
Synergistic Antimicrobial Activity of Ceftriaxone and Polyalthia longifolia Methanol (MEPL) Leaf Extract against Methicillin-Resistant Staphylococcus aureus and Modulation of mecA Gene Presence

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Ceftriaxone and MEPL Antibacterial Activity against MRSA Isolates
2.2. Determination of the MIC and MBC Concentration of Ceftriaxone and MEPL against the MRSA Isolate
2.3. Synergistic Activity of Antibiotic with MEPL
- MIC of ceftriaxone alone = 8000 µg/mL
- MIC of ceftriaxone in combination = 1000 µg/mL
- MIC of MEPL alone = 16,000 µg/mL
- MIC of MEPL in combination = 2000 µg/mL
- FICceftriaxone = 1000 µg/mL ÷ 8000 µg/mL = 0.125
- FICMEPL = 2000 µg/mL ÷ 16000 µg/mL = 0.125
- ΣFIC = FICceftriaxone + FICMEPL
- = 0.125 + 0.125
- = 0.25
2.4. Presence of the mecA Gene in MRSA Treated with Different Combinations of MEPL and Ceftriaxone
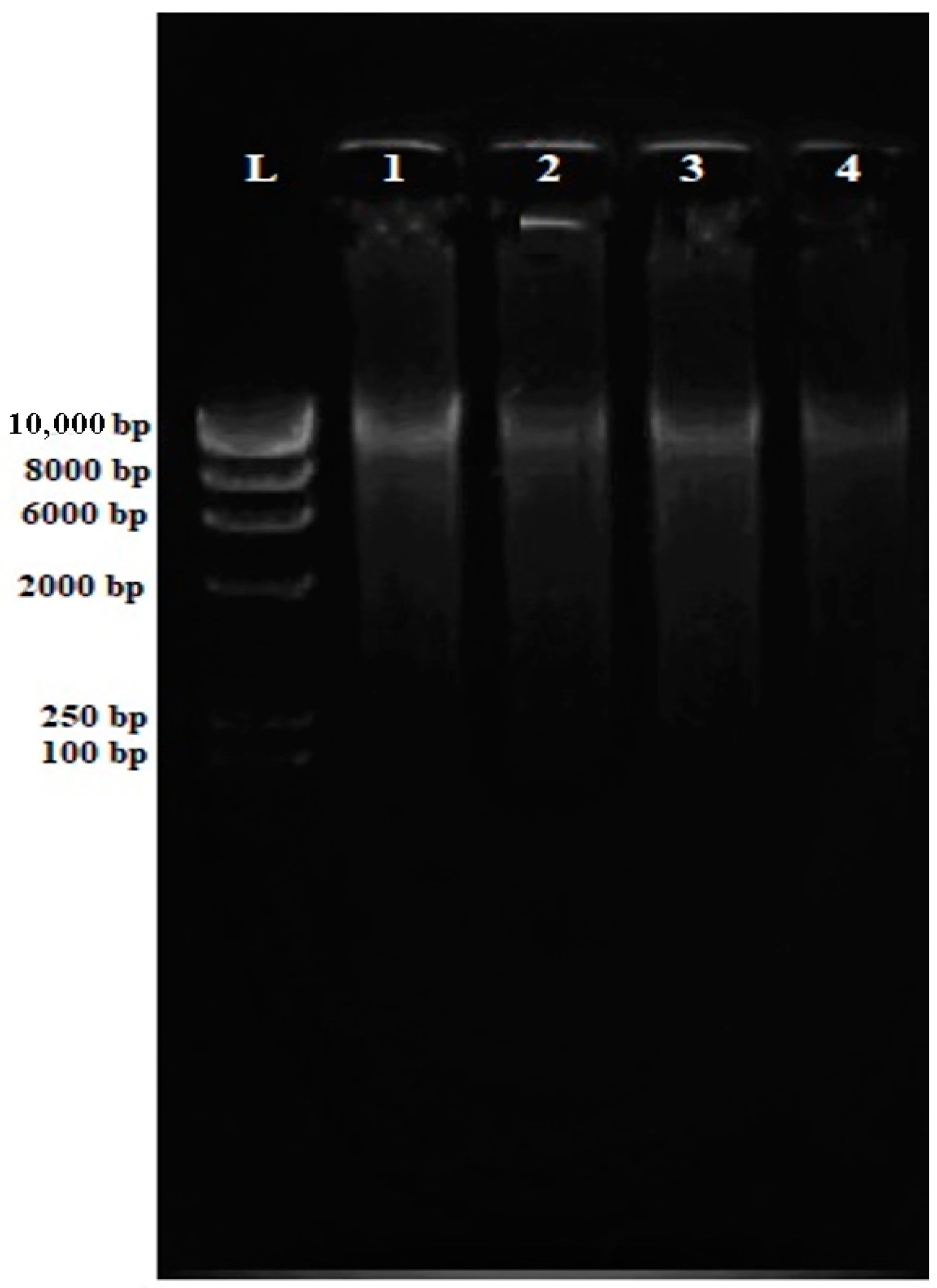
2.4.1. Purity of Genomic DNA
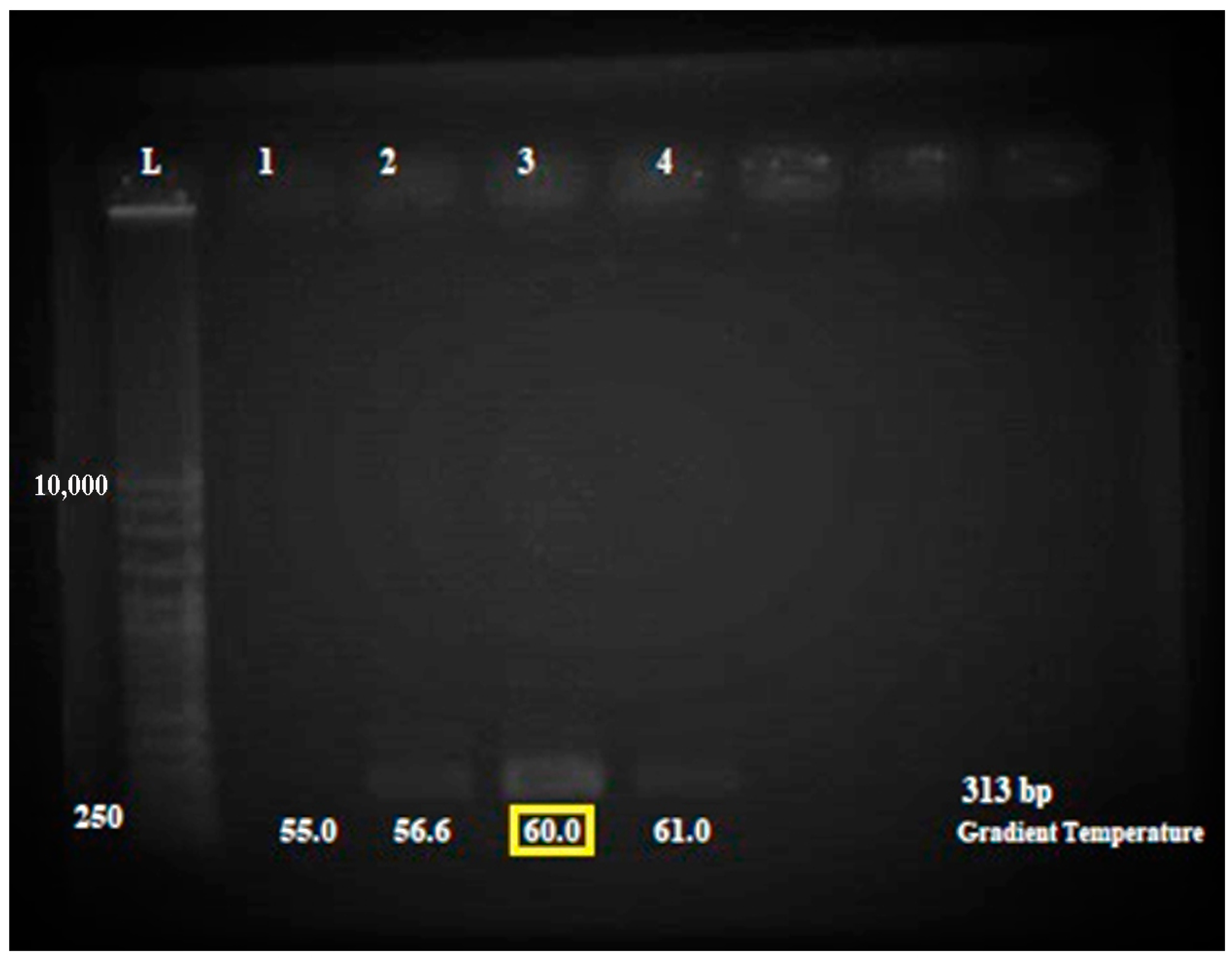
2.4.2. Optimization of mecA Gene Amplification
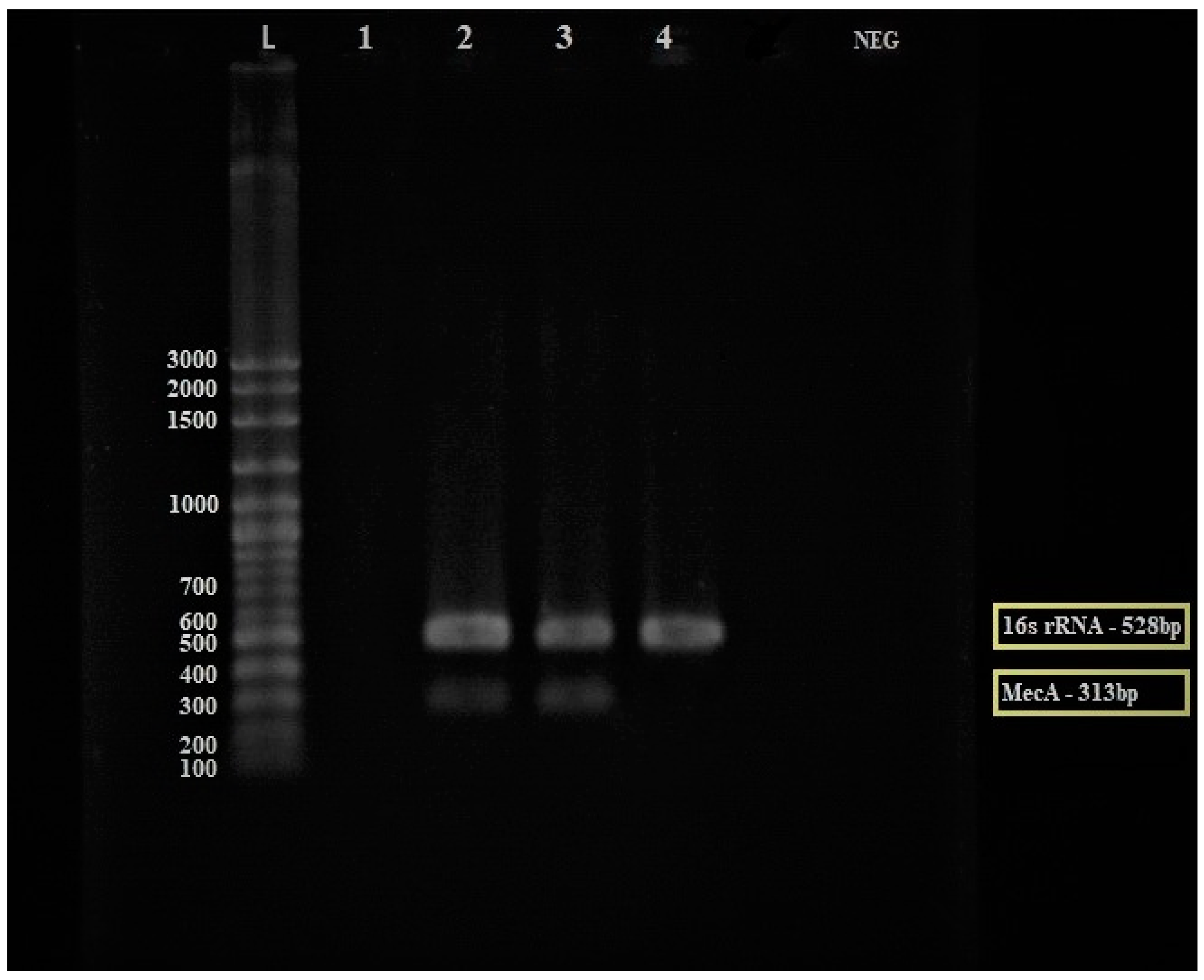
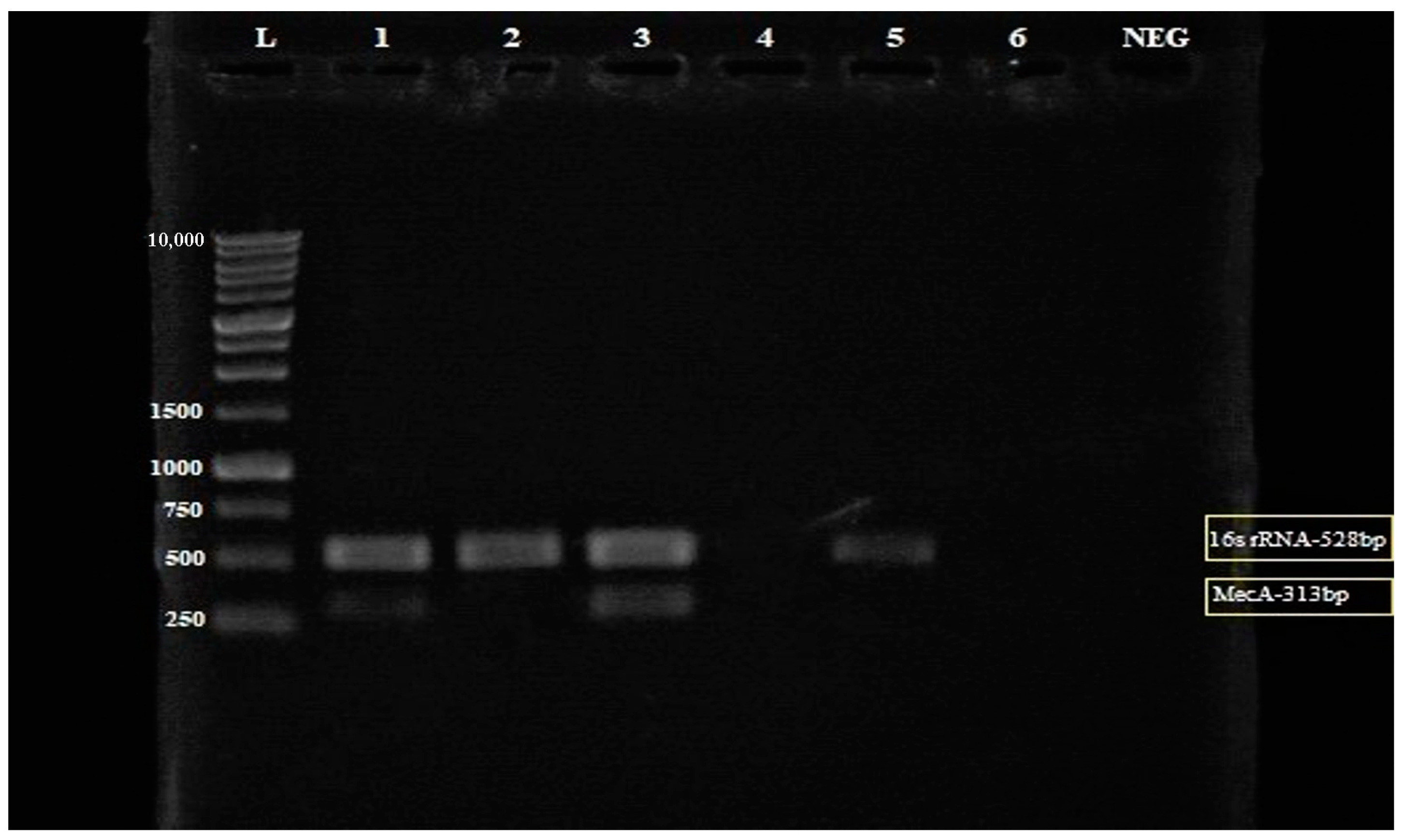
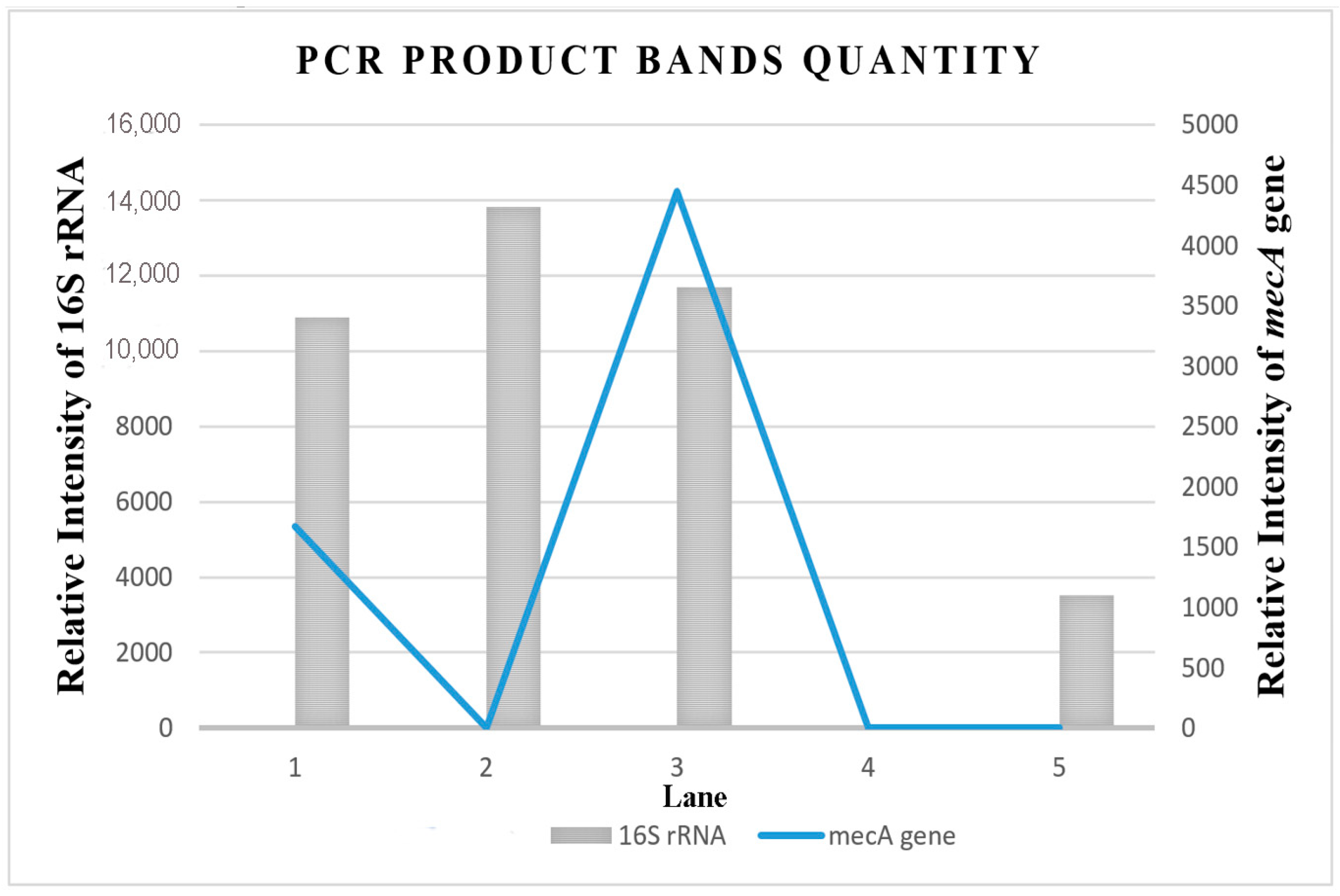
2.4.3. Detection of mecA Gene by Multiplex PCR
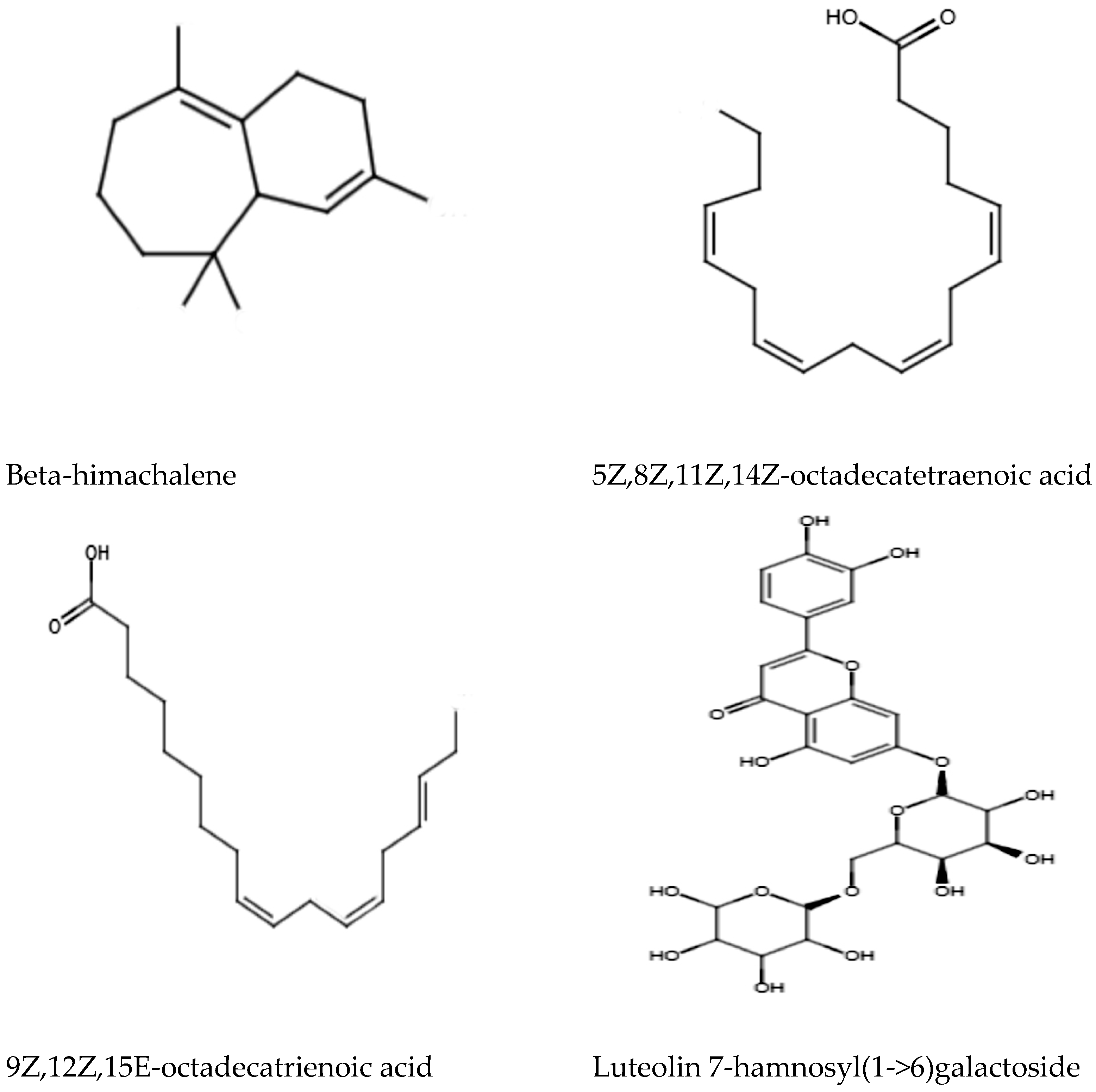
2.4.4. Antimicrobial Compounds in MEPL
3. Discussion
4. Materials and Methods
4.1. Polyalthia longifolia Leaf
4.2. Preparation of Polyalthia longifolia Leaf Extract
4.3. Test Microorganism Collection and Maintenance
Antimicrobial Susceptibility Test
4.4. Culture Media and Inoculum Preparation
4.5. Agar Disc Diffusion Assay of Ceftriaxone
4.6. Agar Well Diffusion Method of Antibacterial Susceptibility Test for MEPL
4.7. Evaluation of the Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) of the MRSA Isolate against Ceftriaxone and MEPL
4.7.1. Determination of the MIC of Ceftriaxone and MEPL against the MRSA Isolate
4.7.2. Determination of the MBC of Ceftriaxone and MEPL against the MRSA Isolate
4.8. Investigation of the Synergistic Properties of MEPL with Ceftriaxone
4.8.1. Preparation of MEPL and Ceftriaxone for Synergistic Study
4.8.2. Measurement of the Fractional Inhibitory Concentration (FIC) by Checkerboard Analysis
Calculation of the Fractional Inhibitory Concentration (FIC) Index
- FIC of plant extracts = MIC of MEPL in combination/MIC of MEPL alone
- FIC of antibiotic = MIC of antibiotic in combination/MIC of antibiotic alone
4.9. Presence of the mecA Gene in MRSA Treated with Different Combinations of MEPL and Ceftriaxone
4.9.1. Concentration-Dependent Assay of Ceftriaxone and MEPL against MRSA and MSSA Isolates
4.9.2. Genomic DNA Extraction
4.9.3. DNA Quantification
4.9.4. Multiplex Polymerase Chain Reaction (PCR)
4.9.5. Agarose Gel Electrophoresis Analysis of the PCR Product
4.10. LC–ESI–MS/MS Identification of Antimicrobial Compounds in MEPL
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garoy, E.Y.; Gebreab, Y.B.; Achila, O.O.; Tekeste, D.G.; Kesete, R.; Ghirmay, R.; Kiflay, R.; Tesfu, T. Methicillin-Resistant, Staphylococcus aureus (MRSA): Prevalence and Antimicrobial Sensitivity Pattern among Patients-A Multicenter Study in Asmara, Eritrea. Can. J. Infect. Dis. Med. Microbiol. 2019, 2019, 8321834. [Google Scholar] [CrossRef] [Green Version]
- Fair, R.J.; Tor, Y. Antibiotics and Bacterial Resistance in the 21st Century. Perspect. Med. Chem. 2014, 6, PMC-S14459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, D.C.; Chate, S.S. Study of antibiotic resistance pattern in methicillin resistant Staphylococcus aureus with special reference to newer antibiotic. J. Glob. Infect. Dis. 2015, 7, 78. [Google Scholar] [CrossRef]
- Wielders, C.L.; Fluit, A.C.; Brisse, S.; Verhoef, J.; Schmitz, F.J. mecA gene is widely disseminated in Staphylococcus aureus population. J. Clin. Microbiol. 2002, 40, 3970–3975. [Google Scholar] [CrossRef] [Green Version]
- Stapleton, P.D.; Taylor, P.W. Methicillin resistance in Staphylococcus aureus: Mechanisms and modulation. Sci Prog 2002, 85 Pt 1, 57–72. [Google Scholar] [CrossRef]
- Katkar, K.V.; Suthar, A.C.; Chauhan, V.S. The chemistry, pharmacologic, and therapeutic applications of Polyalthia longifolia. Pharmacogn. Rev. 2010, 4, 62. [Google Scholar] [PubMed] [Green Version]
- Parchman, M.L.; Munoz, A. Risk Factors for Methicillin-Resistant Staphylococcal aureus Skin and Soft Tissue Infections Presenting in Primary Care: A South Texas Ambulatory Research Network (STARNet) Study. J. Am. Board Fam. Med. 2009, 22, 375–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, K.W.; Zelbst, P.J.; Meacham, J.; Bhadauria, V.S. Green supply chain management practices: Impact on performance. Supply chain management Supply Chain Manag. 2012, 17, 290–305. [Google Scholar] [CrossRef]
- Dumyati, G.; Stevens, V.; Hannett, G.E.; Thompson, A.D.; Long, C.; MacCannell, D.; Limbago, B. Community-associated Clostridium difficile infections, Monroe County, New York, USA. Emerg. Infect. Dis. 2012, 18, 392. [Google Scholar] [CrossRef]
- Kirubakari, B.; Chen, Y.; Kanwar, J.R.; Shin, L.N.; Sasidharan, S. Studies on In Vitro Interaction of Ampicillin and Polyalthia longifolia Leaf Ethyl Acetate Fraction (PLEAF) by Checkerboard Method Against Methicillin Resistant Staphylococcus aureus (MRSA). Curr. Bioact. Compd. 2020, 16, 1049–1062. [Google Scholar] [CrossRef]
- Kirubakari, B.; Chen, Y.; Sasidharan, S. Synergistic effect of Polyalthia longifolia leaf and antibiotics against clinical isolates of methicillin-resistant Staphylococcus aureus (MRSA) by microscopic technique. Antiinflamm. Antiallergy Agents Med. Chem. 2020, 19, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Van Duin, D.; Paterson, D.L. Multidrug-resistant bacteria in the community: Trends and lessons learned. Dis. Clin. 2016, 30, 377–390. [Google Scholar]
- Gube, A.A.; Gonfa, R.; Tadesse, T. Evaluation of Antibiotic Use in Medical Ward of Fitche District Hospital, North Showa Zone, Oromia Region, Ethiopia. Adv. Pharmacoepidemiol. Drug Saf. 2017, 6, 217. [Google Scholar]
- Lee, H.; Jung, D.; Yeom, J.S.; Son, J.S.; Jung, S.I.; Kim, Y.S.; Kim, C.K.; Chang, H.H.; Kim, S.W.; Ki, H.K.; et al. Evaluation of ceftriaxone utilization at multicenter study. Korean J. Intern. Med. 2009, 24, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Richards, D.G.; Wolz, J.P.; Herman, L.M. Vocal mimicry of computer-generated sounds and vocal labeling of objects by a bottlenosed dolphin, Tursiops truncatus. J. Comp. Psychol. 1984, 98, 10. [Google Scholar] [CrossRef] [PubMed]
- Patel, U.C.; Mckissic, E.L.; Kasper, D.; Lentino, J.R.; Pachucki, C.T.; Lee, T.; Lopansri, B.K. Outcomes of ceftriax-one use compared to standard of therapy in Methicillin Susceptible Staphylococcal aureus (MSSA) blood-stream infections. Int. J. Clin. Pharm. 2014, 36, 1282–1289. [Google Scholar] [CrossRef] [PubMed]
- Kamfose, M.M.; Muriiti, F.G.; Knight, T.; Lasserson, D.; Hayward, G.J.A. Intravenous ceftriaxone versus multiple dosing regimens of intravenous anti-Staphylococcal antibiotics for Methicillin-Susceptible Staphylococcus aureus (MSSA): A Systematic Review. Antibiotic 2020, 9, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lother, S.A.; Press, N. Once-Daily Treatments for Methicillin-Susceptible Staphylococcus aureus Bacteremia: Are They Good Enough? Curr. Infect. Dis. Rep. 2017, 19, 43. [Google Scholar] [CrossRef] [PubMed]
- McGowan, J.E. Antimicrobial resistance in hospital organisms and its relation to antibiotic use. Rev. Infect. Dis. 1983, 5, 1033–1048. [Google Scholar] [CrossRef]
- Bbosa, G.; Mwebaza, N.; Odda, J.; Kyegombe, D.; Ntale, M. Antibiotics/antibacterial drug use, their marketing and promotion during the post-antibiotic golden age and their role in emergence of bacterial resistance. Health 2014, 6, 410–425. [Google Scholar] [CrossRef] [Green Version]
- Wooford, N.; Ellington, M.J. The emergence of antibiotic resistance by mutation. Clin. Microbiol. Infect. 2007, 13, 5–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Wintersdorff, C.J.; Penders, J.; van Niekerk, J.M.; Mills, N.D.; Majumder, S.; van Alphen, L.B.; Savelkoul, P.H.; Wolffs, P.F. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holcomb, H.G.; Durbin, K.J.; Cho, M.; Choi, K.J.; Darling, N.D.; Angerio, A.D. Methicillin-resistant Staphylococcus aureus as a threat to public health: A cellular approach. Georgetown Undergraduate. J. Health. Sci. 2008, 5, 2008. [Google Scholar]
- Al-Zoubi, M.S.; Al-Tayyar, I.A.; Hussein, E.; Jabali, A.A.; Khudairat, S. Antimicrobial susceptibility pattern of Staphylococcus aureus isolated from clinical specimens in Northern area of Jordan. Iran. J. Microbiol. 2015, 7, 265–272. [Google Scholar] [PubMed]
- Davis, J. Inactivation of antibiotics and dissemination of resistance genes. Science 1994, 264, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Mehmood, Z.; Mohammad, F. Screening of Some Indian Medicinal Plants for their Antimicrobial Properties. J. Ethnopharmacol. 1998, 62, 183–193. [Google Scholar] [CrossRef]
- Loper, J.E.; Henkels, M.D.; Roberts, R.G.; Grove, G.G.; Willett, M.J.; Smith, T.J. Evaluation of Streptomycin, Oxytetracycline and Copper Resistance of Erwinia amylavora isolated from pear orchards in Washington State. Plant Dis. 1991, 75, 287–290. [Google Scholar] [CrossRef]
- Service, R.F. Antibiotics That Resist Resistance. Science 1991, 270, 724–727. [Google Scholar] [CrossRef]
- Liu, C.S.; Cham, T.M.; Yang, C.H.; Chang, H.W.; Chen, C.H.; Chuang, L.Y. Antibacterial properties of Chinese herbal medicines against nosocomial antibiotic resistant strains of Pseudomonas aeruginosa in Taiwan. Am. J. Chin. Med. 2007, 35, 1047–1060. [Google Scholar] [CrossRef]
- Alexopoulos, A.; Kimbaris, A.C.; Plessas, S.; Mantzourani, I.; Theodoridou, I.; Stavropoulou, E.; Polissiou, M.G.; Bezirtzoglou, E. Antibacterial activities of essential oils from eight Greek aromatic plants against clinical isolates of Staphylococcus aureus. Anaerobe 2011, 17, 399–402. [Google Scholar] [CrossRef]
- Toroglu, S. In-vitro antimicrobial activity and synergistic/antagonistic effect of interactions between antibiotics and some spice essential oils. J. Environ. Biol. 2011, 32, 23–29. [Google Scholar] [PubMed]
- Chung, P.Y.; Navaratnam, P.; Chung, L.Y. Synergistic antimicrobial activity between pentacyclic triterpenoids and antibiotics against Staphylococcus aureus strains. Ann. Clin. Microbiol. Antimicrob. 2011, 10, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inui, T.; Wang, Y.; Deng, S.; Smith, D.C.; Franzblau, S.G.; Paul, G.F. Counter-current chromatography-based analysis of synergy in an anti-tuberculosis ethnobotanical. J. Chromatogr. A 2008, 1151, 211–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adwan, G.; Mhanna, M. Synergistic effects of plant extracts and antibiotics on Staphylococcus aureus strains isolated from clinical specimen. Asian Pac. J. Trop. Med. 2009, 2, 46–51. [Google Scholar]
- Ahmed, Z.; Khan, S.S.; Khan, M.; Tanveer, A.; Lone, Z.A. Synergistic effect of Salvadora persica extracts, tetracycline and penicillin against Staphylococcus aureus. Afr. J. Basic Appl. Sci. 2010, 2, 25–29. [Google Scholar]
- Aboulmagd, E.; Al-Mohammed, H.I.; Al-Badry, S. Synergism and post-antibiotic effect of green tea ex-tract and imipenem against methicillin-resistant Staphylococcus aureus. Microbiol. J. 2011, 1, 89–96. [Google Scholar] [CrossRef]
- Betoni, J.E.; Mantovani, R.P.; Barbosa, L.N.; Di Stasi, L.C.; Fernandes Junior, A. Synergism between plant ex-tract and antimicrobial drugs used on Staphylococcus aureus diseases. Mem. Inst. Oswaldo Cruz 2006, 101, 387–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, D.; Liu, Y.; Jiang, F.; Ji, F.; Wang, H.; Han, X. Synergistic Antibacterial Activity of Designed Trp-Containing Antibacterial Peptides in Combination with Antibiotics Against Multidrug-Resistant Staphylococcus epidermidis. Front. Microbiol. 2019, 10, 2719. [Google Scholar] [CrossRef] [Green Version]
- AL-Ali, K.; Abdelrazik, M.; Hemeg, H.; Ozbak, H. Antibacterial activity of four herbal extracts against methicillin resistant bacteria isolates collected from Almadinah Hospitals. Saudi Arabia. Int. J. Acad. Res. 2014, 2, 27–34. [Google Scholar]
- Chanda, S.; Nair, R. Antimicrobial activity of Polyalthia longifolia (sonn.) thw. var. pendula leaf extracts against 91 clinically important pathogenic microbial strains. Chin. Med. 2010, 1, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Phadnis, A.P.; Patwardhan, S.A.; Dhaneshwar, N.N. Clerodane diterpenoids from Polyalthia longifolia. Phytochemistry 1988, 27, 2899–2901. [Google Scholar] [CrossRef]
- Chaudhary, A.; Sood, S.; Das, P.; Kaur, P.; Mahajan, I.; Gulati, A.; Singh, B. Synthesis of novel antimicrobial aryl himachalene derivatives from naturally occurring himachalenes. EXCLI J. 2014, 13, 1216. [Google Scholar]
- Silva, A.; Silva, S.A.; Carpena, M.; Garcia-Oliveira, P.; Gullón, P.; Barroso, M.F.; Prieto, M.A.; Simal-Gandara, J. Macroalgae as a Source of Valuable Antimicrobial Compounds: Extraction and Applications. Antibiotics 2020, 9, 642. [Google Scholar] [CrossRef]
- Rahman, M.M.; Ahmad, S.H.; Mohamed, M.T.; Ab Rahman, M.Z. Antimicrobial compounds from leaf extracts of Jatropha curcas, Psidium guajava, and Andrographis paniculata. Sci. World J. 2014, 2014, 635240. [Google Scholar] [CrossRef] [Green Version]
- Qian, W.; Liu, M.; Fu, Y.; Zhang, J.; Liu, W.; Li, J.; Li, X.; Li, Y.; Wang, T. Antimicrobial mechanism of luteolin against Staphylococcus aureus and Listeria monocytogenes and its antibiofilm properties. Microb. Pathog. 2020, 142, 104056. [Google Scholar] [CrossRef] [PubMed]
- Arima, H.; Ashida, H.; Danno, G. Rutin enhanced antibacterial activities of flavonoids against Bacillus cereus and Salmonella enteritidis. Biosci. Biotechnol. Biochem. 2002, 66, 1009–1014. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.H.; Wang, M.J.; Chen, P.Y.; Wu, T.Y.; Wen, W.C.; Tsai, F.Y.; Lee, C.K. Constituents of Polyalthia longifolia var. pendula. J. Nat. Prod. 2009, 72, 1960–1963. [Google Scholar] [CrossRef]
- Murthy, M.M.; Subramanyam, M.; Bindu, M.H.; Annapurna, J. Antimicrobial activity of clerodane diterpenoids from Polyalthia longifolia seeds. Fitoterapia 2005, 76, 336–339. [Google Scholar] [CrossRef]
- Sashidhara, K.V.; Singh, S.P.; Sarkar, J.; Sinha, S.J. Cytotoxic clerodane diterpenoids from the leaves of Polyalthia longifolia. Nat. Prod. Res. 2010, 24, 1687–1694. [Google Scholar] [CrossRef]
- Nahari, D.S.; Prasetyawan, S.; Beltran, M.A.G.; Aulanni’am, A. Separation of flavonoids in the extract Polyalthia longifolia (Sonn.) Thw. leaves from Indonesia and the Philippines. J. Phys. Conf. Ser. 2019, 1374, 012001. [Google Scholar] [CrossRef]
- Gupta, V.K.; Tiwari, N.; Gupta, P.; Verma, S.; Pal, A.; Srivastava, S.K.; Darokar, M.P. A clerodane diterpene from Polyalthia longifolia as a modifying agent of the resistance of methicillin resistant Staphylococcus aureus. Phytomedicine 2016, 23, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Verma, S.; Pal, A.; Srivastava, S.K.; Srivastava, P.K.; Darokar, M.P. In vivo efficacy and synergistic interaction of 16α-hydroxycleroda-3, 13 (14) Z-dien-15, 16-olide, a clerodane diterpene from Polyalthia longifolia against methicillin-resistant Staphylococcus aureus. Appl. Microbiol. Biotechnol. 2013, 97, 9121–9131. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.M.; Roy, A.; Ghosh, A.K.; Hazra, T.K.; Basak, A.; Franco, O.L. Challenges and future prospects of antibiotic therapy: From peptides to phages utilization. Front. Pharmacol. 2014, 5, 105. [Google Scholar] [CrossRef]
- Li, X.Z.; Nikaido, H. Efflux-mediated drug resistance in bacteria: An update. Drugs 2009, 69, 1555–1623. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.W.; Kilic, A.; Yang, Q.; McAllister, S.K.; Li, H.; Miller, R.S.; McCormac, M.; Tracy, K.D.; Stratton, C.W.; Han, J.; et al. StaphPlex system for rapid and simultaneous identification of antibiotic resistance determinants and Panton-Valentine leukocidin detection of staphylococci from positive blood cultures. J. Clin. Microbiol. 2007, 45, 1867–1873. [Google Scholar] [CrossRef] [Green Version]
- Moisan, H.; Pruneau, M.; Malouin, F. Binding of ceftaroline to penicillin-binding proteins of Staphylococcus aureus and Streptococcus pneumoniae. J. Antimicrob. Chemother. 2010, 65, 713–716. [Google Scholar] [CrossRef]
- Gradelski, E.; Valera, L.; Aleksunes, L.; Bonner, D.; Fung-Tomc, J. Correlation between genotype and phenotypic categorization of Staphylococci based on methicillin susceptibility and resistance. J. Clin. Microbiol. 2001, 39, 2961–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, B.J.; Tomasz, A. Low-affinity penicillin-binding protein associated with beta-lactam resistance in Staphylococcus aureus. J. Bacteriol. 1984, 158, 513–516. [Google Scholar] [CrossRef] [Green Version]
- Özel, G.; Aslan, V.; Erdem, G.B.; Çağatay, M.; Sencan, I.; Mert, A. Comparison of oxacillin, cefoxitin, ceftizoxime, and moxalactam disk diffusion methods for detection of methicillin susceptibility in staphylococci. Mikrobiyoloji Bulteni 2011, 45, 258–265. [Google Scholar]
- Prasad, K.N.; Kumar, R.; Tiwari, D.P.; Mishra, K.K.; Ayyagari, A. Comparison of various conventional methods with a polymerase chain reaction assay for detecting methicillin-resistant & susceptible Staphylococcus aureus strains. Indian J. Med. Res. 2000, 112, 198–202. [Google Scholar]
- Mohd-Fuat, A.R.; Kofi, E.A.; Allan, G.G. Mutagenic and cytotoxic properties of three herbal plants from Southeast Asia. Trop. Biomed. 2007, 24, 49–59. [Google Scholar]
- Allen, J.R.F.; Holmstedt, B.R. The simple β-carboline alkaloids. Phytochemistry 1980, 19, 1573–1582. [Google Scholar] [CrossRef]
- Patel, K.; Gadewar, M.; Tripathi, R.; Prasad, S.; Patel, D.K. A review on medicinal importance, pharmacological activity and bioanalytical aspects of beta-carboline alkaloid “Harmine”. Asian Pac. J. Trop. Biomed. 2012, 2, 660–664. [Google Scholar] [CrossRef] [Green Version]
- Boeira, J.M.; Viana, A.F.; Picada, J.N.; Henriques, J.A. Genotoxic and recombinogenic activities of the two beta-carboline alkaloids harman and harmine in Saccharomyces cerevisiae. Mutat. Res. 2002, 500, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Rietjens, I.M.; Boersma, M.G.; Van Der Woude, H.; Jeurissen, S.M.; Schutte, M.E.; Alink, G.M.J.; Mutagenesis, M.M.O. Flavonoids and alkenylbenzenes: Mechanisms of mutagenic action and carcinogenic risk. Mutat. Res. 2005, 574, 124–138. [Google Scholar] [CrossRef] [PubMed]
- Faizi, S.; Khan, R.A.; Azher, S.; Khan, S.A.; Tauseef, S.; Ahmad, A. New antimicrobial alkaloids from the roots of Polyalthia longifolia var. pendula. Planta Med. 2003, 69, 350–355. [Google Scholar] [CrossRef]
- Jothy, S.L.; Chen, Y.; Kanwar, J.R.; Sasidharan, S. Evaluation of the Genotoxic Potential againstH2O2-Radical-Mediated DNA Damage and Acute Oral Toxicity of Standardized Extract of Polyalthia longifolia Leaf. Evid. Based Complement. Altern. Med. 2013, 2013, 925380. [Google Scholar] [CrossRef] [Green Version]
- Jothy, S.L.; Aziz, A.; Chen, Y.; Sasidharan, S. Antioxidant Activity and Hepatoprotective Potential of Polyalthia longifolia and Cassia spectabilis Leaves against Paracetamol-Induced Liver Injury. Evid. Based Complement. Altern. Med. 2012, 2012, 561284. [Google Scholar] [CrossRef] [Green Version]
- Jothy, S.L.; Saito, T.; Kanwar, J.R.; Chen, Y.; Aziz, A.; Yin-Hui, L.; Sasidharan, S. Radioprotective activity of Polyalthia longifolia standardized extract against X-ray radiation injury in mice. Phys. Med. 2016, 32, 150–161. [Google Scholar] [CrossRef]
- Mostafa, N.M.; Edmond, M.P.; El-Shazly, M.; Fahmy, H.A.; Sherif, N.H.; Singab, A.N.B. Phytoconstituents and renoprotective effect of Polyalthia longifolia leaves extract on radiation-induced nephritis in rats via TGF-β/smad pathway. Nat. Prod. Res. 2021, 36, 4187–4192. [Google Scholar] [CrossRef]
- Vijayarathna, S. Fundamental Studies on the Mechanism of Polyalthia longifolia (Sonn.) Thwaites Polyphenols Action in HeLa Cells in Relation to microRNA Regulation. PhD Thesis, Universiti Sains Malaysia, Pinang, Malaysia, 2017. [Google Scholar]
- Vijayarathna, S.; Chen, Y.; Kanwar, J.R.; Sasidharan, S. Standardized Polyalthia longifolia leaf extract (PLME) inhibits cell proliferation and promotes apoptosis: The anti-cancer study with various microscopy methods. Biomed. Pharmacother. 2017, 91, 366–377. [Google Scholar] [CrossRef] [PubMed]
- Vijayarathna, S.; Oon, C.E.; Chen, Y.; Kanwar, J.R.; Sasidharan, S. Polyalthia longifolia Methanolic Leaf Extracts (PLME) induce apoptosis, cell cycle arrest and mitochondrial potential depolarization by possibly modulating the redox status in hela cells. Biomed. Pharmacother. 2017, 89, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Vijayarathna, S.; Shanmugapriya Khanwar, J.R.; Sasidharan, S. Standardized Polyalthia longifolia methanolic leaf extracts (PLME) inhibits HeLa cells through inducing microRNAs expression and apoptosis. In Proceedings of the International Conference on Traditional & Alternative Medicine, Kerala, India, 27 September 2017. [Google Scholar]
- Hemagirri, M.; Sasidharan, S. In vitro antiaging activity of polyphenol rich Polyalthia longifolia (Annonaceae) leaf extract in Saccharomyces cerevisiae BY611 yeast cells. J. Ethnopharmacol. 2022, 290, 115110. [Google Scholar] [CrossRef]
- Vaghasiya, Y.; Patel, H.; Chanda, S. Antibacterial activity of methanol extract of Mangifera indica against some human pathogens and its phytochemical study. Afr. J. Biotechnol. 2011, 10, 15788–15794. [Google Scholar] [CrossRef]
- Manasa, M.; Vivek, M.N.; Yashoda, K.; Onkarappa, R.; Prashith, K.T.R. Antimicrobial activity of leaf and pericarp extracts of Polyalthia longifolia (Annonaceae). J. Pharm. Sci. 2004, 393, 221–225. [Google Scholar]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- CLSI Standard. M100 Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2020. [Google Scholar]
- Akinyemi, K.O.; Oladapo, O.; Okwara, C.E.; Ibe, C.C.; Fasure, K.A. Screening of crude extracts of six medicinal plants used in South-West Nigerian unorthodox medicine for anti-methicillin resistant Staphylococcus aureus activity. BMC Complement Altern. Med. 2005, 5, 6. [Google Scholar] [CrossRef]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48, 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampaio, F.C.; Pereira, M.d.S.V.; Dias, C.S.; Costa, V.C.; Conde, N.C.; Buzalaf, M.A. In vitro antimicrobial activity of Caesalpinia ferrea Martius fruits against oral pathogens. J. Ethnopharmacol. 2009, 124, 289–294. [Google Scholar] [CrossRef]
- Agarwal, A.; Jain, N.; Jain, A. Synergistic effect of cefixime and cloxacillin combination against common bacterial pathogens causing community acquired pneumonia. Indian J. Pharmacol. 2007, 39, 251–252. [Google Scholar]
- Zhao, W.H.; Hu, Z.Q.; Okubo, S.; Hara, Y.; Shimamura, T. Mechanism of Synergy between Epigallocatechin Gallate and b-Lactams against Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2001, 45, 1737–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration of Ceftriaxone (µg/mL) | Diameter of Zone of Inhibition (mm) | Concentration of MEPL (mg/mL) | Diameter of Zone of Inhibition (mm) |
|---|---|---|---|
| 8 | 0 | 1 | 21 ± 2 |
| 16 | 0 | 2 | 24 ± 1 |
| 32 | 0 | 3 | 26 ± 2 |
| 64 | 0 | 4 | 28 ± 2 |
| 5 | 29 ± 2 | ||
| 6 | 31 ± 1 | ||
| 7 | 32 ± 1 | ||
| 8 | 34 ± 1 |
| MEPL (μg/mL) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 16,000 | 8000 | 4000 | 2000 | 1000 | 500 | 250 | 125 | 62.5 | ||
| Ceftriaxone sodium (μg/mL) | 8000 | No growth | No growth | No growth | No growth | No growth | Mild growth | Heavy Growth | Heavy Growth | Heavy Growth |
| 4000 | No growth | No growth | No growth | No growth | Mild growth | Mild growth | Heavy Growth | Heavy Growth | Heavy Growth | |
| 2000 | No growth | No growth | No growth | No growth | Mild growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | |
| 1000 | No growth | No growth | No growth | No growth | Mild growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | |
| 500 | No growth | Mild Growth | Mild Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | |
| 250 | No growth | Mild Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | |
| 125 | Mild Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | |
| 62.5 | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | Heavy Growth | |
| Diameter of Zone of Inhibition (mm) | ||||
|---|---|---|---|---|
| Potency | Resistant | Intermediate | Sensitive | |
| Ceftriaxone | 30 µg | ≤13 | 14–20 | ≥21 |
| Primers | Oligonucleotide Primer Sequences (5′ to 3′) | Amplicon Size (bp) |
|---|---|---|
| mecA 761R | CTT GTA CCC AAT TTT GAT CCA TTT G | 313 |
| mecA 449F | AAA CTA CGG TAA CAT TGA TCG CAA | |
| 16S rRNA 914R | AAC CTT GCG GTC GTA CTC CC | 528 |
| 16S rRNA 387F | CGA AAG CCT GAC GGA GCA AC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ranjutha, V.; Chen, Y.; Al-Keridis, L.A.; Patel, M.; Alshammari, N.; Adnan, M.; Sahreen, S.; Gopinath, S.C.B.; Sasidharan, S. Synergistic Antimicrobial Activity of Ceftriaxone and Polyalthia longifolia Methanol (MEPL) Leaf Extract against Methicillin-Resistant Staphylococcus aureus and Modulation of mecA Gene Presence. Antibiotics 2023, 12, 477. https://doi.org/10.3390/antibiotics12030477
Ranjutha V, Chen Y, Al-Keridis LA, Patel M, Alshammari N, Adnan M, Sahreen S, Gopinath SCB, Sasidharan S. Synergistic Antimicrobial Activity of Ceftriaxone and Polyalthia longifolia Methanol (MEPL) Leaf Extract against Methicillin-Resistant Staphylococcus aureus and Modulation of mecA Gene Presence. Antibiotics. 2023; 12(3):477. https://doi.org/10.3390/antibiotics12030477
Chicago/Turabian StyleRanjutha, Valiappan, Yeng Chen, Lamya Ahmed Al-Keridis, Mitesh Patel, Nawaf Alshammari, Mohd Adnan, Sumaira Sahreen, Subash C. B. Gopinath, and Sreenivasan Sasidharan. 2023. "Synergistic Antimicrobial Activity of Ceftriaxone and Polyalthia longifolia Methanol (MEPL) Leaf Extract against Methicillin-Resistant Staphylococcus aureus and Modulation of mecA Gene Presence" Antibiotics 12, no. 3: 477. https://doi.org/10.3390/antibiotics12030477