
Staphylococcus Infection: Relapsing Atopic Dermatitis and Microbial Restoration

Gachon Bio-Nano Institute, Gachon University, Seongnam-si 461-701, Republic of Korea
Antibiotics 2023, 12(2), 222; https://doi.org/10.3390/antibiotics12020222
Submission received: 15 December 2022
/
Revised: 15 January 2023
/
Accepted: 18 January 2023
/
Published: 20 January 2023
(This article belongs to the Special Issue Staphylococcus aureus Skin and Soft Tissue Infections Pathogenesis and Therapeutic Approaches)
Abstract
:Atopic Dermatitis (AD) skin is susceptible to Staphylococcus aureus (SA) infection, potentially exposing it to a plethora of toxins and virulent determinants, including Panton-Valentine leukocidin (PVL) (α-hemolysin (Hla) and phenol-soluble modulins (PSMs)), and superantigens. Depending on the degree of infection (superficial or invasive), clinical treatments may encompass permanganate (aq) and bleach solutions coupled with intravenous/oral antibiotics such as amoxicillin, vancomycin, doxycycline, clindamycin, daptomycin, telavancin, linezolid, or tigecycline. However, when the skin is significantly traumatized (sheathing of epidermal sections), an SA infection can rapidly ensue, impairing the immune system, and inducing local and systemic AD presentations in susceptible areas. Furthermore, when AD presents systemically, desensitization can be long (years) and intertwined with periods of relapse. In such circumstances, the identification of triggers (stress or infection) and severity of the flare need careful monitoring (preferably in real-time) so that tailored treatments targeting the underlying pathological mechanisms (SA toxins, elevated immunoglobulins, impaired healing) can be modified, permitting rapid resolution of symptoms.
1. Introduction
AD accounts for up 3–10% and 20% of skin diseases in adults and children. Although the link with processed foods remains unresolved [1], high-income countries are the most afflicted, with Swedish infants exhibiting a prevalence of up to 20% [2]. Depending on its severity and frequency, AD can dictate many aspects of the lives of sufferers and may be considered a disability akin to severe depression, cystic fibrosis, or renal disease. Exposure to a variety of everyday stressors (microbial infection (fungal and bacteria toxins), environmental pollutants (dioxins), heat, foods (acrylamides) [3,4], pressure, skin pH, and shock) can trigger AD [5]. Initial symptoms include pruritus and eczematous lesions in the knees, trunk, elbows, neck (flexural folds), and upper and lower extremities. AD can present as a chronic disease interspersed with lengthy periods of symptomatic resolution [6].
AD severity is evaluated using “The Scoring of Atopic Dermatitis (SCORAD)” and is based on the intensity of clinical signs and disease extent, with values > 50 considered moderate to severe. Comparable alternatives to SCORAD are the 5-point Investigator Global Assessment (IGA) scale and “The Eczema Area and Severity Index” (EASI), the former a modification of the Psoriasis and Severity Index (PASI) developed by Hanifin et al. [7]. All scoring systems are considered valid, with none of the methods demonstrating an advantage over the other [8]. Scoring and evaluation between patient and clinician can be done remotely [9] in real time [10] or using a variety of online or smartphone applications such as EZTrack for Atopic Dermatitis [11]. A recent validation study comparing the reliability of smartphone photographs taken and analyzed by 79 AD patients after software training (Imagine, LEO Innovation Lab, Copenhagen, Denmark) with five dermatologists showed high agreement and reliability in the assessment of mild to moderate AD [12]. Furthermore, automated severity scoring performed by deep neural networks exhibited > 90% accuracy providing further reassurance for dermatologists and patients alike [13].
The major medical co-morbidities associated with AD are microbial infections, specifically SA [14,15,16,17,18], affecting > 90% of patients. SA colonization is related to skin barrier disruption, a predominant Th2 immune response, elevated IgE levels specific to staphylococcal enterotoxin, and a reduction in skin microbial diversity. Other infections include viruses such as herpes simplex virus (HSV-1 or HSV-2) and coxsackie A6, which can result in eczema herpeticum (EH) and eczema coxsackium (EC). A summary of the major microbial pathogens associated with AD and their impact on the host immune response and skin microbiome is shown in Table 1. This review will briefly visit the various risk factors associated with SA infection, then, as a backdrop, utilize a patient case study to highlight the impact of associated toxins and their ability to impair healing and immune function, predisposing the subject to various triggers and AD flaring, further affirming previous correlations (infection severity) [6]. Finally, the review will close by looking at potential therapies that utilize the brain–skin–gut axis and their roles in restoring the skin microbiome and rebalancing the immune response regarding mild and severe AD.
2. Genetic Risk Factors and Epigenetic Regulators
A family history with inherited traits is the strongest risk factor for AD, with gene mapping studies accounting for 20% of the estimated heritability [19]. The term ‘heritability’ refers to the proportion of variation within a clinical feature that is attributable to genetic factors. Single null mutations in the gene encoding for the essential epidermal structural protein filaggrin (FLG) are currently the strongest genetic risk factor for AD, afflicting 10% of individuals of European ancestry, although 50% of those do not develop the disease [20]. In addition to the European mutations (R501X, 2282del4, R2447X, S3247X), copy number variations (intragenic repetitive FLG gene sequences) and the amount of filaggrin monomer expressed are additional risk factors for AD. Other candidate genes such as filaggrin-2 (FLG2) and those involved in the Th2 immune response (chromosomes 16p12.1–p11.2 (IL-4 receptor) and 5q31.1 (IL-13 receptor)) are thought to pose a significant risk as well [20]. FLG and FLG2, along with loricrin (LOR) and involucrin (IVL), are located on chromosome 1q21.3 [21,22,23,24]. Together with the S100A7 gene series, they constitute the bulk of the epidermal differentiation complex (EDC) [25]. Expression of the complex is internally modulated by cytokines (IL-4,13,17A,22) derived from T-helper cells (Th1,2,17,22). On the other hand, external modulation can occur via microorganisms (Malassezia spp. and Staphylococcus epidermidis) and numerous phytochemicals via the activation of the aryl hydrocarbon receptor (AHR). Prolonged AHR activation can result in accelerated keratinocytic differentiation, loss of sebocytes, and keratinous cyst formation [26]. Another modulator of keratinocyte differentiation is intracellular reactive oxygen species (ROS). ROS levels are kept in check by the oxidative stress-prone AHR and the antioxidative nuclear factor E2-related factor 2 (NRF2) systems.
Keratinocytic differentiation is key to stratum corneum formation and epidermal [27,28] stratification. Their differentiation and subsequent development are regulated by the EDC genetic complex, signaling pathways, transcription factors, and epigenetic modifications. Epigenetic modifications can vary with exposure to acute and chronic environmental stressors leading to changes in genetic expression throughout all stages of keratinocytic development. These modifications include changes to the methylation status of DNA (methylation and hydroxymethylation), covalent additions or reductions to histone components (methyl-or acetyl groups), expression of noncoding RNAs (ncRNAs), nucleosome positioning, and ATP-dependent chromatin remodeling. Such changes are conducted via DNA methyltransferase (DNMT)1; histone deacetylases (HDAC)1/2; polycomb proteins (PcGs) Bmi1 and Ezh1/2; ATP remodelers Brg1 and Mi-2β of the chromodomain helicase DNA-binding (CHD) family; and histone demethylases (HDACs) Jmjd3 and Setd8, which together with several other transcription factors (in particular, p63) stimulate the proliferation and maturation of progenitor cells.
Of the many proteins, enzymes, and micro ribonucleic acids (miRNAs) involved in modulating genetic expression, the de/methylases and miRNA are the most prevalent. In a recently published [29] epigenome-wide association adult study, significant differences in methylation at 19 CpG sites and a partial correlation in altered gene transcript levels between the epidermis of healthy controls and epidermal lesions in AD patients were reported. Additional epigenetic studies have shown that high exposure to smoke can lead to hypomethylation of the thymic stromal lymphopoietin (TSLP) 5′CpG island in pregnant women and is positively correlated with AD [30]. Furthermore, tobacco smoke is thought to modify the Forkhead box-p3 (FOXP3) locus in umbilical cord blood and is associated with low Treg counts at birth. In addition to FOXP3, low Treg counts correlate with increased miR-223 expression. Other mRNAs that play a critical role in AD include (i) miR-124, which is downregulated in the lesional skin of patients with atopic eczema [31]; (ii) miR-143, a tumor suppressor that targets the IL-13Rα1 receptor reducing IL13 levels and inflammation in epidermal keratinocytes [32]; (iii) miR-146, reported to inhibit the expression of several proinflammatory factors, including IFN-γ and AD-associated genes CCL5, CCL8, and ubiquitin D (UBD) [33]; (iv) miR-155, which is positively correlated with AD severity and essential for the differentiation of Treg and T helper 7 (Th17) cells, and was recently shown to target protein kinase inhibitor α (PKIα) leading to its downregulation along with TSLP in a murine model [34]. TSLP promoter hypomethylation has been observed in keratinocytes isolated from AD patients resulting in the overexpression of TSLP in skin lesions [35]
The various epigenetic factors contributing to the polarized expression of EDC genes, interleukins, and cytokines can result in numerous functional deficiencies (decreased water retention, altered lipid formation, pH imbalance, ceramides, and sphingosine) involving proteins filaggrin, loricrin (LOR), involucrin (IVL), and desmosomes K1, K10, (envoplakin and periplakin) impairing corneocyte integrity [36,37]. Treatments have sought to address these deficiencies by either targeting inflammatory cytokines (monoclonal antibodies) or utilizing antioxidative AHR ligands and nuclear factor E2-related factor 2 (NRF2) activators (tapinarof, coal and soybean tar, and glyteer) to dampen reactive oxygen species (ROS) [38].
Finally, the epigenetic role of Staphylococcal enterotoxin B (SEB) is worth mentioning. In addition to the pore-forming toxins of Helicobacter pylori, Listeria, and Salmonella that repress a subset of immune genes (CXCL2, MKP2, and IFIT3) [39], SEB is adept at altering the methylation pattern of two gene regions (IKBKB and STAT-5B) both of which play a crucial role in T- cell maturation/activation [40].
2.1. Factors That Influence Staphylococcus Aureus Colonization of AD Skin
The reduced colonization resistance of AD skin, coupled with barrier deficiencies and a polarized Th2 immune response, renders the skin susceptible to SA biofilm formation and infiltration. The first stage in colonization is the covalent attachment of SA (sortase rich) to the skin, involving the recruitment of fibronectin, corneocytes, loricrin, and cytokeratin 10 [41]. Other factors that enhance colonization [42] and the potential for severe AD include IL-13 and IL-4, which down-regulate the expression of antimicrobial peptides (AMPs) such as human beta-defensin (HBD)-3 and cathelicidin (LL-37) [43,44]. Conversely, AMP expression is upregulated in the presence of coagulase-negative staphylococci (CoNS), S. hominis and S. epidermidis, which secrete phenol-soluble modulins (PSMs) γ and δ further amplifying AMP potency, thereby restricting in vitro growth of group A Streptococcus and S. aureus [45]. In addition to AMPs, other factors that aid colonization resistance include numerous acids (urocanic and pyrrolidone carboxylic) and lipids (including free fatty acid, ceramides, and sphingosine), filaggrin, and filaggrin degradation products (FDP) [46,47].
2.2. S. aureus Infection, Impairment of the Epidermal Barrier, and Potential Remodeling
When challenged, SA can secrete a battery of soluble receptor-mediated (α-toxin) and non-receptor-mediated (PSMs, δ-hemolysin) toxins, as well as enterotoxins A(SEA), SEB, and C(SEC), and a 22-kD superantigen, synonymous with toxic shock syndrome (TSS) [46,47,48,49]. In addition, the potency of said toxins is enhanced when delivered via extracellular vesicles (EV), which have been shown to play multiple roles in the development of AD and the activation of inflammatory macrophages. For example, mice subjected to repeated intranasal EV exposure induced a Th1 and Th17 cell response via Toll-like receptor 2 (TL2), resulting in neutrophilic pulmonary inflammation [50,51,52,53,54]. Many stressors (biochemical and physical) can induce SA and methicillin-resistant (MRSA) EV production, including iron depletion, sublethal concentrations of vancomycin or ampicillin, and temperature. The content of bacterial EVs has been shown to vary within a given temperature range (34–40 °C), with lower temperature SA EVs more cytotoxic to THP-1 cells (macrophages) than those secreted at 40 °C. Conversely, those secreted at 40 °C have been shown to contain virulence factors, primarily proteins and lipids, exhibiting amplified toxin-mediated erythrocyte lysis compared to their lower temperature counterparts [55].
Apart from toxins, other virulent factors include protein A coagulase, proteases, aureolysin, Staphylokinase, and collagen adhesins, which are collectively involved in repurposing the hosts’ proteins and the seeding of biofilms. In addition to SA, EVs are produced by other colonizing microbes (Malassezia species) and by the hosts’ repair mechanisms (keratinocytes), suggesting that EVs can modulate infection and AD inflammatory processes.
Impairment of the epidermal barrier occurs via various mechanisms involving numerous pore-forming toxins (Leukocidins ED, SF-PVL, AB, MF, HlgCB, HlgAB, and α-hemolysin). The most notable of these is α-hemolysin [56], which forms a heptameric β-barrel pore in the membranes of keratinocytes, initiating an ion imbalance. Cellular distribution is further enhanced with the binding of Protein A(PA) to tumor necrosis factor (TNF) receptors on the surface of keratinocytes triggering a sustained Th2 polarized inflammatory response via Langerhans cells. In the ensuing milieu, several other SA endogenous (kallikrein (KLK) 6, KLK13, and KLK14) proteases with a penchant for filaggrin and lipoproteins for the host’s fatty acids (linoleic acid (C18:2) [57] further amplify the Th2 inflammatory response by inducing thymic stromal lymphopoietin expression in a TLR2/TLR6-dependent manner in primary human keratinocytes [58]. Finally, SA becomes internalized by the keratinocytes inducing IL-1α production via Toll-like receptor 9 (TLR9). The various factors involved in SA adhesion and its subsequent internalization by keratinocytes are summarized in Figure 1 [59].
Other virulent factors include δ-Toxin and PSMα2 and PSMα3, the latter acting synergistically with IgE in the absence of bound antigen [60]. Last but not least is the role of superantigens (SAgs) and their impact on the corneocytes of the stratum corneum resulting in IgE antibody elevations following antigen presentation to Th2 cells [61]. SAgs can also trigger mast cell degranulation and induce Th2 cells to release IL-31. Children colonized with bacteria expressing SAgs have a higher disease severity than those with strains [62].
3. The Initial Impact of SA Toxins and AD Recurrence
The impact of these toxins on a child’s skin is shown in Figure 2 [63]. At the time of record, the subject also exhibited (not shown) first systemic AD presentations in multiple cutaneous areas (neck, trunk, arms, and ankles). All AD presentations had dissipated within 12 h of administering antibiotics, and skin restoration was observed after 4 weeks.
Without testing, it is debatable whether patient X had subclinical Th2 and Th22 cellular elevations in those areas’ prior infection. However, it seems likely that multiple pools of antigen-primed T and effector memory cells were generated during infection, which could be triggered later [62].
As discussed previously, only 5% with a genetic predisposition (European ancestry) for AD develop chronic disease. Unfortunately, patient X soon embarked on a long course of mild AD interspersed with periods of acute exacerbations (flares). The first of these resulted from a minor knee skin abrasion (no sign of infection), triggering a hypersensitive response in the folds of both arms coupled with intense itching < 12 h. Initial presentation appeared mixed (bacterial and fungi), with the administration of generic antibiotic creams (week 2) only serving to exacerbate. During the next two weeks, contact allergens were minimized by routinely (every morning) disinfecting bed linen (spraying) and bathing skin folds prior to sleep. The course of events is depicted in Figure 3, with asymptomatic resolution achieved in week 4.
A similar case study involving a nine-year-old boy with chronic AD was reported by Kanchongkittiphon et al. [64], in which the total serum IgE level was markedly elevated, and the possibility of allergen (mites) immunotherapy was discussed. Regarding patient X, bleach baths were considered, but a targeted bathing approach concentrating on the folds, in combination with chloroxylenol spraying of bed linens (patient absent), was adopted, with time to resolution akin to generic treatments.
Further exacerbations were recorded at the end of 2017 (travel stress), followed by severe flaring after another fall in early 2018, suggesting post-traumatic stress involvement. The impact of psychological stress on AD severity, frequency of exacerbations, and reduced healing potential is well documented [65]. In such circumstances (Figure 3), the administration of selective serotonin reuptake inhibitors (SSRIs) sertraline and fluoxetine may have been considered, given the drug’s reported ability to reduce levels of anxiety, proinflammatory cytokines and IgE in AD patients [66,67,68,69,70]. In addition, SSRI paroxetine and fluvoxamine administered to AD patients were shown to reduce pruritus, suggesting central and peripheral nervous system involvement and the neurotransmitter 5-hydroxytryptamine (5-HT). However, anxiety is a complex disorder, and side effects can present if the underlying phenotype has not been identified [67,71]; consequently, recent investigations are currently restricted to topical SSRI usage, such as fluoxetine, which was shown to improve wound healing in mice by accelerating keratinocyte migration and rebalancing the local immune milieu [71]. The interplay between immune cells, neurotransmitters, and hormones and the effect on the levels of cortisol, adrenocorticotropic, and corticotropin-releasing hormones released by mature and immature keratinocytes and fibroblasts remains under investigation [72]. As a side note, for the first 24 months post-trauma (in the absence of an AD flare), the healing time for an abrasion remained significantly prolonged (10–14 days), suggesting IL-4 dysregulation. A model that probably best emulates that period might be the Interleukin-4 Transgenic Mouse developed by Zhao et al. [73] regarding aberrant wound healing.
Lengthy and frequent AD recurrences can lead to observable thickening in the flexural folds with papule-nodule transitions, plaques with excoriation, lichenification, and significant remodeling (Figure 4A–C). Changes in epidermal thickness resulting from chronic AD are usually accompanied by a higher density of nerve fibers [74], Th2, and epidermal dendritic cells, increasing regional susceptibility to additional environmental factors such as heat, pressure, and sweat, the latter often associated with soft wound infections. Soft wound treatments range from cold atmospheric pressure plasma [75], silver and zinc nanoparticles, platelet therapies, chitin bandages, and tens of others. The scarring and nodulation resulting from AD recurrence are shown in Figure 4C.
During 2018–2021, patient X relapsed multiple times, putting them at significant risk from the so-called atopic march that can lead to asthma and allergic rhinitis in later life. However, nodulations rooted in the dermis and epidermis did show significant shrinkage following topical povidone administration in early 2022. Moreover, additional wounds or abrasions rapidly treated with povidone (PV) later in the same year did not evoke flaring and healed quicker, which may have been due to limited IgE production leading to reductions in reagin antibody titers or how the treatment (psychological) was administered [76]. Interestingly, a recent pilot phase II study investigating the combinatorial antiseptic and anti-inflammatory action of liposomal polyvinyl-pyrrolidone (PVP)-iodine gel reduced the global clinical severity score (GCSS) for all dermatoses [77,78]. Moreover, PV preventions have been shown to inhibit C. albicans, and SA biofilm formations often encountered with chronic AD [79]. Side effects included itching and burning (9 and 14%). Povidone has been shown to counter various SA and MRSA armaments (α-hemolysin, phospholipase C, lipase, elastase, and β-glucuronidase) [80] and remains the gold standard for biocides in hospital settings [81,82]. A timeline documenting the progression of AD and associated triggers regarding patient X can be found in Table 2.
However, the misuse of biocides can lead to a surge in MRSA cases (cross-resistance), as reported during the COVID-19 pandemic [83]; thus, alternatives treatments need to be developed to limit the emergence of multi-drug resistant microbes (fungi, bacteria, and viruses) [84]. A recent review by Bieber et al. documents a number of approved inhibitor therapies (IL-4Rα “dupilumab”, anti-IL-13 “tralokinumab”, and JAK1/2) and those currently in clinical trials [85], which might have been a better option (dupilumab) for patient X.
4. Restoration of Microbial Homeostasis
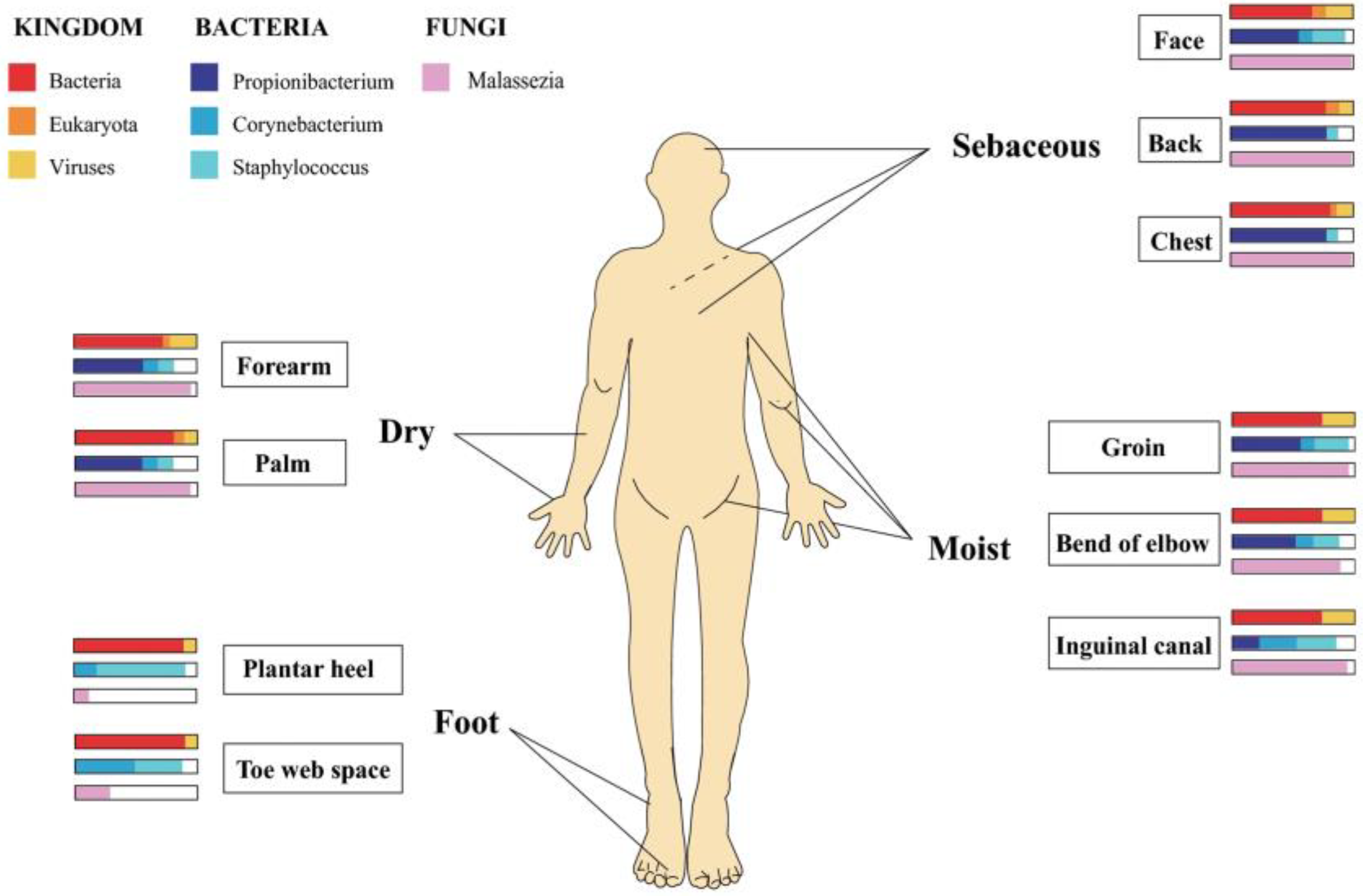
The skin microbiome consists of regionally specific microbial communities hosting over 1000 species from 19 phyla. These communities’ microbial composition depends on on-site skin physiology, and the microenvironments (dry, moist, foot, sebaceous) encountered there (Figure 5). For example, where water loss is relatively high (dry) and follicle density (inner forearm) low, fungal species (Malassezia) predominate; in moist regions (folds), bacteria species (Staphylococcus and Corynebacterium) hold sway and where follicle density and skin surface area are highest (Sebaceous) (head, neck, and chest) lipophilic Propionibacterium species dominate. Finally, various fungal (Aspergillus, Cryptococcus, Rhodotorula, Malassezia, and Epicoccum) species can be found in the feet [86].
Collectively, these microbially diverse communities constitute the bulk of the skin microbiome. The density and diversity of these communities vary with age, sex, and race of the host. Interactions (communications) with the host occur via epithelial and immune cells and are mutually beneficial with the community’s providing protection from pathogenic or translocated species and the host offering “food and board.”
However, these interactions can be perturbed by environmental conditions (skin infection or trauma), providing opportunities for other species, such as SA, that are adept at prioritizing overgrowth and biofilm formations [86,87], leading to prolonged barrier and immunological dysfunction. Of the many SA strains residing on the skin, the clonal complex 1 (CC1) takes precedence during severe AD, whereas CC30 is prominent on the skin of asymptomatic AD individuals.
Numerous therapeutic approaches, including endolysins, bacteriocins (Nisin Z, Lacticin 3147), small molecule inhibitors (2-aminoimidazole), bacteriophages, and nanoparticles or combinations thereof, can be used to prevent SA biofilm formation [80,88]. However, in the case of patient X, concerns extended beyond the skin, as the longevity and frequency of the disease suggested the initial involvement of the gut (microbiome) and, later on, the brain (stress) via the gut–brain–skin axis [89]. Animal model (murine) research regarding factors that affect the axis has revealed the immunoregulatory importance of dietary AHR ligands and hydrophilic bile acids. For example, mice fed an indole-3-Carbinol(I3C)-supplemented diet showed better monocyte-to-dendritic skin differentiation than controls [90], which is consistent with previous reports highlighting the shift from macrophages via the modulation of transcription factors Irf4 and Blimp-1. Moreover, I3C diet supplementation reversed antibiotic-treated mice with impaired differentiation, suggesting a role for microbial-derived AHR ligands [90]. The effects of dietary tryptophan-free deficiencies on an experimental autoimmune encephalomyelitis (EAE) model resulted in higher disease scores, increased expression of pro-neuroinflammatory molecules (Ccl2, Nos2, and Tnfa), and prolonged [91] recovery in mice.
The catabolism of dietary tryptophan represents a major source of indoles for humans. A recent clinical study comparing elevated fasting serum and normal IgE levels in infant AD cohorts showed the former had increased serum indole-3-acetic acid (tryptophan metabolite) and tryptophan levels [92], suggesting impaired tryptophan metabolism. Moreover, the cohort with normal IgE levels also showed a significant decrease in primary conjugated bile acids (taurocholic, taurochenodeoxycholic acid, and glycochenodeoxycholic). In contrast, the high IgE AD cohort showed cholic acid and chenodeoxycholic acid elevations, suggesting reduced conjugation with a potential risk of E. coli intestinal overgrowth [93].
Of the many indole derivatives currently under investigation [94,95,96,97], the work by Yu et al. demonstrated the potential of indole-3-aldehyde treatments for lesional and non-lesional AD patients by attenuating skin inflammation and TSLP keratinocyte expression in an AD-like dermatitis murine model (MC903) [98]. More recently, Fang et al. further emphasized the importance of microbially (Limosilactobacillus reuteri) produced indole derivatives (indole lactic acid and indole propionic acid) in attenuating AD by demonstrating its ability to suppress IgE, TSLP, IL-4 and 5 levels in mice [99]. Regarding the anti-inflammatory potential of hydrophilic bile acids (tauroursodeoxycholic acid (TUDCA)), reports have shown that they reduce the unfolded protein response (UPR) seen in keratinocytes [59,100], hypersensitivity in asthmatic patients, as well as reverse the amyloid beta-oxidation of mitochondrial membranes in Alzheimer’s murine models [101,102]. Whether TUDCA alone or blended bile acids treatments can re-establish skin microbial homeostasis directly or indirectly via changes in the gut microbiota (probiotic interventions or fecal matter transfer) requires further investigation. In addition to indole derivatives, a study by Hwang et al. investigated whether n-3 polyunsaturated fatty acids (PUFA) in red blood cells (RBC) had a protective effect on the atopy of 380 preschoolers. Preschoolers with atopy had lower total RBC n-3 PUFA levels and a greater n-6/n-3 PUFA ratio than the controls, suggesting lower n-3 PUFA may play a role in child atropy [103].
Microbial analysis of AD fecal matter by Reddel [104] and co-workers demonstrated that microbiomes of 19 AD children were characterized by an increase of Faecalibacterium, Sutterella, Oscillospira, Bacteroides, and Parabacteroides and a significant reduction in short-chain fatty acid (SCFA)-producing bacteria (Bifidobacterium, Blautia, Coprococcus, Eubacterium, and Propionibacterium) compared with 18 healthy individuals. More recently [105], Kim et al. showed that fecal matter transfer (FMT) from a healthy murine donor ameliorated AD in mice; resulted in reduced IgE levels and the numbers of mast cells, eosinophils, and basophils; and the restoration of the Th1/Th2 balance. An initial clinical study (n = 9) [106] assessing the clinical efficacy of capsule FMT as a viable treatment in adults with moderate-to-severe AD over eight months demonstrated a significant reduction in the average SCORAD score from baseline pre-FMT as well as a decrease in corticosteroid usage, disease severity, and partial assimilation of the donor’s microbiota.
5. Conclusions
AD is a complex, multifaceted disease with varying degrees of severity (mild, moderate, and severe) and recurrence. Some reports have speculated that a comprehensive understanding of the skin’s ability via microbial modulation to resist SA colonization would reduce AD severity and recurrence, leading to better therapeutics. However, personalized modulation is still in its infancy as the skin virome is yet to be typed, and the ability of species to limit or enhance SA virulence is yet to be elucidated [107,108]. Regarding patient X, a unique set of conditions was presented that allowed for AD monitoring and the changing factors (trauma-stress-temperature and pressure) that initiated flaring over the course of the disease. After numerous therapeutic failures and temporary successes spanning five years, the link between initiation and flaring was finally broken with the topical administration of the biocide povidone. However, the use of biocides has a detrimental effect on commensal microbiota. Thus, alternative treatments that rebalance or partially restore the skin microbiome via bacterial species, such as Bifidobacterium longum or bile acids (TUDCA), that better mediate tryptophan metabolism via the gut–brain axis can be employed regarding mild or non-lesional AD. Concerning chronic or severe AD, in which the dysregulation of the gut immune system is suspected, FMT or partial FMT may be adopted to restore the Th1/Th2 balance and reduce serum IgE levels preventing atopic march [105].
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Thanks to Gachon University, South Korea.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AHR | Aryl hydrocarbon receptor |
| Blimp-1 | B lymphocyte-induced maturation protein-1 |
| Ccl2 | Chemokine ligand 2 |
| CoNS | Coagulase-negative staphylococci |
| E. coli | Escherichia coli |
| FLG | Filaggrin |
| FMT | Fecal matter transfer |
| Gamma | δ-Toxin |
| Hla | α−hemolysin |
| HlgAB | γ-hemolysin |
| HlgCB | γ-hemolysin |
| Jmjd3 | Jumonji domain-containing protein-3 |
| PSM | Phenol-soluble modulin |
| PVL | Panton-Valentine leukocidin |
| PVP | Polyvinyl-pyrrolidone |
| LOR | Loricrin |
| I3C | Indole-3-Carbinol |
| Irf4 | Interferon Regulatory Factor 4 |
| IVL | Involucrin |
| EDC | Epidermal differentiation complex |
| KLK | Kallikrein |
| MRSA | Methicillin-resistant Staphylococcus aureus |
| NRF2 | Nuclear factor E2-related factor 2 |
| Nos2 | Nitric oxide synthase 2 |
| SA | Staphylococcus aureus |
| SCFA | Short-chain fatty acids |
| SEA | Staphylococcal enterotoxins-A |
| SEB | Staphylococcal enterotoxins-B |
| SEC | Staphylococcal enterotoxins-C |
| SSRIs | Selective serotonin reuptake inhibitors |
| TL2 | Toll-like receptor 2 |
| Tnfa | Tumor necrosis factor alpha |
References
- Li, Y.; Su, J.; Luo, D.; Duan, Y.; Huang, Z.; He, M.; Tao, J.; Xiao, S.; Xiao, Y.; Chen, X.; et al. Processed Food and Atopic Dermatitis: A Pooled Analysis of Three Cross-Sectional Studies in Chinese Adults. Front. Nutr. 2021, 8, 754663. [Google Scholar] [CrossRef] [PubMed]
- Urban, K.; Chu, S.; Giesey, R.L.; Mehrmal, S.; Uppal, P.; Nedley, N.; Delost, G.R. The Global, Regional, and National Burden of Atopic Dermatitis in 195 Countries and Territories: An Ecological Study from the Global Burden of Disease Study 2017. JAAD Int. 2021, 2, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Hajimohammadi, B.; Athari, S.M.; Abdollahi, M.; Vahedi, G.; Athari, S.S. Oral Administration of Acrylamide Worsens the Inflammatory Responses in the Airways of Asthmatic Mice Through Agitation of Oxidative Stress in the Lungs. Front. Immunol. 2020, 11, 1940. [Google Scholar] [CrossRef] [PubMed]
- Koszucka, A.; Nowak, A.; Nowak, I.; Motyl, I. Acrylamide in Human Diet, Its Metabolism, Toxicity, Inactivation and the Associated European Union Legal Regulations in Food Industry. Crit. Rev. Food Sci. Nutr. 2019, 60, 1677–1692. [Google Scholar] [CrossRef] [PubMed]
- Thammahong, A.; Kiatsurayanon, C.; Edwards, S.W.; Rerknimitr, P.; Chiewchengchol, D. The Clinical Significance of Fungi in Atopic Dermatitis. Int. J. Derm. 2020, 59, 926–935. [Google Scholar] [CrossRef]
- Girolomoni, G.; Busà, V.M. Flare Management in Atopic Dermatitis: From Definition to Treatment. Ther. Adv. Chronic Dis. 2022, 13, 20406223211066728. [Google Scholar] [CrossRef]
- Hanifin, J.M.; Thurston, M.; Omoto, M.; Cherill, R.; Tofte, S.J.; Graeber, M. The Eczema Area and Severity Index (EASI): Assessment of Reliability in Atopic Dermatitis. EASI Evaluator Group. Exp. Derm. 2001, 10, 11–18. [Google Scholar] [CrossRef]
- Bożek, A.; Reich, A. Assessment of Intra- and Inter-Rater Reliability of Three Methods for Measuring Atopic Dermatitis Severity: EASI, Objective SCORAD, and IGA. DRM 2017, 233, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Lull, C.; von Ahnen, J.A.; Gross, G.; Olsavszky, V.; Knitza, J.; Leipe, J.; Schmieder, A. German Mobile Apps for Patients With Psoriasis: Systematic Search and Evaluation. JMIR Mhealth Uhealth 2022, 10, e34017. [Google Scholar] [CrossRef]
- van Galen, L.S.; Xu, X.; Koh, M.J.A.; Thng, S.; Car, J. Eczema Apps Conformance with Clinical Guidelines: A Systematic Assessment of Functions, Tools and Content. Br. J. Dermatol. 2020, 182, 444–453. [Google Scholar] [CrossRef]
- Xu, X.; Griva, K.; Koh, M.; Lum, E.; Tan, W.S.; Thng, S.; Car, J. Creating a Smartphone App for Caregivers of Children With Atopic Dermatitis With Caregivers, Health Care Professionals, and Digital Health Experts: Participatory Co-Design. JMIR Mhealth Uhealth 2020, 8, e16898. [Google Scholar] [CrossRef]
- Ali, Z.; Chiriac, A.; Bjerre-Christensen, T.; Isberg, A.P.; Dahiya, P.; Manole, I.; Dutei, A.-M.; Deaconescu, I.; Serban, A.; Suru, A.; et al. Mild to Moderate Atopic Dermatitis Severity Can Be Reliably Assessed Using Smartphone-Photographs Taken by the Patient at Home: A Validation Study. Ski. Res. Technol. 2022, 28, 336–341. [Google Scholar] [CrossRef]
- Bang, C.H.; Yoon, J.W.; Ryu, J.Y.; Chun, J.H.; Han, J.H.; Lee, Y.B.; Lee, J.Y.; Park, Y.M.; Lee, S.J.; Lee, J.H. Automated Severity Scoring of Atopic Dermatitis Patients by a Deep Neural Network. Sci. Rep. 2021, 11, 6049. [Google Scholar] [CrossRef]
- Lyons, J.J.; Milner, J.D.; Stone, K.D. Atopic Dermatitis in Children: Clinical Features, Pathophysiology, and Treatment. Immunol. Allergy Clin. 2015, 35, 161–183. [Google Scholar] [CrossRef] [Green Version]
- Shi, B.; Leung, D.Y.M.; Taylor, P.A.; Li, H. Methicillin-Resistant Staphylococcus Aureus Colonization Is Associated with Decreased Skin Commensal Bacteria in Atopic Dermatitis. J. Investig. Derm. 2018, 138, 1668–1671. [Google Scholar] [CrossRef] [Green Version]
- Sugarman, J.L.; Hersh, A.L.; Okamura, T.; Howard, R.; Frieden, I.J. A Retrospective Review of Streptococcal Infections in Pediatric Atopic Dermatitis. Pediatr. Derm. 2011, 28, 230–234. [Google Scholar] [CrossRef]
- Möckel, M.; De La Cruz, N.C.; Rübsam, M.; Wirtz, L.; Tantcheva-Poor, I.; Malter, W.; Zinser, M.; Bieber, T.; Knebel-Mörsdorf, D. Herpes Simplex Virus 1 Can Bypass Impaired Epidermal Barriers upon Ex Vivo Infection of Skin from Atopic Dermatitis Patients. J. Virol. 2022, 96, e0086422. [Google Scholar] [CrossRef]
- Szczepańska, M.; Blicharz, L.; Nowaczyk, J.; Makowska, K.; Goldust, M.; Waśkiel-Burnat, A.; Czuwara, J.; Samochocki, Z.; Rudnicka, L. The Role of the Cutaneous Mycobiome in Atopic Dermatitis. J. Fungi 2022, 8, 1153. [Google Scholar] [CrossRef]
- Weidinger, S.; Novak, N. Atopic Dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Pellerin, L.; Henry, J.; Hsu, C.Y.; Balica, S.; Jean-Decoster, C.; Méchin, M.C.; Hansmann, B.; Rodriguez, E.; Weindinger, S.; Schmitt, A.M.; et al. Defects of Filaggrin-like Proteins in Both Lesional and Nonlesional Atopic Skin. J. Allergy Clin. Immunol. 2013, 131, 1094–1102. [Google Scholar] [CrossRef]
- Elmose, C.; Thomsen, S.F. Twin Studies of Atopic Dermatitis: Interpretations and Applications in the Filaggrin Era. J. Allergy 2015, 2015, 902359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furue, M. Regulation of Filaggrin, Loricrin, and Involucrin by IL-4, IL-13, IL-17A, IL-22, AHR, and NRF2: Pathogenic Implications in Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 5382. [Google Scholar] [CrossRef] [PubMed]
- Scharschmidt, T.C.; Man, M.Q.; Hatano, Y.; Crumrine, D.; Gunathilake, R.; Sundberg, J.P.; Silva, K.A.; Mauro, T.M.; Hupe, M.; Cho, S.; et al. Filaggrin Deficiency Confers a Paracellular Barrier Abnormality That Reduces Inflammatory Thresholds to Irritants and Haptens. J. Allergy Clin. Immunol. 2009, 124, 496–506. [Google Scholar] [CrossRef] [Green Version]
- Jungersted, J.M.; Scheer, H.; Mempel, M.; Baurecht, H.; Cifuentes, L.; Høgh, J.K.; Hellgren, L.I.; Jemec, G.B.E.; Agner, T.; Weidinger, S. Stratum Corneum Lipids, Skin Barrier Function and Filaggrin Mutations in Patients with Atopic Eczema. Allergy 2010, 65, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.J.; Elias, M.S.; Bradley, M. Genetics in Atopic Dermatitis: Historical Perspective and Future Prospects. Acta Derm. Venereol. 2020, 100, adv00163. [Google Scholar] [CrossRef]
- Ju, Q.; Fimmel, S.; Hinz, N.; Stahlmann, R.; Xia, L.; Zouboulis, C.C. 2,3,7,8-Tetrachlorodibenzo-p-Dioxin Alters Sebaceous Gland Cell Differentiation in Vitro. Exp. Derm. 2011, 20, 320–325. [Google Scholar] [CrossRef]
- Rodríguez, E.; Baurecht, H.; Wahn, A.F.; Kretschmer, A.; Hotze, M.; Zeilinger, S.; Klopp, N.; Illig, T.; Schramm, K.; Prokisch, H.; et al. An Integrated Epigenetic and Transcriptomic Analysis Reveals Distinct Tissue-Specific Patterns of DNA Methylation Associated with Atopic Dermatitis. J. Investig. Derm. 2014, 134, 1873–1883. [Google Scholar] [CrossRef] [Green Version]
- Moltrasio, C.; Romagnuolo, M.; Marzano, A.V. Epigenetic Mechanisms of Epidermal Differentiation. Int. J. Mol. Sci. 2022, 23, 4874. [Google Scholar] [CrossRef]
- Mu, Z.; Zhang, J. The Role of Genetics, the Environment, and Epigenetics in Atopic Dermatitis. Adv. Exp. Med. Biol. 2020, 1253, 107–140. [Google Scholar] [CrossRef]
- Martin, M.J.; Estravís, M.; García-Sánchez, A.; Dávila, I.; Isidoro-García, M.; Sanz, C. Genetics and Epigenetics of Atopic Dermatitis: An Updated Systematic Review. Genes 2020, 11, 442. [Google Scholar] [CrossRef]
- Yang, Z.; Zeng, B.; Wang, C.; Wang, H.; Huang, P.; Pan, Y. MicroRNA-124 Alleviates Chronic Skin Inflammation in Atopic Eczema via Suppressing Innate Immune Responses in Keratinocytes. Cell Immunol. 2017, 319, 53–60. [Google Scholar] [CrossRef]
- Zeng, Y.-P.; Nguyen, G.H.; Jin, H.-Z. MicroRNA-143 Inhibits IL-13-Induced Dysregulation of the Epidermal Barrier-Related Proteins in Skin Keratinocytes via Targeting to IL-13Rα1. Mol. Cell. Biochem. 2016, 416, 63–70. [Google Scholar] [CrossRef]
- Rebane, A.; Runnel, T.; Aab, A.; Maslovskaja, J.; Rückert, B.; Zimmermann, M.; Plaas, M.; Kärner, J.; Treis, A.; Pihlap, M.; et al. MicroRNA-146a Alleviates Chronic Skin Inflammation in Atopic Dermatitis through Suppression of Innate Immune Responses in Keratinocytes. J. Allergy Clin. Immunol. 2014, 134, 836–847.e11. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Y.; Yuan, W.; Yao, L.; Wang, S.; Jia, Z.; Wu, P.; Li, L.; Wei, P.; Wang, X.; et al. MicroRNA-155-5p Is a Key Regulator of Allergic Inflammation, Modulating the Epithelial Barrier by Targeting PKIα. Cell Death Dis. 2019, 10, 884. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Zhou, B.; Zhao, M.; Tang, J.; Lu, Q. Promoter Demethylation Contributes to TSLP Overexpression in Skin Lesions of Patients with Atopic Dermatitis. Clin. Exp. Derm. 2014, 39, 48–53. [Google Scholar] [CrossRef]
- Kim, B.E.; Leung, D.Y.M.; Boguniewicz, M.; Howell, M.D. Loricrin and Involucrin Expression Is Down-Regulated by Th2 Cytokines through STAT-6. Clin. Immunol. 2008, 126, 332–337. [Google Scholar] [CrossRef] [Green Version]
- Takei, K.; Mitoma, C.; Hashimoto-Hachiya, A.; Takahara, M.; Tsuji, G.; Nakahara, T.; Furue, M. Galactomyces Fermentation Filtrate Prevents T Helper 2-Mediated Reduction of Filaggrin in an Aryl Hydrocarbon Receptor-Dependent Manner. Clin. Exp. Dermatol. 2015, 40, 786–793. [Google Scholar] [CrossRef]
- Guttman-Yassky, E.; Bissonnette, R.; Ungar, B.; Suárez-Fariñas, M.; Ardeleanu, M.; Esaki, H.; Suprun, M.; Estrada, Y.; Xu, H.; Peng, X.; et al. Dupilumab Progressively Improves Systemic and Cutaneous Abnormalities in Patients with Atopic Dermatitis. J. Allergy Clin. Immunol. 2019, 143, 155–172. [Google Scholar] [CrossRef] [Green Version]
- Bierne, H.; Hamon, M.; Cossart, P. Epigenetics and Bacterial Infections. Cold Spring Harb. Perspect. Med. 2012, 2, a010272. [Google Scholar] [CrossRef]
- Pérez-Novo, C.A.; Zhang, Y.; Denil, S.; Trooskens, G.; De Meyer, T.; Van Criekinge, W.; Van Cauwenberge, P.; Zhang, L.; Bachert, C. Staphylococcal Enterotoxin B Influences the DNA Methylation Pattern in Nasal Polyp Tissue: A Preliminary Study. Allergy Asthma Clin. Immunol. 2013, 9, 48. [Google Scholar] [CrossRef]
- Geoghegan, J.A.; Foster, T.J. Cell Wall-Anchored Surface Proteins of Staphylococcus Aureus: Many Proteins, Multiple Functions. Curr. Top Microbiol. Immunol. 2017, 409, 95–120. [Google Scholar] [CrossRef] [PubMed]
- Tauber, M.; Balica, S.; Hsu, C.-Y.; Jean-Decoster, C.; Lauze, C.; Redoules, D.; Viodé, C.; Schmitt, A.-M.; Serre, G.; Simon, M.; et al. Staphylococcus Aureus Density on Lesional and Nonlesional Skin Is Strongly Associated with Disease Severity in Atopic Dermatitis. J. Allergy Clin. Immunol. 2016, 137, 1272–1274.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirofski, L.; Casadevall, A. The Meaning of Microbial Exposure, Infection, Colonisation, and Disease in Clinical Practice. Lancet Infect. Dis. 2002, 2, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.J.; McLean, W.H.I. One Remarkable Molecule: Filaggrin. J. Investig. Derm. 2012, 132, 751–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.E.; Leung, D.Y.M. Significance of Skin Barrier Dysfunction in Atopic Dermatitis. Allergy Asthma Immunol. Res. 2018, 10, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Benenson, S.; Zimhony, O.; Dahan, D.; Solomon, M.; Raveh, D.; Schlesinger, Y.; Yinnon, A.M. Atopic Dermatitis—A Risk Factor for Invasive Staphylococcus Aureus Infections: Two Cases and Review. Am. J. Med. 2005, 118, 1048–1051. [Google Scholar] [CrossRef]
- Goleva, E.; Berdyshev, E.; Leung, D.Y. Epithelial Barrier Repair and Prevention of Allergy. J. Clin. Investig. 2019, 129, 1463–1474. [Google Scholar] [CrossRef] [Green Version]
- Otto, M. Staphylococcal Infections: Mechanisms of Biofilm Maturation and Detachment as Critical Determinants of Pathogenicity. Annu. Rev. Med. 2013, 64, 175–188. [Google Scholar] [CrossRef]
- Wang, X.; Koffi, P.F.; English, O.F.; Lee, J.C. Staphylococcus Aureus Extracellular Vesicles: A Story of Toxicity and the Stress of 2020. Toxins 2021, 13, 75. [Google Scholar] [CrossRef]
- Hong, S.-W.; Choi, E.-B.; Min, T.-K.; Kim, J.-H.; Kim, M.-H.; Jeon, S.G.; Lee, B.-J.; Gho, Y.S.; Jee, Y.-K.; Pyun, B.-Y.; et al. An Important Role of α-Hemolysin in Extracellular Vesicles on the Development of Atopic Dermatitis Induced by Staphylococcus Aureus. PLoS ONE 2014, 9, e100499. [Google Scholar] [CrossRef]
- Kim, M.-R.; Hong, S.-W.; Choi, E.-B.; Lee, W.-H.; Kim, Y.-S.; Jeon, S.G.; Jang, M.H.; Gho, Y.S.; Kim, Y.-K. Staphylococcus Aureus-Derived Extracellular Vesicles Induce Neutrophilic Pulmonary Inflammation via Both Th1 and Th17 Cell Responses. Allergy 2012, 67, 1271–1281. [Google Scholar] [CrossRef]
- White, J.R.; Dauros-Singorenko, P.; Hong, J.; Vanholsbeeck, F.; Phillips, A.; Swift, S. The Complex, Bidirectional Role of Extracellular Vesicles in Infection. Biochem. Soc. Trans. 2021, 49, 881–891. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, Q.; Tang, J.; Xiong, L.; Li, L. Extracellular Vesicles: Emerging Therapeutics in Cutaneous Lesions. Int. J. Nanomed. 2021, 16, 6183–6202. [Google Scholar] [CrossRef]
- Wang, X.; Eagen, W.J.; Lee, J.C. Orchestration of Human Macrophage NLRP3 Inflammasome Activation by Staphylococcus Aureus Extracellular Vesicles. Proc. Natl. Acad. Sci. USA 2020, 117, 3174–3184. [Google Scholar] [CrossRef]
- Luz, B.S.R.D.; Nicolas, A.; Chabelskaya, S.; Rodovalho, V.D.R.; Le Loir, Y.; Azevedo, V.A.D.C.; Felden, B.; Guédon, E. Environmental Plasticity of the RNA Content of Staphylococcus Aureus Extracellular Vesicles. Front. Microbiol. 2021, 12, 634226. [Google Scholar] [CrossRef]
- Briaud, P.; Frey, A.; Marino, E.C.; Bastock, R.A.; Zielinski, R.E.; Wiemels, R.E.; Keogh, R.A.; Murphy, E.R.; Shaw, L.N.; Carroll, R.K. Temperature Influences the Composition and Cytotoxicity of Extracellular Vesicles in Staphylococcus Aureus. mSphere 2021, 6, e0067621. [Google Scholar] [CrossRef]
- Nguyen, M.T.; Hanzelmann, D.; Härtner, T.; Peschel, A.; Götz, F. Skin-Specific Unsaturated Fatty Acids Boost the Staphylococcus Aureus Innate Immune Response. Infect. Immun. 2016, 84, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Paller, A.S.; Kong, H.H.; Seed, P.; Naik, S.; Scharschmidt, T.C.; Gallo, R.L.; Luger, T.; Irvine, A.D. The Microbiome in Patients with Atopic Dermatitis. J. Allergy Clin. Immunol. 2019, 143, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, K.; Moriwaki, M.; Miyake, R.; Hide, M. Staphylococcus Aureus in Atopic Dermatitis: Strain-Specific Cell Wall Proteins and Skin Immunity. Allergol. Int. 2019, 68, 309–315. [Google Scholar] [CrossRef]
- Williams, M.R.; Nakatsuji, T.; Sanford, J.A.; Vrbanac, A.F.; Gallo, R.L. Staphylococcus Aureus Induces Increased Serine Protease Activity in Keratinocytes. J. Investig. Dermatol. 2017, 137, 377–384. [Google Scholar] [CrossRef]
- Beck, L.A.; Cork, M.J.; Amagai, M.; De Benedetto, A.; Kabashima, K.; Hamilton, J.D.; Rossi, A.B. Type 2 Inflammation Contributes to Skin Barrier Dysfunction in Atopic Dermatitis. JID Innov. 2022, 2, 100131. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.A.; Luster, A.D. T Cell Homing to Epithelial Barriers in Allergic Disease. Nat. Med. 2012, 18, 705–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulme, J. Recent Advances in the Detection of Methicillin Resistant Staphylococcus Aureus (MRSA). BioChip J. 2017, 11, 89–100. [Google Scholar] [CrossRef]
- Kanchongkittiphon, W.; Gaffin, J.M.; Phipatanakul, W. Child with Atopic Dermatitis. Ann. Allergy Asthma Immunol. 2015, 114, 6–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pondeljak, N.; Lugović-Mihić, L. Stress-Induced Interaction of Skin Immune Cells, Hormones, and Neurotransmitters. Clin. Ther. 2020, 42, 757–770. [Google Scholar] [CrossRef]
- Kiecka, A.; Szczepanik, M. The Potential Action of SSRIs in the Treatment of Skin Diseases Including Atopic Dermatitis and Slow-Healing Wounds. Pharm. Rep. 2022, 74, 947–955. [Google Scholar] [CrossRef]
- Oliveira, A.S.; Martinez-de-Oliveira, J.; Donders, G.G.G.; Palmeira-de-Oliveira, R.; Palmeira-de-Oliveira, A. Anti-Candida Activity of Antidepressants Sertraline and Fluoxetine: Effect upon Pre-Formed Biofilms. Med. Microbiol. Immunol. 2018, 207, 195–200. [Google Scholar] [CrossRef]
- Kalaycı, S.; Demirci, S.; Sahin, F. Antimicrobial Properties of Various Psychotropic Drugs Against Broad Range Microorganisms. Curr. Psychopharmacol. 2014, 3, 195–202. [Google Scholar] [CrossRef]
- Ait Chait, Y.; Mottawea, W.; Tompkins, T.A.; Hammami, R. Unravelling the Antimicrobial Action of Antidepressants on Gut Commensal Microbes. Sci. Rep. 2020, 10, 17878. [Google Scholar] [CrossRef]
- Shen, Y.; Yang, X.; Li, G.; Gao, J.; Liang, Y. The Change of Gut Microbiota in MDD Patients under SSRIs Treatment. Sci. Rep. 2021, 11, 14918. [Google Scholar] [CrossRef]
- Nguyen, C.M.; Tartar, D.M.; Bagood, M.D.; So, M.; Nguyen, A.V.; Gallegos, A.; Fregoso, D.; Serrano, J.; Nguyen, D.; Degovics, D.; et al. Topical Fluoxetine as a Novel Therapeutic That Improves Wound Healing in Diabetic Mice. Diabetes 2019, 68, 1499–1507. [Google Scholar] [CrossRef] [Green Version]
- Gittler, J.K.; Shemer, A.; Suárez-Fariñas, M.; Fuentes-Duculan, J.; Gulewicz, K.J.; Wang, C.Q.F.; Mitsui, H.; Cardinale, I.; de Guzman, C.S.; Krueger, J.G.; et al. Progressive Activation of T(H)2/T(H)22 Cytokines and Selective Epidermal Proteins Characterizes Acute and Chronic Atopic Dermatitis. J. Allergy Clin. Immunol. 2012, 130, 1344–1354. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Bao, L.; Chan, L.S.; DiPietro, L.A.; Chen, L. Aberrant Wound Healing in an Epidermal Interleukin-4 Transgenic Mouse Model of Atopic Dermatitis. PLoS ONE 2016, 11, e0146451. [Google Scholar] [CrossRef]
- Stutte, S.; Quast, T.; Gerbitzki, N.; Savinko, T.; Novak, N.; Reifenberger, J.; Homey, B.; Kolanus, W.; Alenius, H.; Förster, I. Requirement of CCL17 for CCR7- and CXCR4-Dependent Migration of Cutaneous Dendritic Cells. Proc. Natl. Acad. Sci. USA 2010, 107, 8736–8741. [Google Scholar] [CrossRef] [Green Version]
- Kolimi, P.; Narala, S.; Nyavanandi, D.; Youssef, A.A.A.; Dudhipala, N. Innovative Treatment Strategies to Accelerate Wound Healing: Trajectory and Recent Advancements. Cells 2022, 11, 2439. [Google Scholar] [CrossRef]
- Sugimoto, K.; Ishikawa, N.; Sugioka, T.; Koseki, H.; Kubosawa, H.; Kagawa, S.; Shimojo, N.; Ito, S.; Hattori, T. The Importance of Disinfection Therapy Using Povidone-Iodine Solution in Atopic Dermatitis. Dermatology 2002, 204 (Suppl. S1), 63–69. [Google Scholar] [CrossRef]
- Augustin, M.; Goepel, L.; Jacobi, A.; Bosse, B.; Mueller, S.; Hopp, M. Efficacy and Tolerability of Liposomal Polyvinylpyrrolidone-Iodine Hydrogel for the Localized Treatment of Chronic Infective, Inflammatory, Dermatoses: An Uncontrolled Pilot Study. Clin. Cosmet. Investig. Derm. 2017, 10, 373–384. [Google Scholar] [CrossRef] [Green Version]
- Murugaiyan, J.; Kumar, P.A.; Rao, G.S.; Iskandar, K.; Hawser, S.; Hays, J.P.; Mohsen, Y.; Adukkadukkam, S.; Awuah, W.A.; Jose, R.A.M.; et al. Progress in Alternative Strategies to Combat Antimicrobial Resistance: Focus on Antibiotics. Antibiotics 2022, 11, 200. [Google Scholar] [CrossRef]
- König, B.; Reimer, K.; Fleischer, W.; König, W. Effects of Betaisodona® on Parameters of Host Defense. DRM 1997, 195, 42–48. [Google Scholar] [CrossRef]
- Barakat, N.A.; Rasmy, S.A.; Hosny, A.E.-D.M.S.; Kashef, M.T. Effect of Povidone-Iodine and Propanol-Based Mecetronium Ethyl Sulphate on Antimicrobial Resistance and Virulence in Staphylococcus Aureus. Antimicrob. Resist. Infect. Control 2022, 11, 139. [Google Scholar] [CrossRef]
- Bigliardi, P.L.; Alsagoff, S.A.L.; El-Kafrawi, H.Y.; Pyon, J.-K.; Wa, C.T.C.; Villa, M.A. Povidone Iodine in Wound Healing: A Review of Current Concepts and Practices. Int. J. Surg. 2017, 44, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, M.J.; Westgate, S.J.; Mueller, S. Povidone-Iodine Ointment Demonstrates in Vitro Efficacy against Biofilm Formation. Int. Wound. J. 2017, 14, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Hulme, J. Application of Nanomaterials in the Prevention, Detection, and Treatment of Methicillin-Resistant Staphylococcus Aureus (MRSA). Pharmaceutics 2022, 14, 805. [Google Scholar] [CrossRef] [PubMed]
- Giau, V.V.; An, S.S.A.; Hulme, J. Recent Advances in the Treatment of Pathogenic Infections Using Antibiotics and Nano-Drug Delivery Vehicles. Drug Des. Dev. Ther. 2019, 13, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieber, T. Atopic Dermatitis: An Expanding Therapeutic Pipeline for a Complex Disease. Nat. Rev. Drug Discov. 2022, 21, 21–40. [Google Scholar] [CrossRef]
- Ito, Y.; Amagai, M. Controlling Skin Microbiome as a New Bacteriotherapy for Inflammatory Skin Diseases. Inflamm. Regen. 2022, 42, 26. [Google Scholar] [CrossRef]
- Yang, Y.; Qu, L.; Mijakovic, I.; Wei, Y. Advances in the Human Skin Microbiota and Its Roles in Cutaneous Diseases. Microb. Cell Factories 2022, 21, 176. [Google Scholar] [CrossRef]
- Giau, V.V.; Lee, H.; An, S.S.A.; Hulme, J. Recent Advances in the Treatment of C. Difficile Using Biotherapeutic Agents. Infect. Drug Resist. 2019, 12, 1597–1615. [Google Scholar] [CrossRef] [Green Version]
- Trzeciak, M.; Sakowicz-Burkiewicz, M.; Wesserling, M.; Dobaczewska, D.; Gleń, J.; Nowicki, R.; Pawelczyk, T. Expression of Cornified Envelope Proteins in Skin and Its Relationship with Atopic Dermatitis Phenotype. Acta Derm. Venereol. 2017, 97, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The Human Skin Microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- De Juan, A.; Segura, E. Modulation of Immune Responses by Nutritional Ligands of Aryl Hydrocarbon Receptor. Front. Immunol. 2021, 12, 1948. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, G.; Liu, X.; Shao, Y.; Gao, P.; Xin, C.; Cui, Z.; Zhao, X.; Xu, G. Serum Metabolomics Study and Eicosanoid Analysis of Childhood Atopic Dermatitis Based on Liquid Chromatography-Mass Spectrometry. J. Proteome Res. 2014, 13, 5715–5723. [Google Scholar] [CrossRef]
- Rothhammer, V.; Borucki, D.M.; Tjon, E.C.; Takenaka, M.C.; Chao, C.-C.; Ardura-Fabregat, A.; de Lima, K.A.; Gutiérrez-Vázquez, C.; Hewson, P.; Staszewski, O.; et al. Microglial Control of Astrocytes in Response to Microbial Metabolites. Nature 2018, 557, 724–728. [Google Scholar] [CrossRef]
- Schjødt, M.S.; Gürdeniz, G.; Chawes, B. The Metabolomics of Childhood Atopic Diseases: A Comprehensive Pathway-Specific Review. Metabolites 2020, 10, 511. [Google Scholar] [CrossRef]
- Kirjavainen, P.V.; Arvola, T.; Salminen, S.J.; Isolauri, E. Aberrant Composition of Gut Microbiota of Allergic Infants: A Target of Bifidobacterial Therapy at Weaning? Gut 2002, 51, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Staley, C.; Weingarden, A.R.; Khoruts, A.; Sadowsky, M.J. Interaction of Gut Microbiota with Bile Acid Metabolism and Its Influence on Disease States. Appl. Microbiol. Biotechnol. 2017, 101, 47–64. [Google Scholar] [CrossRef] [Green Version]
- Guzior, D.V.; Quinn, R.A. Review: Microbial Transformations of Human Bile Acids. Microbiome 2021, 9, 140. [Google Scholar] [CrossRef]
- Yu, J.; Luo, Y.; Zhu, Z.; Zhou, Y.; Sun, L.; Gao, J.; Sun, J.; Wang, G.; Yao, X.; Li, W. A Tryptophan Metabolite of the Skin Microbiota Attenuates Inflammation in Patients with Atopic Dermatitis through the Aryl Hydrocarbon Receptor. J. Allergy Clin. Immunol. 2019, 143, 2108–2119.e12. [Google Scholar] [CrossRef] [Green Version]
- Fang, Z.; Pan, T.; Wang, H.; Zhu, J.; Zhang, H.; Zhao, J.; Chen, W.; Lu, W. Limosilactobacillus Reuteri Attenuates Atopic Dermatitis via Changes in Gut Bacteria and Indole Derivatives from Tryptophan Metabolism. Int. J. Mol. Sci. 2022, 23, 7735. [Google Scholar] [CrossRef]
- Sugiura, K. Unfolded Protein Response in Keratinocytes: Impact on Normal and Abnormal Keratinization. J. Dermatol. Sci. 2013, 69, 181–186. [Google Scholar] [CrossRef]
- Nakada, E.M.; Bhakta, N.R.; Korwin-Mihavics, B.R.; Kumar, A.; Chamberlain, N.; Bruno, S.R.; Chapman, D.G.; Hoffman, S.M.; Daphtary, N.; Aliyeva, M.; et al. Conjugated Bile Acids Attenuate Allergen-Induced Airway Inflammation and Hyperresponsiveness by Inhibiting UPR Transducers. JCI Insight 2019, 4, e98101. [Google Scholar] [CrossRef] [PubMed]
- Van Giau, V.; Wu, S.Y.; Jamerlan, A.; An, S.S.A.; Kim, S.Y.; Hulme, J. Gut Microbiota and Their Neuroinflammatory Implications in Alzheimer’s Disease. Nutrients 2018, 10, 1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, I.; Cha, A.; Lee, H.; Yoon, H.; Yoon, T.; Cho, B.; Lee, S.; Park, Y. N-3 Polyunsaturated Fatty Acids and Atopy in Korean Preschoolers. Lipids 2007, 42, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Reddel, S.; Del Chierico, F.; Quagliariello, A.; Giancristoforo, S.; Vernocchi, P.; Russo, A.; Fiocchi, A.; Rossi, P.; Putignani, L.; El Hachem, M. Gut Microbiota Profile in Children Affected by Atopic Dermatitis and Evaluation of Intestinal Persistence of a Probiotic Mixture. Sci. Rep. 2019, 9, 4996. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kim, K.; Kim, W. Gut Microbiota Restoration through Fecal Microbiota Transplantation: A New Atopic Dermatitis Therapy. Exp. Mol. Med. 2021, 53, 907–916. [Google Scholar] [CrossRef]
- Mashiah, J.; Karady, T.; Fliss-Isakov, N.; Sprecher, E.; Slodownik, D.; Artzi, O.; Samuelov, L.; Ellenbogen, E.; Godneva, A.; Segal, E.; et al. Clinical Efficacy of Fecal Microbial Transplantation Treatment in Adults with Moderate-to-Severe Atopic Dermatitis. Immun. Inflamm. Dis. 2022, 10, e570. [Google Scholar] [CrossRef]
- Graham, E.H.; Clarke, J.L.; Fernando, S.C.; Herr, J.R.; Adamowicz, M.S. The Application of the Skin Virome for Human Identification. Forensic Sci. Int. Genet. 2022, 57, 102662. [Google Scholar] [CrossRef]
- Mujtaba, M.G.; Johnson, H.M.; Parrish, J.M. Staphylococcal Enterotoxin Superantigens Induce Prophylactic Antiviral Activity Against Encephalomyocarditis Virus In Vivo and In Vitro. Viral. Immunol. 2021, 34, 392–400. [Google Scholar] [CrossRef]
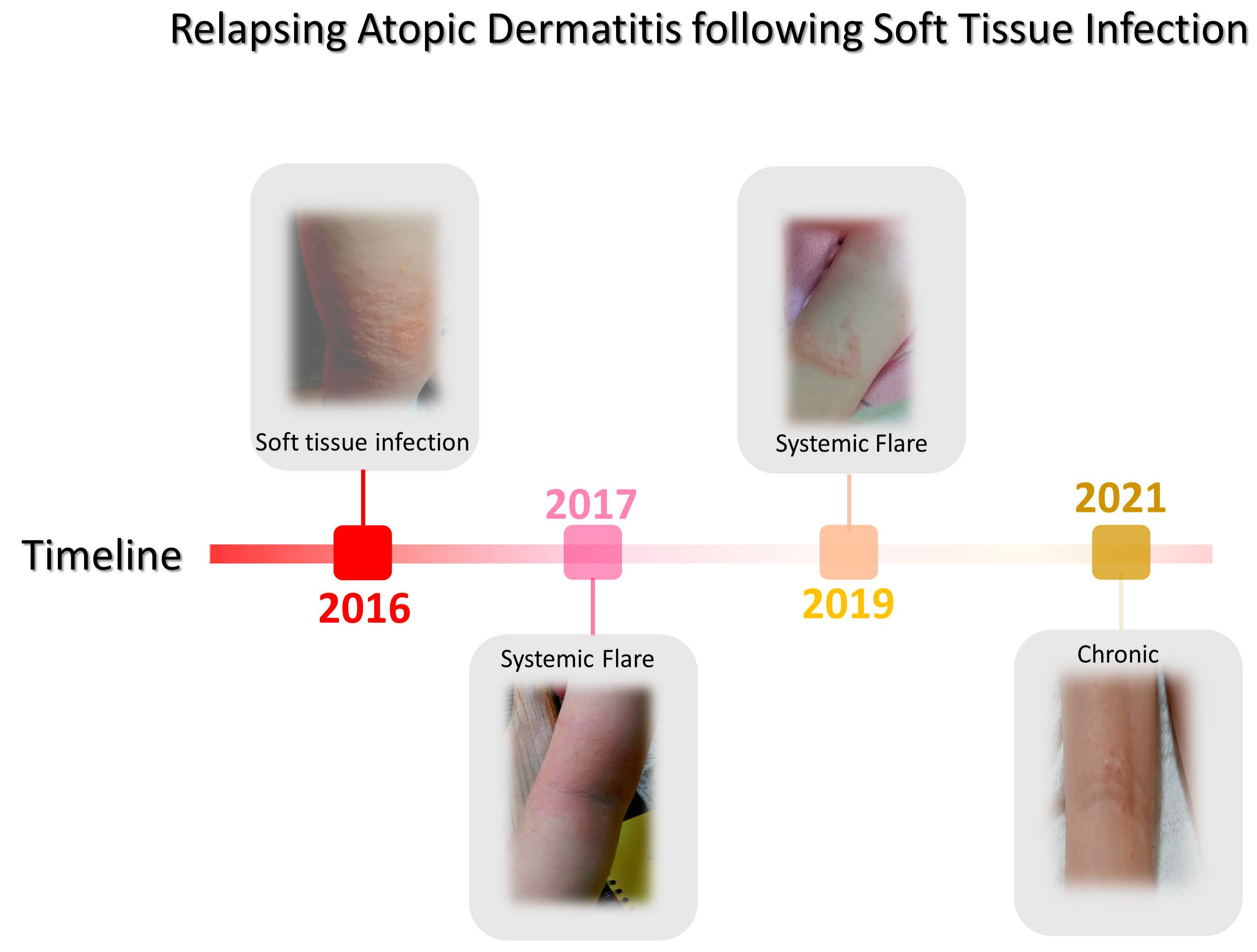
Figure 1.
Interactions between SA cell wall proteins and keratinocytes. Cell wall proteins, clumping factor A (ClfA), fibronectin-binding proteins (FnBP), autolysin (Atl), and serine aspartate repeat-containing protein D(SdrD) involved in adhesion and keratinocyte internalization via respective receptors (heat shock protein 60 (Hsp69), Integrinα5β1, Annexin2, with fibronectin bridging, Desmoglein1, and Heat shock cognate 71 kDa protein (Hsc70)). In addition, PA induces inflammation by binding to tumor necrosis factor receptor-1 (TNFR1), and Toll-like receptor 2 (TLR2) senses lipoprotein following the activation of immune cells. Reproduced with permission from Elsevier [59].
Figure 1.
Interactions between SA cell wall proteins and keratinocytes. Cell wall proteins, clumping factor A (ClfA), fibronectin-binding proteins (FnBP), autolysin (Atl), and serine aspartate repeat-containing protein D(SdrD) involved in adhesion and keratinocyte internalization via respective receptors (heat shock protein 60 (Hsp69), Integrinα5β1, Annexin2, with fibronectin bridging, Desmoglein1, and Heat shock cognate 71 kDa protein (Hsc70)). In addition, PA induces inflammation by binding to tumor necrosis factor receptor-1 (TNFR1), and Toll-like receptor 2 (TLR2) senses lipoprotein following the activation of immune cells. Reproduced with permission from Elsevier [59].

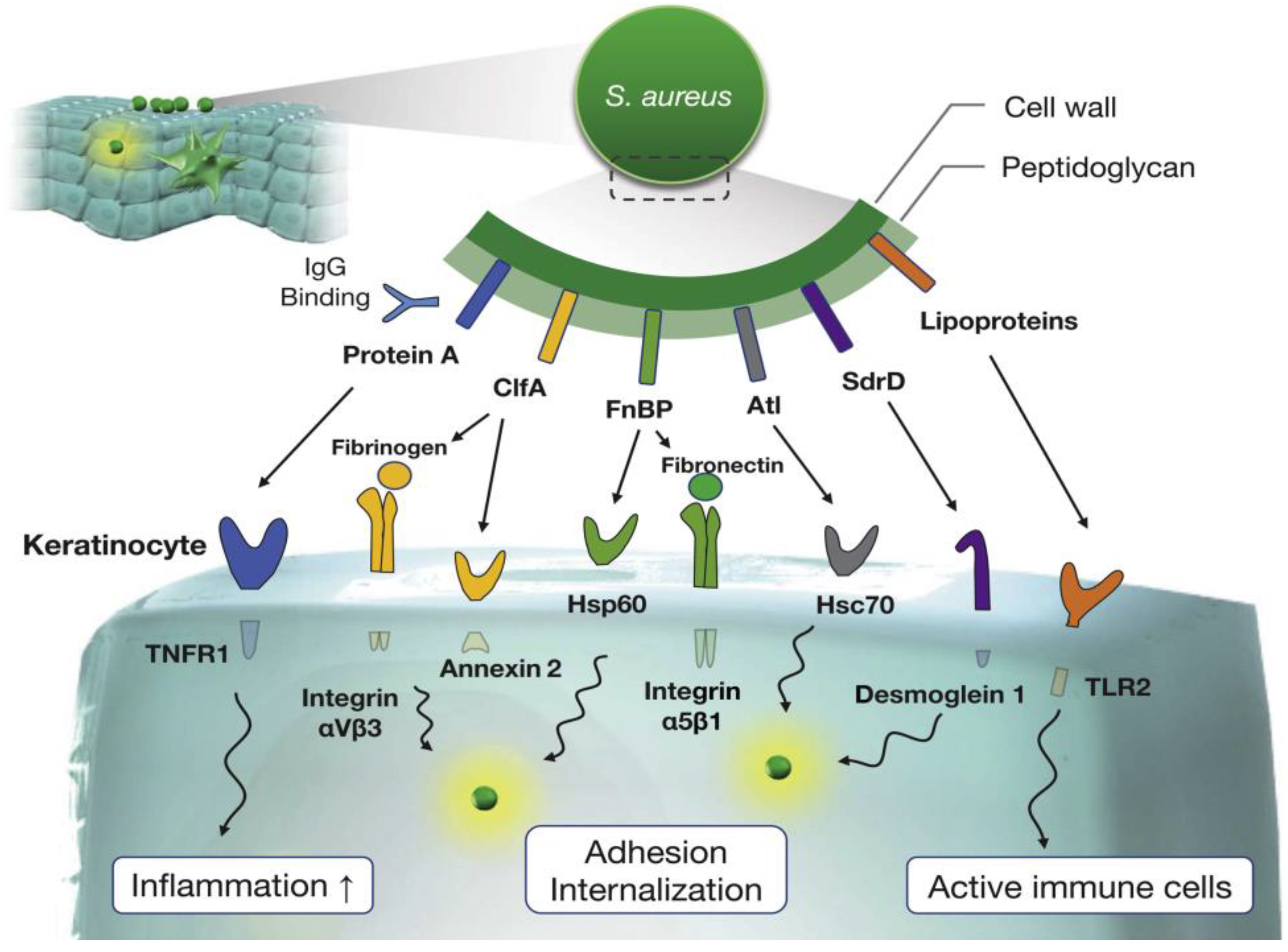
Figure 2.
Photograph of SA-infected abrasion (patient X (Age 9-female); 4 October 2016) following administration (1 October 2016) of low-dose erythromycin and corticosteroids (A). Permanganate (aq) cleansing, followed by administration of high-strength oral antibiotics (amoxicillin) (5 October 2016); overnight resolution of non-localized symptoms (neck, head, and trunk) highest recorded patient temperature 38.5 °C. Day 30 barrier restored (B). (i) Infected area, (ii) immunological reaction to SA toxins, (iii) honey-colored fluid, (iv) skin repaired, sporadic folliculitis. Modified with permission from Springer [63].
Figure 2.
Photograph of SA-infected abrasion (patient X (Age 9-female); 4 October 2016) following administration (1 October 2016) of low-dose erythromycin and corticosteroids (A). Permanganate (aq) cleansing, followed by administration of high-strength oral antibiotics (amoxicillin) (5 October 2016); overnight resolution of non-localized symptoms (neck, head, and trunk) highest recorded patient temperature 38.5 °C. Day 30 barrier restored (B). (i) Infected area, (ii) immunological reaction to SA toxins, (iii) honey-colored fluid, (iv) skin repaired, sporadic folliculitis. Modified with permission from Springer [63].

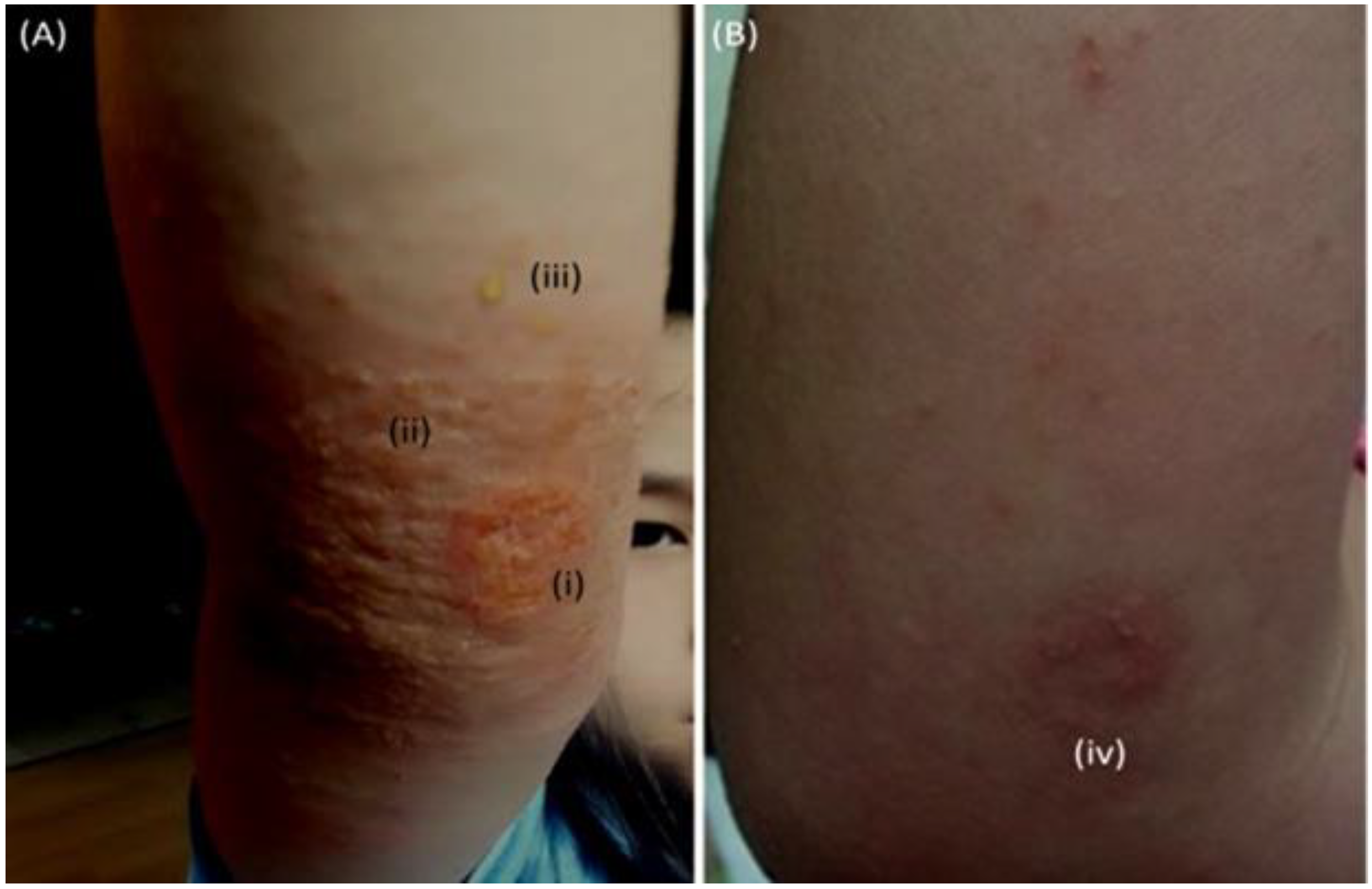
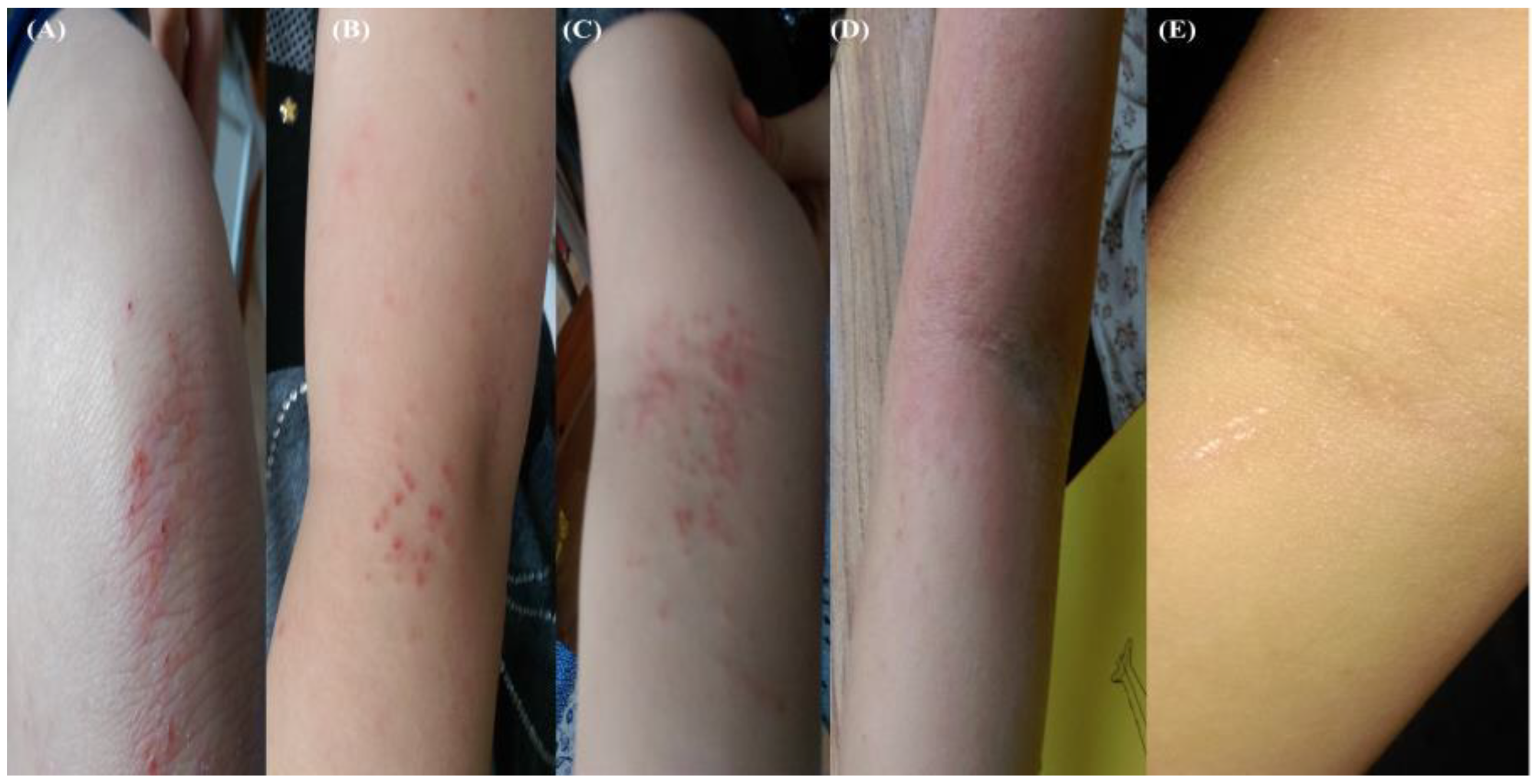
Figure 3.
Suspected stress-induced flare without infection. Photograph (10 October 2017) of grazed knee, healing time 10 days (A). Photograph (11 October 2017): flaring, outer right (B) and inner left forearms (C). Photograph (26 October 2017): severe inflammation, Fucidin and mupirocin treatments discontinued, bacterial and fungal (Candida and Malassezia) involvement suspected (D). Resolution of AD symptoms following daily disinfection (1% Chloroxylenol spraying) of bedding covers (E).
Figure 3.
Suspected stress-induced flare without infection. Photograph (10 October 2017) of grazed knee, healing time 10 days (A). Photograph (11 October 2017): flaring, outer right (B) and inner left forearms (C). Photograph (26 October 2017): severe inflammation, Fucidin and mupirocin treatments discontinued, bacterial and fungal (Candida and Malassezia) involvement suspected (D). Resolution of AD symptoms following daily disinfection (1% Chloroxylenol spraying) of bedding covers (E).

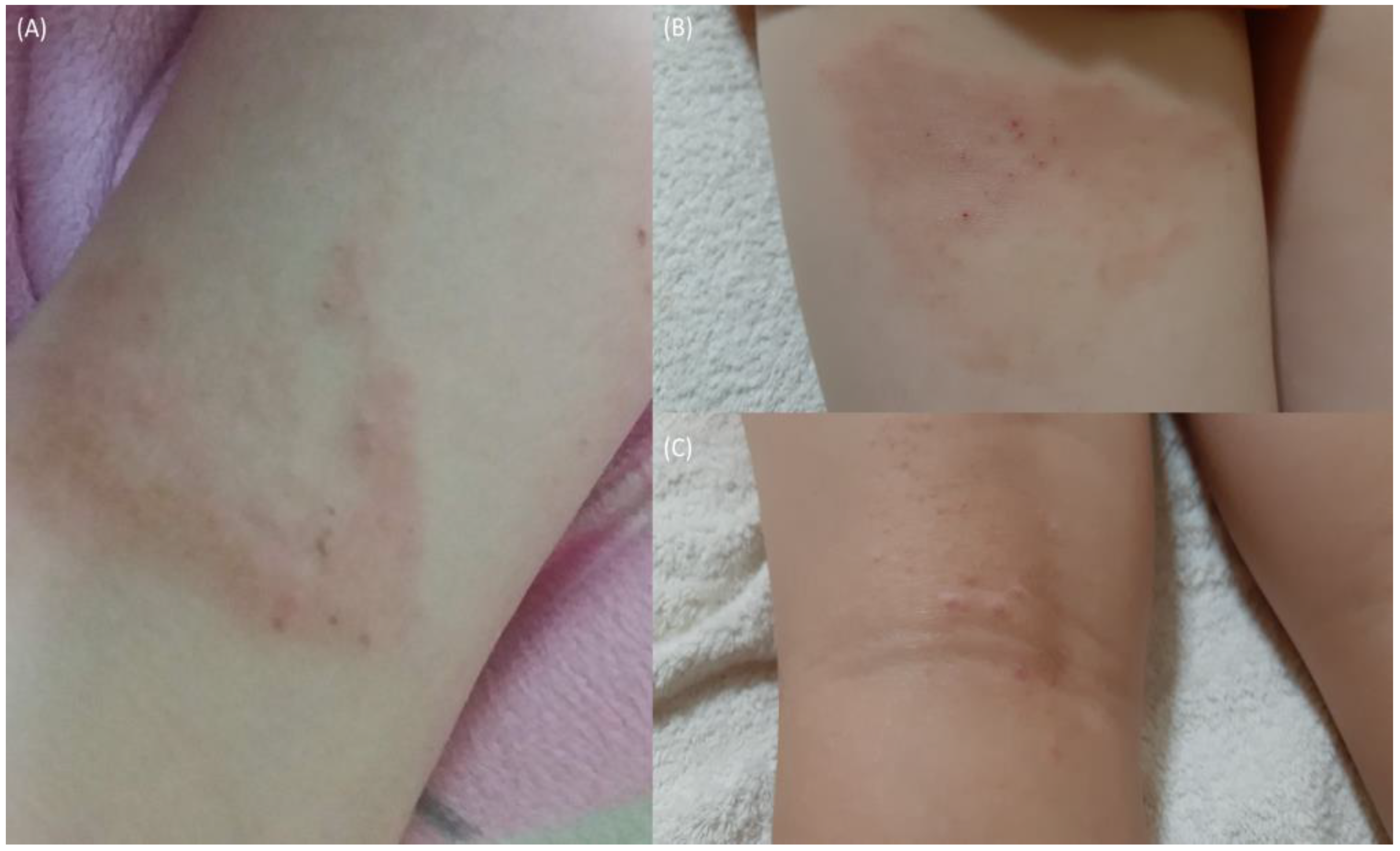
Figure 4.
Recurrent AD flaring. (A) (16 March 2019) heat and pressure-triggered flare (posterior left knee). (B) (16 June 2021) heat and pressure-triggered flare (posterior left thigh). (C) (16 June 2021) scarring and nodulation (posterior left knee) following repeated flaring.
Figure 4.
Recurrent AD flaring. (A) (16 March 2019) heat and pressure-triggered flare (posterior left knee). (B) (16 June 2021) heat and pressure-triggered flare (posterior left thigh). (C) (16 June 2021) scarring and nodulation (posterior left knee) following repeated flaring.

Figure 5.
Regional distribution of microorganisms in human skin. Dry (forearm and palm), sebaceous (head, back, and chest), moist (groin, bend of elbow, and inguinal canal), and foot (plantar heel and toe web space). Wide and narrow bar charts represent the anatomical location and relative abundance of microorganisms distributed in those areas. Reproduced with permission from Springer [87].
Figure 5.
Regional distribution of microorganisms in human skin. Dry (forearm and palm), sebaceous (head, back, and chest), moist (groin, bend of elbow, and inguinal canal), and foot (plantar heel and toe web space). Wide and narrow bar charts represent the anatomical location and relative abundance of microorganisms distributed in those areas. Reproduced with permission from Springer [87].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Microbial skin infections and resultant changes in immune function and microbial composition associated with AD.
Table 1.
Microbial skin infections and resultant changes in immune function and microbial composition associated with AD.
| Infectious Species | Clinical Features | Immune Dysfunction | Microbiome | Reference |
|---|---|---|---|---|
| S. aureus and Methicillin-resistant Staphylococcus aureus (MRSA) | Weeping, honey-colored crusts and pustules, both interfollicular and follicular-based (folliculitis) Abscesses, fever, and lymphadenopathy | ↓Antimicrobial peptides ↑IL-13, IL-4 B-cell Ig class switching to IgE ↑type 2–related chemokines (CCL13, CCL17, CCL18, and CCL22) ↑Degradation of immunoglobulin G (IgG) | ↓ coagulase-negative Staphylococci (CoNS) (S. epidermidis, S. hominis, and S. lugdunensis) ↑ S. aureus | [14,15] |
| Beta-hemolytic streptococcal | Bright red erythema, thick-walled pustules, and heavy crusting | ↑Degradation of IgA, IgM, IgD, and IgE | ↑ S. aureus | [16] |
| Herpes simplex virus (HSV) molluscum contagiosum (MC), eczema vaccinia (EV), and eczema coxsackium (EC) | Superficial clusters of dome-shaped vesicles and/or small, round, punched-out erosions | ↑IL-13 and IL-4 ↓ IFN-γ and TNF-α | ↑ S. aureus | [17] |
| Malassezia globosa and Malassezia restricta nanovesicles | Pruritic monomorphous papules and/or pustules. Hypo- or hyper-pigmented non-inflammatory lesions | ↑ IgE↑ auto-reactive T cells induces autoreactivity to human proteins | ↓S. aureus | [18] |
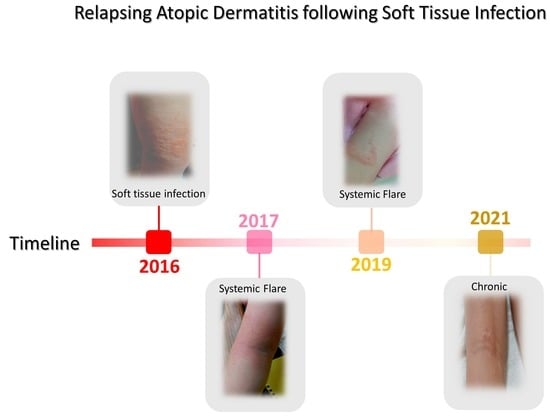
Table 2.
Changes in the SCORAD value of patient X in response to various triggers.
| Date & Location | Trigger | Morphological Description | SCORAD | Treatment | Time to Resolution | Reference |
|---|---|---|---|---|---|---|
| 4 October 2016 A* | Fall and SA-infected abrasion | Marked erythema (deep or bright red), papulation; disease is widespread in extent | 56.87 | Permanganate (aq) cleansing high-strength oral antibiotics (amoxicillin) | 30 days | Figure 2A,B [63] |
| 26 October 2017 B* | Fall and abrasion | Perceptible erythema clearly perceptible induration/papulation | 43.7 | 1% Chloroxylenol bathing prior sleep | 16 days | Figure 3A–E |
| 27 December 2017 B* | Stress | Slight but definite erythema (pink), slight but definite induration | 19.45 | 1% Chloroxylenol bathing prior sleep | 12 days | N/A |
| 27 January 2018 A* | Fall and abrasion | Perceptible erythema induration/papulation | 26.9 | 1% Chloroxylenol bathing prior sleep | 16 days | N/A |
| 16 March 2019 A* | Heat and pressure | Perceptible scarring and Skin thickening (lichenification), itching | 27.8 | UV-B and exercise | 14 days | Figure 4A |
| 16 June 2021 C* | Heat and pressure | Perceptible scarring, persistent nodulation, itching Skin thickening (lichenification) | 21.8 | UV-B and exercise | 10 days | Figure 4B,C |
| 10 March 2022 C* | Foot wound | No inflammatory signs of local or systemic atopic dermatitis; nodulation and occasional itching | 7.4 | Topical application of Povidone | 5 days | N/A |
SCORAD values were assessed using photographs (Figure 2, Figure 3 and Figure 4) and at A* (Eulji University Hospital Daejeon South Korea), B* (Royal Manchester Children’s Hospital UK), C*, and at home by guardians utilizing online software (https://scorad.corti.li/ accessed on 16 March 2019). Permission for other photographs for 27 December 2017, 27 January 2018, and 10 March 2022 was not granted (N/A).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hulme, J. Staphylococcus Infection: Relapsing Atopic Dermatitis and Microbial Restoration. Antibiotics 2023, 12, 222. https://doi.org/10.3390/antibiotics12020222
AMA Style
Hulme J. Staphylococcus Infection: Relapsing Atopic Dermatitis and Microbial Restoration. Antibiotics. 2023; 12(2):222. https://doi.org/10.3390/antibiotics12020222
Chicago/Turabian StyleHulme, John. 2023. "Staphylococcus Infection: Relapsing Atopic Dermatitis and Microbial Restoration" Antibiotics 12, no. 2: 222. https://doi.org/10.3390/antibiotics12020222
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.