
Brazilian Clinical Strains of Actinobacillus pleuropneumoniae and Pasteurella multocida: Capsular Diversity, Antimicrobial Susceptibility (In Vitro) and Proof of Concept for Prevention of Natural Colonization by Multi-Doses Protocol of Tildipirosin
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
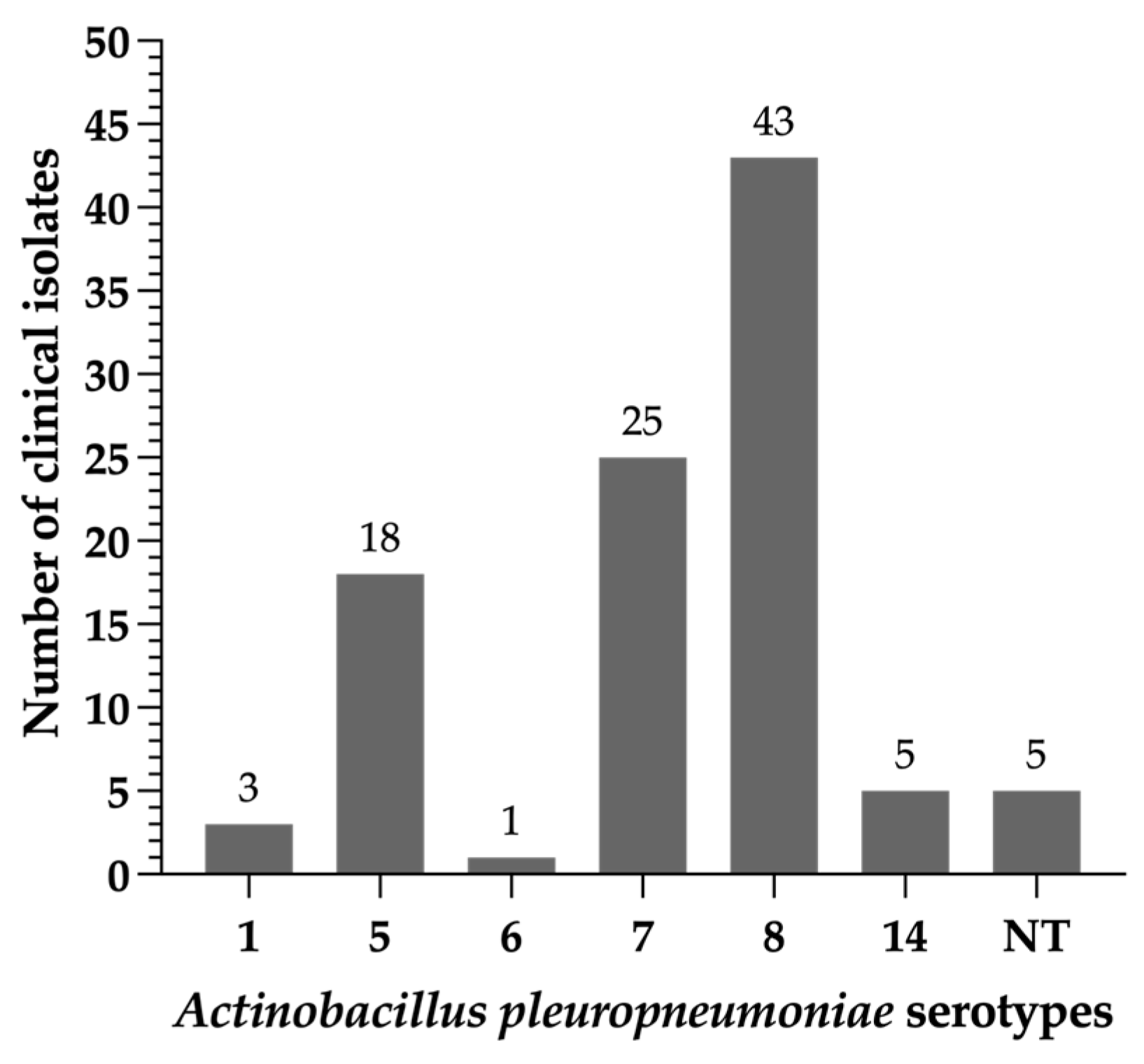
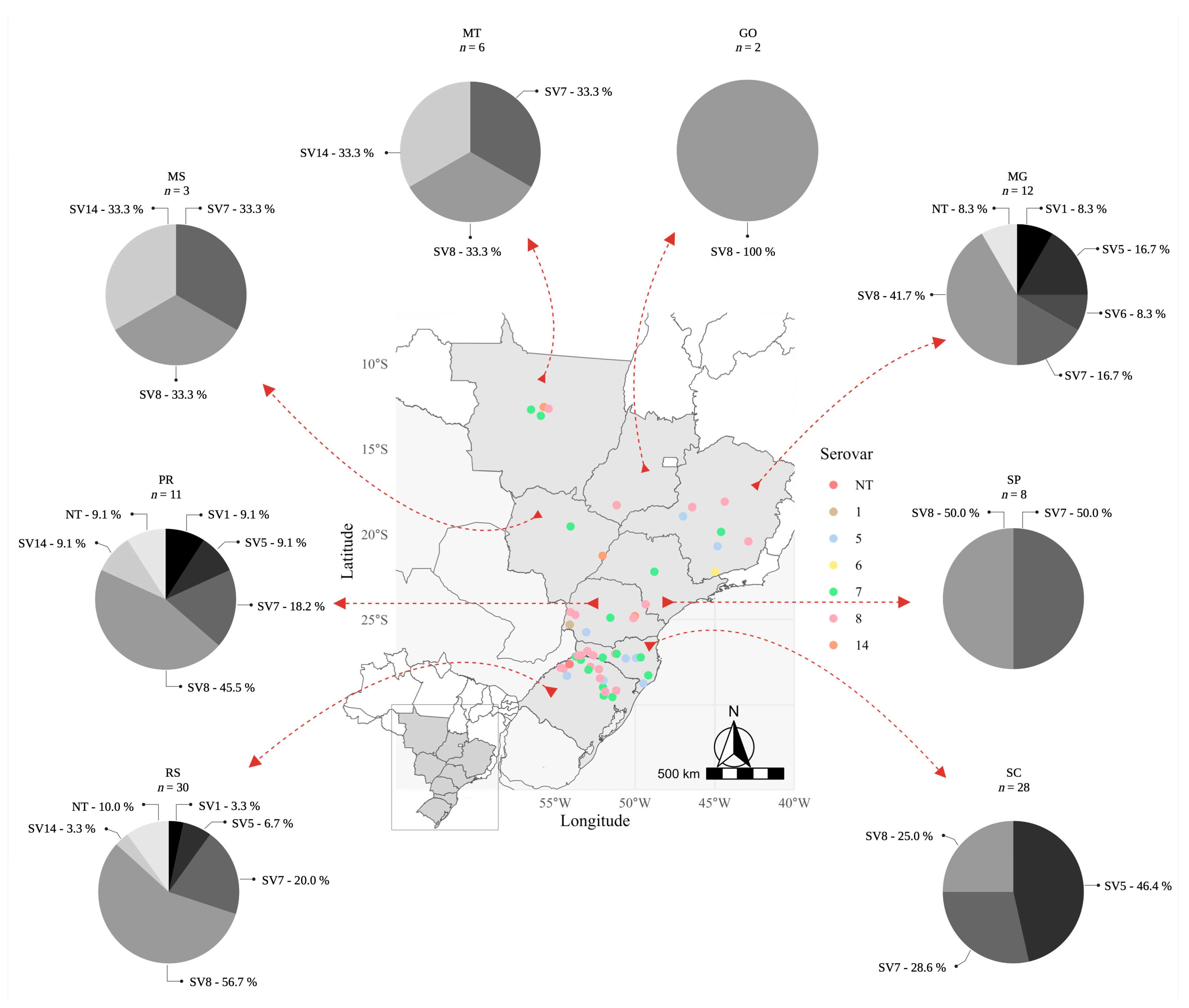
2.1. Typing of Actinobacillus pleuropneumoniae Clinical Isolates
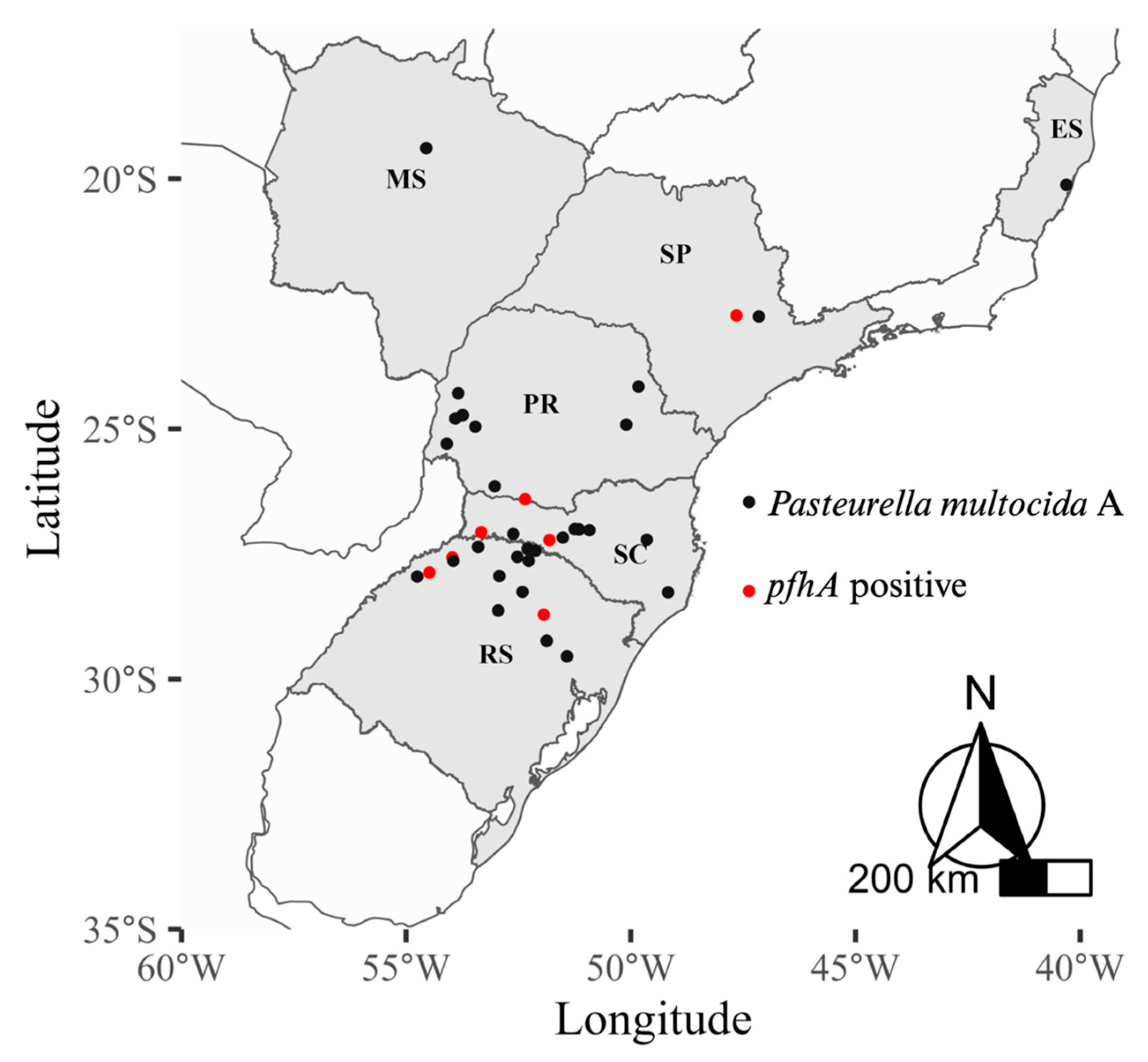
2.2. Determination of Virulence-Associated Genes in Pasteurella multocida Isolates
2.3. Antimicrobial Susceptibility Testing in Actinobacillus pleuropneumoniae and Pasteurella multocida Isolates to Tildipirosin
2.4. In Silico Association Analysis
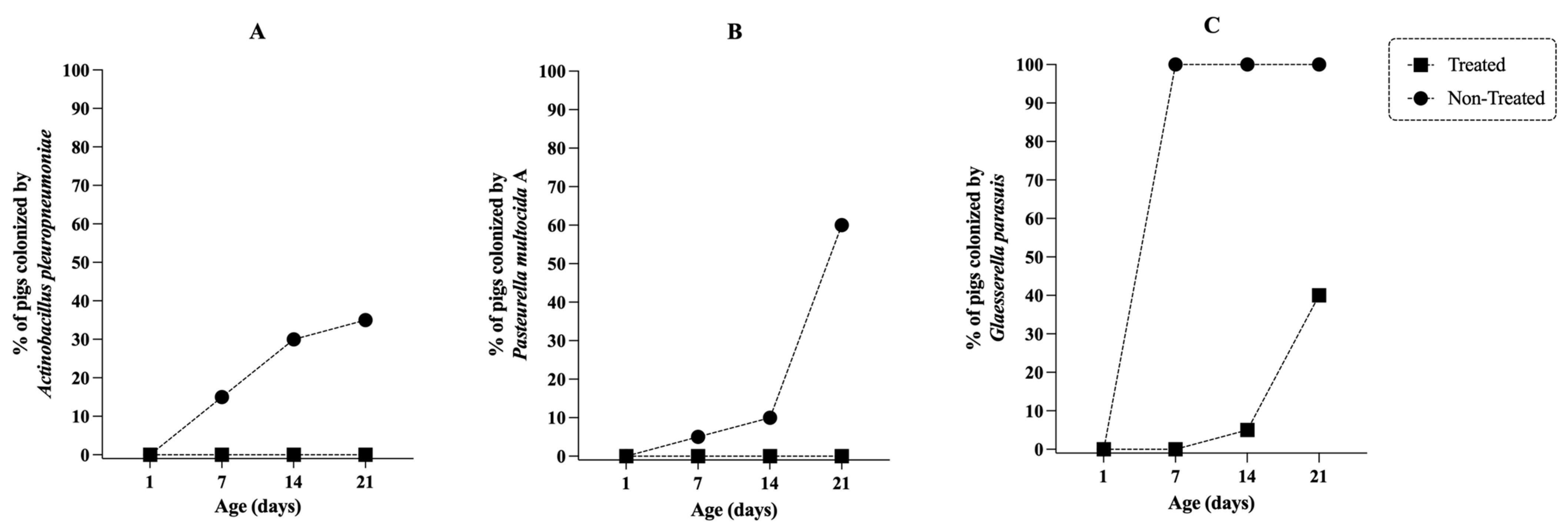
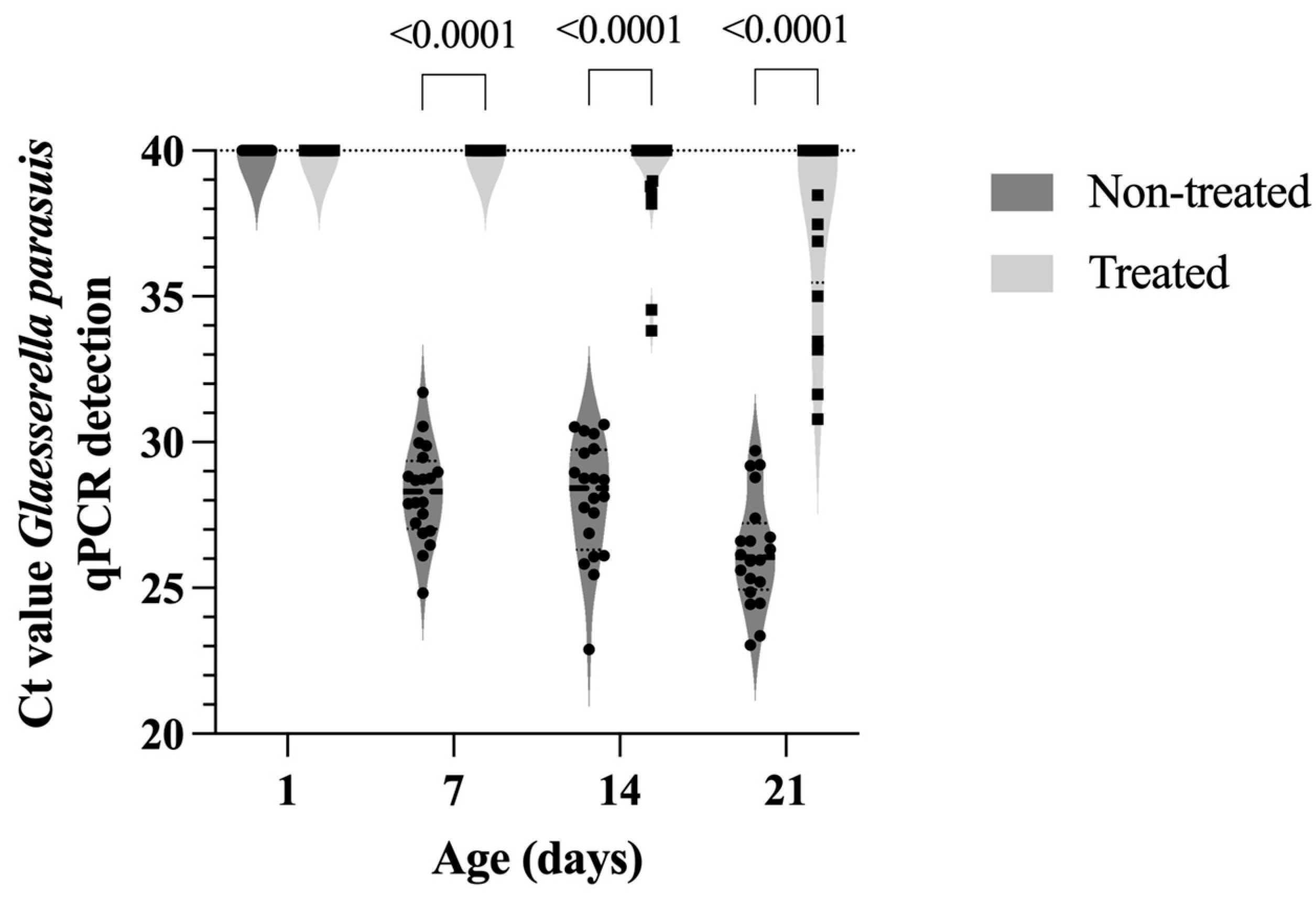
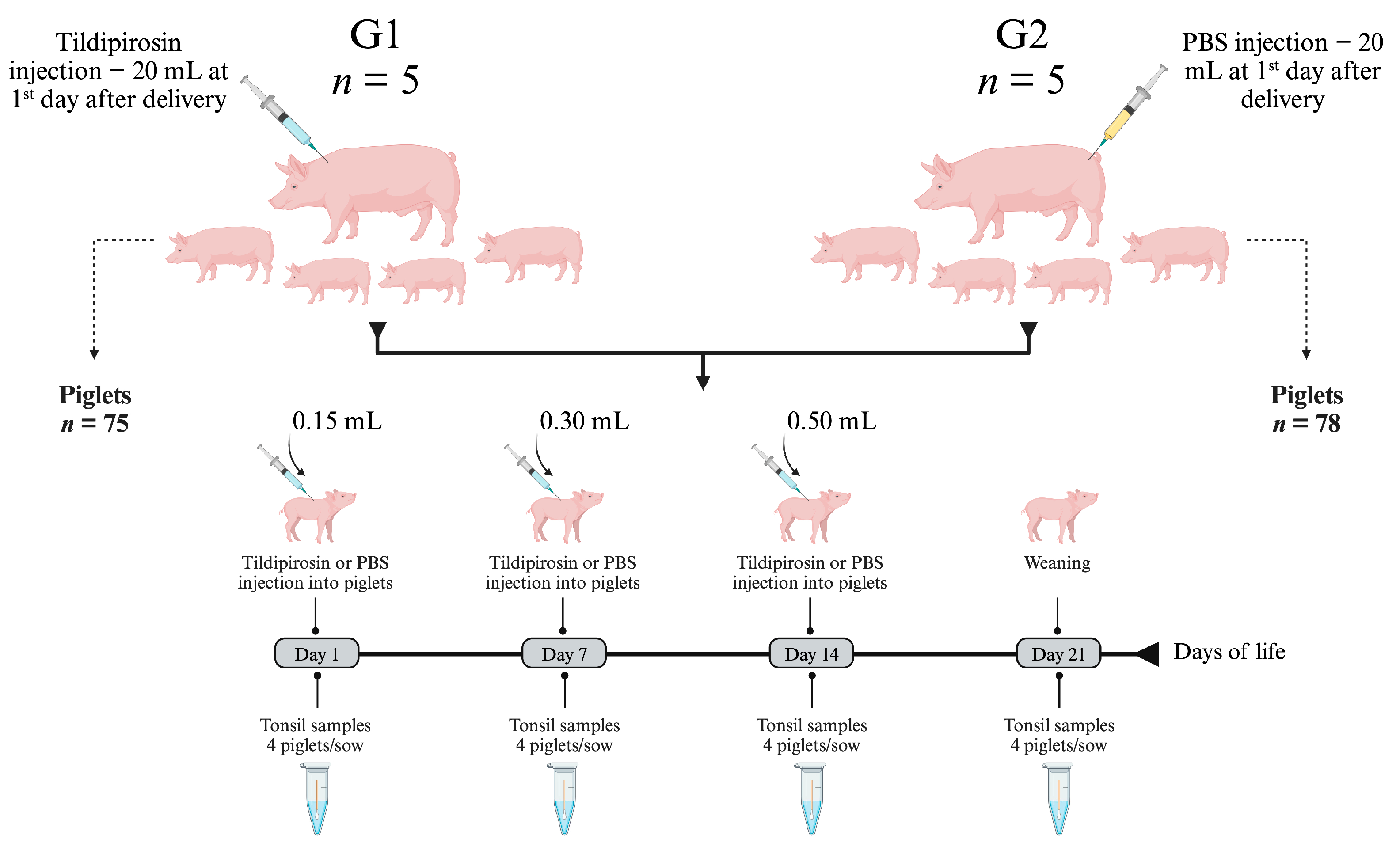
2.5. In Vivo Efficacy of Tildipirosin
3. Discussion
4. Material and Methods
4.1. Actinobacillus pleuropneumoniae Isolates and Molecular Capsule Typing
4.2. Pasteurella multocida Isolates, Capsular Typing, Molecular Confirmation, and Detection of Virulence-Associated Genes
4.3. Antimicrobial Sensitivity Testing
4.4. In Vivo Effect of Tildipirosin on Bacterial Colonization of Tonsils
4.5. Molecular Detection of Tonsillar Bacteria
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Losinger, W.C. Economic impacts of reduced pork production associated with the diagnosis of Actinobacillus pleuropneumoniae on grower/finisher swine operations in the United States. Prev. Vet. Med. 2005, 68, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Sassu, E.L.; Bosse, J.T.; Tobias, T.J.; Gottschalk, M.; Langford, P.R.; Hennig-Pauka, I. Update on Actinobacillus pleuropneumoniae-knowledge, gaps and challenges. Transbound. Emerg. Dis. 2018, 65 (Suppl. 1), 72–90. [Google Scholar] [CrossRef] [PubMed]
- Monger, X.C.; Gilbert, A.A.; Saucier, L.; Vincent, A.T. Antibiotic Resistance: From Pig to Meat. Antibiotics 2021, 10, 1209. [Google Scholar] [CrossRef] [PubMed]
- Stringer, O.W.; Bosse, J.T.; Lacouture, S.; Gottschalk, M.; Fodor, L.; Angen, O.; Velazquez, E.; Penny, P.; Lei, L.; Langford, P.R.; et al. Proposal of Actinobacillus pleuropneumoniae serovar 19, and reformulation of previous multiplex PCRs for capsule-specific typing of all known serovars. Vet. Microbiol. 2021, 255, 109021. [Google Scholar] [CrossRef] [PubMed]
- Carr, J.; Cehn, S.P.; Conner, J.F.; Kirkwood, R.N.; Segalés, J. Respiratory disorders. In Pig health; Carr, J., Cehn, S.P., Conner, J.F., Kirkwood, R.N., Segalés, J., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2018; pp. 103–152. [Google Scholar]
- Opriessnig, T.; Gimenez-Lirola, L.G.; Halbur, P.G. Polymicrobial respiratory disease in pigs. Anim. Health Res. Rev. 2011, 12, 133–148. [Google Scholar] [CrossRef]
- Oliveira Filho, J.X.d.; Morés, M.A.Z.; Rebelatto, R.; Agnol, A.M.D.; Plieski, C.L.A.; Klein, C.S.; Barcellos, D.E.S.N.; Morés, N. Pasteurella multocida type A as the primary agent of pneumonia and septicaemia in pigs. Pesqui. Veterinária Bras. 2015, 35, 716–724. [Google Scholar] [CrossRef]
- Tang, X.; Zhao, Z.; Hu, J.; Wu, B.; Cai, X.; He, Q.; Chen, H. Isolation, antimicrobial resistance, and virulence genes of Pasteurella multocida strains from swine in China. J. Clin. Microbiol. 2009, 47, 951–958. [Google Scholar] [CrossRef]
- Ewers, C.; Lubke-Becker, A.; Bethe, A.; Kiebling, S.; Filter, M.; Wieler, L.H. Virulence genotype of Pasteurella multocida strains isolated from different hosts with various disease status. Vet. Microbiol. 2006, 114, 304–317. [Google Scholar] [CrossRef]
- Rocha, N.; Mores, M.A.Z.; Dezen, D.; Mores, N.; Coldebella, A.; Rebelatto, R.; Kich, D.J. Economic impact of chronic pleural lesions and consequent disqualification of carcasses for export during inspection in swine slaughterhouses. Braz. J. Vet. Res. 2022, 42, e07118. [Google Scholar] [CrossRef]
- Dutra, M.C.; Moreno, L.Z.; Dias, R.A.; Moreno, A.M. Antimicrobial Use in Brazilian Swine Herds: Assessment of Use and Reduction Examples. Microorganisms 2021, 9, 881. [Google Scholar] [CrossRef]
- Tang, K.L.; Caffrey, N.P.; Nobrega, D.B.; Cork, S.C.; Ronksley, P.E.; Barkema, H.W.; Polachek, A.J.; Ganshorn, H.; Sharma, N.; Kellner, J.D.; et al. Restricting the use of antibiotics in food-producing animals and its associations with antibiotic resistance in food-producing animals and human beings: A systematic review and meta-analysis. Lancet Planet. Health 2017, 1, e316–e327. [Google Scholar] [CrossRef] [PubMed]
- Schlunzen, F.; Zarivach, R.; Harms, J.; Bashan, A.; Tocilj, A.; Albrecht, R.; Yonath, A.; Franceschi, F. Structural basis for the interaction of antibiotics with the peptidyl transferase centre in eubacteria. Nature 2001, 413, 814–821. [Google Scholar] [CrossRef]
- Andersen, N.M.; Poehlsgaard, J.; Warrass, R.; Douthwaite, S. Inhibition of protein synthesis on the ribosome by tildipirosin compared with other veterinary macrolides. Antimicrob. Agents Chemother. 2012, 56, 6033–6036. [Google Scholar] [CrossRef] [PubMed]
- Peres, P.R.; Prigol, S.R.; Martín, C.B.G.; Feronatod, C.; Suriñach, M.C.; Kreutz, L.C.; Frandoloso, R. Tildipirosin: An effective antibiotic against Glaesserella parasuis from an in vitro analysis. Vet. Anim. Sci. 2020, 10, 100136. [Google Scholar] [CrossRef] [PubMed]
- Vilaro, A.; Novell, E.; Enrique-Tarancon, V.; Balielles, J.; Vilalta, C.; Martinez, S.; Fraile Sauce, L.J. Antimicrobial Susceptibility Pattern of Porcine Respiratory Bacteria in Spain. Antibiotics 2020, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.; Liu, Q.; Qi, Y.; Yang, B.; Khaliq, H.; Xiong, J.; Moku, G.K.; Ahmed, S.; Li, K.; Zhang, H.; et al. Optimal Regimens and Cutoff Evaluation of Tildipirosin Against Pasteurella multocida. Front. Pharmacol. 2018, 9, 765. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, C.; Jones, S.C.; Bosse, J.T.; Watson, C.M.; Williamson, S.M.; Rycroft, A.N.; Kroll, J.S.; Hartley, H.M.; Langford, P.R. Prevalence of Actinobacillus pleuropneumoniae serovars in England and Wales. Vet. Rec. 2010, 167, 661–662. [Google Scholar] [CrossRef]
- Li, Y.; Bosse, J.T.; Williamson, S.M.; Maskell, D.J.; Tucker, A.W.; Wren, B.W.; Rycroft, A.N.; Langford, P.R.; Consortium, B.T. Actinobacillus pleuropneumoniae serovar 8 predominates in England and Wales. Vet. Rec. 2016, 179, 276. [Google Scholar] [CrossRef]
- Sarkozi, R.; Makrai, L.; Fodor, L. Actinobacillus pleuropneumoniae serotypes in Hungary. Acta Vet. Hung. 2018, 66, 343–349. [Google Scholar] [CrossRef]
- Lacouture, S.; Gottschalk, M. Distribution of Actinobacillus pleuropneumoniae (from 2015 to June 2020) and Glaesserella parasuis (from 2017 to June 2020) serotypes isolated from diseased pigs in Quebec. Can. Vet. J. 2020, 61, 1261–1263. [Google Scholar]
- Gottschalk, M.; Lacouture, S. Canada: Distribution of Streptococcus suis (from 2012 to 2014) and Actinobacillus pleuropneumoniae (from 2011 to 2014) serotypes isolated from diseased pigs. Can. Vet. J. 2015, 56, 1093–1094. [Google Scholar] [PubMed]
- Kim, B.; Hur, J.; Lee, J.Y.; Choi, Y.; Lee, J.H. Molecular serotyping and antimicrobial resistance profiles of Actinobacillus pleuropneumoniae isolated from pigs in South Korea. Vet. Q. 2016, 36, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Min, K.; Chae, C. Serotype and apx genotype profiles of Actinobacillus pleuropneumoniae field isolates in Korea. Vet. Rec. 1999, 145, 251–254. [Google Scholar] [CrossRef] [PubMed]
- McGregor, G.F.; Gottschalk, M.; Godson, D.L.; Wilkins, W.; Bollinger, T.K. Disease risks associated with free-ranging wild boar in Saskatchewan. Can. Vet. J. 2015, 56, 839–844. [Google Scholar] [PubMed]
- Blackall, P.J.; Klaasen, H.L.; van den Bosch, H.; Kuhnert, P.; Frey, J. Proposal of a new serovar of Actinobacillus pleuropneumoniae: Serovar 15. Vet. Microbiol. 2002, 84, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Reimer, D.; Frey, J.; Jansen, R.; Veit, H.P.; Inzana, T.J. Molecular investigation of the role of ApxI and ApxII in the virulence of Actinobacillus pleuropneumoniae serotype 5. Microb. Pathog. 1995, 18, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.; Kim, B.; Cho, W.S.; Kim, J.; Kwon, D.; Cheon, D.S.; Chae, C. Capsular serotype, toxA gene, and antimicrobial susceptibility profiles of Pasteurella multocida isolated from pigs with pneumonia in Korea. Vet. Rec. 2001, 149, 210–212. [Google Scholar] [CrossRef]
- Bethe, A.; Wieler, L.H.; Selbitz, H.J.; Ewers, C. Genetic diversity of porcine Pasteurella multocida strains from the respiratory tract of healthy and diseased swine. Vet. Microbiol. 2009, 139, 97–105. [Google Scholar] [CrossRef]
- Garcia, N.; Fernandez-Garayzabal, J.F.; Goyache, J.; Dominguez, L.; Vela, A.I. Associations between biovar and virulence factor genes in Pasteurella multocida isolates from pigs in Spain. Vet. Rec. 2011, 169, 362. [Google Scholar] [CrossRef]
- Sarangi, L.N.; Priyadarshini, A.; Kumar, S.; Thomas, P.; Gupta, S.K.; Nagaleekar, V.K.; Singh, V.P. Virulence genotyping of Pasteurella multocida isolated from multiple hosts from India. Sci. World J. 2014, 2014, 814109. [Google Scholar] [CrossRef]
- Oliveira Filho, J.X.; Mores, M.A.Z.; Rebellato, R.; Kich, J.D.; Cantao, M.E.; Klein, C.S.; Guedes, R.M.C.; Coldebella, A.; Barcellos, D.; Mores, N. Pathogenic variability among Pasteurella multocida type A isolates from Brazilian pig farms. BMC Vet. Res. 2018, 14, 244. [Google Scholar] [CrossRef] [PubMed]
- ABPA. Annual Report. 2022. Available online: https://abpa-br.org/wp-content/uploads/2023/01/ABPA-Annual-Report-2022.pdf (accessed on 3 September 2023).
- Hillerton, J.E.; Irvine, C.R.; Bryan, M.A.; Scott, D.; Merchant, S.C. Use of antimicrobials for animals in New Zealand, and in comparison with other countries. N. Z. Vet. J. 2017, 65, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.; Liu, Q.; Yang, B.; Ahmed, S.; Cao, J.; He, Q. The pharmacokinetic-pharmacodynamic modeling and cut-off values of tildipirosin against Haemophilus parasuis. Oncotarget 2018, 9, 1673–1690. [Google Scholar] [CrossRef] [PubMed]
- Poehlsgaard, J.; Andersen, N.M.; Warrass, R.; Douthwaite, S. Visualizing the 16-membered ring macrolides tildipirosin and tilmicosin bound to their ribosomal site. ACS Chem. Biol. 2012, 7, 1351–1355. [Google Scholar] [CrossRef] [PubMed]
- Michael, G.B.; Eidam, C.; Kadlec, K.; Meyer, K.; Sweeney, M.T.; Murray, R.W.; Watts, J.L.; Schwarz, S. Increased MICs of gamithromycin and tildipirosin in the presence of the genes erm(42) and msr(E)-mph(E) for bovine Pasteurella multocida and Mannheimia haemolytica. J. Antimicrob. Chemother. 2012, 67, 1555–1557. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, T.S.; Felizardo, M.R.; Sena de Gobbi, D.D.; Gomes, C.R.; Nogueira Filsner, P.H.; Moreno, M.; Paixao, R.; Pereira Jde, J.; Micke Moreno, A. Virulence genes and antimicrobial resistance profiles of Pasteurella multocida strains isolated from rabbits in Brazil. Sci. World J. 2012, 2012, 685028. [Google Scholar] [CrossRef]
- Dellagostin, D.; Klein, R.L.; Giacobbo, I.; Guizzo, J.A.; Dazzi, C.C.; Prigol, S.R.; Martin, C.B.G.; Kreutz, L.C.; Schryvers, A.B.; Frandoloso, R. TbpB(Y)(167A)-based vaccine is safe in pregnant sows and induces high titers of maternal derived antibodies that reduce Glaesserella parasuis colonization in piglets. Vet. Microbiol. 2023, 276, 109630. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.; Menge, M.; Bohland, C.; Zschiesche, E.; Wilhelm, C.; Kilp, S.; Metz, W.; Allan, M.; Ropke, R.; Nurnberger, M. Pharmacokinetics of tildipirosin in porcine plasma, lung tissue, and bronchial fluid and effects of test conditions on in vitro activity against reference strains and field isolates of Actinobacillus pleuropneumoniae. J. Vet. Pharmacol. Ther. 2013, 36, 140–153. [Google Scholar] [CrossRef]
- Torres, F.; Santamaria, R.; Jimenez, M.; Menjon, R.; Ibanez, A.; Collell, M.; Azlor, O.; Fraile, L. Pharmacokinetics of tildipirosin in pig tonsils. J. Vet. Pharmacol. Ther. 2016, 39, 199–201. [Google Scholar] [CrossRef]
- Townsend, K.M.; Boyce, J.D.; Chung, J.Y.; Frost, A.J.; Adler, B. Genetic organization of Pasteurella multocida cap Loci and development of a multiplex capsular PCR typing system. J. Clin. Microbiol. 2001, 39, 924–929. [Google Scholar] [CrossRef]
- Atashpaz, S.; Shayegh, J.; Hejazi, M.S. Rapid virulence typing of Pasteurella multocida by multiplex PCR. Res. Vet. Sci. 2009, 87, 355–357. [Google Scholar] [CrossRef]
- CLSI. CLSI Supplement M100. In Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; Wayne, P., Ed.; Clinical and Laboratory Standards Institute: Philadelphia, PN, USA, 2018. [Google Scholar]
- CLSI. CLSI Supplement VET08. In Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 4th ed.; Wayne, P., Ed.; Clinical and Laboratory Standards Institute: Philadelphia, PN, USA, 2018. [Google Scholar]
- Turni, C.; Pyke, M.; Blackall, P.J. Validation of a real-time PCR for Haemophilus parasuis. J. Appl. Microbiol. 2010, 108, 1323–1331. [Google Scholar] [CrossRef]
- Tocqueville, V.; Kempf, I.; Paboeuf, F.; Marois-Crehan, C. Quantification of Pasteurella multocida in experimentally infected pigs using a real-time PCR assay. Res. Vet. Sci. 2017, 112, 177–184. [Google Scholar] [CrossRef]
- Tobias, T.J.; Bouma, A.; Klinkenberg, D.; Daemen, A.J.; Stegeman, J.A.; Wagenaar, J.A.; Duim, B. Detection of Actinobacillus pleuropneumoniae in pigs by real-time quantitative PCR for the apxIVA gene. Vet. J. 2012, 193, 557–560. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tildipirosin Concentration and Clinical Parameter | Number and Accumulative% of Strains with MIC Equal to or Less than the Indicated Concentration | |
|---|---|---|
| App Isolates (n = 100) | PmA Isolates (n = 60) | |
| ≤0.06 μg/mL | 1 (1.0%) | - |
| 0.12 μg/mL | 17 (18.0%) | - |
| 0.25 μg/mL | 32 (50.0%) | - |
| 0.5 μg/mL | 27 (77.0%) | 9 (15.0%) |
| 1 μg/mL | 8 (85.0%) | 28 (61.7%) |
| 2 μg/mL | 4 (89.0%) | 4 (68.3%) |
| 4 μg/mL | 2 (91.0%) | 3 (73.3%) |
| 8 μg/mL | 3 (94.0%) | - |
| 16 μg/mL | 1 (95.0%) | - |
| 32 μg/mL | 1 (96.0%) | 1 (75.0%) |
| 64 μg/mL | - | - |
| >64 μg/mL | 4 (100.0%) | 15 (100.0%) |
| MIC range (μg/mL) | ≤0.06–>64 | 0.5–>64 |
| MIC50 (μg/mL) | 0.25 | 1 |
| MIC90 (μg/mL) | 4 | ≥64 |
| Clinical breakpoint (μg/mL) | ≤16 $ | ≤4 $ |
| Susceptibility rate | 95.0 | 73.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuchiishi, S.S.; Ramos Prigol, S.; Bresolin, E.; Fernandes Lenhard, B.; Pissetti, C.; García-Iglesias, M.-J.; Gutiérrez-Martín, C.-B.; Martínez-Martínez, S.; Kreutz, L.C.; Frandoloso, R. Brazilian Clinical Strains of Actinobacillus pleuropneumoniae and Pasteurella multocida: Capsular Diversity, Antimicrobial Susceptibility (In Vitro) and Proof of Concept for Prevention of Natural Colonization by Multi-Doses Protocol of Tildipirosin. Antibiotics 2023, 12, 1658. https://doi.org/10.3390/antibiotics12121658
Kuchiishi SS, Ramos Prigol S, Bresolin E, Fernandes Lenhard B, Pissetti C, García-Iglesias M-J, Gutiérrez-Martín C-B, Martínez-Martínez S, Kreutz LC, Frandoloso R. Brazilian Clinical Strains of Actinobacillus pleuropneumoniae and Pasteurella multocida: Capsular Diversity, Antimicrobial Susceptibility (In Vitro) and Proof of Concept for Prevention of Natural Colonization by Multi-Doses Protocol of Tildipirosin. Antibiotics. 2023; 12(12):1658. https://doi.org/10.3390/antibiotics12121658
Chicago/Turabian StyleKuchiishi, Suzana Satomi, Simone Ramos Prigol, Eduarda Bresolin, Bianca Fernandes Lenhard, Caroline Pissetti, María-José García-Iglesias, César-Bernardo Gutiérrez-Martín, Sonia Martínez-Martínez, Luiz Carlos Kreutz, and Rafael Frandoloso. 2023. "Brazilian Clinical Strains of Actinobacillus pleuropneumoniae and Pasteurella multocida: Capsular Diversity, Antimicrobial Susceptibility (In Vitro) and Proof of Concept for Prevention of Natural Colonization by Multi-Doses Protocol of Tildipirosin" Antibiotics 12, no. 12: 1658. https://doi.org/10.3390/antibiotics12121658