
Whole Genome Sequencing and Phenotypic Analysis of Antibiotic Resistance in Filifactor alocis Isolates
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Identification of Isolates and Sequencing Analysis
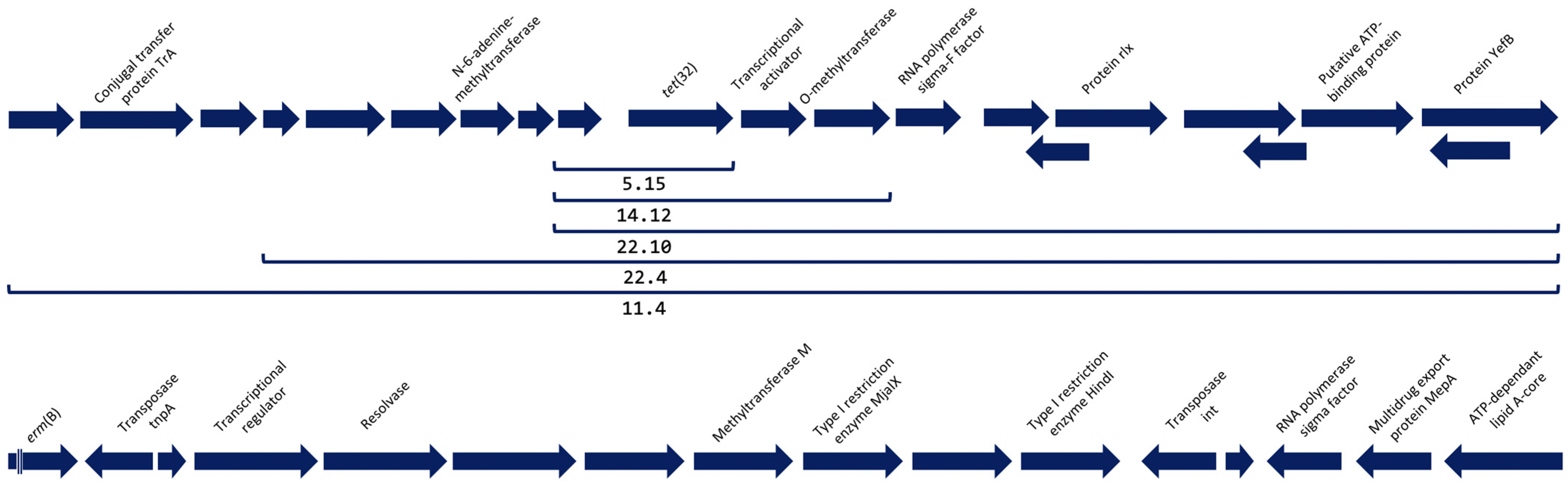
2.2. Detection and Analysis of Antimicrobial Resistance Genes
2.3. Antimicrobial Resistance Testing
3. Discussion
4. Material & Methods
4.1. Sample Collection and Culturing Conditions
4.2. DNA Isolation and Polymerase Chain Reaction-Based Characterization
4.3. Antibiotic Susceptibility Testing
4.4. Whole-Genome Sequencing
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jalava, J.; Eerola, E. Phylogenetic analysis of Fusobacterium alocis and Fusobacterium sulci based on 16S rRNA gene sequences: Proposal of Filifactor alocis (Cato, Moore and Moore) comb. nov. and Eubacterium sulci (Cato, Moore and Moore) comb. nov. Int. J. Syst. Evol. Microbiol. 1999, 49, 1375–1379. [Google Scholar] [CrossRef] [Green Version]
- Aja, E.; Mangar, M.; Fletcher, H.M.; Mishra, A. Filifactor alocis: Recent Insights and Advances. J. Dent. Res. 2021, 100, 790–797. [Google Scholar] [CrossRef]
- Cato, E.P.; Moore, L.V.H.; Moore, W.E.C. Fusobacterium alocis sp. nov. and Fusobacterium sulci sp. nov. from the Human Gingival Sulcus. Int. J. Syst. Evol. Microbiol. 1985, 35, 475–477. [Google Scholar] [CrossRef] [Green Version]
- Hutter, G.; Schlagenhauf, U.; Valenza, G.; Horn, M.; Burgemeister, S.; Claus, H.; Vogel, U. Molecular analysis of bacteria in periodontitis: Evaluation of clone libraries, novel phylotypes and putative pathogens. Microbiology 2003, 149, 67–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jr, J.F.S.; Rôças, I.N. Detection of Filifactor alocis in endodontic infections associated with different forms of periradicular diseases. Oral Microbiol. Immunol. 2003, 18, 263–265. [Google Scholar] [CrossRef]
- Gomes, B.P.; Jacinto, R.C.; Pinheiro, E.; Sousa, E.L.; Zaia, A.A.; Ferraz, C.; Souza-Filho, F.J. Molecular Analysis of Filifactor alocis, Tannerella forsythia, and Treponema denticola Associated with Primary Endodontic Infections and Failed Endodontic Treatment. J. Endod. 2006, 32, 937–940. [Google Scholar] [CrossRef]
- Kumar, P.S.; Leys, E.J.; Bryk, J.M.; Martinez, F.J.; Moeschberger, M.L.; Griffen, A.L. Changes in Periodontal Health Status Are Associated with Bacterial Community Shifts as Assessed by Quantitative 16S Cloning and Sequencing. J. Clin. Microbiol. 2006, 44, 3665–3673. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Liu, Y.; Zhang, M.; Wang, G.; Qi, Z.; Bridgewater, L.; Zhao, L.; Tang, Z.; Pang, X. A Filifactor alocis-centered co-occurrence group associates with periodontitis across different oral habitats. Sci. Rep. 2015, 5, srep09053. [Google Scholar] [CrossRef] [Green Version]
- Griffen, A.L.; Beall, C.J.; Campbell, J.H.; Firestone, N.D.; Kumar, P.S.; Yang, Z.K.; Podar, M.; Leys, E.J. Distinct and complex bacterial profiles in human periodontitis and health revealed by 16S pyrosequencing. ISME J. 2011, 6, 1176–1185. [Google Scholar] [CrossRef] [Green Version]
- Moffatt, C.; Whitmore, S.; Griffen, A.; Leys, E.; Lamont, R. Filifactor alocis interactions with gingival epithelial cells. Mol. Oral Microbiol. 2011, 26, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Aruni, A.W.; Zhang, K.; Dou, Y.; Fletcher, H. Proteome Analysis of Coinfection of Epithelial Cells with Filifactor alocis and Porphyromonas gingivalis Shows Modulation of Pathogen and Host Regulatory Pathways. Infect. Immun. 2014, 82, 3261–3274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miralda, I.; Vashishta, A.; Rogers, M.N.; Lamont, R.J.; Uriarte, S.M. The emerging oral pathogen, Filifactor alocis, extends the functional lifespan of human neutrophils. Mol. Microbiol. 2022, 117, 1340–1351. [Google Scholar] [CrossRef]
- Bao, K.; Claesson, R.; Belibasakis, G.N.; Oscarsson, J. Extracellular Vesicle Subproteome Differences among Filifactor alocis Clinical Isolates. Microorganisms 2022, 10, 1826. [Google Scholar] [CrossRef]
- Mishra, A.; Aja, E.; Fletcher, H.M. Role of Superoxide Reductase FA796 in Oxidative Stress Resistance in Filifactor alocis. Sci. Rep. 2020, 10, 9178. [Google Scholar] [CrossRef] [PubMed]
- Aruni, A.W.; Roy, F.; Fletcher, H.M. Filifactor alocis Has Virulence Attributes That Can Enhance Its Persistence under Oxidative Stress Conditions and Mediate Invasion of Epithelial Cells by Porphyromonas gingivalis. Infect. Immun. 2011, 79, 3872–3886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aruni, A.W.; Roy, F.; Sandberg, L.; Fletcher, H.M. Proteome variation among Filifactor alocis strains. Proteomics 2012, 12, 3343–3364. [Google Scholar] [CrossRef] [Green Version]
- Schlafer, S.; Riep, B.; Griffen, A.L.; Petrich, A.; Hübner, J.; Berning, M.; Friedmann, A.; Göbel, U.B.; Moter, A. Filifactor alocis—involvement in periodontal biofilms. BMC Microbiol. 2010, 10, 66. [Google Scholar] [CrossRef] [Green Version]
- Gray, R.; Vidwans, M. Mixed anaerobic thoracic empyema: The first report of Filifactor alocis causing extra-oral disease. New Microbes New Infect. 2019, 29, 100528. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, Y.; Wan, H. Case report:Multiple abscesses caused by Porphyromonas gingivalis diagnosed by metagenomic next-generation sequencing. Front. Med. 2023, 9, 1089863. [Google Scholar] [CrossRef]
- Yusuf, K.; Sampath, V.; Umar, S. Bacterial Infections and Cancer: Exploring This Association And Its Implications for Cancer Patients. Int. J. Mol. Sci. 2023, 24, 3110. [Google Scholar] [CrossRef]
- Mesturino, M.A.; Bitetti, C.; Clemente, A.; Krzysztofiak, A.; Lancella, L.; Lombardi, R.; Cursi, L.; Boccuzzi, E.; Musolino, A.M.; Villani, A. Aggregatibacter actinomycetemcomitans infection in a 15-year-old boy with pulmonary empyema: A case report and review of literature. Ital. J. Pediatr. 2023, 49, 42. [Google Scholar] [CrossRef] [PubMed]
- Inubushi, J.; Liang, K. Update on minocycline in vitro activity against odontogenic bacteria. J. Infect. Chemother. 2020, 26, 1334–1337. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, M.I.; Römling, U.; Nadeem, F.; Bilal, H.M.; Zafar, M.; Jahan, H.; Ur-Rahman, A. Innovative Strategies to Overcome Antimicrobial Resistance and Tolerance. Microorganisms 2022, 11, 16. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 13.0. EFSA Journal. Available online: http://www.eucast.org (accessed on 20 February 2023).
- Àlvarez, G.; Arredondo, A.; Isabal, S.; Teughels, W.; Laleman, I.; Contreras, M.J.; Isbej, L.; Huapaya, E.; Mendoza, G.; Mor, C.; et al. Association of nine pathobionts with periodontitis in four South American and European countries. J. Oral Microbiol. 2023, 15, 2188630. [Google Scholar] [CrossRef] [PubMed]
- Lafaurie, G.I.; Neuta, Y.; Ríos, R.; Pacheco-Montealegre, M.; Pianeta, R.; Castillo, D.M.; Herrera, D.; Reyes, J.; Diaz, L.; Castillo, Y.; et al. Differences in the subgingival microbiome according to stage of periodontitis: A comparison of two geographic regions. PLoS ONE 2022, 17, e0273523. [Google Scholar] [CrossRef]
- Thaker, M.; Spanogiannopoulos, P.; Wright, G.D. The tetracycline resistome. Cell. Mol. Life Sci. 2010, 67, 419–431. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, X.; Zong, Z. Genome sequence and virulence factors of a group G Streptococcus dysgalactiae subsp. equisimilis strain with a new element carrying erm(B). Sci. Rep. 2016, 6, 20389. [Google Scholar] [CrossRef] [Green Version]
- Arredondo, A.; Blanc, V.; Mor, C.; Nart, J.; León, R. Tetracycline and multidrug resistance in the oral microbiota: Differences between healthy subjects and patients with periodontitis in Spain. J. Oral Microbiol. 2021, 13, 1847431. [Google Scholar] [CrossRef]
- Warburton, P.; Roberts, A.P.; Allan, E.; Seville, L.; Lancaster, H.; Mullany, P. Characterization of tet (32) Genes from the Oral Metagenome. Antimicrob. Agents Chemother. 2009, 53, 273–276. [Google Scholar] [CrossRef] [Green Version]
- Speer, B.S.; Shoemaker, N.B.; Salyers, A.A. Bacterial Resistance to Tetracycline: Mechanisms, Transfer, and Clinical Significance. Clin. Microbiol. Rev. 1992, 5, 387–399. [Google Scholar] [CrossRef]
- Roberts, M.C. Resistance to Tetracycline, Macrolide-Lincosamide-Streptogramin, Trimethoprim, and Sulfonamide Drug Classes. Appl. Biochem. Biotechnol. Part B Mol. Biotechnol. 2002, 20, 261–284. [Google Scholar] [CrossRef] [PubMed]
- Weisblum, B. Erythromycin resistance by ribosome modification. Antimicrob. Agents Chemother. 1995, 39, 577–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciric, L.; Ellatif, M.; Sharma, P.; Patel, R.; Song, X.; Mullany, P.; Roberts, A.P. Tn916-like elements from human, oral, commensal streptococci possess a variety of antibiotic and antiseptic resistance genes. Int. J. Antimicrob. Agents 2012, 39, 360–361. [Google Scholar] [CrossRef] [PubMed]
- Arredondo, A.; Blanc, V.; Mor, C.; Nart, J.; León, R. Azithromycin and erythromycin susceptibility and macrolide resistance genes in Prevotella from patients with periodontal disease. Oral Dis. 2019, 25, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.C. Location of the Various Genes Table. Available online: http://faculty.washington.edu/marilynr (accessed on 20 February 2023).
- Tansirichaiya, S.; Rahman, A.; Roberts, A.P. The Transposon Registry. Mob. DNA 2019, 10, 40. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC); European Food Safety Authority (EFSA); European Medicines Agency (EMA). ECDC/EFSA/EMA Second Joint Report on the Integrated Analysis of the Consumption of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Bacteria from Humans and Food-Producing Animals. EFSA J. 2017, 15, e04872. [Google Scholar] [CrossRef]
- Oscarsson, J.; Claesson, R.; Bao, K.; Brundin, M.; Belibasakis, G. Phylogenetic Analysis of Filifactor alocis Strains Isolated from Several Oral Infections Identified a Novel RTX Toxin, FtxA. Toxins 2020, 12, 687. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E.; et al. AMRFinderPlus and the Reference Gene Catalog facilitate examination of the genomic links among antimicrobial resistance, stress response, and virulence. Sci. Rep. 2021, 11, 12728. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Isolate | Gene | Class of Antimicrobial | Coverage of Sequence (%) | Identity of Sequence (%) |
|---|---|---|---|---|
| 5.15 | tet(32) | Tetracycline | 68.39 | 99.54 |
| 11.40 | tet(32) | Tetracycline | 100 | 100 |
| 14.12 | tet(32) | Tetracycline | 100 | 100 |
| 22.10 | tet(32) | Tetracycline | 100 | 100 |
| 22.40 | tet(32) | Tetracycline | 100 | 100 |
| 30.27 | erm(B) | Macrolide | 72.24 | 100 |
| Isolate | AMC | AMX | AZM | CDM | CIP | DOX | MIN | MTZ | TET |
|---|---|---|---|---|---|---|---|---|---|
| 5.15 | <0.016 | <0.016 | 1 | 0.047 | 0.008 | 0.047 | 0.032 | <0.016 | 0.25 |
| 8.25 | 0.016 | 0.023 | 0.032 | 0.016 | 0.023 | 0.023 | 0.016 | 0.016 | 0.016 |
| 11.40 | 0.032 | 0.023 | 0.047 | <0.016 | 0.032 | 0.125 | 0.094 | <0.016 | 0.75 |
| 14.12 | 0.25 | <0.016 | 0.032 | <0.016 | 0.012 | 0.5 | 0.125 | <0.016 | 0.75 |
| 22.10 | 0.016 | 0.064 | 0.047 | 0.016 | 0.023 | 0.25 | 0.19 | 0.016 | 1 |
| 22.40 | 0.023 | 0.032 | 0.032 | <0.016 | 0.016 | 0.25 | 0.032 | <0.016 | 0.75 |
| 30.27 | <0.016 | <0.016 | >256 | 1 | 0.125 | <0.016 | < 0.016 | <0.016 | 0.016 |
| 48B | 0.032 | 0.016 | 0.064 | <0.016 | 0.047 | 0.047 | < 0.016 | 0.016 | 0.016 |
| ATCC 35896 | 0.064 | 0.032 | 0.032 | <0.016 | 0.023 | 1 | 1 | 0.016 | 2 |
| Range | <0.016–0.094 | <0.016–0.25 | 0.032–>256 | <0.016–0.19 | 0.08–1 | <0.016–0.75 | <0.016–0.19 | <0.016–>256 | 0.016–1 |
| MIC50 | 0.016 | 0.023 | 0.047 | <0.016 | 0.023 | 0.047 | 0.032 | <0.016 | 0.25 |
| MIC90 | 0.032 | 0.064 | 1 | 0.047 | 0.047 | 0.25 | 0.125 | 0.016 | 0.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero-Martínez, R.; Maher, A.; Àlvarez, G.; Figueiredo, R.; León, R.; Arredondo, A. Whole Genome Sequencing and Phenotypic Analysis of Antibiotic Resistance in Filifactor alocis Isolates. Antibiotics 2023, 12, 1059. https://doi.org/10.3390/antibiotics12061059
Romero-Martínez R, Maher A, Àlvarez G, Figueiredo R, León R, Arredondo A. Whole Genome Sequencing and Phenotypic Analysis of Antibiotic Resistance in Filifactor alocis Isolates. Antibiotics. 2023; 12(6):1059. https://doi.org/10.3390/antibiotics12061059
Chicago/Turabian StyleRomero-Martínez, Rosa, Anushiravan Maher, Gerard Àlvarez, Rui Figueiredo, Rubén León, and Alexandre Arredondo. 2023. "Whole Genome Sequencing and Phenotypic Analysis of Antibiotic Resistance in Filifactor alocis Isolates" Antibiotics 12, no. 6: 1059. https://doi.org/10.3390/antibiotics12061059