
Use of Probiotics to Control Biofilm Formation in Food Industries

 , , and
, , and
Abstract
:
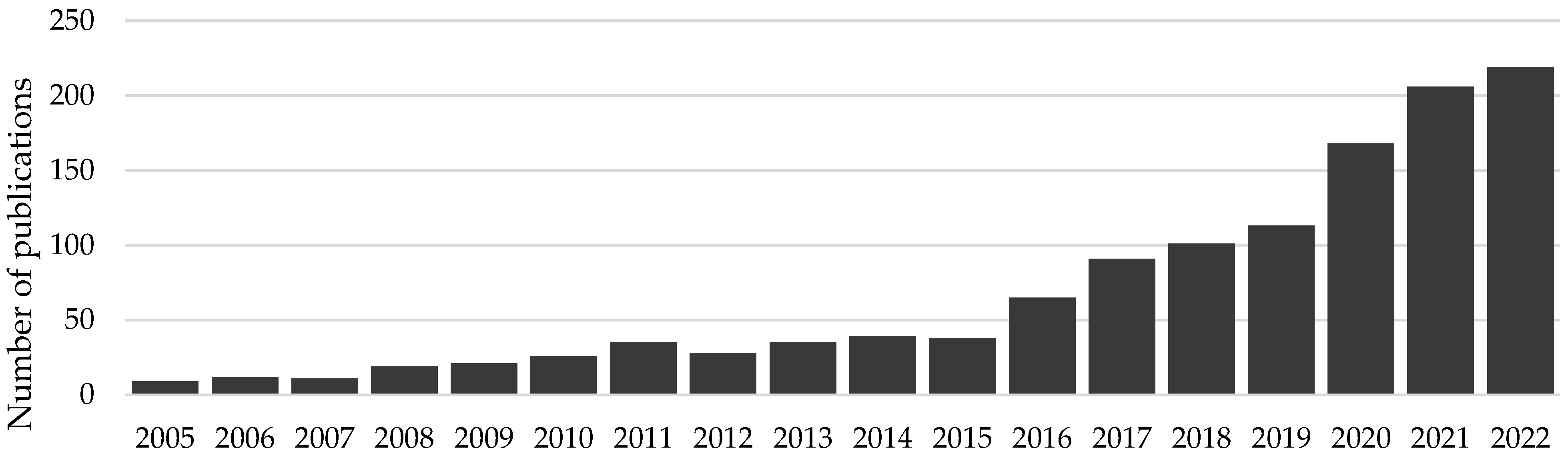
1. Introduction
2. The Nature and Extent of Foodborne Diseases
3. The Role of Biofilms in Food Contamination
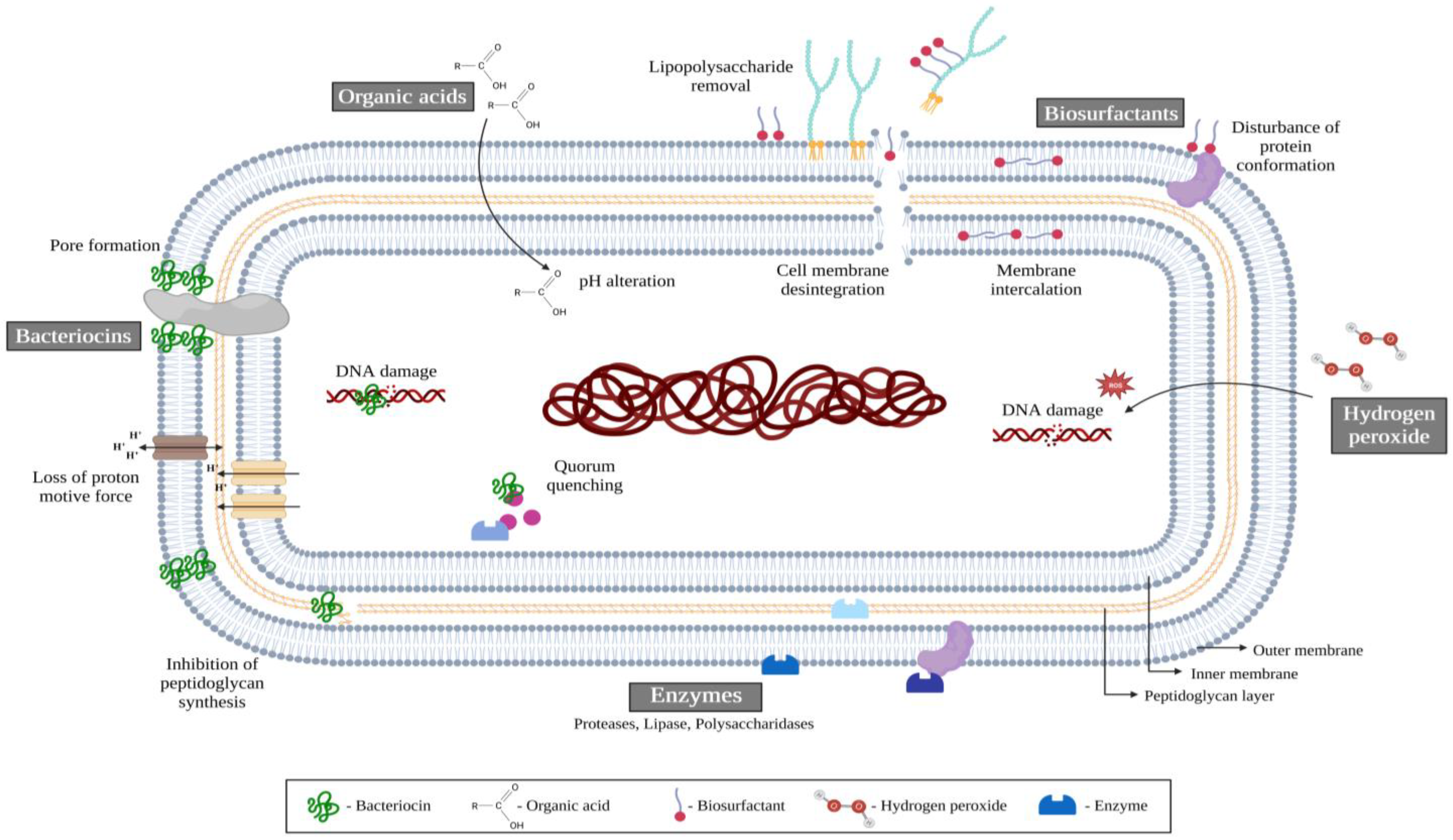
4. Biofilm Prevention and Control Strategies in the Food Industry
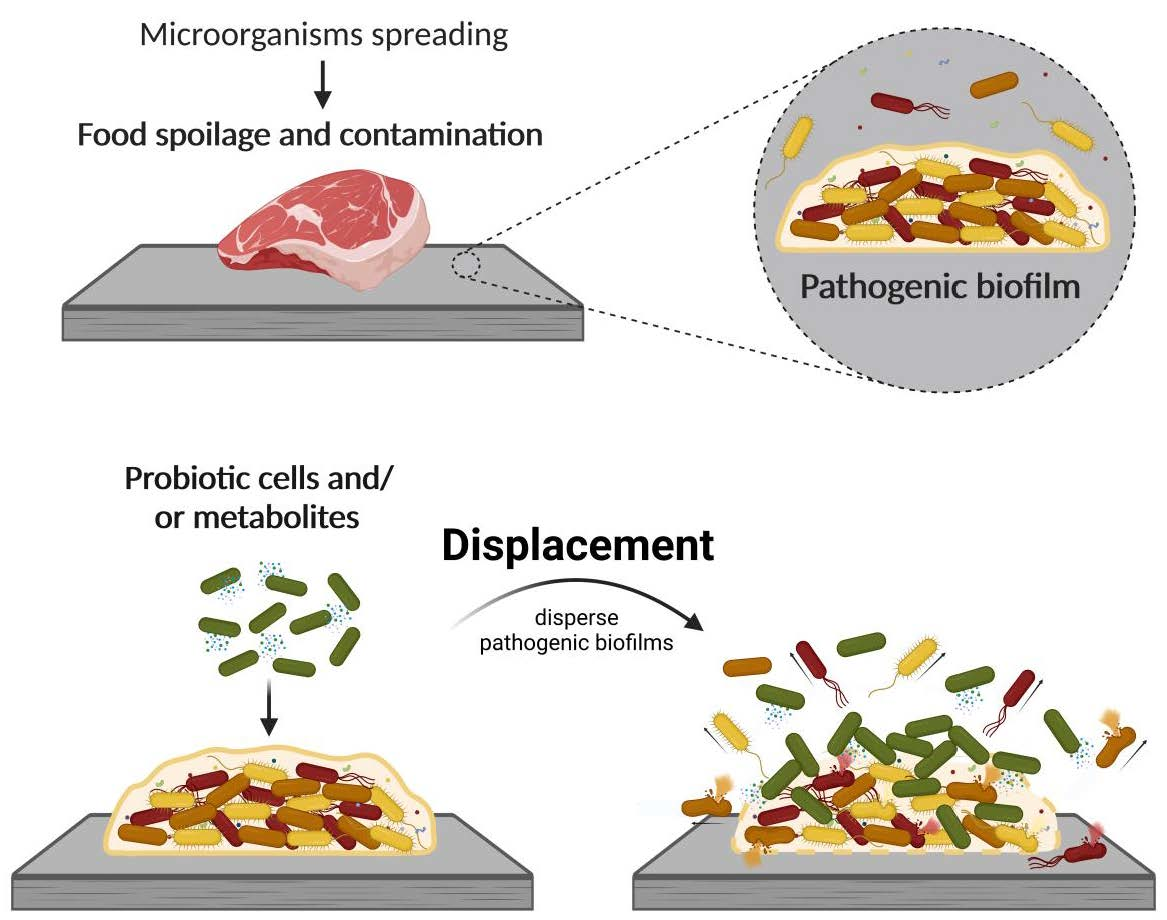
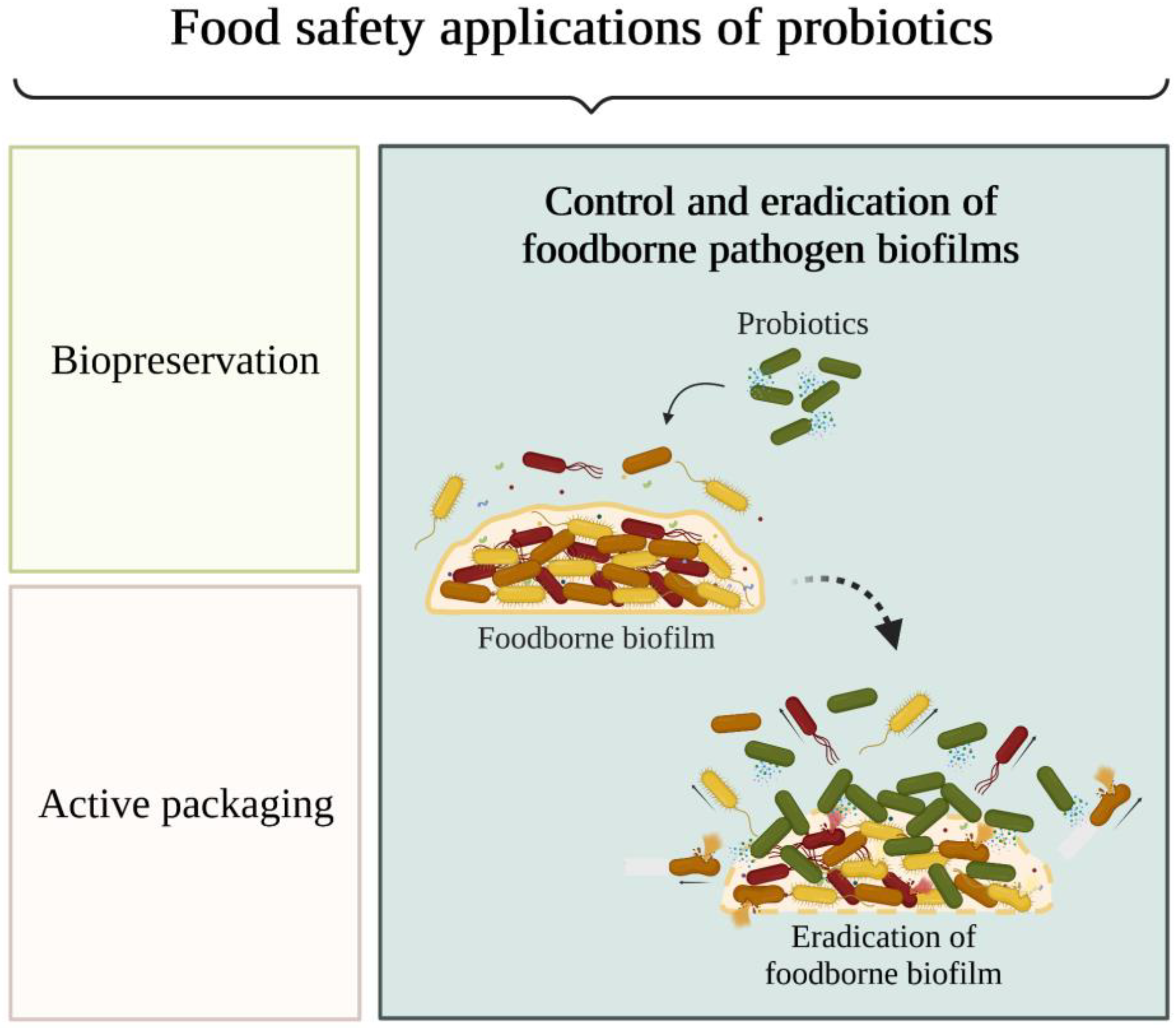
Probiotics as an Anti-Biofilm Approach
5. Probiotic Displacement Effects
5.1. Lactiplantibacillus spp.
5.2. Lacticaseibacillus spp.
5.3. Lactobacillus spp.
5.4. Limosilactobacillus spp.
5.5. Ligilactobacillus spp., Lactilactobacillus spp., Levilactobacillus spp. and Companilactobacillus spp.
5.6. Pediococcus spp., Leuconostoc spp., Lactococcus spp. and Enterococcus spp.
5.7. Bifidobacterium spp. and Saccharomyces spp.
5.8. Probiotic Cocktails
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tazehabadi, M.H.; Algburi, A.; Popov, I.V.; Ermakov, A.M.; Chistyakov, V.A.; Prazdnova, E.V.; Weeks, R.; Chikindas, M.L. Probiotic Bacilli Inhibit Salmonella Biofilm Formation Without Killing Planktonic Cells. Front. Microbiol. 2021, 12, 615328. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, S.; Martínez-Blanco, H.; Rodríguez-Aparicio, L.B.; Ferrero, M.A. Effect of fermented broth from lactic acid bacteria on pathogenic bacteria proliferation. J. Dairy Sci. 2016, 99, 2654–2665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sornsenee, P.; Chatatikun, M.; Mitsuwan, W.; Kongpol, K.; Kooltheat, N.; Sohbenalee, S.; Pruksaphanrat, S.; Mudpan, A.; Romyasamit, C. Lyophilized cell-free supernatants of Lactobacillus isolates exhibited antibiofilm, antioxidant, and reduces nitric oxide activity in lipopolysaccharide-stimulated RAW 264.7 cells. PeerJ 2021, 9, e12586. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic Mechanisms of Action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef]
- Divyashree, S.; Anjali, P.G.; Somashekaraiah, R.; Sreenivasa, M.Y. Probiotic properties of Lactobacillus casei—MYSRD 108 and Lactobacillus plantarum—MYSRD 71 with potential antimicrobial activity against Salmonella paratyphi. Biotechnol. Rep. 2021, 32, e00672. [Google Scholar] [CrossRef]
- Qualified Presumption of Safety (QPS): EFSA Journal. Available online: https://efsa.onlinelibrary.wiley.com/doi/toc/10.1002/(ISSN)1831-4732.QPS (accessed on 20 January 2023).
- EUR-Lex—Regulation (EC) No 1924/2006 of the European Parliament and of the Council of 20 December 2006 on Nutrition and Health Claims Made on Foods. Available online: https://eur-lex.europa.eu/legal-content/en/ALL/?uri=CELEX%3A32006R1924 (accessed on 20 January 2023).
- Global Overview for Probiotics: Trends, Markets, and Harmonization|RAPS. Available online: https://www.raps.org/news-and-articles/news-articles/2022/9/global-overview-for-probiotics-trends-markets-and?GA_network=x&GA_device=c&GA_campaign=18448087812&GA_adgroup=&GA_target=&GA_placement=&GA_creative=&GA_extension=&GA_keyword=&GA_loc_physical_ms=1011759&GA_landingpage=https\%3a\%2fwww.raps.org\%2fnews-and-articles\%2fnews-articles\%2f2022\%2f9\%2fglobal-overview-for-probiotics-trends-markets-and&gclid=Cj0KCQiAiJSeBhCCARIsAHnAzT-P6bWY8j7jWWPwV1asExzYsInprUJ6LfBJ3im7LfLWkMb7cPA_OQAaAm_UEALw_wcB (accessed on 20 January 2023).
- Moradi, M.; Kousheh, S.A.; Almasi, H.; Alizadeh, A.; Guimarães, J.T.; Yılmaz, N.; Lotfi, A. Postbiotics produced by lactic acid bacteria: The next frontier in food safety. Compr. Rev. Food Sci. 2020, 19, 3390–3415. [Google Scholar] [CrossRef]
- Asaithambi, N.; Singh, S.K.; Singha, P. Current status of non-thermal processing of probiotic foods: A review. J. Food Eng. 2021, 303, 110567. [Google Scholar] [CrossRef]
- Global and European Probiotic Market Insights 2018–2021. Available online: https://www.ipaeurope.org/wp-content/uploads/2022/05/Market-data-probiotics-2018-2021.pdf (accessed on 20 January 2023).
- The Burden of Foodborne Diseases in the WHO European Region. Available online: https://www.euro.who.int/__data/assets/pdf_file/0005/402989/50607-WHO-Food-Safety-publicationV4_Web.pdf (accessed on 27 October 2022).
- Hossain, M.I.; Mizan, M.F.R.; Roy, P.K.; Nahar, S.; Toushik, S.H.; Ashrafudoulla, M.; Jahid, I.K.; Lee, J.; Ha, S.-D. Listeria monocytogenes biofilm inhibition on food contact surfaces by application of postbiotics from Lactobacillus curvatus B.67 and Lactobacillus plantarum M.2. Food Res. Int. 2021, 148, 110595. [Google Scholar] [CrossRef]
- Qiao, Z.; Chen, J.; Zhou, Q.; Wang, X.; Shan, Y.; Yi, Y.; Liu, B.; Zhou, Y.; Lü, X. Purification, characterization, and mode of action of a novel bacteriocin BM173 from Lactobacillus crustorum MN047 and its effect on biofilm formation of Escherichia coli and Staphylococcus aureus. J. Dairy Sci. 2021, 104, 1474–1483. [Google Scholar] [CrossRef]
- Jaffee, S.; Henson, S.; Unnevehr, L.; Grace, D.; Cassou, E. The Safe Food Imperative: Accelerating Progress in Low- and Middle-Income Countries; The World Bank: Washington, DC, USA, 2019. [Google Scholar]
- USDA ERS—Cost Estimates of Foodborne Illnesses. Available online: https://www.ers.usda.gov/data-products/cost-estimates-of-foodborne-illnesses.aspx (accessed on 27 October 2022).
- The European Union One Health 2020 Zoonoses Report. Available online: https://efsa.onlinelibrary.wiley.com/doi/full/10.2903/j.efsa.2021.6971 (accessed on 20 January 2023).
- Tan, X.; Han, Y.; Xiao, H.; Zhou, Z. Pediococcus acidilactici Inhibit Biofilm Formation of Food-Borne Pathogens on Abiotic Surfaces. Trans. Tianjin Univ. 2017, 23, 70–77. [Google Scholar] [CrossRef]
- Sahoo, M.; Panigrahi, C.; Aradwad, P. Management strategies emphasizing advanced food processing approaches to mitigate food borne zoonotic pathogens in food system. Food Front. 2022, 3, 641–665. [Google Scholar] [CrossRef]
- EUR-Lex—Council Regulation (EEC) No 315/93 of 8 February 1993 Laying down Community Procedures for Contaminants in Food. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=celex%3A31993R0315 (accessed on 20 January 2023).
- Hadawey, A.; Savvas, A.T.; Chaer, I.; Sundararajan, R. Unwrapped food product display shelf life assessment. Energy Procedia 2017, 123, 62–69. [Google Scholar] [CrossRef]
- Coughlan, L.M.; Cotter, P.D.; Hill, C.; Alvarez-Ordóñez, A. New Weapons to Fight Old Enemies: Novel Strategies for the (Bio)control of Bacterial Biofilms in the Food Industry. Front. Microbiol. 2016, 7, 1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrascosa, C.; Raheem, D.; Ramos, F.; Saraiva, A.; Raposo, A. Microbial Biofilms in the Food Industry—A Comprehensive Review. Int. J. Environ. Res. Public Health 2021, 18, 2014. [Google Scholar] [CrossRef] [PubMed]
- Jara, J.; Pérez-Ramos, A.; del Soar, G.; Rodríguez, J.M.; Fernández, L.; Orgaz, B. Role of Lactobacillus biofilms in Listeria monocytogenes adhesion to glass surfaces. Int. J. Food Microbiol. 2020, 334, 108804. [Google Scholar] [CrossRef]
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the Food Industry: Health Aspects and Control Methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef]
- Dhivya, R.; Rajakrishnapriya, V.C.; Sruthi, K.; Chidanand, D.V.; Sunil, C.K.; Rawson, A. Biofilm combating in the food industry: Overview, non-thermal approaches, and mechanisms. J. Food Process. Preserv. 2022, 46, e16282. [Google Scholar] [CrossRef]
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.; Bonaventura, G.D.; Hébraud, M.; Jaglic, Z.; et al. Critical review on biofilm methods. Crit. Rev. Microbiol. 2016, 43, 313–351. [Google Scholar] [CrossRef] [Green Version]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as Complex Differentiated Communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef] [Green Version]
- Petrova, O.E.; Sauer, K. Sticky Situations: Key Components That Control Bacterial Surface Attachment. J. Bacteriol. 2012, 194, 2413–2425. [Google Scholar] [CrossRef] [Green Version]
- Toushik, S.H.; Kim, K.-S.; Ashrafudoulla, M.; Mizan, M.F.R.; Roy, P.K.; Nahar, S.; Kim, Y.; Ha, S.D. Korean kimchi-derived lactic acid bacteria inhibit foodborne pathogenic biofilm growth on seafood and food processing surface materials. Food Control 2021, 129, 108276. [Google Scholar] [CrossRef]
- Kim, N.-N.; Kim, W.J.; Kang, S.-S. Anti-biofilm effect of crude bacteriocin derived from Lactobacillus brevis DF01 on Escherichia coli and Salmonella Typhimurium. Food Control 2019, 98, 274–280. [Google Scholar] [CrossRef]
- Cisneros, L.; Cattelan, N.; Villalba, M.I.; Rodriguez, C.; Serra, D.O.; Yantorno, O.; Fadda, S. Lactic acid bacteria biofilms and their ability to mitigate Escherichia coli O157:H7 surface colonization. Lett. Appl. Microbiol. 2021, 73, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Yang, C.; Bao, X.; Chen, F.; Guo, X. Strategies for controlling biofilm formation in food industry. Grain Oil Sci. Technol. 2022, 5, 179–186. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Sauer, K.; Stoodley, P.; Goeres, D.M.; Hall-Stoodley, L.; Burmlle, M.; Stewart, P.S.; Bjarnsholt, T. The biofilm life cycle: Expanding the conceptual model of biofilm formation. Nat. Rev. Microbiol. 2022, 20, 608–620. [Google Scholar] [CrossRef]
- Merino, L.; Procura, F.; Trejo, F.M.; Bueno, D.J.; Golowczyc, M.A. Biofilm formation by Salmonella sp. in the poultry industry: Detection, control and eradication strategies. Food Res. Int. 2019, 119, 530–540. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Yuan, L.; Sadiq, F.A.; He, G. Inhibitory effect of Lactobacillus plantarum metabolites against biofilm formation by Bacillus licheniformis isolated from milk powder products. Food Control 2019, 106, 106721. [Google Scholar] [CrossRef]
- Kıran, F.; Akoğlu, A.; Çakır, İ. Control of Listeria monocytogenes biofilm on industrial surfaces by cell-free extracts of Lactobacillus plantarum. J. Food Process. Preserv. 2021, 45, e15042. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; El-Saadony, M.T.; Alagawany, M.; Arif, M.; Batiha, G.E.; Khafaga, A.F.; Elwan, H.A.M.; Elnesr, S.S.; Abd El-Hack, E.M. Using essential oils to overcome bacterial biofilm formation and their antimicrobial resistance. Saudi J. Biol. Sci. 2021, 28, 5145–5156. [Google Scholar] [CrossRef]
- Dobson, A.; Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocin Production: A Probiotic Trait? Appl. Environ. Microbiol. 2012, 78, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltani, S.; Biron, E.; Said, L.B.; Subirade, M.; Fliss, I. Bacteriocin-Based Synergetic Consortia: A Promising Strategy to Enhance Antimicrobial Activity and Broaden the Spectrum of Inhibition. Microbiol. Spectr. 2022, 10, e00406–e00421. [Google Scholar] [CrossRef] [PubMed]
- Salman, M.K.; Abuqwider, J.; Mauriello, G. Anti-Quorum Sensing Activity of Probiotics: The Mechanism and Role in Food and Gut Health. Microorganisms 2023, 11, 793. [Google Scholar] [CrossRef] [PubMed]
- Toushik, S.H.; Mizan, M.F.R.; Hossain, M.I.; Ha, S.-D. Fighting with old foes: The pledge of microbe-derived biological agents to defeat mono- and mixed-bacterial biofilms concerning food industries. Trends Food Sci. Technol. 2020, 99, 413–425. [Google Scholar] [CrossRef]
- Carvalho, F.M.; Teixeira-Santos, R.; Mergulhão, F.J.M.; Gomes, L.C. Targeting biofilms in medical devices using probiotic cells: A systematic review. AIMS Mater. Sci. 2021, 8, 501–523. [Google Scholar] [CrossRef]
- Carvalho, F.M.; Teixeira-Santos, R.; Mergulhão, F.J.M.; Gomes, L.C. The Use of Probiotics to Fight Biofilms in Medical Devices: A Systematic Review and Meta-Analysis. Microorganisms 2021, 9, 27. [Google Scholar] [CrossRef]
- Gavrilova, E.; Anisimova, E.; Gabdelkhadieva, A.; Nikitina, E.; Vafina, A.; Yarullina, D.; Bogachev, M.; Kayumov, A. Newly isolated lactic acid bacteria from silage targeting biofilms of foodborne pathogens during milk fermentation. BMC Microbiol. 2019, 19, 248. [Google Scholar] [CrossRef]
- Ruiz, M.J.; García, M.D.; Padola, N.L.; Etcheverría, A.I. Ability of Lactiplantibacillus plantarum to reduce biofilms of pathogens involved in foodborne diseases. Rev. Vet. 2022, 31, 48–52. [Google Scholar] [CrossRef]
- Todorov, S.D.; de Paula, O.A.L.; Camargo, A.C.; Lopes, D.A.; Nero, L.A. Combined effect of bacteriocin produced by Lactobacillus plantarum ST8SH and vancomycin, propolis or EDTA for controlling biofilm development by Listeria monocytogenes. Rev. Argent. Microbiol. 2018, 50, 48–55. [Google Scholar] [CrossRef]
- Liu, Y.; Bu, Y.; Li, J.; Liu, Y.; Liu, A.; Gong, P.; Liu, T.; Zhang, L.; Wang, S.; Yi, H. Inhibition Activity of Plantaricin Q7 Produced by Lactobacillus plantarum Q7 against Listeria monocytogenes and Its Biofilm. Fermentation 2022, 8, 75. [Google Scholar] [CrossRef]
- Göksel, Ş.; Akçelik, N.; Özdemir, C.; Akçelik, M. The Effects of Lactic Acid Bacteria on Salmonella Biofilms. J. Microbiol. 2022, 91, 278–285. [Google Scholar] [CrossRef]
- Rao, K.P.; Kumar, N.H.; Somashekaraiah, R.; Murali, M.; Sreenivasa, M.Y. Probiotic Attributes and Inhibitory Effects of Lactobacillus plantarum MYS84 against the Growth and Biofilm Formation of Pseudomonas aeruginosa. Microbiology 2021, 90, 361–369. [Google Scholar] [CrossRef]
- Ben Slama, R.; Kouidhi, B.; Zmantar, T.; Chaieb, K.; Bakhrouf, A. Anti-listerial and Anti-biofilm Activities of Potential Probiotic Lactobacillus Strains Isolated from Tunisian Traditional Fermented Food. J. Food Saf. 2013, 33, 8–16. [Google Scholar] [CrossRef]
- Lv, X.; Cui, T.; Du, H.; Sun, M.; Bai, F.; Li, J.; Zhang, D. Lactobacillus plantarum CY 1-1: A novel quorum quenching bacteria and anti-biofilm agent against Aeromonas sobria. LWT 2021, 137, 110439. [Google Scholar] [CrossRef]
- Woo, J.; Ahn, J. Probiotic-mediated competition, exclusion and displacement in biofilm formation by food-borne pathogens. Lett. Appl. Microbiol. 2013, 56, 307–313. [Google Scholar] [CrossRef]
- Shangguan, W.; Xie, T.; Zhang, R.; Lu, C.; Han, X.; Zhong, Q. Anti-biofilm potential of kefir-derived Lactobacillus paracasei L10 against Vibrio parahaemolyticus. Lett. Appl. Microbiol. 2021, 73, 750–758. [Google Scholar] [CrossRef]
- Koohestani, M.; Moradi, M.; Tajik, H.; Badali, A. Effects of cell-free supernatant of Lactobacillus acidophilus LA5 and Lactobacillus casei 431 against planktonic form and biofilm of Staphylococcus aureus. Vet. Res. Forum 2018, 9, 301–306. [Google Scholar]
- Singh, N.; Kaur, R.; Singh, B.P.; Rokana, N.; Goel, G.; Puniya, A.K.; Panwar, H. Impairment of Cronobacter sakazakii and Listeria monocytogenes biofilms by cell-free preparations of lactobacilli of goat milk origin. Folia Microbiol. 2020, 65, 185–196. [Google Scholar] [CrossRef]
- Moradi, M.; Mardani, K.; Tajik, H. Characterization and application of postbiotics of Lactobacillus spp. on Listeria monocytogenes in vitro and in food models. LWT 2019, 111, 457–464. [Google Scholar] [CrossRef]
- Nataraj, B.H.; Ramesh, C.; Mallappa, R.H. Characterization of biosurfactants derived from probiotic lactic acid bacteria against methicillin-resistant and sensitive Staphylococcus aureus isolates. LWT 2021, 151, 112195. [Google Scholar] [CrossRef]
- Patel, M.; Siddiqui, A.J.; Hamadou, W.S.; Surti, M.; Awadelkareem, A.M.; Ashraf, S.A.; Alreshidi, M.; Snoussi, M.; Rizvi, S.M.D.; Bardakci, F.; et al. Inhibition of Bacterial Adhesion and Antibiofilm Activities of a Glycolipid Biosurfactant from Lactobacillus rhamnosus with Its Physicochemical and Functional Properties. Antibiotics 2021, 10, 1546. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Sharma, C.; Gulhane, R.D.; Rokana, N.; Singh, B.P.; Puniya, A.K.; Attri, S.; Goel, G.; Panwar, H. Inhibitory effects of lactobacilli of goat’s milk origin against growth and biofilm formation by pathogens: An in vitro study. Food Biosci. 2018, 22, 129–138. [Google Scholar] [CrossRef]
- Shokri, D.; Khorasgani, M.R.; Mohkam, M.; Fatemi, S.M.; Ghasemi, Y.; Taheri-Kafrani, A. The Inhibition Effect of Lactobacilli Against Growth and Biofilm Formation of Pseudomonas aeruginosa. Probiotics Antimicrob. Proteins 2018, 10, 34–42. [Google Scholar] [CrossRef]
- Aman, M.; Aneeqha, N.; Bristi, K.; Deeksha, J.; Afza, N.; Sindhuja, V.; Shastry, R.P. Lactic acid bacteria inhibits quorum sensing and biofilm formation of Pseudomonas aeruginosa strain JUPG01 isolated from rancid butter. Biocatal. Agric. Biotechnol. 2021, 36, 102115. [Google Scholar] [CrossRef]
- Pérez-Ibarreche, M.; Castellano, P.; Leclercq, A.; Vignolo, G. Control of Listeria monocytogenes biofilms on industrial surfaces by the bacteriocin-producing Lactobacillus sakei CRL1862. FEMS Microbiol. Lett. 2016, 363, fnw118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, T.; Bai, F.; Sun, M.; Lv, X.; Li, X.; Zhang, D.; Du, H. Lactobacillus crustorum ZHG 2-1 as novel quorum-quenching bacteria reducing virulence factors and biofilms formation of Pseudomonas aeruginosa. LWT 2020, 117, 108696. [Google Scholar] [CrossRef]
- Abid, Y.; Casillo, A.; Gharsallah, H.; Joulak, I.; Lanzetta, R.; Corsaro, M.M.; Attia, H.; Azabou, S. Production and structural characterization of exopolysaccharides from newly isolated probiotic lactic acid bacteria. Int. J. Biol. Macromol. 2018, 108, 719–728. [Google Scholar] [CrossRef]
- Zhao, T.; Podtburg, T.C.; Zhao, P.; Chen, D.; Baker, D.A.; Cords, B.; Doyle, M.P. Reduction by Competitive Bacteria of Listeria monocytogenes in Biofilms and Listeria Bacteria in Floor Drains in a Ready-to-Eat Poultry Processing Plant. J. Food Prot. 2013, 76, 601–607. [Google Scholar] [CrossRef]
- García-Almendárez, B.E.; Cann, I.K.O.; Martin, S.E.; Guerrero-Legarreta, I.; Regalado, C. Effect of Lactococcus lactis UQ2 and its bacteriocin on Listeria monocytogenes biofilms. Food Control 2008, 19, 670–680. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yu, H.H.; Song, Y.J.; Park, Y.J.; Lee, N.-K.; Paik, H.-D. Anti-biofilm effect of the cell-free supernatant of probiotic Saccharomyces cerevisiae against Listeria monocytogenes. Food Control 2021, 121, 107667. [Google Scholar] [CrossRef]
- Ndahetuye, J.B.; Koo, O.K.; O’Bryan, C.A.; Ricke, S.C.; Crandall, P.G. Role of Lactic Acid Bacteria as a Biosanitizer To Prevent Attachment of Listeria monocytogenes F6900 on Deli Slicer Contact Surfaces. J. Food Prot. 2012, 75, 1429–1436. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, G.P.; Rossi, D.A.; Valadares, E.C.; Peres, P.A.B.M.; Braz, R.F.; Notário, F.O.; Gomes, M.M.; Silva, R.R.; Carrijo, K.F.; Fonseca, B.B. Lactic Bacterium and Bacillus Sp. Biofilms Can Decrease the Viability of Salmonella gallinarum, Salmonella heidelberg, Campylobacter jejuni and Methicillin Resistant Staphylococcus aureus on Different Substrates. Braz. J. Poult. Sci. 2021, 23, 1408. [Google Scholar] [CrossRef]
- Hossain, M.I.; Mizan, M.F.R.; Ashrafudoulla, M.; Nahar, S.; Joo, H.-J.; Jahid, I.K.; Park, S.H.; Kim, K.-S.; Ha, S.-D. Inhibitory effects of probiotic potential lactic acid bacteria isolated from kimchi against Listeria monocytogenes biofilm on lettuce, stainless-steel surfaces, and MBEC™ biofilm device. LWT 2020, 118, 108864. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, J.; Liu, Z.; Pei, J.; Brennan, C.; Abd El-Aty, A.M. Isolation and Characterization of Bacteriocin-Producing Lacticaseibacillus rhamnosus XN2 from Yak Yoghurt and Its Bacteriocin. Molecules 2022, 27, 2066. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.M.; Teixeira-Santos, R.; Mergulhão, F.J.M.; Gomes, L.C. Effect of Lactobacillus plantarum Biofilms on the Adhesion of Escherichia coli to Urinary Tract Devices. Antibiotics 2021, 10, 966. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probiotic Genus | Anti-Biofilm Compound | Foodborne Microorganism | Outcomes | References |
|---|---|---|---|---|
| Lactiplantibacillus | Cells | Bacillus cereus | No repression of B. cereus was observed. In contrast, treated S. aureus biofilms were reduced by up to 100%. Likewise, the removal rates of established biofilms of E. coli and P. aeruginosa were higher than 93.7% and 99.9%, respectively. A remarkable biofilm reduction of 99.6% was observed for S. typhimurium. | [46,47] |
| Staphylococcus aureus | ||||
| Escherichia coli | ||||
| Pseudomonas aeruginosa | ||||
| Salmonella typhimurium | ||||
| Bacteriocin | Listeria monocytogenes | Treatment with bacteriocins completely disintegrated L. monocytogenes biofilms and caused extensive cell membrane damage. High concentrations of bacteriocins were more effective in removing biofilms. | [48,49] | |
| CFS | S. typhimurium | CFS promoted biofilm growth. | [50] | |
| P. aeruginosa | P. aeruginosa biofilms were reduced by 15–78% in a concentration-dependent manner. | [51] | ||
| L. monocytogenes | Biofilm reductions of up to 90% were obtained upon treatment with CFS. | [52] | ||
| Crude extract | Aeromonas sobria | Biofilm removal of up to 66% was observed, possibly due to the reduction in the levels of some virulence factors, thus suppressing QS. | [53] | |
| Lacticaseibacillus | Cells | S. typhimurium | The displacement activity of the probiotic planktonic cells was not effective against S. typhimurium (up to 0.4 log reduction). Biofilm dispersion was more pronounced for L. monocytogenes (99.9%). | [54] |
| L. monocytogenes | ||||
| CFS | Vibrio parahaemolyticus | Biofilm biomass and metabolic activity were reduced by up to 41%. | [55] | |
| S. aureus | CFS significantly removed the pre-established biofilm in the range of 65–77%. | [56] | ||
| E. coli | Significant eradication of established biofilms of both E. coli (58–84%) and A. baumannii (28–63%) was observed after treatment with the CFS. | [3] | ||
| Acinetobacter baumannii | ||||
| Cronobacter sakazakii | The reduction of pre-formed biofilms was around 10–51% for C. sakazakii and 16–52% for L. monocytogenes, depending on the contact time and CFS concentration. | [57,58] | ||
| L. monocytogenes | ||||
| Biosurfactant | S. aureus | Biosurfactants extracted from Lacticaseibacillus dispersed the pre-formed biofilms in a dose-dependent manner, ranging from 48% to 76%. | [59,60] | |
| E. coli | ||||
| Bacillus subtilis | ||||
| P. aeruginosa | ||||
| Lactobacillus | Cells | S. typhimurium | The displacement activity of probiotic cells was not effective against S. typhimurium (0.8 log reduction). Biofilm dispersion was more pronounced against L. monocytogenes (97%). | [54] |
| L. monocytogenes | ||||
| CFS | S. aureus | CFS reduced the population of sessile cells of P. aeruginosa by up to 77% and removed S. aureus biofilms by approximately 18–87%. Following neutralisation, a marked reduction in biofilm inhibition was observed. | [56,61] | |
| P. aeruginosa | ||||
| L. monocytogenes | CFS reduced the biofilm amount of L. monocytogenes by 48%. | [58] | ||
| Biosurfactant | S. aureus | Biosurfactants extracted from the probiotic strains dispersed the pre-formed biofilms around 45–63%. | [59] | |
| Limosilactobacillus | Cells | B. cereus | Whereas B. cereus biofilms were not affected, viable S. aureus cells were not detected in the biofilms exposed to the probiotics. The removal values of the pre-established E. coli and P. aeruginosa biofilms were higher than 6 and 5 log CFU/mL, respectively. | [46] |
| S. aureus | ||||
| E. coli | ||||
| P. aeruginosa | ||||
| CFS | P. aeruginosa | The CFS of Limosilactobacillus totally removed the pre-formed biofilms of P. aeruginosa. | [62] | |
| E. coli | Significant eradication of established biofilms of both E. coli (58–84%) and A. baumannii (28–63%) was observed after treatment with CFS. | [3] | ||
| A. baumannii | ||||
| Crude extract | Chromobacterium violaceum | The crude extract exhibited significant QS inhibitory and anti-biofilm properties, reducing C. violaceum biofilm by 3 to 40% and P. aeruginosa biofilm by up to 32%. | [63] | |
| P. aeruginosa | ||||
| Ligilactobacillus | CFS | L. monocytogenes | L. monocytogenes biofilm was depleted by 63% with Ligilactobacillus CFS. | [58] |
| Lactilactobacillus | Cells | L. monocytogenes | Lactilactobacillus cells were able to displace the pre-established biofilm by 1.8–2.2 log CFU/cm2. An enhanced pathogen inhibition was observed when the semi-purified bacteriocin extract was added (reductions of 3.1–3.6 log CFU/cm2). | [64] |
| Bacteriocin | ||||
| CFS | S. typhimurium | CFS increased the S. typhimurium biofilm due to the inability of the anti-biofilm substances to diffuse through the biofilm matrix. | [50] | |
| Levilactobacillus | Bacteriocin | E. coli | Bacteriocins did not significantly remove the biofilms of E. coli or S. typhimurium (only 16%). | [31] |
| S. typhimurium | ||||
| CFS | E. coli | Significant eradication of established biofilms of both E. coli (58–84%) and A. baumannii (28–63%) was observed. | [3] | |
| A. baumannii | ||||
| Companilactobacillus | Crude extract | P. aeruginosa | Crude extract of Companilactobacillus had a strong removal effect on P. aeruginosa biofilms, achieving up to 39% biomass removal and 98% CFU reduction. | [65] |
| Pediococcus | Cells | S. aureus | Pediococcus had a significant inhibition effect in the displacement strategy, with a reduction of 3–4 log CFU/coupon. | [18] |
| S. typhimurium | ||||
| L. monocytogenes | ||||
| E. coli | ||||
| CFS | S. typhimurium | Pediococcus caused a statistically significant removal (33%) of mature S. typhimurium biofilms. | [50] | |
| Crude extract | C. violaceum | The crude extract exhibited significant QS inhibitory and anti-biofilm properties, reducing C. violaceum and P. aeruginosa biofilms by up to 40% and 32%, respectively. | [63] | |
| P. aeruginosa | ||||
| EPS | S. aureus | S. aureus, E. coli, and E. faecalis mature biofilms were reduced by up to 75%, 52% and 50%, respectively, and the disruptive activity increased with increasing EPS concentration. | [66] | |
| E. coli | ||||
| Enterococcus faecalis | ||||
| Lactococcus | Cells | L. monocytogenes | L. monocytogenes biofilms decreased by 1–6 log and 2.7 log when exposed to cells and bacteriocin from Lactococcus, respectively. | [67,68] |
| Bacteriocin | ||||
| Leuconostoc | EPS | S. aureus | S. aureus, E. coli, and E. faecalis biofilms were reduced by up to 77%, 62% and 53%, respectively, and the disruptive activity increased with increasing EPS concentration. | [66] |
| E. coli | ||||
| E. faecalis | ||||
| Enterococcus | Cells | L. monocytogenes | L. monocytogenes biofilms decreased by 1–5 log. | [67] |
| Bifidobacterium | CFS | S. typhimurium | The CFS of Bifidobacterium did not affect mature S. typhimurium biofilms. | [50] |
| Saccharomyces | CFS | L. monocytogenes | The structure of CFS-treated L. monocytogenes biofilms was dispersed, and the number of cells attached to the surface decreased by 52–77%. | [69] |
| Cocktail (Lactobacillus and Pediococcus) | Cells | L. monocytogenes | The LAB cocktail significantly displaced L. monocytogenes biofilms after a 24 h contact time (98%) and was still able to reduce mature biofilms after 72 h. | [70] |
| Cocktail (Bacillus and Pediococcus) | Cells | Salmonella gallinarum | Biofilm reductions of up to 99.9% were detected. | [71] |
| Salmonella heidelberg | ||||
| S. aureus | ||||
| Campylobacter jejuni |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomé, A.R.; Carvalho, F.M.; Teixeira-Santos, R.; Burmølle, M.; Mergulhão, F.J.M.; Gomes, L.C. Use of Probiotics to Control Biofilm Formation in Food Industries. Antibiotics 2023, 12, 754. https://doi.org/10.3390/antibiotics12040754
Tomé AR, Carvalho FM, Teixeira-Santos R, Burmølle M, Mergulhão FJM, Gomes LC. Use of Probiotics to Control Biofilm Formation in Food Industries. Antibiotics. 2023; 12(4):754. https://doi.org/10.3390/antibiotics12040754
Chicago/Turabian StyleTomé, Andreia R., Fábio M. Carvalho, Rita Teixeira-Santos, Mette Burmølle, Filipe J. M. Mergulhão, and Luciana C. Gomes. 2023. "Use of Probiotics to Control Biofilm Formation in Food Industries" Antibiotics 12, no. 4: 754. https://doi.org/10.3390/antibiotics12040754