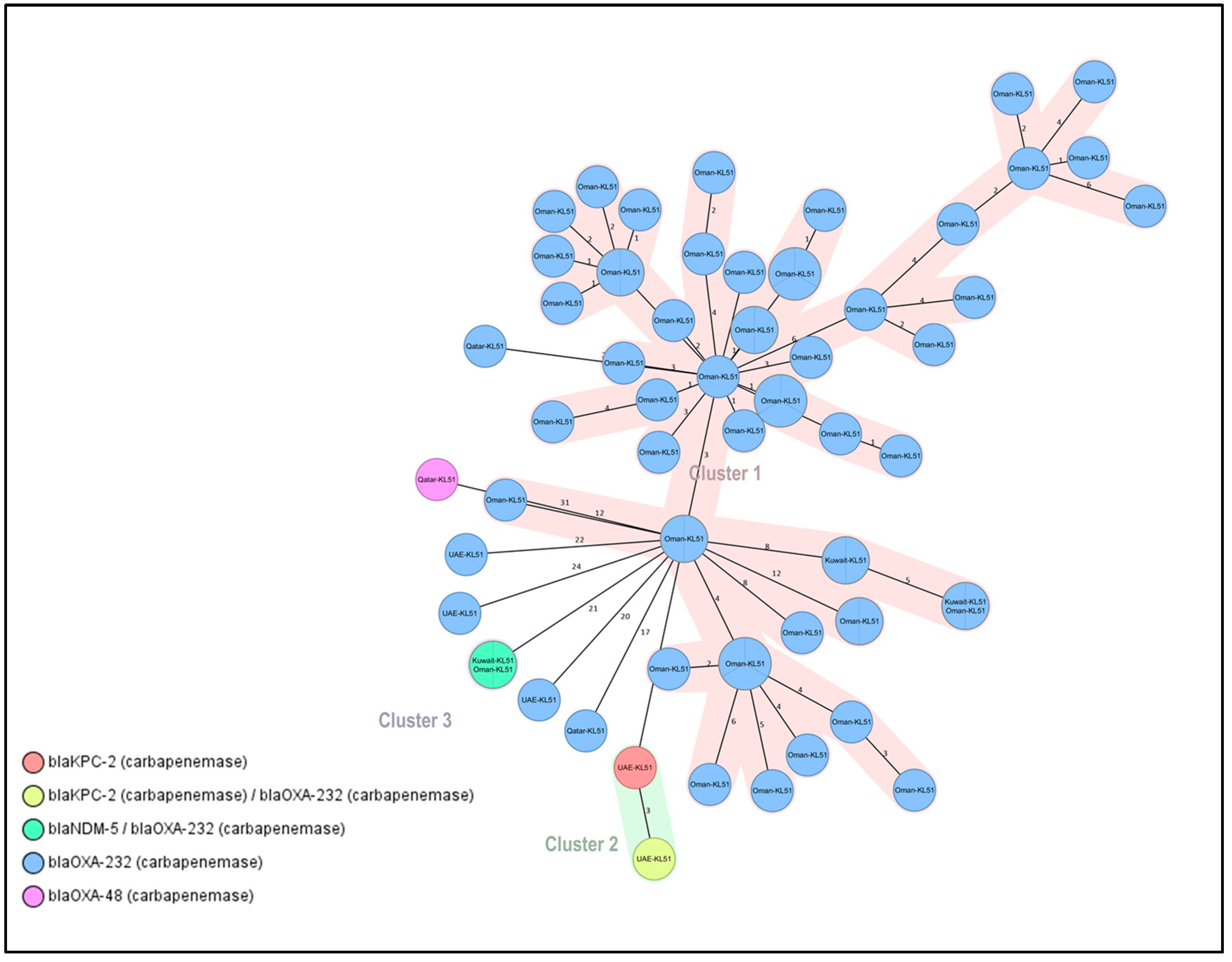
2.6. Outbreaks Associated with ST-231 and ST-2096 CRKP
2.7. Clustering of Isolates among ST-15, ST-101, and ST-45 Lineages
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vivas, R.; Dolabella, S.S.; Barbosa, A.A.T.; Jain, S. Prevalence of Klebsiella pneumoniae carbapenemase—And New Delhi metallo-beta-lactamase-positive K. pneumoniae in Sergipe, Brazil, and combination therapy as a potential treatment option. Rev. Soc. Bras. Med. Trop. 2020, 53, e20200064. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Review on Antimicrobial Resistance. Wellcome Trust and HM Government. 2016. Available online: https://amr-review.org/sites/default/files/160525_Final%20paper_with%20cover.pdf (accessed on 14 May 2023).
- Feil, E.J. Enterobacteriaceae: Joining the dots with pan-European epidemiology. Lancet Infect. Dis. 2017, 17, 118–119. [Google Scholar] [CrossRef] [PubMed]
- Perez, F.; Villegas, M.V. The role of surveillance systems in confronting the global crisis of antibiotic-resistant bacteria. Curr. Opin. Infect. Dis. 2015, 28, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Han, R.; Shi, Q.; Wu, S.; Yin, D.; Peng, M.; Dong, D.; Zheng, Y.; Guo, Y.; Zhang, R.; Hu, F. Dissemination of Carbapenemases (KPC, NDM, OXA-48, IMP, and VIM) Among Carbapenem-Resistant Enterobacteriaceae Isolated From Adult and Children Patients in China. Front. Cell. Infect. Microbiol. 2020, 10, 314. [Google Scholar] [CrossRef] [PubMed]
- Tischendorf, J.; de Avila, R.A.; Safdar, N. Risk of infection following colonization with carbapenem-resistant Enterobactericeae: A systematic review. Am. J. Infect. Control 2016, 44, 539–543. [Google Scholar] [CrossRef] [Green Version]
- Hammoudi Halat, D.; Ayoub Moubareck, C. The Current Burden of Carbapenemases: Review of Significant Properties and Dissemination among Gram-Negative Bacteria. Antibiotics 2020, 9, 186. [Google Scholar] [CrossRef] [Green Version]
- Naas, T.; Dortet, L.; Iorga, B.I. Structural and Functional Aspects of Class A Carbapenemases. Curr. Drug Targets 2016, 17, 1006–1028. [Google Scholar] [CrossRef]
- Moghnia, O.H.; Rotimi, V.O.; Al-Sweih, N.A. Preponderance of blaKPC-Carrying Carbapenem-Resistant Enterobacterales among Fecal Isolates from Community Food Handlers in Kuwait. Front. Microbiol. 2021, 12, 737828. [Google Scholar] [CrossRef]
- Jeon, J.H.; Lee, J.H.; Lee, J.J.; Park, K.S.; Karim, A.M.; Lee, C.R.; Jeong, B.C.; Lee, S.H. Structural basis for carbapenem-hydrolyzing mechanisms of carbapenemases conferring antibiotic resistance. Int. J. Mol. Sci. 2015, 16, 9654–9692. [Google Scholar] [CrossRef] [Green Version]
- Walsh, T.R.; Toleman, M.A.; Poirel, L.; Nordmann, P. Metallo-beta-lactamases: The quiet before the storm? Clin. Microbiol. Rev. 2005, 18, 306–325. [Google Scholar] [CrossRef] [Green Version]
- Jamal, W.Y.; Albert, M.J.; Rotimi, V.O. High Prevalence of New Delhi Metallo-β-Lactamase-1 (NDM-1) Producers among Carbapenem-Resistant Enterobacteriaceae in Kuwait. PLoS ONE 2016, 11, e0152638. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Hao, Q. Crystal structure of NDM-1 reveals a common β-lactam hydrolysis mechanism. FASEB J. 2011, 25, 2574–2582. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; McNally, A.; Zong, Z. NDM Metallo-β-Lactamases and Their Bacterial Producers in Health Care Settings. Clin. Microbiol. Rev. 2019, 32, e00115-8. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Potron, A.; Nordmann, P. OXA-48-like carbapenemases: The phantom menace. Int. J. Antimicrob. Agents 2012, 67, 1597–1606. [Google Scholar] [CrossRef] [Green Version]
- Potron, A.; Rondinaud, E.; Poirel, L.; Belmonte, O.; Boyer, S.; Camiade, S.; Nordmann, P. Genetic and biochemical characterisation of OXA-232, a carbapenem-hydrolysing class D β-lactamase from Enterobacteriaceae. Int. J. Antimicrob. Agents 2013, 41, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Pitout, J.D.D.; Peirano, G.; Kock, M.M.; Strydom, K.A.; Matsumura, Y. The Global Ascendency of OXA-48-Type Carbapenemases. Clin. Microbiol. Rev. 2019, 33, e00102-19. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Yang, X.; Chan, E.W.; Zhang, R.; Chen, S. Klebsiella species: Taxonomy, hypervirulence and multidrug resistance. EbioMedicine 2022, 79, 103998. [Google Scholar] [CrossRef]
- Abd El Ghany, M.; Sharaf, H.; Al-Agamy, M.H.; Shibl, A.; Hill-Cawthorne, G.A.; Hong, P.Y. Genomic characterization of NDM-1 and 5, and OXA-181 carbapenemases in uropathogenic Escherichia coli isolates from Riyadh, Saudi Arabia. PLoS ONE 2018, 13, e0201613. [Google Scholar] [CrossRef]
- Al-Agamy, M.H.; Aljallal, A.; Radwan, H.H.; Shibl, A.M. Characterization of carbapenemases, ESBLs, and plasmid-mediated quinolone determinants in carbapenem-insensitive Escherichia coli and Klebsiella pneumoniae in Riyadh hospitals. J. Infect. Public Health 2018, 11, 64–68. [Google Scholar] [CrossRef]
- Al-Baloushi, A.E.; Pál, T.; Ghazawi, A.; Sonnevend, A. Genetic support of carbapenemases in double carbapenemase producer Klebsiella pneumoniae isolated in the Arabian Peninsula. Acta Microbiol. Immunol. Hung. 2018, 65, 135–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alotaibi, F.E.; Bukhari, E.E.; Al-Mohizea, M.M.; Hafiz, T.; Essa, E.B.; AlTokhais, Y.I. Emergence of carbapenem-resistant Enterobacteriaceae isolated from patients in a university hospital in Saudi Arabia. Epidemiology, clinical profiles and outcomes. J. Infect. Public Health 2017, 10, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Memish, Z.A.; Assiri, A.; Almasri, M.; Roshdy, H.; Hathout, H.; Kaase, M.; Gatermann, S.G.; Yezli, S. Molecular characterization of carbapenemase production among gram-negative bacteria in Saudi Arabia. Microb. Drug Resist. 2015, 21, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Moubareck, C.A.; Mouftah, S.F.; Pál, T.; Ghazawi, A.; Halat, D.H.; Nabi, A.; AlSharhan, M.A.; AlDeesi, Z.O.; Peters, C.C.; Celiloglu, H.; et al. Clonal emergence of Klebsiella pneumoniae ST14 co-producing OXA-48-type and NDM carbapenemases with high rate of colistin resistance in Dubai, United Arab Emirates. Int. J. Antimicrob. Agents 2018, 52, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Pál, T.; Ghazawi, A.; Darwish, D.; Villa, L.; Carattoli, A.; Hashmey, R.; Aldeesi, Z.; Jamal, W.; Rotimi, V.; Al-Jardani, A.; et al. Characterization of NDM-7 Carbapenemase-Producing Escherichia coli Isolates in the Arabian Peninsula. Microb. Drug Resist. 2017, 23, 871–878. [Google Scholar] [CrossRef]
- Sonnevend, Á.; Ghazawi, A.A.; Hashmey, R.; Jamal, W.; Rotimi, V.O.; Shibl, A.M.; Al-Jardani, A.; Al-Abri, S.S.; Tariq, W.U.; Weber, S.; et al. Characterization of Carbapenem-Resistant Enterobacteriaceae with High Rate of Autochthonous Transmission in the Arabian Peninsula. PLoS ONE 2015, 10, e0131372. [Google Scholar] [CrossRef]
- Uz Zaman, T.; Albladi, M.; Siddique, M.I.; Aljohani, S.M.; Balkhy, H.H. Insertion element mediated mgrB disruption and presence of ISKpn28 in colistin-resistant Klebsiella pneumoniae isolates from Saudi Arabia. Infect. Drug Resist. 2018, 11, 1183–1187. [Google Scholar] [CrossRef] [Green Version]
- Zowawi, H.M.; Sartor, A.L.; Balkhy, H.H.; Walsh, T.R.; Al Johani, S.M.; AlJindan, R.Y.; Alfaresi, M.; Ibrahim, E.; Al-Jardani, A.; Al-Abri, S.; et al. Molecular characterization of carbapenemase-producing Escherichia coli and Klebsiella pneumoniae in the countries of the Gulf cooperation council: Dominance of OXA-48 and NDM producers. Antimicrob. Agents Chemother. 2014, 58, 3085–3090. [Google Scholar] [CrossRef] [Green Version]
- Al-Abdely, H.; AlHababi, R.; Dada, H.M.; Roushdy, H.; Alanazi, M.M.; Alessa, A.A.; Gad, N.M.; Alasmari, A.M.; Radwan, E.E.; Al-Dughmani, H.; et al. Molecular characterization of carbapenem-resistant Enterobacterales in thirteen tertiary care hospitals in Saudi Arabia. Ann. Saudi Med. 2021, 41, 63–70. [Google Scholar] [CrossRef]
- Sonnevend, Á.; Abdulrazzaq, N.; Ghazawi, A.; Thomsen, J.; Bharathan, G.; Makszin, L.; Rizvi, T.A.; Pál, T.; UAE CRE Study Group. The first nationwide surveillance of carbapenem-resistant Enterobacterales in the United Arab Emirates-increased association of Klebsiella pneumoniae CC14 clone with Emirati patients. Int. J. Infect. Dis. 2022, 120, 103–112. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, M.; Qiu, S.; Wang, J.; Peng, J.; Zhao, P.; Zhu, R.; Wang, H.; Li, Y.; Wang, K.; et al. Antimicrobial activity and stability of the D-amino acid substituted derivatives of antimicrobial peptide polybia-MPI. AMB Express 2016, 6, 122. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Xiong, Y.; Rogers, L.; Carter, G.P.; French, N. Genome-by-genome approach for fast bacterial genealogical relationship evaluation. Bioinformatics 2018, 34, 3025–3027. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef] [PubMed]
- Pei, N.; Li, Y.; Liu, C.; Jian, Z.; Liang, T.; Zhong, Y.; Sun, W.; He, J.; Cheng, X.; Li, H.; et al. Large-Scale Genomic Epidemiology of Klebsiella pneumoniae Identified Clone Divergence with Hypervirulent Plus Antimicrobial-Resistant Characteristics Causing Within-Ward Strain Transmissions. Microbiol. Spectr. 2022, 10, e0269821. [Google Scholar] [CrossRef] [PubMed]
- Findlay, J.; Hopkins, K.L.; Loy, R.; Doumith, M.; Meunier, D.; Hill, R.; Pike, R.; Mustafa, N.; Livermore, D.M.; Woodford, N. OXA-48-like carbapenemases in the UK: An analysis of isolates and cases from 2007 to 2014. J. Antimicrob. Chemother. 2017, 72, 1340–1349. [Google Scholar] [CrossRef]
- Mouftah, S.F.; Pál, T.; Higgins, P.G.; Ghazawi, A.; Idaghdour, Y.; Alqahtani, M.; Omrani, A.S.; Rizvi, T.A.; Sonnevend, Á. Diversity of carbapenem-resistant Klebsiella pneumoniae ST14 and emergence of a subgroup with KL64 capsular locus in the Arabian Peninsula. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Giske, C.G.; Fröding, I.; Hasan, C.M.; Turlej-Rogacka, A.; Toleman, M.; Livermore, D.; Woodford, N.; Walsh, T.R. Diverse sequence types of Klebsiella pneumoniae contribute to the dissemination of blaNDM-1 in India, Sweden, and the United Kingdom. Antimicrob. Agents Chemother. 2012, 56, 2735–2738. [Google Scholar] [CrossRef] [Green Version]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 4, 252–275. [Google Scholar] [CrossRef]
- Yoon, E.J.; Yang, J.W.; Kim, J.O.; Lee, H.; Lee, K.J.; Jeong, S.H. Carbapenemase-producing Enterobacteriaceae in South Korea: A report from the National Laboratory Surveillance System. Future Microbiol. 2018, 13, 771–783. [Google Scholar] [CrossRef]
- Holt, K.E.; Wertheim, H.; Zadoks, R.N.; Baker, S.; Whitehouse, C.A.; Dance, D.; Jenney, A.; Connor, T.R.; Hsu, L.Y.; Severin, J.; et al. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. USA 2015, 112, E3574–E3581. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.R.; Molton, J.S.; Wyres, K.L.; Gorrie, C.; Wong, J.; Hoh, C.H.; Teo, J.; Kalimuddin, S.; Lye, D.C.; Archuleta, S.; et al. Differential host susceptibility and bacterial virulence factors driving Klebsiella liver abscess in an ethnically diverse population. Sci. Rep. 2016, 6, 29316. [Google Scholar] [CrossRef] [Green Version]
- Shankar, C.; Mathur, P.; Venkatesan, M.; Pragasam, A.K.; Anandan, S.; Khurana, S.; Veeraraghavan, B. Rapidly disseminating blaOXA-232 carrying Klebsiella pneumoniae belonging to ST231 in India: Multiple and varied mobile genetic elements. BMC Microbiol. 2019, 19, 137. [Google Scholar] [CrossRef]
- Arabaghian, H.; Salloum, T.; Alousi, S.; Panossian, B.; Araj, G.F.; Tokajian, S. Molecular Characterization of Carbapenem Resistant Klebsiella pneumoniae and Klebsiella quasipneumoniae Isolated from Lebanon. Sci. Rep. 2019, 9, 531. [Google Scholar] [CrossRef] [Green Version]
- Blundell-Hunter, G.; Enright, M.C.; Negus, D.; Dorman, M.J.; Beecham, G.E.; Pickard, D.J.; Wintachai, P.; Voravuthikunchai, S.P.; Thomson, N.R.; Taylor, P.W. Characterisation of Bacteriophage-Encoded Depolymerases Selective for Key Klebsiella pneumoniae Capsular Exopolysaccharides. Front. Cell. Infect. Microbiol. 2021, 11, 686090. [Google Scholar] [CrossRef]
- Koskinen, K.; Penttinen, R.; Örmälä-Odegrip, A.M.; Giske, C.G.; Ketola, T.; Jalasvuori, M. Systematic Comparison of Epidemic and Non-Epidemic Carbapenem Resistant Klebsiella pneumoniae Strains. Front. Cell. Infect. Microbiol. 2021, 11, 599924. [Google Scholar] [CrossRef]
- Nagaraj, G.; Shamanna, V.; Govindan, V.; Rose, S.; Sravani, D.; Akshata, K.P.; Shincy, M.R.; Venkatesha, V.T.; Abrudan, M.; Argimón, S.; et al. High-Resolution Genomic Profiling of Carbapenem-Resistant Klebsiella pneumoniae Isolates: A Multicentric Retrospective Indian Study. Clin. Infect. Dis. 2021, 73 (Suppl. S4), S300–S307. [Google Scholar] [CrossRef] [PubMed]
- Hala, S.; Antony, C.P.; Alshehri, M.; Alsaedi, A.; Thaqafi, O.A.; Al-Ahmadi, G.J.; Kaaki, M.; Alazmi, M.A.; Alhaj-Hussein, T.B.; Yasen, M.; et al. An Emerging Clone (ST2096) of Klebsiella pneumoniae Clonal Complex 14 With Enhanced Virulence Causes an Outbreak in Saudi Arabia. J. Infect. Public Health 2020, 13, 363–364. [Google Scholar] [CrossRef]
- Gu, D.; Dong, N.; Zheng, Z.; Gu, D.; Dong, N.; Zheng, Z.; Lin, D.; Huang, M.; Wang, L.; Chan, E.W.; et al. A fatal outbreak of ST11 carbapenem-resistant hypervirulent Klebsiella pneumoniae in a Chinese hospital: A molecular epidemiological study. Lancet Infect. Dis. 2018, 18, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Balushi, M.A.; Kumar, R.; Al-Rashdi, A.; Ratna, A.; Al-Jabri, A.; Al-Shekaili, N.; Rani, R.; Sumri, S.A.; Al-Ghabshi, L.; Al-Abri, S.; et al. Genomic analysis of the emerging carbapenem-resistant Klebsiella pneumoniae sequence type 11 harbouring Klebsiella pneumoniae carbapenemase (KPC) in Oman. J. Infect. Public Health 2022, 15, 1089–1096. [Google Scholar] [CrossRef]
- Safavi, M.; Bostanshirin, N.; Hajikhani, B.; Yaslianifard, S.; van Belkum, A.; Goudarzi, M.; Hashemi, A.; Darban-Sarokhalil, D.; Dadashi, M. Global genotype distribution of human clinical isolates of New Delhi metallo-β-lactamase-producing Klebsiella pneumoniae; A systematic review. J. Glob. Antimicrob. 2020, 23, 420–429. [Google Scholar] [CrossRef]
- Jiang, Y.; Wei, Z.; Wang, Y.; Hua, X.; Feng, Y.; Yu, Y. Tracking a hospital outbreak of KPC-producing ST11 Klebsiella pneumoniae with whole genome sequencing. Clin. Microbiol. Infect. 2015, 21, 1001–1007. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.M.; Guo, M.K.; Ke, S.C.; Lin, Y.P.; Li, C.R.; Vy Nguyen, H.T.; Wu, L.T. Emergence and nosocomial spread of ST11 carbapenem-resistant Klebsiella pneumoniae co-producing OXA-48 and KPC-2 in a regional hospital in Taiwan. J. Med. Microbiol. 2018, 67, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Hu, L.; Zhong, Q.; Hang, Y.; Liu, Y.; Hu, X.; Ding, H.; Chen, Y.; Xu, X.; Fang, X.; et al. Dissemination of Klebsiella pneumoniae ST11 isolates with carbapenem resistance in integrated and emergency intensive care units in a Chinese tertiary hospital. J. Med. Microbiol. 2019, 68, 882–889. [Google Scholar] [CrossRef]
- Liu, C.; Yang, P.; Zheng, J.; Yi, J.; Lu, M.; Shen, N. Convergence of two serotypes within the epidemic ST11 KPC-producing Klebsiella pneumoniae creates the “Perfect Storm” in a teaching hospital. BMC Genom. 2022, 23, 693. [Google Scholar] [CrossRef] [PubMed]
- Boyd, S.E.; Livermore, D.M.; Hooper, D.C.; Hope, W.W. Metallo-β-Lactamases: Structure, Function, Epidemiology, Treatment Options, and the Development Pipeline. Antimicrob. Agents Chemother. 2020, 64, e00397-20. [Google Scholar] [CrossRef] [PubMed]
- Abid, F.B.; Tsui, C.; Doi, Y.; Deshmukh, A.; McElheny, C.L.; Bachman, W.C.; Fowler, E.L.; Albishawi, A.; Mushtaq, K.; Ibrahim, E.B.; et al. Molecular characterization of clinical carbapenem-resistant Enterobacterales from Qatar. Eur. J. Clin. Microbiol. 2021, 40, 1779–1785. [Google Scholar] [CrossRef]
- Aris, P.; Robatjazi, S.; Nikkhahi, F.; Amin Marashi, S.M. Molecular mechanisms and prevalence of colistin resistance of Klebsiella pneumoniae in the Middle East region: A review over the last 5 years. J. Glob. Antimicrob. Resist. 2020, 22, 625–630. [Google Scholar] [CrossRef]
- Siri, G.P.; Sithebe, N.P.; Ateba, C.N. Identification of Klebsiella species isolated from Modimola dam (Mafikeng) Northwest Province South Africa. Afr. J. Microbiol. Res. 2011, 5, 3958–3963. [Google Scholar]
- Fang, C.T.; Lai, S.Y.; Yi, W.C.; Hsueh, P.R.; Liu, K.L.; Chang, S.C. Klebsiella pneumoniae genotype K1, An emerging pathogen that causes septic ocular or central nervous system complications from pyogenic liver abscess. Clin. Infect. Dis. 2007, 45, 284–293. [Google Scholar] [CrossRef] [Green Version]
- Vila, A.; Cassata, A.; Pagella, H.; Amadio, C.; Yeh, K.M.; Chang, F.Y.; Siu, L.K. Appearance of Klebsiella pneumoniae liver abscess syndrome in Argentina: Case report and review of molecular mechanisms of pathogenesis. Open Microbiol. J. 2011, 5, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, G.J.; Mendonça, N. Association between antimicrobial resistance and virulence in Escherichia coli. Virulence 2012, 3, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Wyres, K.L.; Lam, M.M.C.; Holt, K.E. Population genomics of Klebsiella pneumoniae. Nat. Rev. Microbiol. 2020, 18, 344–359. [Google Scholar] [CrossRef]
- Al Fadhli, A.H.; Jamal, W.Y.; Rotimi, V.O. Elucidating the virulence genes harboured by carbapenemase- and non-carbapenemase-producing carbapenem-resistant Klebsiella pneumoniae rectal isolates from patients admitted to intensive care units using whole-genome sequencing in Kuwait. J. Med. Microbiol. 2022, 71, 10. [Google Scholar] [CrossRef] [PubMed]
- Roe, C.C.; Vazquez, A.J.; Esposito, E.P.; Zarrilli, R.; Sahl, J.W. Diversity, Virulence, and Antimicrobial Resistance in Isolates From the Newly Emerging Klebsiella pneumoniae ST101 Lineage. Front. Microbiol. 2019, 10, 542. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, P.F.; Lin, T.L.; Yang, F.L.; Wu, M.C.; Pan, Y.J.; Wu, S.H.; Wang, J.T. Lipopolysaccharide O1 antigen contributes to the virulence in Klebsiella pneumoniae causing pyogenic liver abscess. PLoS ONE 2012, 7, e33155. [Google Scholar] [CrossRef]
- Castanheira, M.; Costello, A.J.; Deshpande, L.M.; Jones, R.N. Expansion of clonal complex 258 KPC-2-producing Klebsiella pneumoniae in Latin American hospitals: Report of the SENTRY Antimicrobial Surveillance Program. Antimicrob. Agents Chemother. 2012, 56, 1668–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argimón, S.; David, S.; Underwood, A.; Abrudan, M.; Wheeler, N.E.; Kekre, M.; Abudahab, K.; Yeats, C.A.; Goater, R.; Taylor, B.; et al. Rapid Genomic Characterization and Global Surveillance of Klebsiella Using Pathogenwatch. Clin. Infect. Dis. 2021, 73, S325–S335. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Wick, R.R.; Gorrie, C.; Jenney, A.; Follador, R.; Thomson, N.R.; Holt, K.E. Identification of Klebsiella capsule synthesis loci from whole genome data. Microb. Genom. 2016, 2, e000102. [Google Scholar] [CrossRef] [PubMed]
- Lam, M.M.C.; Wick, R.R.; Watts, S.C.; Cerdeira, L.T.; Wyres, K.L.; Holt, K.E. A genomic surveillance framework and genotyping tool for Klebsiella pneumoniae and its related species complex. Nat. Commun. 2021, 12, 4188. [Google Scholar] [CrossRef] [PubMed]
- Jünemann, S.; Sedlazeck, F.J.; Prior, K.; Albersmeier, A.; John, U.; Kalinowski, J.; Mellmann, A.; Goesmann, A.; von Haeseler, A.; Stoye, J.; et al. Updating benchtop sequencing performance comparison. Nat. Biotechnol. 2013, 31, 294–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Country | Total Number (n) | Carbapenemase Genes | Sequence Types (ST) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NDM | KPC | OXA | IMP | VIM | Dual | None | ST 101 | ST 11 | ST 14 | ST 147 | ST 15 | ST 2096 | ST 231 | ST 307 | ST 45 | ST 48 | ||
| UAE | 98 | 27 | 6 | 30 | 0 | 0 | 21 | 14 | 0 | 6 | 54 | 10 | 7 | 0 | 9 | 0 | 0 | 1 |
| Saudi | 230 | 21 | 1 | 130 | 0 | 2 | 17 | 59 | 15 | 3 | 43 | 8 | 4 | 98 | 0 | 8 | 6 | 7 |
| Qatar | 164 | 38 | 4 | 27 | 0 | 0 | 10 | 85 | 2 | 5 | 4 | 14 | 2 | 2 | 6 | 8 | 6 | 2 |
| Oman | 212 | 77 | 0 | 76 | 0 | 0 | 21 | 38 | 8 | 62 | 3 | 37 | 10 | 0 | 68 | 2 | 0 | 0 |
| Kuwait | 15 | 2 | 0 | 3 | 0 | 0 | 1 | 9 | 0 | 0 | 0 | 0 | 2 | 0 | 8 | 0 | 0 | 0 |
| Bahrain | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 720 | 165 | 11 | 266 | 0 | 2 | 70 | 206 | 25 | 76 | 105 | 69 | 25 | 100 | 91 | 18 | 12 | 10 |
| CRKp % | 71% (514/720) | 32% (165/514) | 2% (11/514) | 51.7% (266/514) | 0 | 0.38% (2/514) | 13.6% (70/514) | 28.6% (206/720) | 88% (22/25) | 94.7% (72/76) | 80% (84/105) | 84% (58/69) | 84% (21/25) | 87% (87/100) | 73.6% (67/91) | 38.8% (7/18) | 50% (6/12) | 40% (4/10) |
| Carbapenem Resistance Genes | Total Isolates (n) | Percentage (%) |
|---|---|---|
| KPC-2 | 10 | 1.94 |
| KPC-2/OXA-232 | 1 | 0.19 |
| KPC-3 | 1 | 0.19 |
| NDM-1 | 159 | 30.93 |
| NDM-1/OXA-162 | 4 | 0.77 |
| NDM-1/OXA-181 | 1 | 0.19 |
| NDM-1/OXA-232 | 31 | 6.03 |
| NDM-1/OXA-48 | 21 | 4.08 |
| NDM-5 | 5 | 0.97 |
| NDM-5/OXA-181 | 3 | 0.58 |
| NDM-5/OXA-232 | 6 | 1.16 |
| NDM-5/OXA-48 | 3 | 0.58 |
| NDM-7 | 1 | 0.19 |
| OXA-162 | 5 | 0.97 |
| OXA-181 | 19 | 3.69 |
| OXA-232 | 143 | 27.82 |
| OXA-48 | 99 | 19.26 |
| VIM-29 | 2 | 0.38 |
| ST-14 | ST-231 | ST-2096 | ST-11 | ST-147 | ST-15 | ST-101 | ST-45 | ||
|---|---|---|---|---|---|---|---|---|---|
| Capsular type | KL5 | 1% | -- | -- | -- | -- | -- | -- | -- |
| KL2 | 61% | -- | -- | -- | -- | 4% | -- | -- | |
| KL10 | -- | -- | -- | -- | 4% | -- | -- | -- | |
| KL14 | -- | -- | -- | 76% | -- | -- | -- | -- | |
| KL15 | -- | -- | -- | 8% | -- | -- | -- | -- | |
| KL16 | 1% | -- | -- | -- | -- | -- | -- | -- | |
| KL17 | -- | -- | -- | -- | -- | -- | 88% | -- | |
| KL19 | -- | -- | -- | -- | -- | 24% | -- | -- | |
| KL24 | -- | -- | -- | 12% | -- | 20% | -- | 8% | |
| KL43 | -- | -- | -- | -- | -- | -- | 50% | ||
| KL47 | -- | -- | -- | 1% | -- | -- | -- | -- | |
| KL48 | -- | -- | -- | -- | -- | 24% | -- | -- | |
| KL50 | 1% | -- | 2% | -- | -- | -- | -- | -- | |
| KL51 | -- | 100% | -- | -- | -- | -- | -- | -- | |
| KL52 | -- | -- | -- | -- | -- | -- | -- | 8% | |
| KL64 | 30% | -- | 93% | 1% | 94% | 4% | 4% | ||
| KL102 | -- | -- | 4% | 8% | |||||
| KL107 | -- | -- | 4% | 1% | 1% | -- | 4% | 8% | |
| KL112 | -- | -- | -- | -- | -- | 16% | -- | -- | |
| KL127 | -- | -- | -- | -- | -- | -- | 8% | ||
| KL135 | 1% | -- | -- | -- | -- | -- | -- | ||
| KL166 | -- | -- | 1% | -- | -- | 4% | -- | -- | |
| O antigen | O1 | 85% | 99% | 96% | -- | -- | 92% | 92% | -- |
| O2a | 10% | 1% | 3% | 13% | 91% | 4% | 4% | 67% | |
| O3 | 1% | -- | -- | 41% | 4% | 4% | -- | 8% | |
| O4 | -- | -- | -- | 8% | -- | -- | -- | ||
| OL101 | -- | -- | -- | 1% | -- | -- | -- | 25% | |
| OL102 | -- | -- | 1% | 1% | 3% | -- | -- | -- | |
| OL104 | -- | -- | -- | 36% | -- | -- | -- | -- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Fadhli, A.H.; Mouftah, S.F.; Jamal, W.Y.; Rotimi, V.O.; Ghazawi, A. Cracking the Code: Unveiling the Diversity of Carbapenem-Resistant Klebsiella pneumoniae Clones in the Arabian Peninsula through Genomic Surveillance. Antibiotics 2023, 12, 1081. https://doi.org/10.3390/antibiotics12071081
Al Fadhli AH, Mouftah SF, Jamal WY, Rotimi VO, Ghazawi A. Cracking the Code: Unveiling the Diversity of Carbapenem-Resistant Klebsiella pneumoniae Clones in the Arabian Peninsula through Genomic Surveillance. Antibiotics. 2023; 12(7):1081. https://doi.org/10.3390/antibiotics12071081
Chicago/Turabian StyleAl Fadhli, Amani H, Shaimaa F. Mouftah, Wafaa Y. Jamal, Vincent O. Rotimi, and Akela Ghazawi. 2023. "Cracking the Code: Unveiling the Diversity of Carbapenem-Resistant Klebsiella pneumoniae Clones in the Arabian Peninsula through Genomic Surveillance" Antibiotics 12, no. 7: 1081. https://doi.org/10.3390/antibiotics12071081