
Diclofenac and Meloxicam Exhibited Anti-Virulence Activities Targeting Staphyloxanthin Production in Methicillin-Resistant Staphylococcus aureus

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. Identification of Clinical Isolates
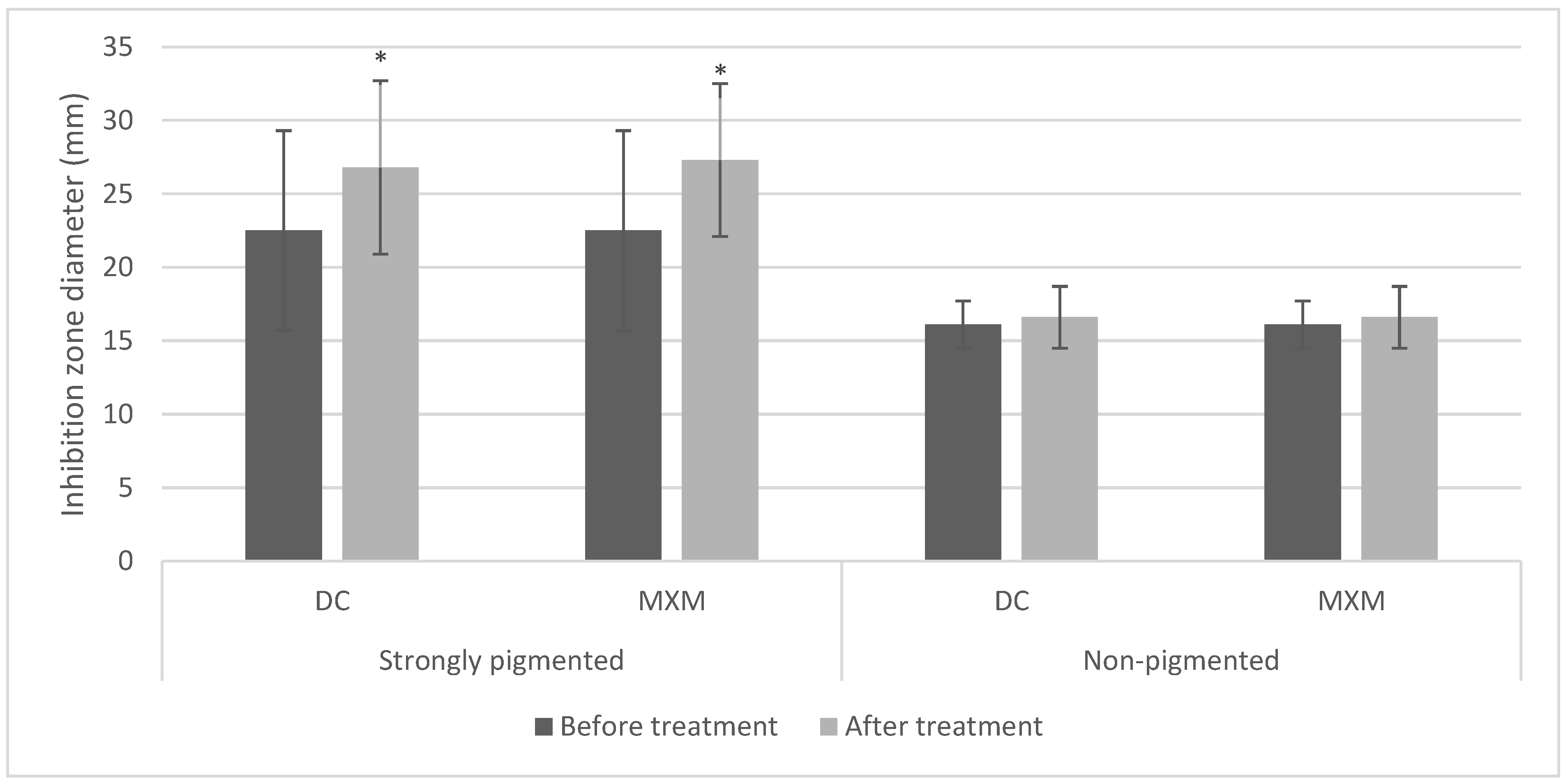
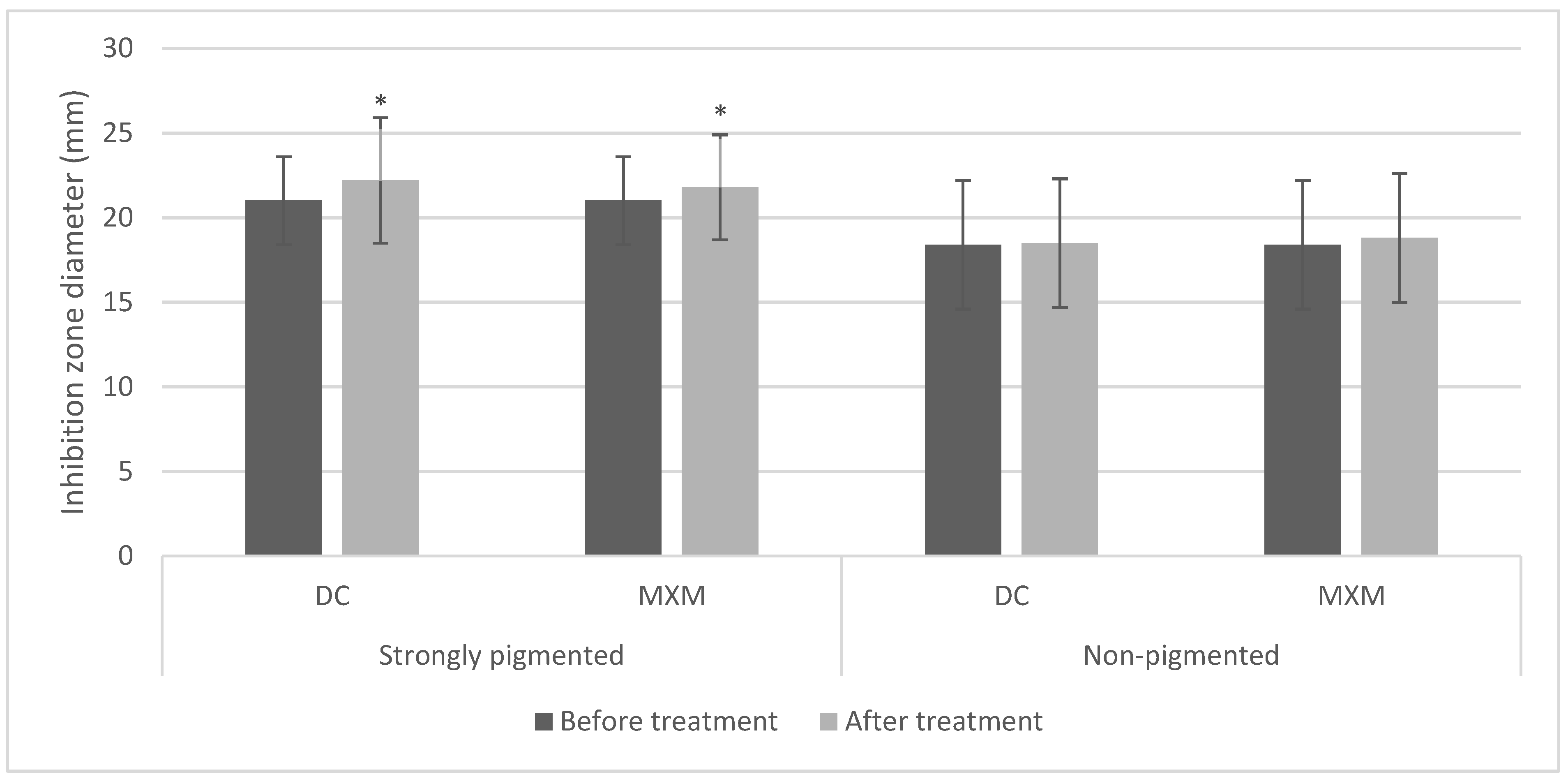
2.2. Antibiotic Resistance Pattern
2.3. Staphyloxanthin Production Assay
2.4. Quantitative Estimation of Staphyloxanthin Production
2.5. Determination of MICs of Tested Drugs
2.6. The Effect of the Tested Drugs on Staphyloxanthin Synthesis
2.7. Effect of Staphyloxanthin Inhibition on Protease Production
2.8. Effect of Staphyloxanthin Inhibition on DNase Activity
2.9. ERIC-PCR Analysis
2.10. Detection of mecA Gene among MRSA Isolates
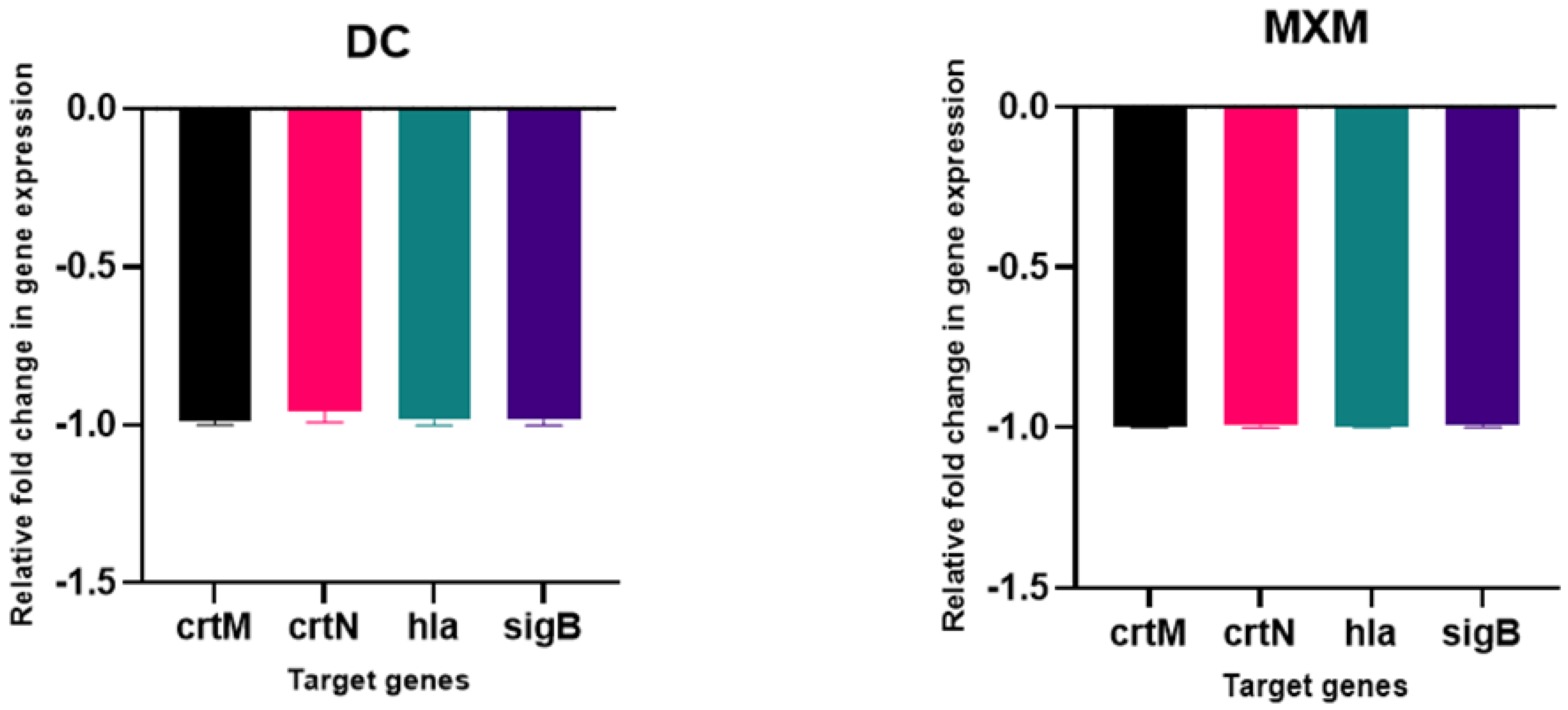
2.11. DC/MXM Treatment Effect on Virulent Genes Expression
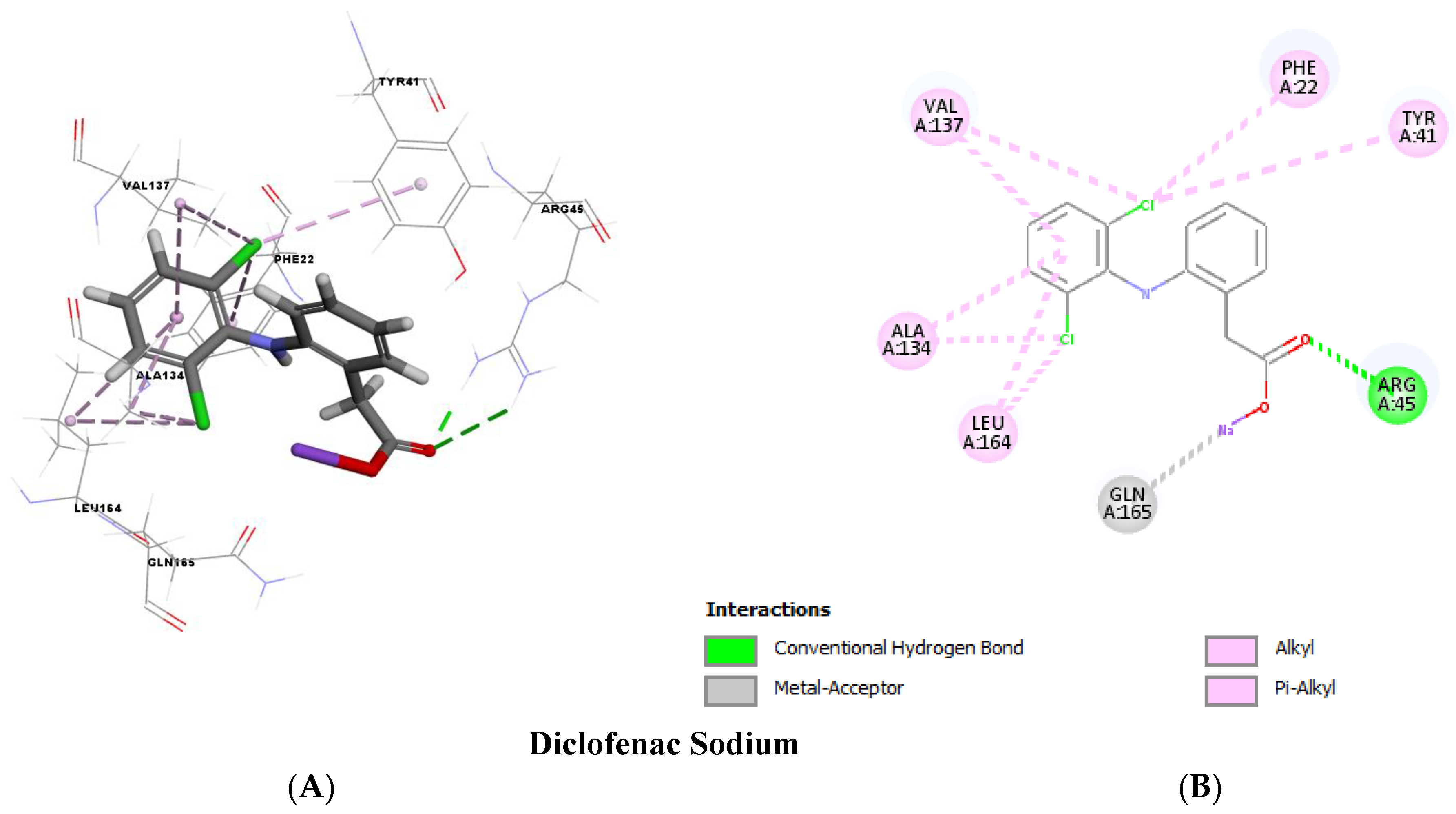
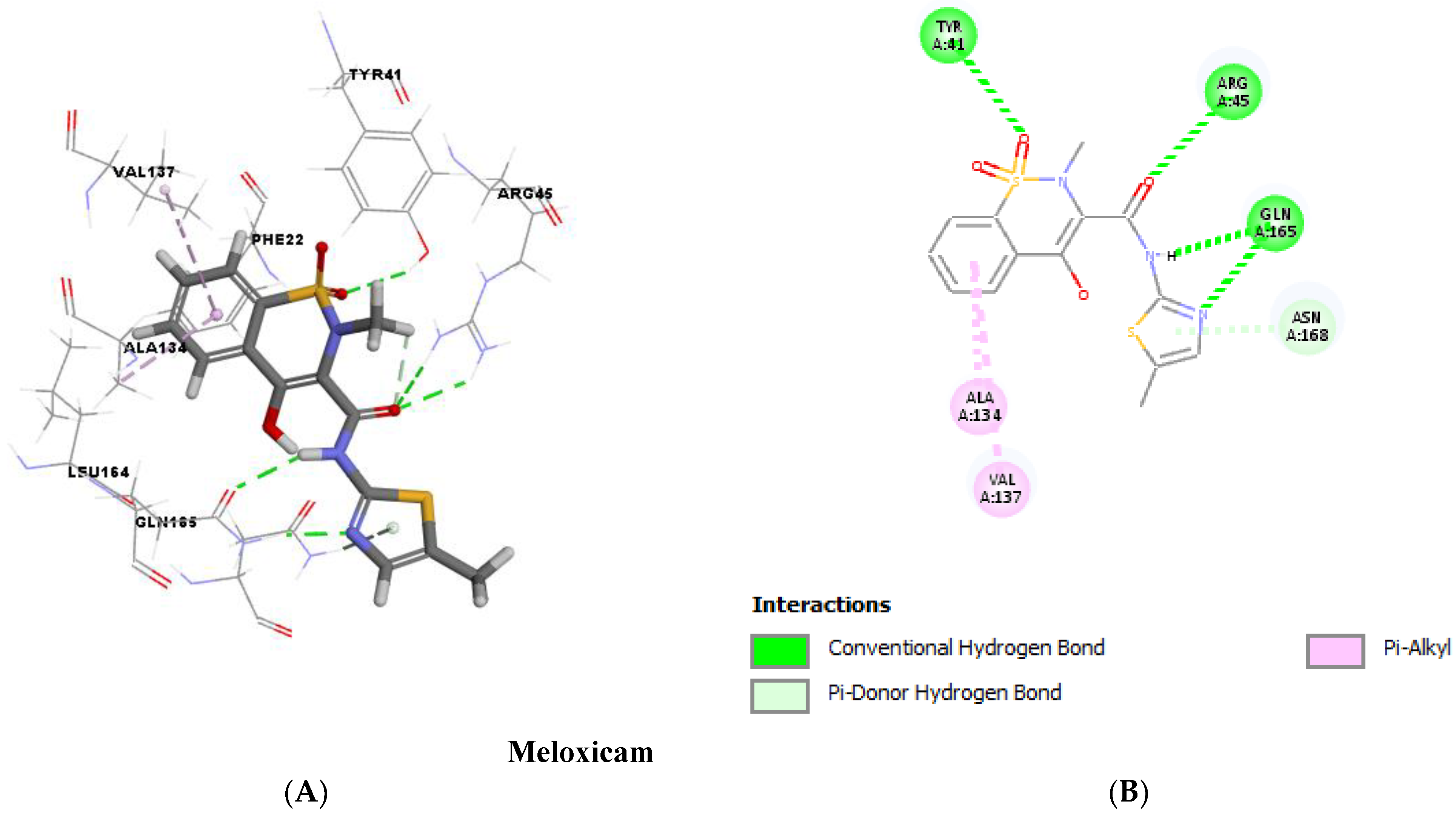
2.12. Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Stock Solutions
4.3. Antibiotic Susceptibility of Clinical Isolates
4.4. Staphyloxanthin Production Assay
4.5. Determination of MICs of Tested Drugs
4.6. Staphyloxanthin Inhibition Assay
4.7. Extracellular Protease Assay
4.8. Extracellular Nuclease Assay
4.9. Determination of Genetic Diversity Using ERIC-PCR
4.10. Detection of mecA Gene by cPCR
4.11. Real-Time PCR (qPCR) Analysis
4.12. Molecular Docking Analysis
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Troeman, D.; Van Hout, D.; Kluytmans, J.A.J.W. Antimicrobial approaches in the prevention of Staphylococcus aureus infections: A review. J. Antimicrob. Chemother. 2019, 74, 281–294. [Google Scholar] [CrossRef] [Green Version]
- Hiramatsu, K. The emergence of Staphylococcus aureus with reduced susceptibility to vancomycin in Japan. Am. J. Med. 1998, 104, 7S–10S. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Outbreaks of community-associated methicillin-resistant Staphylococcus aureus skin infections--Los Angeles County, California, 2002–2003. MMWR Morb. Mortal. Wkly Rep. 2003, 52, 88. Available online: https://pubmed.ncbi.nlm.nih.gov/12588006/ (accessed on 5 December 2022).
- Shrivastava, S.R.; Shrivastava, P.S.; Ramasamy, J. World health organization releases global priority list of antibiotic-resistant bacteria to guide research, discovery, and development of new antibiotics. J. Med. Soc. 2018, 32, 76. [Google Scholar] [CrossRef]
- Kong, C.; Neoh, H.-m.; Nathan, S. Targeting Staphylococcus aureus toxins: A potential form of anti-virulence therapy. J. Toxins 2016, 8, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Boudry, P.; Bohn, C.; Bouloc, P. Staphylococcus aureus pigmentation is not controlled by Hfq. BMC Res. Notes 2020, 13, 63. [Google Scholar] [CrossRef]
- Clauditz, A.; Resch, A.; Wieland, K.-P.; Peschel, A.; Götz, F. Staphyloxanthin plays a role in the fitness of Staphylococcus aureus and its ability to cope with oxidative stress. Infect. Immun. 2006, 74, 4950–4953. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.Y.; Essex, A.; Buchanan, J.T.; Datta, V.; Hoffman, H.M.; Bastian, J.F.; Fierer, J.; Nizet, V. Staphylococcus aureus golden pigment impairs neutrophil killing and promotes virulence through its antioxidant activity. J. Exp. Med. 2005, 202, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-I.; Liu, G.Y.; Song, Y.; Yin, F.; Hensler, M.E.; Jeng, W.-Y.; Nizet, V.; Wang, A.H.-J.; Oldfield, E. A cholesterol biosynthesis inhibitor blocks Staphylococcus aureus virulence. J. Sci. 2008, 319, 1391–1394. [Google Scholar] [CrossRef] [Green Version]
- Xue, L.; Chen, Y.Y.; Yan, Z.; Lu, W.; Wan, D.; Zhu, H. Staphyloxanthin: A potential target for antivirulence therapy. Infect. Drug Resist. 2019, 12, 2151. [Google Scholar] [CrossRef]
- Lin, F.-Y.; Liu, C.-I.; Liu, Y.-L.; Zhang, Y.; Wang, K.; Jeng, W.-Y.; Ko, T.-P.; Cao, R.; Wang, A.H.-J.; Oldfield, E. Mechanism of action and inhibition of dehydrosqualene synthase. Proc. Natl. Acad. Sci. USA 2010, 107, 21337–21342. [Google Scholar] [CrossRef] [PubMed]
- Raisig, A.; Sandmann, G. 4, 4′-Diapophytoene desaturase: Catalytic properties of an enzyme from the C30 carotenoid pathway of Staphylococcus aureus. J. Bacteriol. Res. 1999, 181, 6184–6187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Di, H.; Wang, Y.; Cao, Q.; Xu, B.; Zhang, X.; Yang, N.; Liu, G.; Yang, C.-G.; Xu, Y. Small-molecule targeting of a diapophytoene desaturase inhibits S. aureus virulence. Nat. Chem. Biol. 2016, 12, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Elmesseri, R.A.; Saleh, S.E.; Elsherif, H.M.; Yahia, I.S.; Aboshanab, K.M. Staphyloxanthin as a Potential Novel Target for Deciphering Promising Anti-Staphylococcus aureus Agents. J. Antibiot. Res. 2022, 11, 298. [Google Scholar] [CrossRef]
- Abbas, H.A.; Atallah, H.; El-Sayed, M.A.; El-Ganiny, A.M. Diclofenac mitigates virulence of multidrug-resistant Staphylococcus aureus. Arch. Microbiol. 2020, 202, 2751–2760. [Google Scholar] [CrossRef]
- Dutta, N.K.; Mazumdar, K.; Seok, S.-H.; Park, J.H. The anti-inflammatory drug Diclofenac retains anti-listerial activity in vivo. Appl. Microbiol. 2008, 47, 106–111. [Google Scholar] [CrossRef]
- Ghalehnoo, Z.R.; Rashki, A.; Najimi, M.; Dominguez, A. The role of diclofenac sodium in the dimorphic transition in Candida albicans. Microb. Pathog. 2010, 48, 110–115. [Google Scholar] [CrossRef]
- Mazumdar, K.; Dastidar, S.; Park, J.; Dutta, N.K. The anti-inflammatory non-antibiotic helper compound diclofenac: An antibacterial drug target. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 881–891. [Google Scholar] [CrossRef]
- She, P.; Wang, Y.; Luo, Z.; Chen, L.; Tan, R.; Wang, Y.; Wu, Y. Meloxicam inhibits biofilm formation and enhances antimicrobial agents efficacy by Pseudomonas aeruginosa. MicrobiologyOpen 2018, 7, e00545. [Google Scholar] [CrossRef] [Green Version]
- Alfeky, A.-A.E.; Tawfick, M.M.; Ashour, M.S.; El-Moghazy, A. High Prevalence of Multi-drug Resistant Methicillin-Resistant Staphylococcus aureus in Tertiary Egyptian Hospitals. J. Infect. Dev. Ctries. 2022, 16, 795–806. [Google Scholar] [CrossRef]
- Shorr, A.F. Epidemiology and economic impact of meticillin-resistant Staphylococcus aureus. Pharmacoeconomics 2007, 25, 751–768. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.S.; El-Baghdady, K.; El-All, A.; Said, M.; Warda, M.A.; Prince, A.M.; Ibrahim, M. Prevalence of multidrug resistance in the Egyptian methicillin-resistant Staphylococcus aureus isolates. Afr. J. Biol. Sci. 2020, 16, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Koupahi, H.; Jahromy, S.H.; Rahbar, M. Evaluation of different phenotypic and genotypic methods for detection of methicillin resistant Staphylococcus aureus (MRSA). Iran. J. Pathol. 2016, 11, 370. [Google Scholar] [PubMed]
- Shady, H.M.A.; Bakr, A.E.A.; Hashad, M.E.; Alzohairy, M.A. Staphylococcus aureus nasal carriage among outpatients attending primary health care centers: A comparative study of two cities in Saudi Arabia and Egypt. Braz. J. Microbiol. 2015, 19, 68–76. [Google Scholar]
- Liu, F.; Rajabi, S.; Shi, C.; Afifirad, G.; Omidi, N.; Kouhsari, E.; Khoshnood, S.; Azizian, K. Antibacterial activity of recently approved antibiotics against methicillin-resistant Staphylococcus aureus (MRSA) strains: A systematic review and meta-analysis. Ann. Clin. Microbiol. Antimicrob. 2022, 21, 37. [Google Scholar] [CrossRef]
- Kishk, R.M.; Anani, M.M.; Nemr, N.A.; Soliman, N.M.; Fouad, M.M. Inducible clindamycin resistance in clinical isolates of Staphylococcus aureus in Suez Canal University Hospital, Ismailia, Egypt. J. Infect. Dev. Ctries. 2020, 14, 1281–1287. [Google Scholar] [CrossRef]
- Hashem, R.A.; Yassin, A.S.; Zedan, H.H.; Amin, M.A. Fluoroquinolone resistant mechanisms in methicillin-resistant Staphylococcus aureus clinical isolates in Cairo, Egypt. J. Infect. Dev. Ctries. 2013, 7, 796–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alseqely, M.; Newton-Foot, M.; Khalil, A.; El-Nakeeb, M.; Whitelaw, A.; Abouelfetouh, A. Association between fluoroquinolone resistance and MRSA genotype in Alexandria, Egypt. Sci. Rep. 2021, 11, 4253. [Google Scholar] [CrossRef]
- Rasmi, A.H.; Ahmed, E.F.; Darwish, A.M.A.; Gad, G.F.M. Virulence genes distributed among Staphylococcus aureus causing wound infections and their correlation to antibiotic resistance. BMC Infect. 2022, 22, 652. [Google Scholar] [CrossRef]
- Zhang, J.; Suo, Y.; Zhang, D.; Jin, F.; Zhao, H.; Shi, C. Genetic and virulent difference between pigmented and non-pigmented Staphylococcus aureus. Front. Microbiol. 2018, 9, 598. [Google Scholar] [CrossRef]
- Askoura, M.; Saleh, M.; Abbas, H. An innovative role for tenoxicam as a quorum sensing inhibitor in Pseudomonas aeruginosa. Arch. Microbiol. 2020, 202, 555–565. [Google Scholar] [CrossRef] [PubMed]
- El-Ganiny, A.M.; Gad, A.I.; El-Sayed, M.A.; Shaldam, M.A.; Abbas, H.A. The promising anti-virulence activity of candesartan, domperidone, and miconazole on Staphylococcus aureus. Braz. J. Microbiol. 2022, 53, 1–18. [Google Scholar] [CrossRef]
- Selvaraj, A.; Jayasree, T.; Valliammai, A.; Pandian, S.K. Myrtenol attenuates MRSA biofilm and virulence by suppressing sarA expression dynamism. Front. Microbiol. 2019, 10, 2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valliammai, A.; Selvaraj, A.; Muthuramalingam, P.; Priya, A.; Ramesh, M.; Pandian, S.K. Staphyloxanthin inhibitory potential of thymol impairs antioxidant fitness, enhances neutrophil mediated killing and alters membrane fluidity of methicillin resistant Staphylococcus aureus. Biomed. Pharmacother. 2021, 141, 111933. [Google Scholar] [CrossRef]
- Valliammai, A.; Sethupathy, S.; Priya, A.; Selvaraj, A.; Bhaskar, J.P.; Krishnan, V.; Pandian, S.K. 5-Dodecanolide interferes with biofilm formation and reduces the virulence of Methicillin-resistant Staphylococcus aureus (MRSA) through up regulation of agr system. Sci. Rep. 2019, 9, 13744. [Google Scholar] [CrossRef] [Green Version]
- Vijayakumar, K.; Muhilvannan, S.; Arun Vignesh, M. Hesperidin inhibits biofilm formation, virulence and staphyloxanthin synthesis in methicillin resistant Staphylococcus aureus by targeting SarA and CrtM: An in vitro and in silico approach. World J. Microbiol. Biotechnol. 2022, 38, 44. [Google Scholar] [CrossRef] [PubMed]
- Abbas, H.A.; Elsherbini, A.M.; Shaldam, M.A. Glyceryl trinitrate blocks staphyloxanthin and biofilm formation in Staphylococcus aureus. Afr. Health Sci. 2019, 19, 1376–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boles, B.R.; Horswill, A.R. agr-Mediated dispersal of Staphylococcus aureus biofilms. PLoS Pathog. 2008, 4, e1000052. [Google Scholar] [CrossRef]
- Deniz Candan, E.; Idil, N.; Seyis Bilkay, I. Usefulness of REP and ERIC-PCR combination for tracking the spread of Staphylococcus aureus strains. Minerva Biotecnol. 2013, 25, 245–250. [Google Scholar]
- Hunter, P.R. Reproducibility and indices of discriminatory power of microbial typing methods. J. Clin. Microbiol. 1990, 28, 1903–1905. [Google Scholar] [CrossRef] [Green Version]
- Selvaraj, A.; Valliammai, A.; Muthuramalingam, P.; Priya, A.; Suba, M.; Ramesh, M.; Karutha Pandian, S. Carvacrol targets SarA and CrtM of methicillin-resistant Staphylococcus aureus to mitigate biofilm formation and staphyloxanthin synthesis: An in vitro and in vivo approach. ACS Omega 2020, 5, 31100–31114. [Google Scholar] [CrossRef] [PubMed]
- Yehia, F.A.-z.A.; Yousef, N.; Askoura, M. Celastrol mitigates staphyloxanthin biosynthesis and biofilm formation in Staphylococcus aureus via targeting key regulators of virulence; in vitro and in vivo approach. BMC Microbiol. 2022, 22, 106. [Google Scholar] [CrossRef] [PubMed]
- Khodaverdian, V.; Pesho, M.; Truitt, B.; Bollinger, L.; Patel, P.; Nithianantham, S.; Yu, G.; Delaney, E.; Jankowsky, E.; Shoham, M. Discovery of antivirulence agents against methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2013, 57, 3645–3652. [Google Scholar] [CrossRef] [PubMed]
- Procop, G.W.; Church, D.L.; Hall, G.S.; Janda, W.M. Koneman’s Color Atlas and Textbook of Diagnostic Microbiology, 7th ed.; Jones & Bartlett Publishers: Burlington, MA, USA, 2020; pp. 670–732. [Google Scholar]
- Satlin, M.J.; Lewis, J.S.; Weinstein, M.P.; Patel, J.; Humphries, R.M.; Kahlmeter, G.; Giske, C.G.; Turnidge, J. Clinical and Laboratory Standards Institute and European Committee on Antimicrobial Susceptibility Testing position statements on polymyxin B and colistin clinical breakpoints. Arch. Clin. Infect. Dis. 2020, 71, e523–e529. [Google Scholar] [CrossRef]
- Zhang, S.; Qu, X.; Tang, H.; Wang, Y.; Yang, H.; Yuan, W.; Yue, B. Diclofenac Resensitizes Methicillin-Resistant Staphylococcus aureus to β-Lactams and Prevents Implant Infections. Adv. Sci. Lett. 2021, 8, 2100681. [Google Scholar] [CrossRef]
- Sakai, K.; Koyama, N.; Fukuda, T.; Mori, Y.; Onaka, H.; Tomoda, H. Search method for inhibitors of staphyloxanthin production by methicillin-resistant Staphylococcus aureus. Biol. Pharm. Bull. 2012, 35, 48–53. [Google Scholar] [CrossRef] [Green Version]
- AL-Kazaz, E.J.; Melconian, A.K.; Kandela, N.J. Extraction of Staphyloxanthin from Staphylococcus aureus isolated from clinical sources to determine its antibacterial activity against other bacteria. Iraqi J. Sci. 2014, 55, 1823–1832. [Google Scholar]
- Lagacé-Wiens, P.R.; Alfa, M.J.; Manickam, K.; Karlowsky, J.A. Thermostable DNase is superior to tube coagulase for direct detection of Staphylococcus aureus in positive blood cultures. J. Clin. Microbiol. 2007, 45, 3478–3479. [Google Scholar] [CrossRef] [Green Version]
- Versalovic, J.; Koeuth, T.; Lupski, R. Distribution of repetitive DNA sequences in eubacteria and application to finerpriting of bacterial enomes. Nucleic Acids Res. 1991, 19, 6823–6831. [Google Scholar] [CrossRef]
- Anjay, A.K.; Agarwal, R.; Ramees, T.; Dubal, Z.; Kaushik, P.; Kumar, M.S.; Dudhe, N.; Milton, A.; Abhishek, B.; Shagufta, B. Molecular typing of Salmonella typhimurium and S. enteritidis serovars from diverse origin by ERIC-PCR. J. Pure Appl. Microbiol. 2015, 9, 2627–2634. [Google Scholar]
- McClure, J.-A.; Conly, J.M.; Lau, V.; Elsayed, S.; Louie, T.; Hutchins, W.; Zhang, K. Novel multiplex PCR assay for detection of the staphylococcal virulence marker Panton-Valentine leukocidin genes and simultaneous discrimination of methicillin-susceptible from-resistant staphylococci. J. Clin. Microbiol. 2006, 44, 1141–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Agents | Susceptibility Pattern of the Clinical Isolates, (N = 80) | |||||
|---|---|---|---|---|---|---|
| Sensitive (S) | Intermediate (I) | Resistant (R) | ||||
| N | % | N | % | N | % | |
| Cefoxitin | 0 | 0 | 0 | 0 | 80 | 100 |
| Clindamycin | 50 | 62.5 | 1 | 1.25 | 29 | 36.25 |
| Ciprofloxacin | 44 | 55 | 3 | 3.75 | 33 | 41.25 |
| Linezolid | 80 | 100 | 0 | 0 | 0 | 0 |
| Vancomycin | 80 | 100 | 0 | 0 | 0 | 0 |
| STX Inhibitors | cDOCKER Energy (Kcal/mol) | cDOCKER Interaction Energy (Kcal/mol) | Interactions with Amino Acid Residues of CrtM | |
|---|---|---|---|---|
| Diclofenac | −28.5335 | −36.6979 | Hydrogen bonds | Hydrophobic interactions |
| 2 H-bonds with ARG45 | 8 hydrophobic bonds with PHE22, TYR41, ALA134, VAL137 and LEU164 | |||
| Meloxicam | −23.4162 | −40.4063 | 6 H-bonds with TYR41, ARG45, GLN165 and ASN168 | 2 hydrophobic bonds with ALA134 and VAL137 |
| Target Gene | Primer Sequence (5′–3′) |
|---|---|
| crtM | F-CTGCTAATTCTATGATTGGTTGTGC |
| R-TGGGAATATTATGCAGCTATMGCAG | |
| crtN | F-GATGAAGCTTTGACGCAACA |
| R-TTCGCATGATACGTTTGCTC | |
| hla | F-GAA AGG TAC CAT TGC TGG TCA |
| R-AAG GCC AGG CTA AAC CAC TT | |
| sigB | F-AAG TGA TTC GTA AGG ACG TCT |
| R-TCG ATA ACT ATA ACC AAA GCC T |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elmesseri, R.A.; Saleh, S.E.; Ghobish, S.A.; Majrashi, T.A.; Elsherif, H.M.; Aboshanab, K.M. Diclofenac and Meloxicam Exhibited Anti-Virulence Activities Targeting Staphyloxanthin Production in Methicillin-Resistant Staphylococcus aureus. Antibiotics 2023, 12, 277. https://doi.org/10.3390/antibiotics12020277
Elmesseri RA, Saleh SE, Ghobish SA, Majrashi TA, Elsherif HM, Aboshanab KM. Diclofenac and Meloxicam Exhibited Anti-Virulence Activities Targeting Staphyloxanthin Production in Methicillin-Resistant Staphylococcus aureus. Antibiotics. 2023; 12(2):277. https://doi.org/10.3390/antibiotics12020277
Chicago/Turabian StyleElmesseri, Rana A., Sarra E. Saleh, Sarah A. Ghobish, Taghreed A. Majrashi, Heba M. Elsherif, and Khaled M. Aboshanab. 2023. "Diclofenac and Meloxicam Exhibited Anti-Virulence Activities Targeting Staphyloxanthin Production in Methicillin-Resistant Staphylococcus aureus" Antibiotics 12, no. 2: 277. https://doi.org/10.3390/antibiotics12020277