
The Prevalence and Molecular Biology of Staphylococcus aureus Isolated from Healthy and Diseased Equine Eyes in Egypt

 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Design
2.2. Sample Collection
2.3. Isolation and Identification of Staphylococcus Species
2.4. Antibiotic Susceptibility Testing
2.5. DNA Extraction and PCR
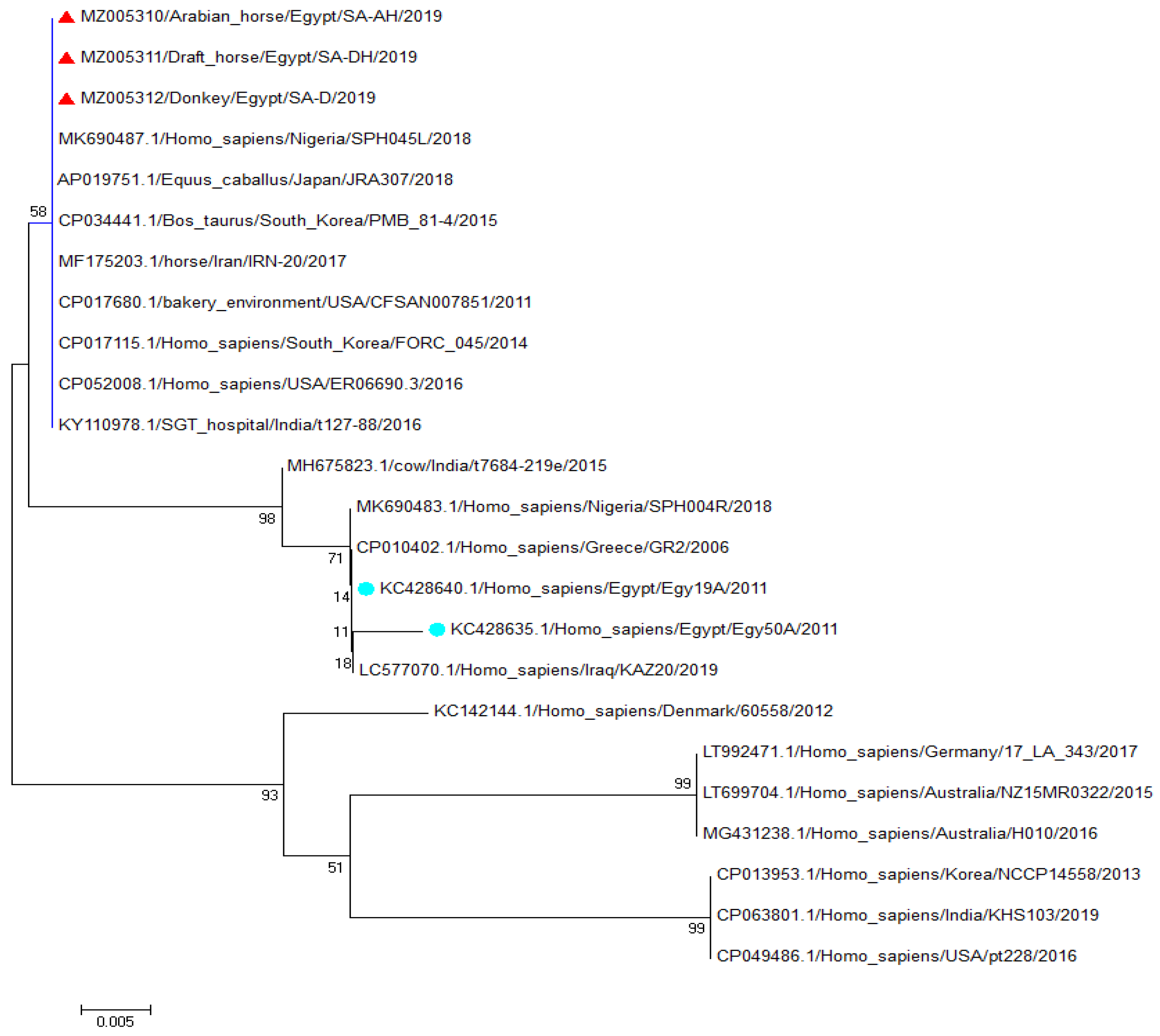
2.6. Sequencing and Phylogenetic Analysis

{kind=link}
| Target Gene | Primer Sequences | Amplified Segment (bp) | Primary Denaturation | Amplification (35 Cycles) | Final Extension | Reference | ||
|---|---|---|---|---|---|---|---|---|
| Secondary Denaturation | Annealing | Extension | ||||||
| spa (x region) | For CAA GCA CCA AAA GAG GAA | Variable | 94 °C 5 min. | 94 °C 30 s. | 60 °C 40 s. | 72 °C 40 s. | 72 °C 7 min. | [29] |
| Rev CAC CAG GTT TAA CGA CAT | ||||||||
| nuc | For ATATGTATGGCAATCGTTTCAAT | 395 | 94 °C 5 min. | 94 °C 30 s. | 55 °C 40 s. | 72 °C 40 s. | 72 °C 7 min. | [30] |
| Rev GTAAATGCACTTGCTTCAGGAC | ||||||||
| mecA | For GTA GAA ATG ACT GAA CGT CCG ATA A | 310 | 94 °C 5 min. | 94 °C 30 s. | 50 °C 30 s. | 72 °C 30 s. | 72 °C 7 min. | [31] |
| Rev CCA ATT CCA CAT TGT TTC GGT CTA A | ||||||||
| hlg | For GCCAATCCGTTATTAGAAAATGC | 937 | 94 °C 5 min. | 94 °C 30 s. | 55 °C 40 s. | 72 °C 1 min. | 72 °C 10 min. | [32] |
| Rev CCATAGACGTAGCAACGGAT | ||||||||
| Staph 16sRNA | For AAC TCT GTT ATT AGG GAA GAA CA | 756 | 94 °C 5 min. | 94 °C 30 s. | 60 °C 40 s. | 72 °C 1 min | 72 °C 10 min | [33] |
| Rev CCA CCT TCC TCC GGT TTG TCA CC | ||||||||
2.7. Statistical Analysis
3. Results
3.1. Identification and Epidemiology of S. aureus in Healthy and Diseased Equine Eyes
3.2. Sequencing and Phylogenetic Analysis of the Spa (x Region) Gene Fragment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hsiao, C.-H.; Chuang, C.-C.; Tan, H.Y.; Ma, D.H.K.; Lin, K.-K.; Chang, C.-J.; Huang, Y.C. Methicillin-resistant Staphylococcus aureus ocular infection: A 10-year hospital-based study. Ophthalmology 2012, 119, 522–527. [Google Scholar] [CrossRef]
- Parhizgari, N.; Khoramrooz, S.S.; Hosseini, S.A.A.M.; Marashifard, M.; Yazdanpanah, M.; Emaneini, M.; Gharibpour, F.; Mirzaii, M.; Darban-Sarokhalil, D.; Moein, M. High frequency of multidrug-resistant Staphylococcus aureus with Scc, Mec, Type III, and Spa types T037 and T631 isolated from burn patients in southwest Iran. Apmis 2016, 124, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Paschalis-Trela, K.; Cywinska, A. The prevalence of ocular diseases in Polish Arabian horses. BMC Vet. Res. 2017, 13, 319. [Google Scholar] [CrossRef]
- Gilger, B.C.; Deeg, C. Equine recurrent uveitis. In Equine Ophthalmology; Elsevier: Amsterdam, The Netherlands, 2011; pp. 317–349. [Google Scholar]
- Johns, I.C.; Baxter, K.; Booler, H.; Hicks, C.; Menzies-Gow, N. Conjunctival bacterial and fungal flora in healthy horses in the UK. Vet. Ophthalmol. 2011, 14, 195–199. [Google Scholar] [CrossRef]
- Foti, M.; Fisichella, V.; Giacopello, C. Detection of methicillin-resistant Staphylococcus aureus (MRSA) in the microbial flora from the conjunctiva of healthy donkeys from Sicily (Italy). Vet. Ophthalmol. 2013, 16, 89–92. [Google Scholar] [CrossRef]
- Camargo, I.L.B.C.; Gilmore, M.S. Staphylococcus aureus—Probing for host weakness? J. Bacteriol. 2008, 190, 2253–2256. [Google Scholar] [CrossRef] [Green Version]
- Wieler, L.H.; Ewers, C.; Guenther, S.; Walther, B.; Lübke-Becker, A. Methicillin-resistant staphylococci (MRS) and extended-spectrum beta-lactamases (ESBL)-producing enterobacteriaceae in companion animals: Nosocomial infections as one reason for the rising prevalence of these potential zoonotic pathogens in clinical samples. Int. J. Med. Microbiol. 2011, 301, 635–641. [Google Scholar] [PubMed]
- Walther, B.; Tedin, K.; Lübke-Becker, A. Multidrug-resistant opportunistic pathogens challenging veterinary infection control. Vet. Microbiol. 2017, 200, 71–78. [Google Scholar] [CrossRef]
- Baptiste, K.E.; Williams, K.; Willams, N.J.; Wattret, A.; Clegg, P.D.; Dawson, S.; Corkill, J.E.; O’Neill, T.; Hart, C.A. Methicillin-resistant staphylococci in companion animals. Emerg. Infect. Dis. 2005, 11, 1942. [Google Scholar] [CrossRef]
- Vengust, M.; Anderson, M.E.C.; Rousseau, J.; Weese, J.S. Methicillin-resistant staphylococcal colonization in clinically normal dogs and horses in the community. Lett. Appl. Microbiol. 2006, 43, 602–606. [Google Scholar] [CrossRef]
- Weese, J.S.; Rousseau, J.; Traub-Dargatz, J.L.; Willey, B.M.; McGeer, A.J.; Low, D.E. Community-associated methicillin-resistant Staphylococcus aureus in horses and humans who work with horses. J. Am. Vet. Med. Assoc. 2005, 226, 580–583. [Google Scholar] [CrossRef]
- Leonard, F.C.; Markey, B.K. Methicillin-resistant Staphylococcus aureus in animals: A review. Vet. J. 2008, 175, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Strommenger, B.; Kehrenberg, C.; Kettlitz, C.; Cuny, C.; Verspohl, J.; Witte, W.; Schwarz, S. Molecular characterization of methicillin-resistant Staphylococcus aureus strains from pet animals and their relationship to human isolates. J. Antimicrob. Chemother. 2006, 57, 461–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Duijkeren, E.; Moleman, M.; van Oldruitenborgh-Oosterbaan, M.M.S.; Multem, J.; Troelstra, A.; Fluit, A.C.; Van Wamel, W.B.J.; Houwers, D.J.; De Neeling, A.J.; Wagenaar, J.A. Methicillin-resistant Staphylococcus aureus in horses and horse personnel: An investigation of several outbreaks. Vet. Microbiol. 2010, 141, 96–102. [Google Scholar] [CrossRef]
- Waqar, N.; Amin, Q.; Munir, T.; Ikram, M.S.; Shahzad, N.; Mirza, A.; Ali, A.; Arshad, M.I. A cross-sectional study of methicillin-resistant Staphylococcus aureus at the equine-human interface. Trop. Anim. Health Prod. 2019, 51, 1927–1933. [Google Scholar] [CrossRef]
- Plummer, C.E. Corneal response to injury and infection in the horse. Vet. Clin. Equine Pract. 2017, 33, 439–463. [Google Scholar] [CrossRef] [PubMed]
- Walther, B.; Wieler, L.H.; Friedrich, A.W.; Hanssen, A.M.; Kohn, B.; Brunnberg, L.; Lübke-Becker, L. Methicillin-resistant Staphylococcus aureus (MRSA) isolated from small and exotic animals at a university hospital during routine microbiological examinations. Vet. Microbiol. 2008, 127, 171–178. [Google Scholar] [CrossRef]
- Vincze, S.; Stamm, I.; Kopp, P.A.; Hermes, J.; Adlhoch, C.; Semmler, T.; Wieler, L.H.; Lübke-Becker, A.; Walther, B. Alarming proportions of methicillin-resistant Staphylococcus aureus (MRSA) in wound samples from companion animals, Germany 2010–2012. PLoS ONE 2014, 9, e85656. [Google Scholar] [CrossRef] [Green Version]
- Diene, S.M.; Corvaglia, A.R.; François, F.; van der Mee-Marquet, N. Prophages and adaptation of Staphylococcus aureus St398 to the human clinic. BMC Genom. 2017, 18, 133. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.R.d.A.; Santana, A.F.; Almeida, A.C.d.V.R.; Sousa, R.F.; Perecmanis, S.; Galera, P.D. Bacterial culture and antibiotic sensitivity from the ocular conjunctiva of horses. Ciência Rural 2017, 47, 19. [Google Scholar] [CrossRef]
- Kahlmeter, G.; Brown, D.F.; Goldstein, F.W.; MacGowan, A.P.; Mouton, J.W.; Odenholt, I.; Rodloff, A.; Soussy, C.J.; Steinbakk, M.; Soriano, F.; et al. European Committee on Antimicrobial Susceptibility Testing (EUCAST) technical notes on antimicrobial susceptibility testing. Clin. Microbiol. Infect. 2006, 12, 501–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauer, P.; Andrew, S.E.; Lassaline, M.; Gelatt, K.N.; Denis, H.M. Changes in antibiotic resistance in equine bacterial ulcerative keratitis (1991–2000): 65 horses. Vet. Ophthalmol. 2003, 6, 309–313. [Google Scholar] [CrossRef]
- Wada, S.; Hobo, S.; Niwa, H. Ulcerative keratitis in thoroughbred racehorses in Japan from 1997 to 2008. Vet. Ophthalmol. 2010, 13, 99–105. [Google Scholar] [CrossRef]
- Kuroda, T.; Kinoshita, Y.; Niwa, H.; Mizobe, F.; Ueno, T.; Kuwano, A.; Hatazoe, T.; Hobo, S. Methicillin-resistant Staphylococcus aureus ulcerative keratitis in a thoroughbred racehorse. J. Equine Sci. 2015, 26, 95–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. Clustal W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Momtaz, H.; Dehkordi, F.S.; Rahimi, E.; Asgarifar, A.; Momeni, M. Virulence genes and antimicrobial resistance profiles of Staphylococcus aureus isolated from chicken meat in Isfahan Province, Iran. J. Appl. Poult. Res. 2013, 22, 913–921. [Google Scholar] [CrossRef]
- Gao, J.; Ferreri, M.; Liu, X.Q.; Chen, L.B.; Su, J.L.; Han, B. Development of multiplex polymerase chain reaction assay for rapid detection of Staphylococcus aureus and selected antibiotic resistance genes in bovine mastitic milk samples. J. Vet. Diagn. Investig. 2011, 23, 894–901. [Google Scholar] [CrossRef] [Green Version]
- McClure, J.A.; Conly, J.M.; Lau, V.; Elsayed, S.; Louie, T.; Hutchins, W.; Zhang, K. Novel multiplex PCR assay for detection of the staphylococcal virulence marker Panton-Valentine leukocidin genes and simultaneous discrimination of methicillin-susceptible from resistant staphylococci. J. Clin. Microbiol. 2006, 44, 1141–1144. [Google Scholar] [CrossRef] [Green Version]
- Kumar, J.; Dinesh, Y.K.N.; Gaur, A.; Khanna, D. Detection of virulence genes in Staphylococcus aureus isolated from paper currency. Int. J. Infect. Dis. 2009, 13, e450–e455. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Sparling, J.; Chow, B.L.; Elsayed, S.; Hussain, Z.; Church, D.L.; Gregson, D.B.; Louie, T.; Conly, J.M. New quadriplex PCR assay for detection of methicillin and mupirocin resistance and simultaneous discrimination of Staphylococcus aureus from coagulase-negative staphylococci. J. Clin. Microbiol. 2004, 42, 4947–4955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamilmahan, P.; Zama, M.M.S.; Pathak, R.; Muneeswaran, N.S.; Karthik, K. A retrospective study of ocular occurrence in domestic animals: 799 cases. Vet. World 2013, 6, 274–276. [Google Scholar] [CrossRef]
- Scantlebury, C.E.; Aklilu, N.; Reed, K.; Knottenbelt, D.C.; Gebreab, F.; Pinchbeck, G.L. Ocular disease in working horses in Ethiopia: A cross-sectional study. Vet. Rec. 2013, 172, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vola, M.E.; Moriyama, A.S.; Lisboa, R.; Vola, M.M.; Hirai, F.E.; Bispo, P.J.M.; Höfling-Lima, A.L. Prevalence and antibiotic susceptibility of methicillin-resistant Staphylococcus aureus in ocular infections. Arq. Bras. Oftalmol. 2013, 76, 350–353. [Google Scholar] [CrossRef] [Green Version]
- Malalana, F.; McGowan, T.W.; Ireland, J.L.; Pinchbeck, G.L.; McGowan, C.M. Prevalence of owner-reported ocular problems and veterinary ocular findings in a population of horses aged ≥15 years. Equine Vet. J. 2019, 51, 212–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramesh, S.; Ramakrishnan, R.; Bharathi, M.J.; Amuthan, M.; Viswanathan, S. Prevalence of bacterial pathogens causing ocular infections in South India. Indian J. Pathol. Microbiol. 2010, 53, 281. [Google Scholar] [PubMed]
- Timney, B.; Macuda, T. Vision and hearing in horses. J. Am. Vet. Med. Assoc. 2001, 218, 1567–1574. [Google Scholar] [CrossRef]
- Plata, K.; Rosato, A.E.; Wegrzyn, G. Staphylococcus aureus as an infectious agent: Overview of biochemistry and molecular genetics of its pathogenicity. Acta Biochim. Pol. 2009, 56, 597–612. [Google Scholar] [CrossRef]
- Chambers, H.F.; DeLeo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef]
- Soimala, T.; Lübke-Becker, A.; Schwarz, S.; Feßler, A.T.; Huber, C.; Semmler, T.; Merle, R.; Gehlen, H.; Eule, J.C.; Walther, B. Occurrence and molecular composition of methicillin-resistant Staphylococcus aureus isolated from ocular surfaces of horses presented with ophthalmologic disease. Vet. Microbiol. 2018, 222, 1–6. [Google Scholar] [CrossRef]
- Bagga, B.; Reddy, A.K.; Garg, P. Decreased susceptibility to quinolones in methicillin-resistant Staphylococcus aureus isolated from ocular infections at a tertiary eye care centre. Br. J. Ophthalmol. 2010, 94, 1407–1408. [Google Scholar] [CrossRef]
- Schmitz, F.J.; Krey, A.; Sadurski, R.; Verhoef, J.; Milatovic, D.; Fluit, A.C. European Sentry Participants. Resistance to tetracycline and distribution of tetracycline resistance genes in European Staphylococcus aureus isolates. J. Antimicrob. Chemother 2001, 47, 239–240. [Google Scholar] [CrossRef] [Green Version]
- Zilberberg, M.D.; Shorr, A.F.; Kollef, M.F. Growth and geographic variation in hospitalizations with resistant infections, United States, 2000–2005. Emerg. Infect. Dis. 2008, 14, 1756. [Google Scholar] [CrossRef]
- Lakshmi, V.; Ashok, R.; Susmita, J.; Shailaja, V.V. Changing trends in the antibiograms of Salmonella isolates at a tertiary care hospital in Hyderabad. Indian J. Med. Microbiol. 2006, 24, 45–48. [Google Scholar] [CrossRef]
- Hammad, O.M.; Hifnawy, T.; Omran, D.; Tantawi, M.A.E.; Girgis, N.I. Ceftriaxone versus chloramphenicol for treatment of acute typhoid fever. Life Sci. J. 2011, 8, 100–105. [Google Scholar]
- Graille, M.; Stura, E.A.; Corper, A.L.; Sutton, B.J.; Taussig, M.J.; Charbonnier, J.-B.; Silverman, G.J. Crystal structure of a Staphylococcus aureus protein A domain complexed with the Fab fragment of a human IgM antibody: Structural basis for recognition of B-cell receptors and superantigen activity. Proc. Natl. Acad. Sci. USA 2000, 97, 5399–5404. [Google Scholar] [CrossRef] [Green Version]
- Keener, A.B.; Thurlow, L.T.; Kang, S.A.; Spidale, N.A.; Clarke, S.H.; Cunnion, K.M.; Tisch, R.; Richardson, A.R.; Vilen, B.J. Staphylococcus aureus protein A disrupts immunity mediated by long-lived plasma cells. J. Immunol. 2017, 198, 1263–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shopsin, B.; Gomez, M.; Montgomery, S.O.; Smith, D.H.; Waddington, M.; Dodge, D.E.; Bost, D.A.; Riehman, M.; Naidich, S.; Kreiswirth, B.N. Evaluation of protein A gene polymorphic region DNA sequencing for typing of Staphylococcus aureus strains. J. Clin. Microbiol. 1999, 37, 3556–3563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asadollahi, P.; Farahani, N.N.; Mirzaii, M.; Khoramrooz, S.S.; Van Belkum, A.; Asadollahi, K.; Dadashi, M.; Darban-Sarokhalil, D. Distribution of the most prevalent Spa types among clinical isolates of methicillin-resistant and-susceptible Staphylococcus aureus around the world: A review. Front. Microbiol. 2018, 9, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Donk, C.F.M.; Schols, J.M.G.A.; Schneiders, V.; Grimm, K.-H.; Stobberingh, E.E. Antibiotic resistance, population structure and spread of Staphylococcus aureus in nursing homes in the Euregion Meuse-Rhine. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1483–1489. [Google Scholar] [CrossRef] [PubMed]
- Berends, E.T.M.; Horswill, A.R.; Haste, N.M.; Monestier, M.; Nizet, V.; von Köckritz-Blickwede, M. Nuclease expression by Staphylococcus aureus facilitates escape from neutrophil extracellular traps. J. Innate Immun. 2010, 2, 576–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, E.E.; Rice, K.C.; Boles, B.R.; Endres, J.L.; Ranjit, D.; Chandramohan, L.; Tsang, L.H.; Smeltzer, M.S.; Horswill, A.R.; Bayles, K.W. Modulation of eDNA release and degradation affects Staphylococcus aureus biofilm maturation. PLoS ONE 2009, 4, e5822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiedrowski, M.R.; Kavanaugh, J.S.; Malone, C.L.; Mootz, J.M.; Voyich, J.M.; Smeltzer, M.S.; Bayles, K.W.; Horswill, A.R. Nuclease modulates biofilm formation in community-associated methicillin-resistant Staphylococcus aureus. PLoS ONE 2011, 6, e26714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoegh, S.V.; Skov, M.N.; Boye, K.; Worning, P.; Jensen, T.G.; Kemp, M. Variations in the Staphylococcus aureus-specific nuc gene can potentially lead to misidentification of methicillin-susceptible and-resistant S. aureus. J. Med. Microbiol. 2014, 63, 1020–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, X.; Liu, W.; Huang, W.; Fu, Q.; Li, M. Molecular characteristic and virulence gene profiles of community-associated methicillin-resistant Staphylococcus aureus isolates from pediatric patients in Shanghai, China. Front. Microbiol. 2016, 7, 1818. [Google Scholar] [CrossRef] [Green Version]
- Zarizal, S.; Yeo, C.C.; Faizal, G.M.; Hoong, C.; Chew, Z.A.Z.; Al-Obaidi, M.M.J.; Amin, N.S.; Nasir, M.D.M. Nasal colonisation, antimicrobial susceptibility and genotypic pattern of Staphylococcus aureus among agricultural biotechnology students in Besut, Terengganu, east coast of Malaysia. Trop. Med. Int. Health 2018, 23, 905–913. [Google Scholar] [CrossRef] [Green Version]

| S. aureus | Healthy (n = 77) | Diseased (n = 33) | OR | p-Value | 95% CI |
|---|---|---|---|---|---|
| 0.309 | 0.009 | 0.125–0.763 | |||
| Positive | 23 (29.9%) | 19 (57.6%) | |||
| Negative | 54 (70.1%) | 14 (42.4%) |
| Risk Factor | S. aureus | OR | p-Value | 95% CI | |
|---|---|---|---|---|---|
| Negative | Positive | ||||
| Age | 3.368 | 0.003 | 1.412–7.960 | ||
| <5 | 37 (54.4%) | 11(26.2%) | |||
| 5–10 | 31 (45.6%) | 31 (73.8%) | |||
| Breed | 7.053 | 0.06 | 0.07–0.6 | ||
| Arabian | 67 (98.5%) | 38 (90.5%) | |||
| Others | 1 (1.5%) | 4 (9.5%) | |||
| Sex | 2.869 | 0.05 | 1.0–1.798 | ||
| Male | 19 (27.9%) | 5 (11.9%) | |||
| Female | 49 (72.1%) | 37 (88.1%) | |||
| Season | 2.983 | 0.030 | 1.017–8.234 | ||
| Winter | 61 (89.7%) | 32 (76.2%) | |||
| Spring | 7 (10.3%) | 10 (23.8%) | |||
| Antimicrobial Sensitivity for Staphylococcus aureus | Antimicrobial Agent | ||
|---|---|---|---|
| Sensitive | Intermediate | Resistant | |
| 24 (100%) | 0 (0%) | 0 (0%) | Sulfamethoxazole/trimethoprim (25 μg) |
| 2 (8.33%) | 0 (0%) | 22 (91. 67%) | Oxacillin (1 μg) |
| 18 (75%) | 0 (0%) | 6 (25%) | Tetracycline (30 μg) |
| 6 (25%) | 1 (4.16%) | 17 (70.83%) | Rifamycin (5 μg) |
| 17 (70.83%) | 2 (8.33%) | 5 (20.83%) | Ampicillin (10 μg) |
| 8 (33.33%) | 5 (20.83%) | 11 (45.83%) | Gentamycin (10 μg) |
| 1 (4.16%) | 1 (4.16%) | 22 (91.67%) | Cephalosporin (10 μg) |
| 18 (75%) | 1 (4.16%) | 5 (20.83%) | Chloramphenicol (30 μg) |
| Case No. | Sample (Eye) | Equine Source | Age (in Years) | Clinical Signs | spa (x Region) | nuc | mecA | hlg |
|---|---|---|---|---|---|---|---|---|
| 1 | 1(Right) | Arabian | 0.5 | corneal edema, eyelid edema, keratitis | − | + | + | + |
| 2 | 2 (Left) | Arabian | 6 | mucopurulent discharge, corneal edema, eyelid edema, keratitis | − | + | + | + |
| 3 | 3 (Right) | Arabian | 8 | conjunctivitis, watery lacrimation | − | + | + | − |
| 4 | 4 Right) | Arabian | 1.5 | conjunctivitis | − | + | − | − |
| 5 | 5 (Left) | Draft | 1 | mucopurulent discharge, corneal edema, eyelid edema, keratitis e | + | + | + | + |
| 5 | 6 (Right) | Draft | 1 | mucopurulent discharge | − | + | + | + |
| 6 | 7 (Left) | Arabian | 2.5 | corneal opacity | + | + | − | + |
| 7 | 8 (Left) | Draft | 4 | conjunctivitis, watery lacrimation | + | + | + | − |
| 7 | 9 (Right) | Draft | 4 | conjunctivitis, watery lacrimation | + | + | + | − |
| 8 | 10 (Left) | Donkey | 5 | eyelid edema, mucopurulent discharge | + | + | − | + |
| 9 | 11 (Right) | Arabian | 9 | conjunctivitis, watery lacrimation | + | + | + | − |
| 9 | 12 (Left) | Arabian | 9 | conjunctivitis, watery lacrimation | + | + | + | − |
| 10 | 13 (Left) | Arabian | 5 | conjunctivitis, watery lacrimation | + | + | + | − |
| 10 | 14 (Right) | Arabian | 5 | conjunctivitis, watery lacrimation | + | + | + | − |
| 11 | 15 (Left) | Arabian | 11 | corneal opacity | + | + | + | + |
| 12 | 16 (Right) | Arabian | 14 | conjunctivitis, watery lacrimation | + | + | + | − |
| 13 | 17 (Right) | Donkey | 13 | eyelid edema, keratitis, mucopurulent discharge | + | + | + | + |
| 14 | 18 (Left) | Arabian | 6 | corneal edema, eyelid edema, keratitis | + | + | + | + |
| 15 | 19 (Right) | Donkey | 9 | eyelid edema, keratitis, mucopurulent discharge | + | + | + | + |
| 16 | 20 (Left) | Arabian | 12 | lacrimation, conjunctivitis | − | + | + | − |
| 17 | 21 (Left) | Arabian | 7 | corneal edema, eyelid edema, keratitis, mucopurulent discharge | − | + | + | + |
| 18 | 22 (Left) | Donkey | 7 | eyelid edema, keratitis, mucopurulent discharge | + | + | + | + |
| 19 | 23 (Left) | Arabian | 9 | eyelid edema, keratitis, mucopurulent discharge | − | + | + | + |
| 19 | 24 (Right) | Arabian | 9 | eyelid edema, keratitis, mucopurulent discharge | − | + | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tahoun, A.; Elnafarawy, H.K.; El-Sharkawy, H.; Rizk, A.M.; Alorabi, M.; El-Shehawi, A.M.; Youssef, M.A.; Ibrahim, H.M.M.; El-Khodery, S. The Prevalence and Molecular Biology of Staphylococcus aureus Isolated from Healthy and Diseased Equine Eyes in Egypt. Antibiotics 2022, 11, 221. https://doi.org/10.3390/antibiotics11020221
Tahoun A, Elnafarawy HK, El-Sharkawy H, Rizk AM, Alorabi M, El-Shehawi AM, Youssef MA, Ibrahim HMM, El-Khodery S. The Prevalence and Molecular Biology of Staphylococcus aureus Isolated from Healthy and Diseased Equine Eyes in Egypt. Antibiotics. 2022; 11(2):221. https://doi.org/10.3390/antibiotics11020221
Chicago/Turabian StyleTahoun, Amin, Helmy K. Elnafarawy, Hanem El-Sharkawy, Amira M. Rizk, Mohammed Alorabi, Ahmed M. El-Shehawi, Mohamed A. Youssef, Hussam M. M. Ibrahim, and Sabry El-Khodery. 2022. "The Prevalence and Molecular Biology of Staphylococcus aureus Isolated from Healthy and Diseased Equine Eyes in Egypt" Antibiotics 11, no. 2: 221. https://doi.org/10.3390/antibiotics11020221