
Insight into the Clonal Lineage and Antimicrobial Resistance of Staphylococcus aureus from Vascular Access Infections before and during the COVID-19 Pandemic

Abstract
:1. Introduction
2. Results
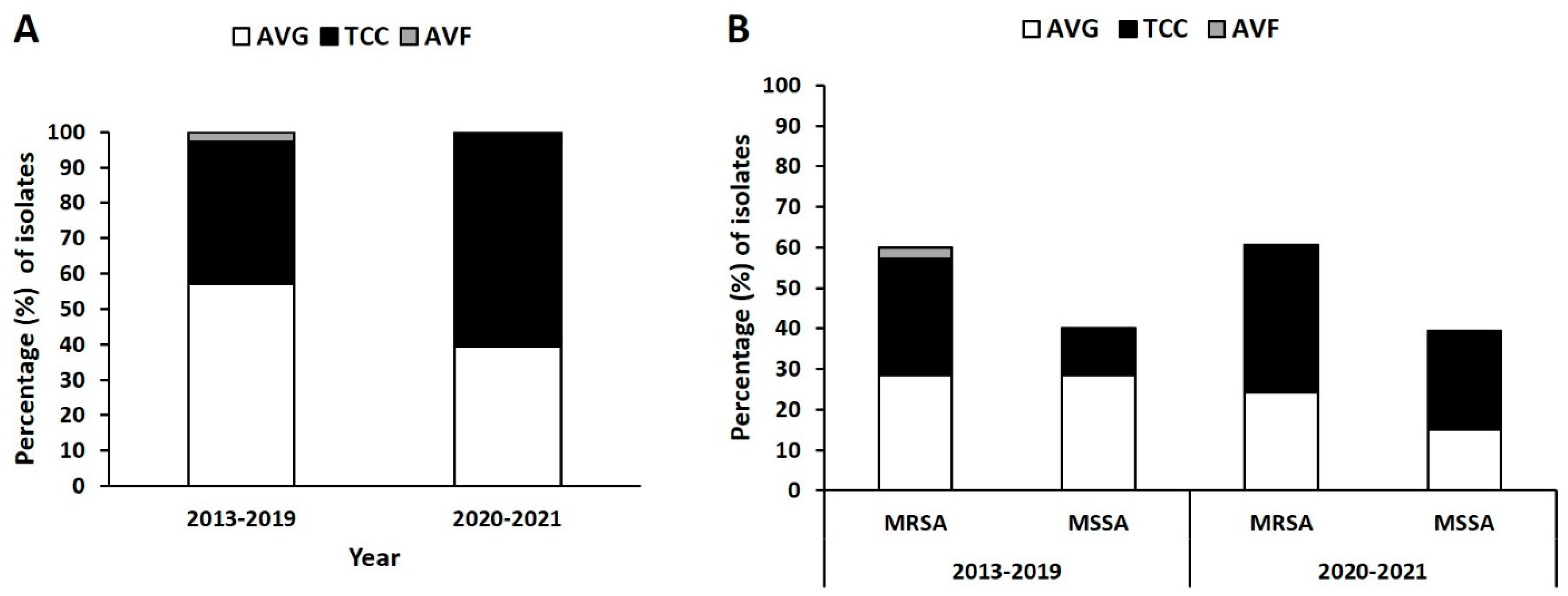
2.1. Distribution of Isolates from Different VAI Types
2.2. Distribution of SCCmec Type in MRSA Isolates
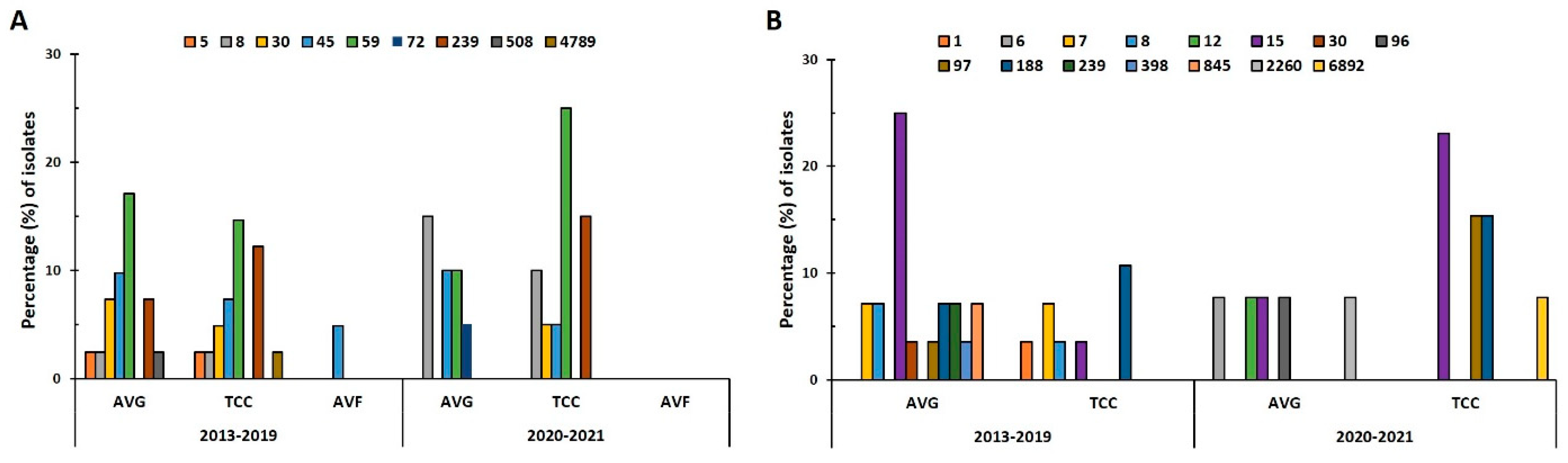
2.3. Distribution of Sequence Types (STs) in VAI Isolates
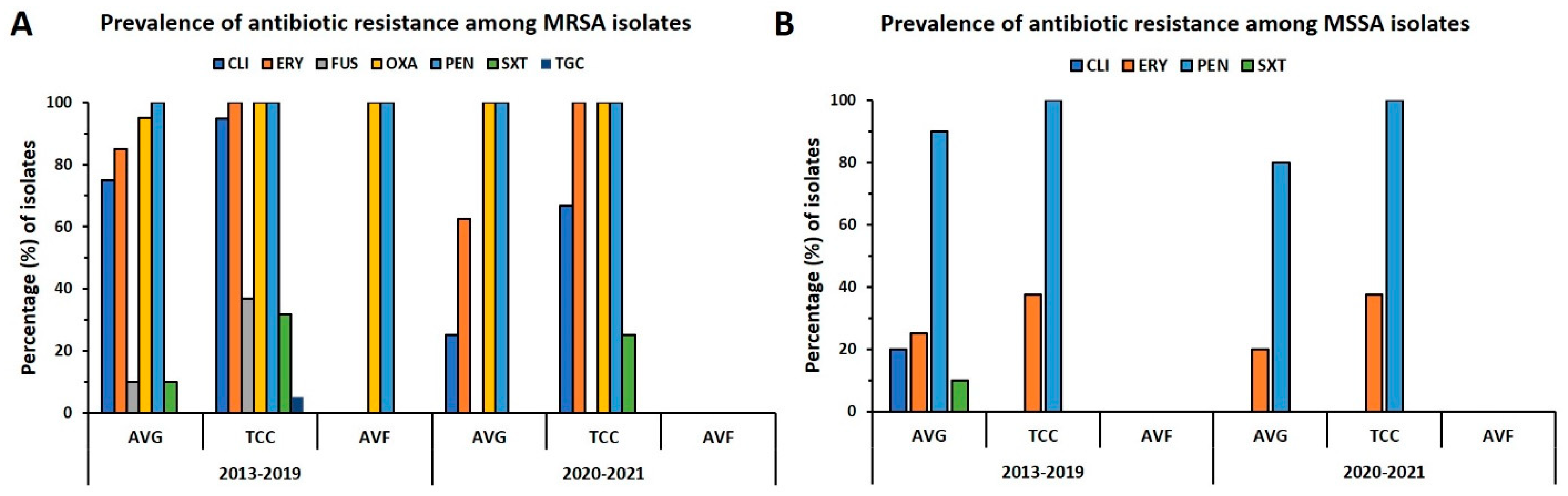
2.4. Prevalence of Antibiotic Resistance in VAI Isolates
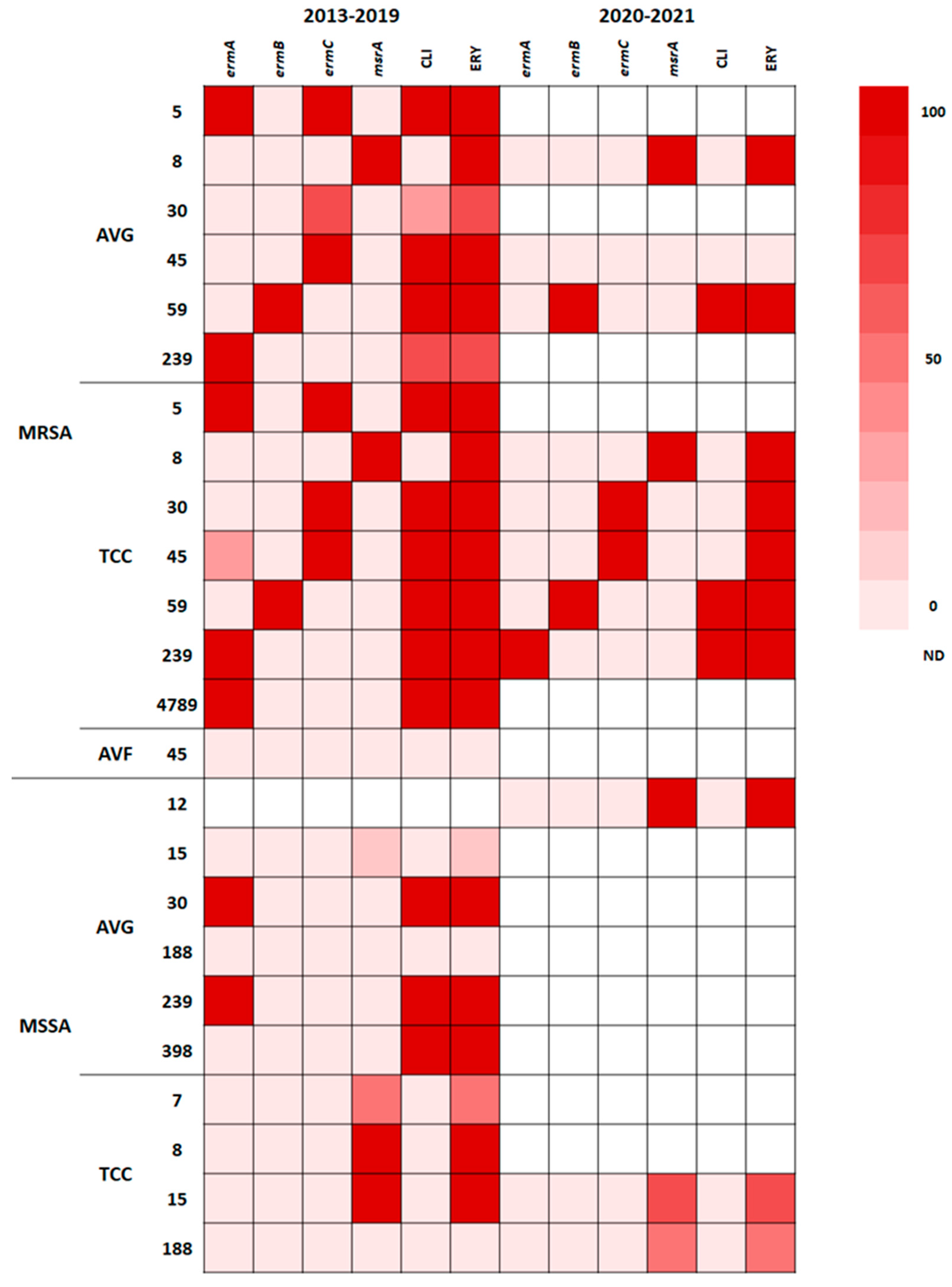
2.5. Correlation of STs with Antibiotic Resistance
2.6. Correlation of Resistance Genotype with Phenotype
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Study Setting, Bacterial Isolate Collection, and Identification
4.3. Antimicrobial Susceptibility Testing
4.4. Molecular Characterization and Antibiotic-Resistant Gene Detection
4.4.1. Identification of Methicillin-Resistant S. aureus and Staphylococcal Chromosomal Cassette mec (SCCmec) Type
4.4.2. Molecular Typing
4.4.3. Erythromycin-Resistant Gene Detection
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ogston, A. Micrococcus poisoning. J. Anat. Physiol. 1882, 16 Pt 4, 526–567. [Google Scholar] [PubMed]
- Kluytmans, J.; van Belkum, A.; Verbrugh, H. Nasal carriage of Staphylococcus aureus: Epidemiology, underlying mechanisms, and associated risks. Clin. Microbiol. Rev. 1997, 10, 505–520. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F. The changing epidemiology of Staphylococcus aureus? Emerg. Infect. Dis. 2001, 7, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.L.; Chang, S.C.; Pan, H.J.; Hsueh, P.R.; Yang, L.S.; Ho, S.W.; Luh, K.T. Longitudinal analysis of methicillin-resistant Staphylococcus aureus isolates at a teaching hospital in Taiwan. J. Formos. Med. Assoc. 1999, 98, 426–432. [Google Scholar]
- Huang, Y.-C.; Chen, C.-J. Community-associated meticillin-resistant Staphylococcus aureus in children in Taiwan, 2000s. Int. J. Antimicrob. Agents 2011, 38, 2–8. [Google Scholar] [CrossRef]
- Chen, C.-J.; Huang, Y.-C.; Su, L.-H.; Wu, T.-L.; Huang, S.-H.; Chien, C.-C.; Chen, P.-Y.; Lu, M.-C.; Ko, W.-C. Molecular epidemiology and antimicrobial resistance of methicillin-resistant Staphylococcus aureus bloodstream isolates in Taiwan, 2010. PLoS ONE 2014, 9, e101184. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-Y.; Fang, Y.-P.; Chang, Y.-F.; Wu, T.-H.; Yang, Y.-Y.; Huang, Y.-C. Comparison of molecular epidemiology of bloodstream methicillin-resistant Staphylococcus aureus isolates between a new and an old hospital in central Taiwan. Int. J. Infect. Dis. 2019, 79, 162–168. [Google Scholar] [CrossRef] [Green Version]
- Emerging Infections Program, Healthcare-Associated Infections—Community Interface Surveillance Report, Invasive Staphylococcus aureus, 2019. Available online: https://www.cdc.gov/hai/eip/pdf/2019-MRSA-Report-508.pdf (accessed on 10 May 2023).
- Vandecasteele, S.J.; Boelaert, J.R.; De Vriese, A.S. Staphylococcus aureus infections in hemodialysis: What a nephrologist should know. Clin. J. Am. Soc. Nephrol. 2009, 4, 1388–1400. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.-Y.; Tu, H.-P.; Chen, T.-C.; Shen, M.-C.; Chiu, Y.-W.; Chen, Y.-H.; Lu, P.-L. Association of bacterial genotypes and epidemiological features with treatment failure in hemodialysis patients with methicillin-resistant Staphylococcus aureus bacteremia. PLoS ONE 2018, 13, e0198486. [Google Scholar] [CrossRef]
- Control, C.F.D. & Prevention. Invasive methicillin-resistant Staphylococcus aureus infections among dialysis pa-tients--United States. MMWR Morb. Mortal. Wkly. Rep. 2005, 56, 197–199. [Google Scholar]
- Akoh, J.A. Vascular access infections: Epidemiology, diagnosis, and management. Curr. Infect. Dis. Rep. 2011, 13, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.-C.; Chen, S.-Y.; Ko, W.-C.; Hsueh, P.-R. Increased antimicrobial resistance during the COVID-19 pandemic. Int. J. Antimicrob. Agents 2021, 57, 106324. [Google Scholar] [CrossRef]
- Weisblum, B. Erythromycin resistance by ribosome modification. Antimicrob. Agents Chemother. 1995, 39, 577–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoro, D.; Benedetto, F.; Mondello, P.; Pipitò, N.; Barillà, D.; Spinelli, F.; Ricciardi, C.A.; Cernaro, V.; Buemi, M. Vascular access for hemodialysis: Current perspectives. Int. J. Nephrol. Renovasc. Dis. 2014, 7, 281–294. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-J.; Huang, Y.-C. New epidemiology of Staphylococcus aureus infection in Asia. Clin. Microbiol. Infect. 2014, 20, 605–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Zhou, W.; Zhan, Q.; Zheng, B.; Chen, Y.; Luo, Q.; Shen, P.; Xiao, Y. Genomic epidemiology and characterization of methicillin-resistant Staphylococcus aureus from bloodstream infections in China. mSystems 2021, 6, e0083721. [Google Scholar] [CrossRef]
- Zhang, H.; Cao, J.; He, Z.; Zong, X.; Sun, B. Molecular epidemiology of Staphylococcus aureus in a Tertiary Hospital in Anhui, China: ST59 Remains a Serious Threat. Infect. Drug Resist. 2023, 16, 961–976. [Google Scholar] [CrossRef]
- Tsao, F.-Y.; Kou, H.-W.; Huang, Y.-C. Dissemination of methicillin-resistant Staphylococcus aureus sequence type 45 among nursing home residents and staff in Taiwan. Clin. Microbiol. Infect. 2015, 21, 451–458. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-C.; Chang, C.-J.; Lin, Y.-T.; Huang, K.-Y.A.; Chen, C.-J. A longitudinal survey of methicillin-resistant Staphylococcus aureus carriage in nursing homes and the long-term care facility in Taiwan. J. Microbiol. Immunol. Infect. 2022, 55, 853–859. [Google Scholar] [CrossRef]
- Wu, T.-H.; Lee, C.-Y.; Yang, H.-J.; Fang, Y.-P.; Chang, Y.-F.; Tzeng, S.-L.; Lu, M.-C. Prevalence and molecular characteristics of methicillin-resistant Staphylococcus aureus among nasal carriage strains isolated from emergency department patients and healthcare workers in central Taiwan. J. Microbiol. Immunol. Infect. 2019, 52, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Diekema, D.J.; Richter, S.S.; Heilmann, K.P.; Dohrn, C.L.; Riahi, F.; Tendolkar, S.; McDanel, J.S.; Doern, G.V. Continued emergence of USA300 methicillin-resistant Staphylococcus aureus in the United States: Results from a nationwide surveillance study. Infect. Control Hosp. Epidemiol. 2014, 35, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Carrel, M.; Perencevich, E.N.; David, M.Z. USA300 methicillin-resistant Staphylococcus aureus, United States, 2000–2013. Emerg. Infect. Dis. 2015, 21, 1973–1980. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Song, E.H.; Park, S.Y.; Lee, S.-R.; Park, S.-J.; Sung, H.; Kim, M.-N.; Kim, S.-H.; Lee, S.-O.; Choi, S.-H.; et al. Emergence of panton-valentine leucocidin-positive ST8-methicillin-resistant Staphylococcus aureus (USA300 Clone) in Korea causing healthcare-associated and hospital-acquired bacteraemia. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1323–1329. [Google Scholar] [CrossRef]
- Nagao, M. First outbreak of methicillin-resistant Staphylococcus aureus USA300 harboring the Panton-Valentine leukocidin genes among Japanese health care workers and hospitalized patients. Am. J. Infect. Control 2010, 38, e37–e39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.-C.; Chen, C.-J.; Kuo, C.-C.; Lu, M.-C. Emergence, transmission and phylogeny of meticillin-resistant Staphylococcus aureus sequence type 8 (USA300) in Taiwan. J. Hosp. Infect. 2018, 100, 355–358. [Google Scholar] [CrossRef]
- Chen, P.-Y.; Chuang, Y.-C.; Wang, J.-T.; Sheng, W.-H.; Chen, Y.-C.; Chang, S.-C. Sequence type 8 as an emerging clone of methicillin-resistant Staphylococcus aureus causing bloodstream infections in Taiwan: Evolving molecular epidemiology of MRSA bacteraemia. Emerg. Microbes Infect. 2021, 10, 1908–1918. [Google Scholar] [CrossRef]
- Chuang, Y.Y.; Huang, Y.C. Molecular epidemiology of community-associated meticillin-resistant Staphylococcus aureus in Asia. Lancet Infect. Dis. 2013, 13, 698–708. [Google Scholar] [CrossRef]
- Huang, Y.C.; Chen, C.J. USA300 (sequence type 8) has become a major clone of methicillin-resistant Staphy-lococcus aureus in northern Taiwan. Int. J. Antimicrob. Agents 2022, 59, 106534. [Google Scholar] [CrossRef]
- Kaku, N.; Sasaki, D.; Ota, K.; Miyazaki, T.; Yanagihara, K. Changing molecular epidemiology and characteristics of MRSA isolated from bloodstream infections: Nationwide surveillance in Japan in 2019. J. Antimicrob. Chemother. 2022, 77, 2130–2141. [Google Scholar] [CrossRef]
- Aktas, Z.; Aridogan, A.; Kayacan, C.B.; Aydin, D. Resistance to macrolide, lincosamide and streptogramin antibiotics in staphylococci isolated in Istanbul, Turkey. J. Microbiol. 2007, 45, 286–290. [Google Scholar] [PubMed]
- Wang, C.-C.; Lo, W.-T.; Chu, M.-L.; Siu, L. Epidemiological typing of community-acquired methicillin-resistant Staphylococcus aureus isolates from children in Taiwan. Clin. Infect. Dis. 2004, 39, 481–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zhuang, H.; Ji, S.; Sun, L.; Zhao, F.; Wu, D.; Shen, P.; Jiang, Y.; Yu, Y.; Chen, Y. Distribution of erm genes among MRSA isolates with resistance to clindamycin in a Chinese teaching hospital. Infect. Genet. Evol. 2021, 96, 105127. [Google Scholar] [CrossRef] [PubMed]
- CLSI. CLSI Document M100-S27; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Tseng, Y.H.; Wong, M.Y.; Huang, T.Y.; Lin, B.S.; Tung, C.W.; Huang, Y.K. Molecular characterization of clinical isolates from vascular access infection: A single-institution study. Microbiologyopen 2020, 9, e1126. [Google Scholar] [CrossRef]
- Pournajaf, A.; Ardebili, A.; Goudarzi, L.; Khodabandeh, M.; Narimani, T.; Abbaszadeh, H. PCR-based identification of methicillin-resistant Staphylococcus aureus strains and their antibiotic resistance profiles. Asian Pac. J. Trop. Biomed. 2014, 4 (Suppl. S1), S293–S297. [Google Scholar] [CrossRef] [Green Version]
- Boye, K.; Bartels, M.D.; Andersen, I.S.; Møller, J.A.; Westh, H. A new multiplex PCR for easy screening of methicillin-resistant Staphylococcus aureus SCCmec types I-V. Clin. Microbiol. Infect. 2007, 13, 725–727. [Google Scholar] [CrossRef] [Green Version]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Ruimy, R.; Armand-Lefevre, L.; Barbier, F.; Ruppé, E.; Cocojaru, R.; Mesli, Y.; Maiga, A.; Benkalfat, M.; Benchouk, S.; Hassaine, H.; et al. Comparisons between geographically diverse samples of carried Staphylococcus aureus. J. Bacteriol. 2009, 191, 5577–5583. [Google Scholar] [CrossRef] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb Software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Duran, N.; Ozer, B.; Duran, G.G.; Onlen, Y.; Demir, C. Antibiotic resistance genes & susceptibility patterns in staphylococci. Indian J. Med. Res. 2012, 135, 389–396. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ST | 2013–2019 | 2020–2021 | ||||
|---|---|---|---|---|---|---|
| AVG | TCC | AVF | AVG | TCC | AVF | |
| MRSA | ||||||
| 5 | CLI, ERY, OXA, PEN (1) | CLI, ERY, FUS, OXA, PEN (1) | ||||
| 8 | ERY, OXA, PEN (1) | ERY, OXA, PEN (1) | ERY, OXA, PEN (3) | ERY, OXA, PEN (2) | ||
| 30 | CLI, ERY, OXA, PEN (1) | CLI, ERY, OXA, PEN (2) | ERY, OXA, PEN (1) | |||
| ERY, OXA, PEN (1) | ||||||
| OXA, PEN (1) | ||||||
| 45 | CLI, ERY, FUS, OXA, PEN (2) | CLI, ERY, FUS, OXA, PEN (3) | OXA, PEN (2) | OXA, PEN (2) | ERY, OXA, PEN (1) | |
| CLI, ERY, OXA, PEN (2) | ||||||
| 59 | CLI, ERY, OXA, PEN (7) | CLI, ERY, OXA, PEN (6) | CLI, ERY, OXA, PEN (2) | CLI, ERY, OXA, PEN (5) | ||
| 72 | OXA, PEN (1) | |||||
| 239 | CLI, ERY, OXA, PEN, SXT (2) | CLI, ERY, FUS, OXA, PEN, SXT (2) | CLI, ERY, OXA, PEN, SXT (3) | |||
| PEN (1) | CLI, ERY, OXA, PEN, SXT (2) | |||||
| CLI, ERY, OXA, PEN, SXT, TGC (1) | ||||||
| 508 | OXA, PEN (1) | |||||
| 4789 | CLI, ERY, FUS, OXA, PEN, SXT (1) | |||||
| MSSA | ||||||
| 1 | PEN (1) | |||||
| 6 | PEN (1) | |||||
| 7 | PEN (2) | ERY, PEN (1) | ||||
| PEN (1) | ||||||
| 8 | PEN (2) | ERY, PEN (1) | ||||
| 12 | ERY, PEN (1) | |||||
| 15 | ERY, PEN (1) | ERY, PEN (1) | PEN (1) | ERY, PEN (2) | ||
| PEN (6) | PEN (1) | |||||
| 30 | CLI, ERY, PEN (1) | |||||
| 96 | PEN (1) | |||||
| 97 | PEN (1) | PEN (2) | ||||
| 188 | PEN (1) | PEN (3) | ERY, PEN (1) | |||
| NONE (1) | PEN (1) | |||||
| 239 | CLI, ERY, PEN, SXT (2) | |||||
| 398 | CLI, ERY (1) | |||||
| 845 | PEN (2) | |||||
| 2260 | NONE (1) | |||||
| 6892 | PEN (1) | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kao, C.-C.; Lai, C.-H.; Wong, M.-Y.; Huang, T.-Y.; Tseng, Y.-H.; Lu, C.-H.; Lin, C.-C.; Huang, Y.-K. Insight into the Clonal Lineage and Antimicrobial Resistance of Staphylococcus aureus from Vascular Access Infections before and during the COVID-19 Pandemic. Antibiotics 2023, 12, 1070. https://doi.org/10.3390/antibiotics12061070
Kao C-C, Lai C-H, Wong M-Y, Huang T-Y, Tseng Y-H, Lu C-H, Lin C-C, Huang Y-K. Insight into the Clonal Lineage and Antimicrobial Resistance of Staphylococcus aureus from Vascular Access Infections before and during the COVID-19 Pandemic. Antibiotics. 2023; 12(6):1070. https://doi.org/10.3390/antibiotics12061070
Chicago/Turabian StyleKao, Chih-Chen, Chi-Hsiang Lai, Min-Yi Wong, Tsung-Yu Huang, Yuan-Hsi Tseng, Chu-Hsueh Lu, Chien-Chao Lin, and Yao-Kuang Huang. 2023. "Insight into the Clonal Lineage and Antimicrobial Resistance of Staphylococcus aureus from Vascular Access Infections before and during the COVID-19 Pandemic" Antibiotics 12, no. 6: 1070. https://doi.org/10.3390/antibiotics12061070