
Prevalence and Molecular Characterization of Methicillin-Resistant Staphylococci (MRS) and Mammaliicocci (MRM) in Dromedary Camels from Algeria: First Detection of SCCmec-mecC Hybrid in Methicillin-Resistant Mammaliicoccus lentus
 ,
,  ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Determination of Staphylococcaceae Flora in the Nose of Dromedary Camels
2.2. Detection of MRS and MRM in Dromedary Camels
2.3. WGS Analysis
2.3.1. WGS and Phylogenetic Analysis
2.3.2. Analysis of Antimicrobial Resistance Genes
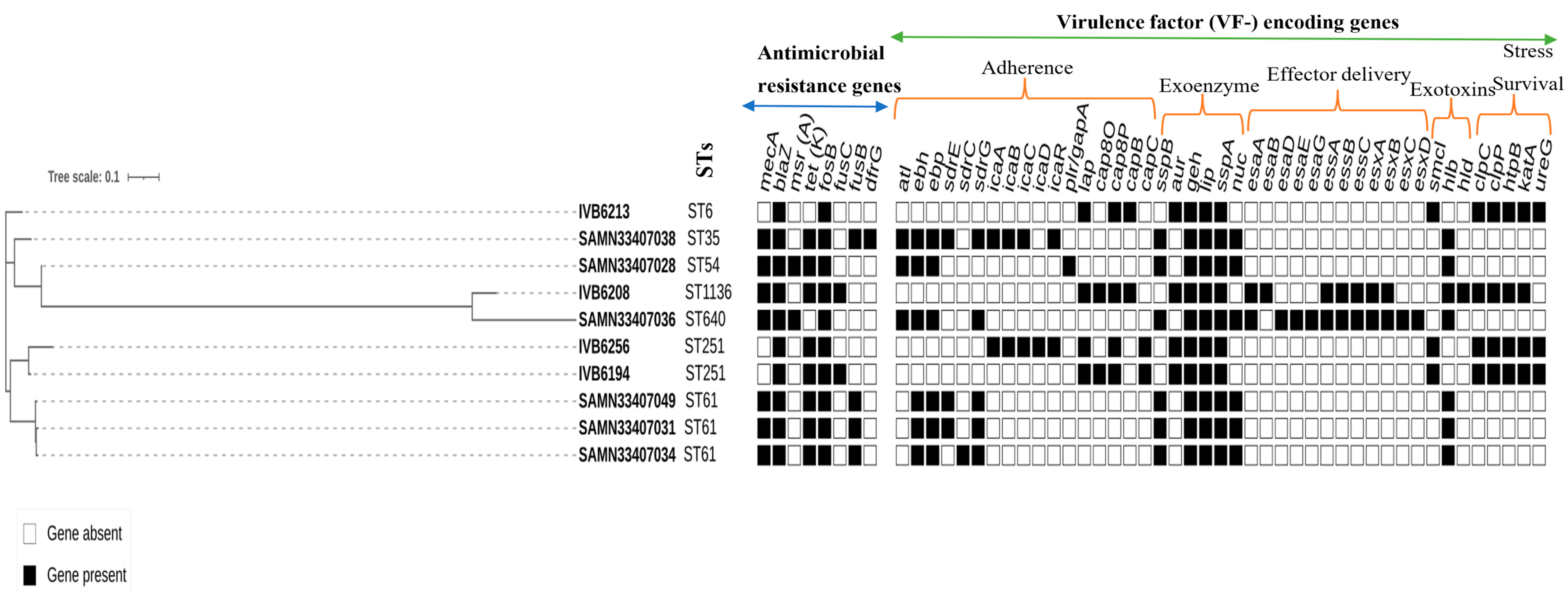
2.3.3. Analysis of Virulence Factors
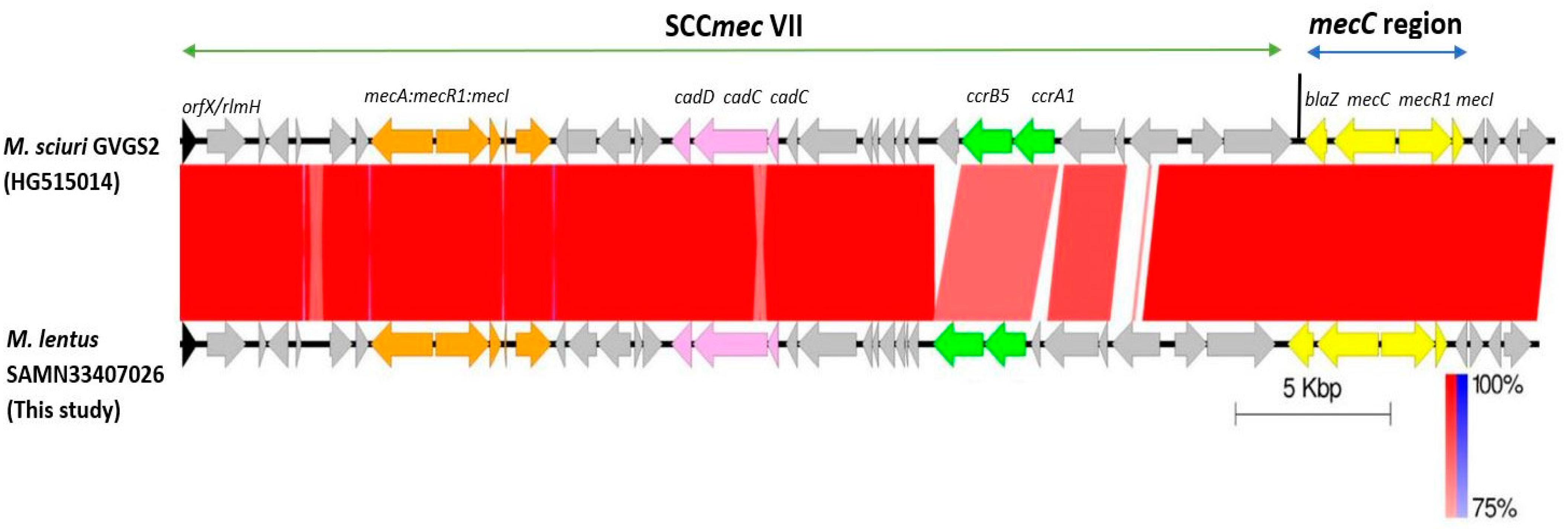
2.3.4. SCCmec Analysis
3. Discussion
4. Materials and Methods
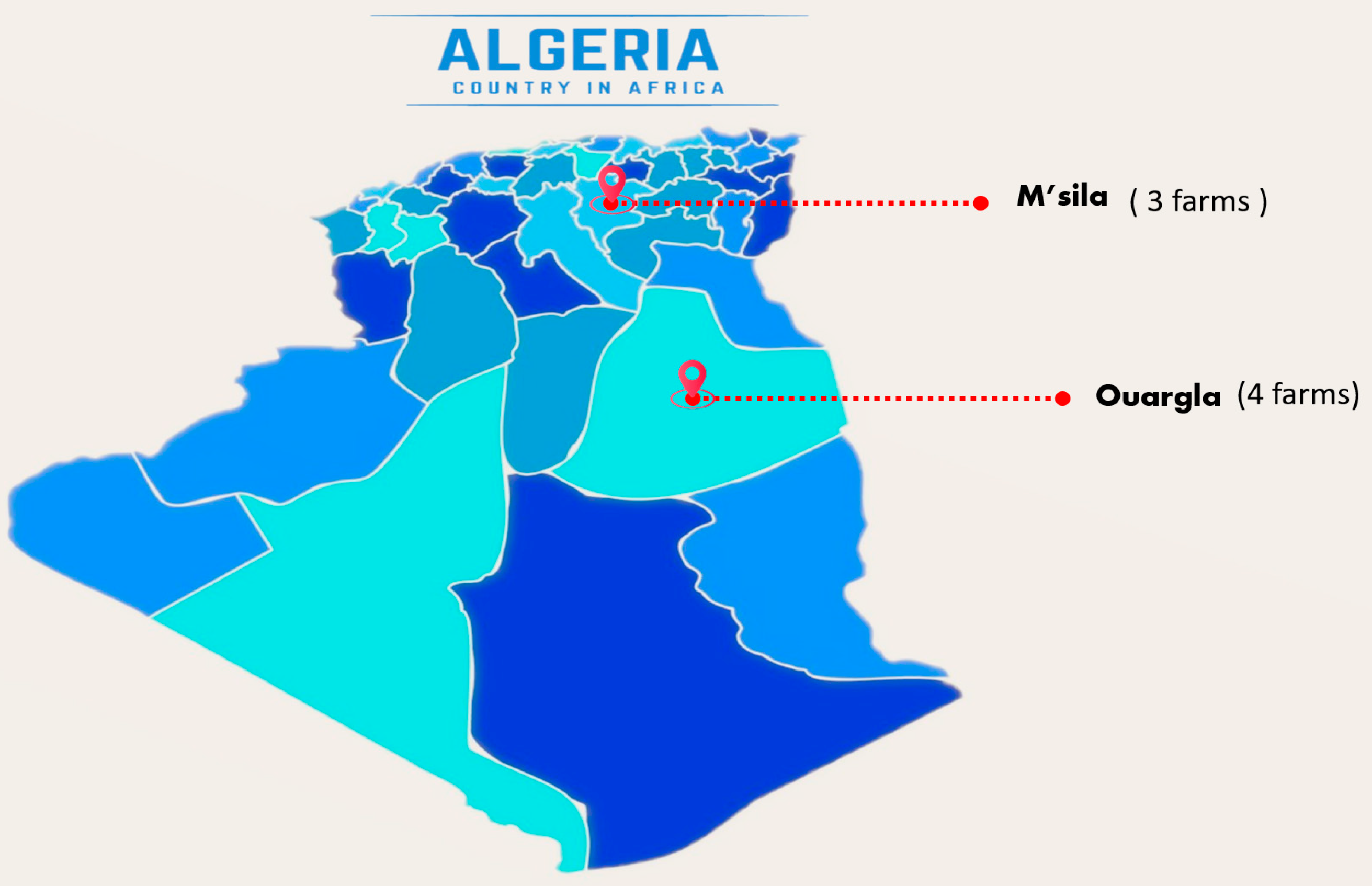
4.1. Sample Origin and Isolation Methods
4.1.1. Isolation of Non-Methicillin-Resistant Staphylococcaceae
4.1.2. Isolation of Methicillin-Resistant Staphylococcaceae
4.2. Bacterial Identification
4.3. PCR for mec Genes Identification
4.4. Whole Genome Sequencing (WGS)
4.4.1. Genome Assembly and Annotation
4.4.2. Core Genome Analysis
4.4.3. Accessory Genome Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Konuspayeva, G.; Faye, B.; Gagaoua, M.; Dib, A.L.; Bererhi, E.; Pulina, G. Recent Advances in Camel Milk Processing. Animals 2021, 11, 1045. [Google Scholar] [CrossRef] [PubMed]
- Al-Amery, K.; Elhariri, M.; Elsayed, A.; El-Moghazy, G.; Elhelw, R.; El-Mahallawy, H.; El Hariri, M.; Hamza, D. Vancomycin-resistant Staphylococcus aureus isolated from camel meat and slaughterhouse workers in Egypt. Antimicrob. Resist. Infect. Control 2019, 8, 129. [Google Scholar] [CrossRef] [Green Version]
- Dioli, M. Dromedary (Camelus dromedarius) and Bactrian Camel (Camelus bactrianus) Crossbreeding Husbandry Practices in Turkey and Kazakhstan: An In-Depth Review. Pastoralism 2020, 10, 6. [Google Scholar] [CrossRef]
- Swanepoel, F.; Stroebel, A.; Moyo, S. The Role of Livestock in Developing Communities: Enhancing Multifunctionality; University of the Free State; Technical Centre for Agricultural and Rural Cooperation: Cape Town, South Africa, 2010; Available online: https://cgspace.cgiar.org/handle/10568/3003 (accessed on 5 March 2023).
- Zarrin, M.; Riveros, J.L.; Ahmadpour, A.; de Almeida, A.M.; Konuspayeva, G.; Vargas-Bello-Pérez, E.; Faye, B.; Hernández-Castellano, L.E. Camelids: New Players in the International Animal Production Context. Trop. Anim. Health Prod. 2020, 52, 903–913. [Google Scholar] [CrossRef]
- Al-Jassim, R.; Sejian, V. Review Paper: Climate Change and Camel Production: Impact and Contribution. J. Camelid Sci. 2015, 8, 1–17. [Google Scholar]
- Harrak, M.E.; Faye, B.; Bengoumi, M. Main Pathologies of Camels, Breeding of Camels, Constraints, Benefits and Perspectives. In Proceedings of the 4th Conference of the International Society of Camelid Research and Development, Djerba, Tunisia, 8–12 December 2011. [Google Scholar]
- Zhu, S.; Zimmerman, D.; Deem, S.L. A Review of Zoonotic Pathogens of Dromedary Camels. EcoHealth 2019, 16, 356–377. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, N.M.; Van Kerkhove, M.D. Identification of MERS-CoV in Dromedary Camels. Lancet Infect. Dis. 2014, 14, 93–94. [Google Scholar] [CrossRef] [Green Version]
- Boukary, A.R.; Saegerman, C.; Abatih, E.; Fretin, D.; Bada, R.A.; De Deken, R.; Thys, E.; Igoho, S.B.; Dicko, A.H.; Van Damme, I.; et al. Seroprevalence and Potential Risk Factors for Brucella spp. Infection in Traditional Cattle, Sheep and Goats Reared in Urban, Periurban and Rural Areas of Niger. PLoS ONE 2013, 8, e83175. [Google Scholar] [CrossRef]
- Babelhadj, B.; Di Bari, M.A.; Pirisinu, L.; Chiappini, B.; Gaouar, S.B.S.; Riccardi, G.; Agrimi, U.; Nonno, R. Prion Disease in Dromedary Camels, Algeria. Emerg. Infect. Dis. 2018, 24, 1029–1036. [Google Scholar] [CrossRef] [Green Version]
- Gossner, C.; Danielson, N.; Gervelmeyer, A.; Berthe, F.; Faye, B.; Kaasik Aaslav, K.; Adlhoch, C.; Zeller, H.; Penttinen, P. Human-Dromedary Camel Interactions and the Risk of Acquiring Zoonotic Middle East Respiratory Syndrome Coronavirus Infection. Zoonoses Public Health 2016, 63, 557–568. [Google Scholar] [CrossRef]
- Watson, E.E.; Kochore, H.H.; Dabasso, B.H. Camels and Climate Resilience: Adaptation in Northern Kenya. Hum. Ecol. 2016, 44, 701–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akunda, I.K.; Kariuki, D.W.; Matulis, G.; Mitema, E.S.; Muya, S.N.; Kariuki, S. Antimicrobial Resistance Patterns and Characterisation of Emerging Beta-Lactamase-Producing Escherichia coli in Camels Sampled from Northern Kenya. Vet. Med. Sci. 2023. [CrossRef]
- Madhaiyan, M.; Wirth, J.S.; Saravanan, V.S. Phylogenomic Analyses of the Staphylococcaceae Family Suggest the Reclassification of Five Species within the Genus Staphylococcus as Heterotypic Synonyms, the Promotion of Five Subspecies to Novel Species, the Taxonomic Reassignment of Five Staphylococcus Species and the Addition of 14 Novel Species to the Genus Staphylococcus. Int. J. Syst. Evol. Microbiol. 2020, 70, 5926–5936. [Google Scholar] [CrossRef] [PubMed]
- Belhout, C.; Elgroud, R.; Butaye, P. Methicillin-Resistant Staphylococcus aureus (MRSA) and Other Methicillin-Resistant Staphylococci and Mammaliicoccus (MRNaS) Associated with Animals and Food Products in Arab Countries: A Review. Vet. Sci. 2022, 9, 317. [Google Scholar] [CrossRef]
- Nemeghaire, S.; Vanderhaeghen, W.; Angeles Argudín, M.; Haesebrouck, F.; Butaye, P. Characterization of methicillin-resistant Staphylococcus sciuri isolates from industrially raised pigs, cattle and broiler chickens. J. Antimicrob. Chemother. 2014, 69, 2928–2934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peton, V.; Le Loir, Y. Staphylococcus aureus in veterinary medicine. Infect. Genet. Evol. 2014, 21, 602–615. [Google Scholar] [CrossRef] [PubMed]
- Nemeghaire, S.; Argudín, M.A.; Feßler, A.T.; Hauschild, T.; Schwarz, S.; Butaye, P. The ecological importance of the Staphylococcus sciuri species group as a reservoir for resistance and virulence genes. Vet. Microbiol. 2014, 171, 342–356. [Google Scholar] [CrossRef] [PubMed]
- Vanderhaeghen, W.; Vandendriessche, S.; Crombé, F.; Nemeghaire, S.; Dispas, M.; Denis, O.; Hermans, K.; Haesebrouck, F.; Butaye, P. Characterization of methicillin-resistant non-Staphylococcus aureus staphylococci carriage isolates from different bovine populations. J. Antimicrob. Chemother. 2013, 68, 300–307. [Google Scholar] [CrossRef] [Green Version]
- Butaye, P.; Argudín, M.A.; Smith, T.C. Livestock-Associated MRSA and Its Current Evolution. Curr. Clin. Microbiol. Rep. 2016, 3, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Schwendener, S.; Perreten, V. The bla and mec families of β-lactam resistance genes in the genera Macrococcus, Mammaliicoccus and Staphylococcus: An in-depth analysis with emphasis on Macrococcus. J. Antimicrob. Chemother. 2022, 77, 2571–2573. [Google Scholar] [CrossRef]
- Kirby, W.M.M. Extraction of a highly potent penicillin inactivator from penicillin resistant staphylococci. Science 1944, 99, 452–453. [Google Scholar] [CrossRef] [PubMed]
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus aureus: Molecular Characterization, Evolution, and Epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.; van Alen, S.; Idelevich, E.A.; Schleimer, N.; Seggewiß, J.; Mellmann, A.; Kempf, V.A.J.; Kriegeskorte, A. Plasmid-Encoded Transferable mecB-Mediated Methicillin Resistance in Staphylococcus aureus. Emerg. Infect. Dis. 2018, 24, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Hu, Y.; Baker, M.; Dottorini, T.; Li, H.; Dong, Y.; Chen, M.; Zheng, X.; Liu, Y. Novel SCCmec Type XV (7A) and Two Pseudo-SCCmec Variants in Foodborne MRSA in China. J. Antimicrob. Chemother. 2022, 77, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Shore, A.C.; Coleman, D.C. Staphylococcal Cassette Chromosome mec: Recent Advances and New Insights. Int. J. Med. Microbiol. 2013, 303, 350–359. [Google Scholar] [CrossRef]
- Wang, L.; Archer, G.L. Roles of CcrA and CcrB in Excision and Integration of Staphylococcal Cassette Chromosome mec, a Staphylococcus aureus Genomic Island. J. Bacteriol. 2010, 192, 3204–3212. [Google Scholar] [CrossRef] [Green Version]
- Misiura, A.; Pigli, Y.Z.; Boyle-Vavra, S.; Daum, R.S.; Boocock, M.R.; Rice, P.A. Roles of Two Large Serine Recombinases in Mobilizing the Methicillin-Resistance Cassette SCCmec. Mol. Microbiol. 2013, 88, 1218–1229. [Google Scholar] [CrossRef] [Green Version]
- García-Álvarez, L.; Holden, M.T.; Lindsay, H.; Webb, C.R.; Brown, D.F.; Curran, M.D.; Walpole, E.; Brooks, K.; Pickard, D.J.; Teale, C.; et al. Meticillin-Resistant Staphylococcus aureus with a Novel mecA Homologue in Human and Bovine Populations in the UK and Denmark: A Descriptive Study. Lancet Infect. Dis. 2011, 11, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Shore, A.C.; Deasy, E.C.; Slickers, P.; Brennan, G.; O’Connell, B.; Monecke, S.; Ehricht, R.; Coleman, D.C. Detection of Staphylococcal Cassette Chromosome mec Type XI Carrying Highly Divergent mecA, mecI, mecR1, blaZ, and ccr Genes in Human Clinical Isolates of Clonal Complex 130 Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 3765–3773. [Google Scholar] [CrossRef] [Green Version]
- Larsen, J.; Raisen, C.L.; Ba, X.; Sadgrove, N.J.; Padilla-González, G.F.; Simmonds, M.S.J.; Bhattacharya, S.; Sandoval-Motta, S.; Gribble, F.M.; Reimann, F.; et al. Emergence of Methicillin Resistance Predates the Clinical Use of Antibiotics. Nature 2022, 602, 135–141. [Google Scholar] [CrossRef]
- Worthing, K.A.; Coombs, G.W.; Pang, S.; Abraham, S.; Saputra, S.; Trott, D.J.; Jordan, D.; Norris, J.M. Isolation of mecC MRSA in Australia. J. Antimicrob. Chemother. 2016, 71, 2348–2349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, T.; Hiramatsu, K.; Tomasz, A.; de Lencastre, H.; Perreten, V.; Holden, M.T.G.; Coleman, D.C.; Goering, R.; Giffard, P.M.; Skov, R.L.; et al. Guidelines for Reporting Novel mecA Gene Homologues. Antimicrob. Agents Chemother. 2012, 56, 4997–4999. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report: 2022; World Health Organization: Geneva, Switzerland, 2022; Available online: https://www.who.int/publications/i/item/9789240062702 (accessed on 5 March 2023).
- Falagas, M.E.; Karageorgopoulos, D.E.; Leptidis, J.; Korbila, I.P. MRSA in Africa: Filling the Global Map of Antimicrobial Resistance. PLoS ONE 2013, 8, e68024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thwala, T.; Madoroba, E.; Maliehe, T.S.; Magwedere, K.; Basson, A.K.; Butaye, P. Antimicrobial Resistance, Enterotoxin and mec Gene Profiles of Staphylococcus aureus Associated with Beef-Based Protein Sources from KwaZulu-Natal Province, South Africa. Microorganisms 2022, 10, 1199. [Google Scholar] [CrossRef] [PubMed]
- Akarsu, H.; Liljander, A.; Younan, M.; Brodard, I.; Overesch, G.; Glücks, I.; Labroussaa, F.; Kuhnert, P.; Perreten, V.; Monecke, S.; et al. Genomic Characterization and Antimicrobial Susceptibility of Dromedary-Associated Staphylococcaceae from the Horn of Africa. Appl. Environ. Microbiol. 2022, 88, e01146-22. [Google Scholar] [CrossRef] [PubMed]
- Harrison, E.M.; Paterson, G.K.; Holden, M.T.; Ba, X.; Rolo, J.; Morgan, F.J.; Pichon, B.; Kearns, A.; Zadoks, R.N.; Peacock, S.J.; et al. A Novel Hybrid SCCmec-mecC Region in Staphylococcus sciuri. J. Antimicrob. Chemother. 2014, 69, 911–918. [Google Scholar] [CrossRef]
- Agabou, A.; Ouchenane, Z.; Ngba Essebe, C.; Khemissi, S.; Chehboub, M.T.E.; Chehboub, I.B.; Sotto, A.; Dunyach-Remy, C.; Lavigne, J.P. Emergence of Nasal Carriage of ST80 and ST152 PVL+ Staphylococcus aureus Isolates from Livestock in Algeria. Toxins 2017, 9, 342. [Google Scholar] [CrossRef] [Green Version]
- Ben Chehida, F.; Gharsa, H.; Tombari, W.; Selmi, R.; Khaldi, S.; Daaloul, M.; Ben Slama, K.; Messadi, L. First Report of Antimicrobial Susceptibility and Virulence Gene Characterization Associated with Staphylococcus aureus Carriage in Healthy Camels from Tunisia. Animals 2021, 11, 2507. [Google Scholar] [CrossRef]
- Silva, V.; Caniça, M.; Manageiro, V.; Verbisck, N.; Tejedor-Junco, M.T.; González-Martin, M.; Fernandes, T.; Belas, A.; Borrego, N.; Simões, J.; et al. Staphylococcus aureus and Methicillin-Resistant Coagulase-Negative Staphylococci in Nostrils and Buccal Mucosa of Healthy Camels Used for Recreational Purposes. Animals 2022, 12, 10. [Google Scholar] [CrossRef]
- Yusuf, S.T.; Kwaga, J.K.P.; Okolocha, E.C.; Bello, M. Phenotypic Occurrence of Methicillin-Resistant Staphylococcus aureus in Camels Slaughtered at Kano Abattoir, Kano, Nigeria. Sokoto J. Vet. Sci. 2017, 15, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Alzohairy, M.A. Colonization and Antibiotic Susceptibility Pattern of Methicillin Resistance Staphylococcus aureus (MRSA) among Farm Animals in Saudi Arabia. J. Bacteriol. Res. 2011, 3, 63–68. [Google Scholar]
- Kaspar, U.; Kriegeskorte, A.; Schubert, T.; Peters, G.; Rudack, C.; Pieper, D.H.; Wos-Oxley, M.L.; Becker, K. The Culturome of the Human Nose Habitats Reveals Individual Bacterial Fingerprint Patterns. Environ. Microbiol. 2016, 18, 2130–2142. [Google Scholar] [CrossRef] [PubMed]
- Asante, J.; Amoako, D.G.; Abia, A.L.K.; Dadzie, I.; Sevilleja, E.A.; Antonio, M. Review of Clinically and Epidemiologically Relevant Coagulase-Negative Staphylococci in Africa. Microb. Drug Resist. 2020, 26, 951–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsubakishita, S.; Kuwahara-Arai, K.; Sasaki, T.; Hiramatsu, K. Origin and Molecular Evolution of the Determinant of Methicillin Resistance in Staphylococci. Antimicrob. Agents Chemother. 2010, 54, 4352–4359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loncaric, I.; Kübber-Heiss, A.; Posautz, A.; Ruppitsch, W.; Lepuschitz, S.; Schauer, B.; Feßler, A.T.; Krametter-Frötscher, R. Characterization of mecC Gene-Carrying Coagulase-Negative Staphylococcus spp. Isolated from Various Animals. Vet. Microbiol. 2019, 230, 138–144. [Google Scholar] [CrossRef]
- Dhaouadi, S.; Bouchami, O.; Soufi, L.; Dhaouadi, F.; Chaari, S.; Bouglita, W.; Slama, K.B.; Messadi, L.; Ben Haj Khalifa, A. Frequent Dissemination and Carriage of an SCCmec-mecC Hybrid in Methicillin-Resistant Mammaliicoccus sciuri in Farm Animals from Tunisia. J. Glob. Antimicrob. Resist. 2022, 31, 228–235. [Google Scholar] [CrossRef]
- Sekizuka, T.; Niwa, H.; Kinoshita, Y.; Uchida-Fujii, E.; Inamine, Y.; Hashino, M.; Kato, H.; Kuroda, M.; Iwaki, M.; Murakami, M.; et al. Identification of a mecA/mecC-Positive MRSA ST1-t127 Isolate from a Racehorse in Japan. J. Antimicrob. Chemother. 2020, 75, 292–295. [Google Scholar] [CrossRef]
- Larsen, J.; Stegger, M.; Andersen, P.S.; Petersen, A.; Larsen, A.R.; Westh, H.; Agersø, Y.; Fetsch, A.; Kraushaar, B.; Käsbohrer, A.; et al. Evidence for Human Adaptation and Foodborne Transmission of Livestock-Associated Methicillin-Resistant Staphylococcus aureus. Clin. Infect. Dis. 2016, 63, 1349–1352. [Google Scholar] [CrossRef] [Green Version]
- Sahin-Tóth, J.; Albert, E.; Juhász, A.; Ghidán, Á.; Juhász, J.; Horváth, A.; Kalmár, Z. Prevalence of Staphylococcus aureus in Wild Hedgehogs (Erinaceus europaeus) and First Report of mecC-MRSA in Hungary. Sci. Total Environ. 2022, 815, 152858. [Google Scholar] [CrossRef]
- Enger, H.; Larssen, K.W.; Damås, E.S.; Aamot, H.V.; Blomfeldt, A.; Elstrøm, P.; Stegger, M.; Johansen, H.K.; Borgen, K.; Sundsfjord, A.; et al. A Tale of Two STs: Molecular and Clinical Epidemiology of MRSA t304 in Norway 2008–2016. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 209. [Google Scholar] [CrossRef]
- Boswihi, S.S.; Udo, E.E.; Monecke, S.; Mathew, B.; Noronha, B.; Verghese, T.; Tappa, A.; Johny, M.; Isloor, S.; Alshrari, A.S.; et al. Emerging Variants of Methicillin-Resistant Staphylococcus aureus Genotypes in Kuwait Hospitals. PLoS ONE 2018, 13, e0195933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartels, M.D.; Worning, P.; Andersen, L.P.; Bes, M.; Enger, H.; Ås, C.G.; Bengtsson, B.; Grønlund, M.M.; Hellmark, B.; Kristinsson, K.G.; et al. Repeated Introduction and Spread of the MRSA Clone t304/ST6 in Northern Europe. Clin. Microbiol. Infect. 2021, 27, e1–e284. [Google Scholar] [CrossRef]
- Pauly, N.; Wichmann-Schauer, H.; Ballhausen, B.; Torres Reyes, N.; Fetsch, A.; Tenhagen, B.-A. Detection and Quantification of Methicillin-Resistant Staphylococcus aureus in Fresh Broiler Meat at Retail in Germany. Int. J. Food Microbiol. 2019, 292, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wu, C.; Wang, Y.; Fan, R.; Schwarz, S.; Zhang, S. Identification of Multiresistance Gene cfr in Methicillin-Resistant Staphylococcus aureus from Pigs: Plasmid Location and Integration into a Staphylococcal Cassette Chromosome mec Complex. Antimicrob. Agents Chemother. 2015, 59, 3641–3644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venla, J.; Ahmad, A.M.; Viivi, H.; Laura, L.; Saara, S.; Sanna, S.; Jaana, P.; Henri, V.; Eija, K.; Timo, S.; et al. Clonal Dissemination of Successful Emerging Clone mecA-MRSA t304/ST6 among Humans and Hedgehogs in the Helsinki Metropolitan Area in Finland. One Health 2023, 16, 100516. [Google Scholar] [CrossRef]
- Aro, T.; Kantele, A. High Rates of Meticillin-Resistant Staphylococcus aureus among Asylum Seekers and Refugees Admitted to Helsinki University Hospital, 2010 to 2017. Eurosurveillance 2018, 23, 180044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mairi, A.; Touati, A.; Pantel, A.; Zenati, K.; Martinez, A.Y.; Dunyach-Remy, C.; Touati, D.; Lavigne, J.P. Distribution of Toxinogenic Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus from Different Ecological Niches in Algeria. Toxins 2019, 11, 498. [Google Scholar] [CrossRef] [Green Version]
- Månsson, E.; Johannesen, T.B.; Nilsdotter-Augustinsson, Å.; Söderquist, B.; Stegger, M. Comparative genomics of Staphylococcus epidermidis from prosthetic-joint infections and nares highlights genetic traits associated with antimicrobial resistance, not virulence. Microb. Genom. 2021, 7, 000506. [Google Scholar] [CrossRef]
- Vitali, L.A.; Petrelli, D.; Lamikanra, A.; Prenna, M.; Akinkunmi, E.O. Diversity of Antibiotic Resistance Genes and Staphylococcal Cassette Chromosome mec Elements in Faecal Isolates of Coagulase-Negative Staphylococci from Nigeria. BMC Microbiol. 2014, 14, 106. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Shang, Q.; Li, W.; Guo, W.; Stojadinovic, A.; Mannion, C.; Bhattacharya, A.; Ding, X. Antibiotics for Cancer Treatment: A Double-Edged Sword. J. Cancer 2020, 11, 5135–5149. [Google Scholar] [CrossRef]
- Dortet, L.; Girlich, D.; Virlouvet, A.L.; Poirel, L.; Nordmann, P.; Iorga, B.I.; Naas, T. Characterization of BRPMBL, the Bleomycin Resistance Protein Associated with the Carbapenemase NDM. Antimicrob. Agents Chemother. 2017, 61, e02304-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderhaeghen, W.; Vandendriessche, S.; Crombé, F.; Dispas, M.; Denis, O.; Hermans, K.; Haesebrouck, F.; Butaye, P. Species and Staphylococcal Cassette Chromosome mec (SCCmec) Diversity among Methicillin-Resistant Non-Staphylococcus aureus Staphylococci Isolated from Pigs. Vet. Microbiol. 2012, 158, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Cameron, M.; Perry, J.; Middleton, J.R.; Chaffer, M.; Lewis, J.; Keefe, G.P. Short communication: Evaluation of MALDI-TOF mass spectrometry and a custom reference spectra expanded database for the identification of bovine-associated coagulase-negative staphylococci. J. Dairy Sci. 2018, 101, 590–595. [Google Scholar] [CrossRef]
- Vanderhaeghen, W.; Cerpentier, T.; Adriaensen, C.; Vicca, J.; Hermans, K.; Butaye, P. Methicillin-resistant Staphylococcus aureus (MRSA) ST398 associated with clinical and subclinical mastitis in Belgian cows. Vet. Microbiol. 2010, 144, 166–171. [Google Scholar] [CrossRef] [Green Version]
- de Melo, D.A.; Soares, B.S.; Motta, C.C.D.; Dubenczuck, F.C.; Barbieri, N.L.; Logue, C.M.; Oliveira, S.D. Accuracy of PCR universal primer for methicillin-resistant Staphylococcus and comparison of different phenotypic screening assays. Braz. J. Microbiol. 2020, 51, 403–407. [Google Scholar] [CrossRef]
- Bokma, J.; Vereecke, N.; Nauwynck, H.; Haesebrouck, F.; Theuns, S.; Pardon, B.; De Bleecker, K.; Boyen, F.; Maes, D. Genome-Wide Association Study Reveals Genetic Markers for Antimicrobial Resistance in Mycoplasma bovis. Microbiol. Spectr. 2021, 9, e0026221. [Google Scholar] [CrossRef] [PubMed]
- Vereecke, N.; Botteldoorn, N.; Brossé, C.; Bonckaert, C.; Nauwynck, H.; Haesebrouck, F.; Boyen, F.; Pasmans, F. Predictive Power of Long-Read Whole-Genome Sequencing for Rapid Diagnostics of Multidrug-Resistant Brachyspira hyodysenteriae Strains. Microbiol. Spectr. 2023, 11, e0412322. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Cerdeira, L.T.; Hawkey, J.; Méric, G.; Vezina, B.; Wyres, K.L.; Holt, K.E. Trycycler: Consensus Long-Read Assemblies for Bacterial Genomes. Genome Biol. 2021, 22, 266. [Google Scholar] [CrossRef] [PubMed]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of Long, Error-Prone Reads Using Repeat Graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Vaser, R.; Šikić, M. Time- and Memory-Efficient Genome Assembly with Raven. Nat. Comput. Sci. 2021, 1, 332–336. [Google Scholar] [CrossRef]
- Ruan, J.; Li, H. Fast and Accurate Long-Read Assembly with wtdbg2. Nat. Methods 2020, 17, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- RAST Annotation Server. RAST Server. Available online: https://rast.nmpdr.org/ (accessed on 19 October 2022).
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- VFDB: Virulence Factors of Bacterial Pathogens. Available online: http://www.mgc.ac.cn/cgi-bin/VFs/v5/main.cgi (accessed on 19 February 2023).




{kind=link}
{kind=link}
{kind=link}
| Staphylococcaceae Species | Number of Strains | Source |
|---|---|---|
| S. aureus | 32 | farm 1 (n = 9), farm 2 (n = 1), farm 3 (n = 8), farm 4 (n = 5), farm 5 (n = 2), farm 6 (n = 3), farm 7 (n = 4) |
| M. sciuri | 30 | farm 1 (n = 10), farm 2 (n = 3), farm 3 (n = 9), farm 5 (n = 3), farm 6 (n = 3), farm 7 (n = 2) |
| S. delphini | 18 | farm 1 (n = 1), farm 2 (n = 1), farm 3 (n = 8), farm 4 (n = 2), farm 5 (n = 1), farm 6 (n = 2), farm 7 (n = 3) |
| S. intermedius | 9 | farm 3 (n = 4), farm 5 (n = 2), farm 7 (n = 3) |
| S. simulans | 7 | farm 3 (n = 3), farm 4 (n = 3), farm 7 (n = 1) |
| S. agnetis | 4 | farm 1 (n = 1), farm 2 (n = 1), farm 3 (n = 1), farm 5 (n = 1) |
| S. epidermidis | 2 | Farm 3 (n = 1), farm 4 (n = 1) |
| S. hyicus | 8 | farm 1 (n = 2), farm 2 (n = 1), farm 6 (n = 3), farm 7 (n = 1) |
| S. chromogenes | 4 | farm 1 (n = 2), farm 3 (n = 1), farm 4 (n = 1) |
| S. lutrae | 1 | farm 5 (n = 1) |
| S. gallinarum | 1 | farm 5 (n = 1) |
| S. muscae | 2 | farm 3 (n = 2) |
| S. xylosus | 2 | farm 4 (n = 1), farm 7 (n = 1) |
| Biosamples Accessions Numbers | Species | Farm (Origin) | Region | Sequence Type ST * | SCCmec | β-Lactams | Other Antibiotic Resistance Genes |
|---|---|---|---|---|---|---|---|
| SAMN33407026 | M. lentus | 1 | Centre (M’sila) | NA | SCCmec VII -mecC | mecA, mecC, blaZ | str, erm(B), mph(C), tet(K) |
| SAMN33407029 | M. lentus | 1 | Centre (M’sila) | NA | SCCmec VII -mecC | mecA, mecC, blaZ | str, erm(B), mph(C), tet(K) |
| SAMN33407050 | M. lentus | 5 | South (Ouargla) | NA | SCCmec VII -mecC | mecA, mecC, blaZ | str, erm(B), mph(C), tet(K) |
| SAMN33407025 | M. lentus | 1 | Centre (M’sila) | NA | SCCmec VII -mecC | mecA, mecC, blaZ | str, erm(B), mph(C) |
| SAMN33407043 | M. lentus | 1 | Centre (M’sila) | NA | SCCmec VII -mecC | mecA, mecC, blaZ | erm(B), mph(C), tet(K) |
| SAMN33407027 | M. lentus | 1 | Centre (M’sila) | NA | SCCmec IIa | mecA | erm(A), mph(C) |
| SAMN33407030 | S. aureus | 4 | South (Ouargla) | ST6 | SCCmec IVa | mecA, blaZ | |
| SAMN33407032 | S. aureus | 4 | South (Ouargla) | ST6 | SCCmec IVa | mecA, blaZ | |
| SAMN33407033 | S. aureus | 5 | South (Ouargla) | ST6 | SCCmec IVa | mecA, blaZ | |
| SAMN33407028 | S. epidermidis | 1 | Centre (M’sila) | ST54 | SCCmec Vb | mecA, blaZ | tet(K), fosB, msr(A) |
| SAMN33407031 | S. epidermidis | 4 | South (Ouargla) | ST61 | SCCmec IIa | mecA, blaZ | tet(K), fosB, fusB |
| SAMN33407034 | S. epidermidis | 5 | South (Ouargla) | ST61 | SCCmec IIa | mecA, blaZ | tet(K), fosB, fusB |
| SAMN33407036 | S. epidermidis | 5 | South (Ouargla) | ST640 | SCCmec IVa | mecA, blaZ | msr(A), fosB |
| SAMN33407038 | S. epidermidis | 6 | South (Ouargla) | ST35 | SCCmec IIa | mecA, blaZ | tet(K), fosB, fusB, dfrG |
| SAMN33407049 | S. epidermidis | 5 | South (Ouargla) | ST61 | SCCmec IIa | mecA, blaZ | tet(K), fosB, fusB |
| SAMN33407035 | S. hominis | 5 | South (Ouargla) | ST1 | SCCmec VIII | mecA, blaZ | bleO, aadD, tet(K), fusC |
| Biosamples Accessions Numbers | Species | Adhesion | Exoenzymes | Immune Evasion | Effector Delivery | Exotoxins |
|---|---|---|---|---|---|---|
| SAMN33407030 | S. aureus | atl, ebh, cna, ebp, eap/map, efb, fnA, fnB, icaA, icaB, icaC, icaR, sdrC, sdrD, sdrE, spa, | sspB, sspC, hysA, geh, lip, sspA, coa, sak, nuc, | adsA, chp, scn, sbi | esaA, esaB, esaE, esaG, essA, essB, essC, esxA, esxB, esxC, esxD | hly/hla, hlb, hld, sea, hlgA, hlgB, hlgC, lukD |
| SAMN33407032 | S. aureus | atl, ebh, cna, ebp, eap/map, efb, fnA, fnB, icaA, icaB, icaC, icaR, sdrC, sdrD, sdrE, spa | sspB, sspC, hysA, geh, lip, sspA, coa, sak, nuc | adsA, chp, scn, sbi | esaA, esaB, esaE, esaG, essA, essB, essC, esxA, esxB, esxC, esxD | hly/hla, hlb, hld, sea, hlgA, hlgB, hlgC, lukD |
| SAMN33407033 | S. aureus | atl, ebh, cna, ebp, eap/map, efb, fnA, fnB, icaA, icaB, icaC, icaR, sdrC, sdrD, sdrE, spa | sspB, sspC, hysA, geh, lip, sspA, coa, sak, nuc | adsA, chp, scn, sbi | esaA, esaB, esaE, esaG, essA, essB, essC, esxA, esxB, esxC, esxD | hly/hla, hlb, hld, sea, hlgA, hlgB, hlgC, lukD |
| SAMN33407028 | S. epidermidis | atl, ebh, ebp, plr/gapA | sspB, geh, lip, sspA, nuc | hlb | ||
| SAMN33407031 | S. epidermidis | ebh, ebp, sdrE, sdrG, | sspB, geh, lip, sspA, nuc | hlb | ||
| SAMN33407034 | S. epidermidis | ebh, ebp, sdrC, sdrG | sspB, geh, lip, sspA, nuc, | hlb | ||
| SAMN33407036 | S. epidermidis | atl, ebh, ebp, sdrG | sspB, geh, lip, sspA, nuc | esaA, esaD, esaE, esaG, essA, essB, essC, esxA, esxB, esxC, esxD | hlb | |
| SAMN33407038 | S. epidermidis | atl, ebh, ebp, sdrE, sdrG, icaA, icaB, icaC, icaR, | sspB, geh, lip, sspA, nuc | hlb | ||
| SAMN33407049 | S. epidermidis | ebh, ebp, sdrE, sdrG | sspB, geh, lip, sspA, nuc | hlb |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belhout, C.; Boyen, F.; Vereecke, N.; Theuns, S.; Taibi, N.; Stegger, M.; de la Fé-Rodríguez, P.Y.; Bouayad, L.; Elgroud, R.; Butaye, P. Prevalence and Molecular Characterization of Methicillin-Resistant Staphylococci (MRS) and Mammaliicocci (MRM) in Dromedary Camels from Algeria: First Detection of SCCmec-mecC Hybrid in Methicillin-Resistant Mammaliicoccus lentus. Antibiotics 2023, 12, 674. https://doi.org/10.3390/antibiotics12040674
Belhout C, Boyen F, Vereecke N, Theuns S, Taibi N, Stegger M, de la Fé-Rodríguez PY, Bouayad L, Elgroud R, Butaye P. Prevalence and Molecular Characterization of Methicillin-Resistant Staphylococci (MRS) and Mammaliicocci (MRM) in Dromedary Camels from Algeria: First Detection of SCCmec-mecC Hybrid in Methicillin-Resistant Mammaliicoccus lentus. Antibiotics. 2023; 12(4):674. https://doi.org/10.3390/antibiotics12040674
Chicago/Turabian StyleBelhout, Chahrazed, Filip Boyen, Nick Vereecke, Sebastiaan Theuns, Nadia Taibi, Marc Stegger, Pedro Yoelvys de la Fé-Rodríguez, Leila Bouayad, Rachid Elgroud, and Patrick Butaye. 2023. "Prevalence and Molecular Characterization of Methicillin-Resistant Staphylococci (MRS) and Mammaliicocci (MRM) in Dromedary Camels from Algeria: First Detection of SCCmec-mecC Hybrid in Methicillin-Resistant Mammaliicoccus lentus" Antibiotics 12, no. 4: 674. https://doi.org/10.3390/antibiotics12040674