
Exploring the Accessory Genome of Multidrug-Resistant Rhodococcus equi Clone 2287

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatic Analysis
2.2. Bacterial Strains and Culture Conditions
2.3. Bacterial Conjugation
2.4. Plasmid Loss Assay
2.5. PCR
3. Results
3.1. The Search for Genetic Elements in MDR R. equi Clone 2287 Selected and Maintained over Time
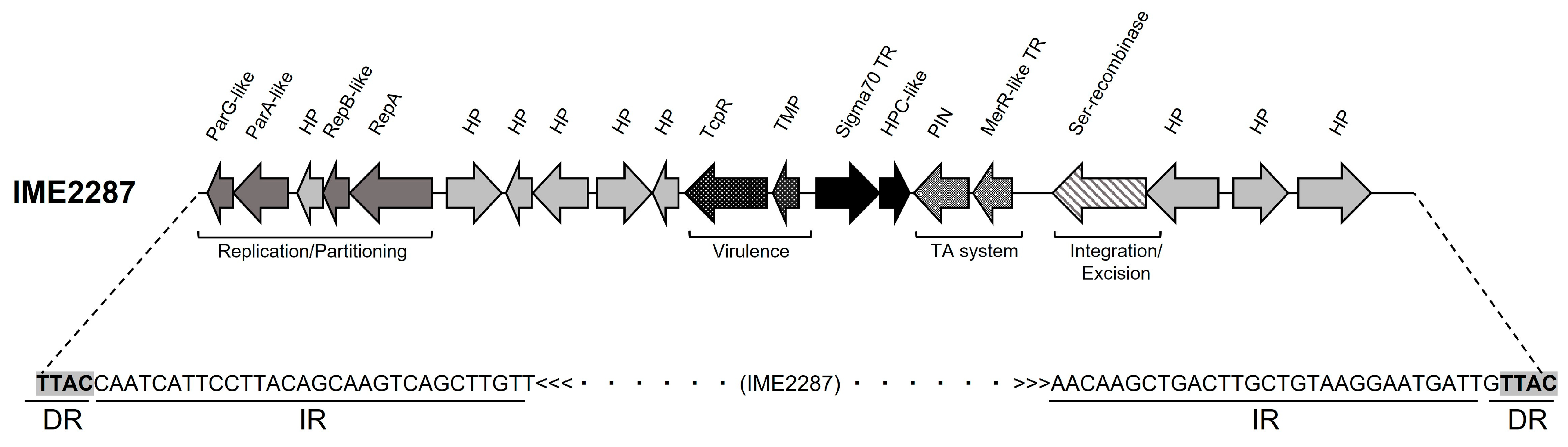
3.2. Molecular Characterization of IME2287, a Putative Mobilizable Element Associated with Macrolide Resistance in MDR R. equi
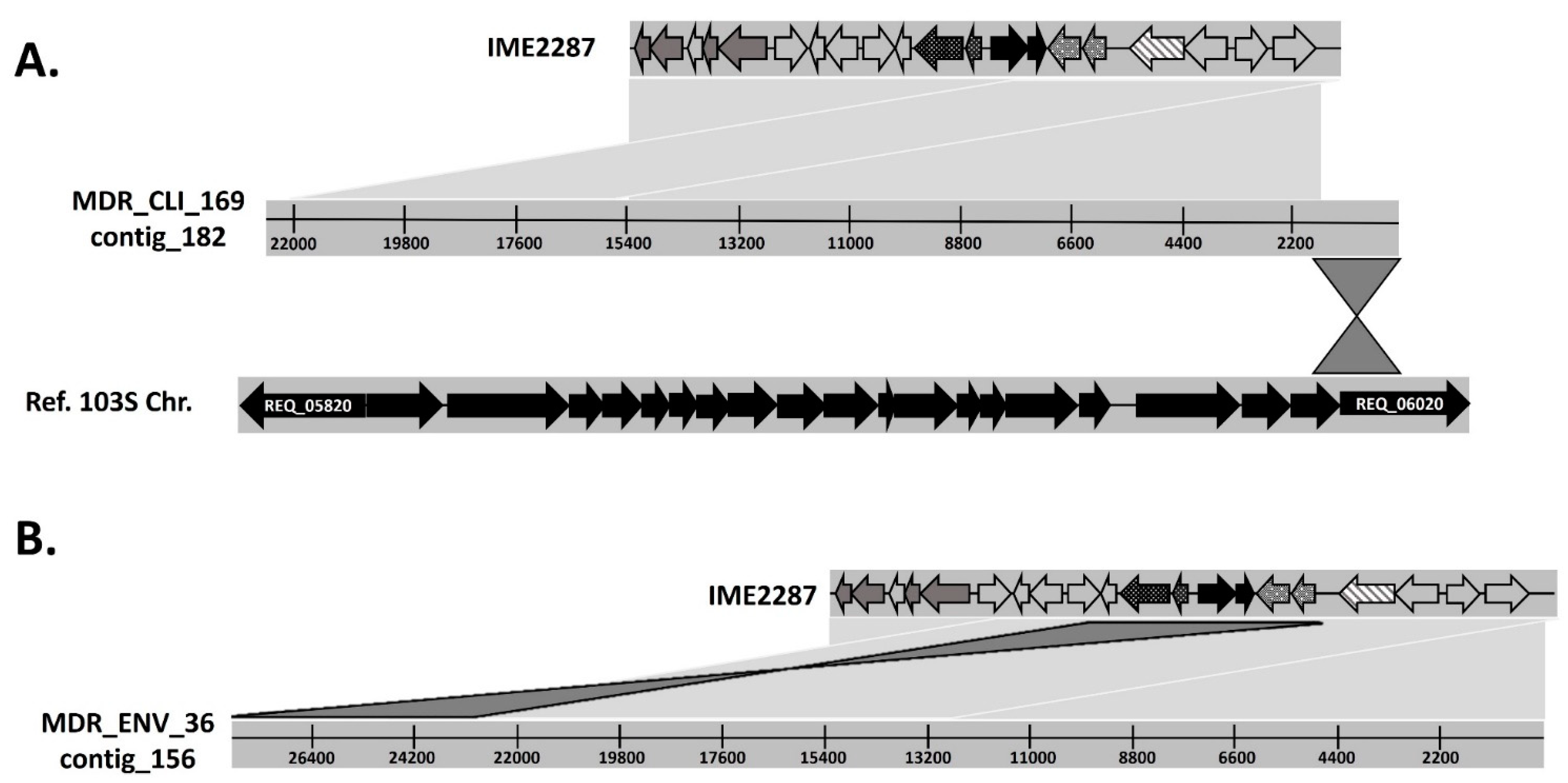
3.3. IME2287 Moved from MDR R. equi Clone 2287 to other R. equi Genetic Backgrounds Associated with Macrolide Resistance
3.4. IME2287 Mobilization Is Independent of Macrolide Resistance Plasmid pRErm46
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prescott, J.F. Rhodococcus equi: An animal and human pathogen. Clin. Microbiol. Rev. 1991, 4, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Bordin, A.I.; Huber, L.; Sanz, M.G.; Cohen, N.D. Rhodococcus equi foal pneumonia: Update on epidemiology, immunity, treatment and prevention. Equine Vet. J. 2022, 54, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Giguere, S.; Hondalus, M.K.; Yager, J.A.; Darrah, P.; Mosser, D.M.; Prescott, J.F. Role of the 85-kilobase plasmid and plasmid-encoded virulence-associated protein A in intracellular survival and virulence of Rhodococcus equi. Infect. Immun. 1999, 67, 3548–3557. [Google Scholar] [CrossRef] [PubMed]
- Valero-Rello, A.; Hapeshi, A.; Anastasi, E.; Alvarez, S.; Scortti, M.; Meijer, W.G.; MacArthur, I.; Vazquez-Boland, J.A. An Invertron-Like Linear Plasmid Mediates Intracellular Survival and Virulence in Bovine Isolates of Rhodococcus equi. Infect. Immun. 2015, 83, 2725–2737. [Google Scholar] [CrossRef] [PubMed]
- Ocampo-Sosa, A.A.; Lewis, D.A.; Navas, J.; Quigley, F.; Callejo, R.; Scortti, M.; Leadon, D.P.; Fogarty, U.; Vazquez-Boland, J.A. Molecular epidemiology of Rhodococcus equi based on traA, vapA, and vapB virulence plasmid markers. J. Infect. Dis. 2007, 196, 763–769. [Google Scholar] [CrossRef]
- Song, Y.; Xu, X.; Huang, Z.; Xiao, Y.; Yu, K.; Jiang, M.; Yin, S.; Zheng, M.; Meng, H.; Han, Y.; et al. Genomic Characteristics Revealed Plasmid-Mediated Pathogenicity and Ubiquitous Rifamycin Resistance of Rhodococcus equi. Front. Cell. Infect. Microbiol. 2022, 12, 807610. [Google Scholar]
- Chaffin, M.K.; Cohen, N.D.; Martens, R.J.; Edwards, R.F.; Nevill, M. Foal-related risk factors associated with development of Rhodococcus equi pneumonia on farms with endemic infection. J. Am. Vet. Med. Assoc. 2003, 223, 1791–1799. [Google Scholar] [CrossRef]
- Chaffin, M.K.; Cohen, N.D.; Martens, R.J.; Edwards, R.F.; Nevill, M.; Smith, R., 3rd. Hematologic and immunophenotypic factors associated with development of Rhodococcus equi pneumonia of foals at equine breeding farms with endemic infection. Vet. Immunol. Immunopathol. 2004, 100, 33–48. [Google Scholar] [CrossRef]
- Giguere, S.; Prescott, J.F. Clinical manifestations, diagnosis, treatment, and prevention of Rhodococcus equi infections in foals. Vet. Microbiol. 1997, 56, 313–334. [Google Scholar] [CrossRef]
- Vazquez-Boland, J.A.; Giguere, S.; Hapeshi, A.; MacArthur, I.; Anastasi, E.; Valero-Rello, A. Rhodococcus equi: The many facets of a pathogenic actinomycete. Vet. Microbiol. 2013, 167, 9–33. [Google Scholar] [CrossRef]
- Muscatello, G.; Gilkerson, J.R.; Browning, G.F. Detection of virulent Rhodococcus equi in exhaled air samples from naturally infected foals. J. Clin. Microbiol. 2009, 47, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Muscatello, G.; Leadon, D.P.; Klayt, M.; Ocampo-Sosa, A.; Lewis, D.A.; Fogarty, U.; Buckley, T.; Gilkerson, J.R.; Meijer, W.G.; Vazquez-Boland, J.A. Rhodococcus equi infection in foals: The science of ‘rattles’. Equine Vet. J. 2007, 39, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Takai, S.; Yoda, A.; Sasaki, Y.; Kakuda, T.; Suzuki, Y.; Oikawa, M. Birth month associated with tracheal colonization of Rhodococcus equi in newborn foals on horse-breeding farms with sporadic rhodococcosis in Japan. Vet. Microbiol. 2022, 267, 109373. [Google Scholar] [CrossRef] [PubMed]
- Giles, C.; Vanniasinkam, T.; Ndi, S.; Barton, M.D. Rhodococcus equi (Prescottella equi) vaccines; the future of vaccine development. Equine Vet. J. 2015, 47, 510–518. [Google Scholar] [CrossRef]
- Kahn, S.K.; Cohen, N.D.; Bordin, A.I.; Coleman, M.C.; Heird, J.C.; Welsh, T.H., Jr. Transfusion of hyperimmune plasma for protecting foals against Rhodococcus equi pneumonia. Equine Vet. J. 2022, 55, 376–388. [Google Scholar] [CrossRef]
- Perkins, G.A.; Yeager, A.; Erb, H.N.; Nydam, D.V.; Divers, T.J.; Bowman, J.L. Survival of foals with experimentally induced Rhodococcus equi infection given either hyperimmune plasma containing R. equi antibody or normal equine plasma. Vet. Ther. 2002, 3, 334–346. [Google Scholar]
- Higuchi, T.; Arakawa, T.; Hashikura, S.; Inui, T.; Senba, H.; Takai, S. Effect of prophylactic administration of hyperimmune plasma to prevent Rhodococcus equi infection on foals from endemically affected farms. Zentralbl Vet. B 1999, 46, 641–648. [Google Scholar] [CrossRef]
- Cohen, N. Control and prevention of Rhodococcus equi foal pneumonia. AAEP Proc. 2010, 56, 129–131. [Google Scholar]
- Giguere, S. Treatment of Infections Caused by Rhodococcus equi. Vet. Clin. N. Am. Equine Pract. 2017, 33, 67–85. [Google Scholar] [CrossRef]
- Erol, E.; Scortti, M.; Fortner, J.; Patel, M.; Vazquez-Boland, J.A. Antimicrobial Resistance Spectrum Conferred by pRErm46 of Emerging Macrolide (Multidrug)-Resistant Rhodococcus equi. J. Clin. Microbiol. 2021, 59, e0114921. [Google Scholar] [CrossRef]
- Hillidge, C.J. Use of erythromycin-rifampin combination in treatment of Rhodococcus equi pneumonia. Vet. Microbiol. 1987, 14, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, C.R.; Sweeney, R.W.; Divers, T.J. Rhodococcus equi pneumonia in 48 foals: Response to antimicrobial therapy. Vet. Microbiol. 1987, 14, 329–336. [Google Scholar] [CrossRef]
- Erol, E.; Shaffer, C.L.; Lubbers, B.V. Synergistic combinations of clarithromycin with doxycycline or minocycline reduce the emergence of antimicrobial resistance in Rhodococcus equi. Equine Vet. J. 2021, 54, 799–806. [Google Scholar] [CrossRef]
- Giguere, S.; Lee, E.; Williams, E.; Cohen, N.D.; Chaffin, M.K.; Halbert, N.; Martens, R.J.; Franklin, R.P.; Clark, C.C.; Slovis, N.M. Determination of the prevalence of antimicrobial resistance to macrolide antimicrobials or rifampin in Rhodococcus equi isolates and treatment outcome in foals infected with antimicrobial-resistant isolates of R. equi. J. Am. Vet. Med. Assoc. 2010, 237, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Huber, L.; Giguere, S.; Slovis, N.M.; Carter, C.N.; Barr, B.S.; Cohen, N.D.; Elam, J.; Erol, E.; Locke, S.J.; Phillips, E.D.; et al. Emergence of resistance to macrolides and rifampicin in clinical isolates of Rhodococcus equi from foals in central Kentucky, USA: 1995 to 2017. Antimicrob. Agents Chemother. 2018, 63, e01714-18. [Google Scholar] [PubMed]
- Alvarez-Narvaez, S.; Huber, L.; Giguere, S.; Hart, K.A.; Berghaus, R.D.; Sanchez, S.; Cohen, N.D. Epidemiology and Molecular Basis of Multidrug Resistance in Rhodococcus equi. Microbiol. Mol. Biol. Rev. 2021, 85, e00011-21. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Y.; Yan, J.; Wang, C.; He, H. Appearance of multidrug-resistant virulent Rhodococcus equi clinical isolates obtained in China. J. Clin. Microbiol. 2014, 52, 703. [Google Scholar] [CrossRef]
- Buckley, T.; McManamon, E.; Stanbridge, S. Resistance studies of erythromycin and rifampin for Rhodococcus equi over a 10-year period. Ir. Vet. J. 2007, 60, 728–731. [Google Scholar] [CrossRef]
- Val-Calvo, J.; Darcy, J.; Gibbons, J.; Creighton, A.; Egan, C.; Buckley, T.; Schmalenberger, A.; Fogarty, U.; Scortti, M.; Vazquez-Boland, J.A. International Spread of Multidrug-Resistant Rhodococcus equi. Emerg. Infect. Dis. 2022, 28, 1899–1903. [Google Scholar] [CrossRef]
- Duchesne, R.; Castagnet, S.; Maillard, K.; Petry, S.; Cattoir, V.; Giard, J.C.; Leon, A. In vitro antimicrobial susceptibility of equine clinical isolates from France, 2006–2016. J. Glob. Antimicrob. Resist. 2019, 19, 144–153. [Google Scholar] [CrossRef]
- Kalinowski, M.; Jarosz, L.; Gradzki, Z. Assessment of Antimicrobial Susceptibility of Virulent Strains of Rhodococcus equi Isolated From Foals and Soil of Horse Breeding Farms With and Without Endemic Infections. J. Equine Vet. Sci. 2020, 91, 103114. [Google Scholar] [CrossRef]
- Alvarez-Narvaez, S.; Giguère, S.; Cohen, N.; Slovis, N.M.; Vazquez-Boland, J. Spread of emerging multidrug-resistant Rhodococcus equi in USA. Emerg. Infect. Dis. 2021, 27, 529. [Google Scholar] [CrossRef] [PubMed]
- Huber, L.; Giguere, S.; Slovis, N.M.; Alvarez-Narvaez, S.; Hart, K.A.; Greiter, M.; Morris, E.R.A.; Cohen, N.D. The novel and transferable erm(51) gene confers macrolides, lincosamides and streptogramins B (MLSB) resistance to clonal Rhodococcus equi in the environment. Environ. Microbiol. 2020, 22, 2858–2869. [Google Scholar] [CrossRef]
- Anastasi, E.; Giguere, S.; Berghaus, L.J.; Hondalus, M.K.; Willingham-Lane, J.M.; MacArthur, I.; Cohen, N.D.; Roberts, M.C.; Vazquez-Boland, J.A. Novel transferable erm(46) determinant responsible for emerging macrolide resistance in Rhodococcus equi. J. Antimicrob. Chemother. 2015, 70, 3184–3190. [Google Scholar] [PubMed]
- Alvarez-Narvaez, S.; Giguere, S.; Anastasi, E.; Hearn, J.; Scortti, M.; Vazquez-Boland, J.A. Clonal Confinement of a Highly Mobile Resistance Element Driven by Combination Therapy in Rhodococcus equi. mBio 2019, 10, e02260-19. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Narvaez, S.; Berghaus, L.J.; Morris, E.R.A.; Willingham-Lane, J.M.; Slovis, N.M.; Giguere, S.; Cohen, N.D. A Common Practice of Widespread Antimicrobial Use in Horse Production Promotes Multi-Drug Resistance. Sci. Rep. 2020, 10, 911. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Blum, M.; Chang, H.Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef]
- Tripathi, V.N.; Harding, W.C.; Willingham-Lane, J.M.; Hondalus, M.K. Conjugal transfer of a virulence plasmid in the opportunistic intracellular actinomycete Rhodococcus equi. J. Bacteriol. 2012, 194, 6790–6801. [Google Scholar] [CrossRef]
- Alvarez-Narvaez, S.; Giguere, S.; Berghaus, L.J.; Dailey, C.; Vazquez-Boland, J.A. Horizontal Spread of Rhodococcus equi Macrolide Resistance Plasmid pRErm46 across Environmental Actinobacteria. Appl. Environ. Microbiol. 2020, 86, e00108-20. [Google Scholar] [CrossRef]
- Berns, A.E.; Philipp, H.; Narres, H.D.; Burauel, P.; Vereecken, H.; Tappe, W. Effect of gamma-sterilization and autoclaving on soil organic matter structure as studied by solid state NMR, UV and fluorescence spectroscopy. Eur. J. Soil. Sci. 2008, 59, 540–550. [Google Scholar] [CrossRef]
- Alvarez-Narvaez, S. Conjugal Transfer of Host-Adaptive Determinants in the Pathogenic Actinobacterium Rhodococcus equi. Ph.D. Thesis, Edinburgh Research Archive, University of Edinburgh, Edinburgh, Scotland, 2017. [Google Scholar]
- Carver, T.J.; Rutherford, K.M.; Berriman, M.; Rajandream, M.-A.; Barrell, B.G.; Parkhill, J. ACT: The Artemis comparison tool. Bioinformatics 2005, 21, 3422–3423. [Google Scholar] [CrossRef]
- Askarian, F.; van Sorge, N.M.; Sangvik, M.; Beasley, F.C.; Henriksen, J.R.; Sollid, J.U.; van Strijp, J.A.; Nizet, V.; Johannessen, M. A Staphylococcus aureus TIR domain protein virulence factor blocks TLR2-mediated NF-kappaB signaling. J. Innate Immun. 2014, 6, 485–498. [Google Scholar] [CrossRef]
- Cirl, C.; Wieser, A.; Yadav, M.; Duerr, S.; Schubert, S.; Fischer, H.; Stappert, D.; Wantia, N.; Rodriguez, N.; Wagner, H.; et al. Subversion of Toll-like receptor signaling by a unique family of bacterial Toll/interleukin-1 receptor domain-containing proteins. Nat. Med. 2008, 14, 399–406. [Google Scholar] [CrossRef]
- Essuman, K.; Summers, D.W.; Sasaki, Y.; Mao, X.; Yim, A.K.Y.; DiAntonio, A.; Milbrandt, J. TIR Domain Proteins Are an Ancient Family of NAD(+)-Consuming Enzymes. Curr. Biol. 2018, 28, 421–430 e424. [Google Scholar] [CrossRef]
- Patot, S.; Imbert, P.R.; Baude, J.; Martins Simoes, P.; Campergue, J.B.; Louche, A.; Nijland, R.; Bes, M.; Tristan, A.; Laurent, F.; et al. The TIR Homologue Lies near Resistance Genes in Staphylococcus aureus, Coupling Modulation of Virulence and Antimicrobial Susceptibility. PLoS Pathog. 2017, 13, e1006092. [Google Scholar]
- Klemm, E.J.; Wong, V.K.; Dougan, G. Emergence of dominant multidrug-resistant bacterial clades: Lessons from history and whole-genome sequencing. Proc. Natl. Acad. Sci. USA 2018, 115, 12872–12877. [Google Scholar] [CrossRef]
- Bobay, L.M.; Ochman, H. The Evolution of Bacterial Genome Architecture. Front. Genet. 2017, 8, 72. [Google Scholar] [CrossRef]
- Takai, S.; Sugawara, T.; Watanabe, Y.; Sasaki, Y.; Tsubaki, S.; Sekizaki, T. Effect of growth temperature on maintenance of virulent Rhodococcus equi. Vet. Microbiol. 1994, 39, 187–192. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [PubMed]
- Coronas-Serna, J.M.; Louche, A.; Rodriguez-Escudero, M.; Roussin, M.; Imbert, P.R.C.; Rodriguez-Escudero, I.; Terradot, L.; Molina, M.; Gorvel, J.P.; Cid, V.J.; et al. The TIR-domain containing effectors BtpA and BtpB from Brucella abortus impact NAD metabolism. PLoS Pathog. 2020, 16, e1007979. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.; Zhang, J.; Fischer, H.; Huang, W.; Lutay, N.; Cirl, C.; Lum, J.; Miethke, T.; Svanborg, C. Inhibition of TIR domain signaling by TcpC: MyD88-dependent and independent effects on Escherichia coli virulence. PLoS Pathog. 2010, 6, e1001120. [Google Scholar] [CrossRef] [PubMed]
- Wagner, T.M.; Janice, J.; Paganelli, F.L.; Willems, R.J.; Askarian, F.; Pedersen, T.; Top, J.; de Haas, C.; van Strijp, J.A.; Johannessen, M.; et al. Enterococcus faecium TIR-Domain Genes Are Part of a Gene Cluster Which Promotes Bacterial Survival in Blood. Int. J. Microbiol. 2018, 2018, 1435820. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.; Horohov, D.W.; Meijer, W.G.; Muscatello, G. Current understanding of the equine immune response to Rhodococcus equi. An immunological review of R. equi pneumonia. Vet. Immunol. Immunopathol. 2010, 135, 1–11. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Comprehensive comparative-genomic analysis of type 2 toxin-antitoxin systems and related mobile stress response systems in prokaryotes. Biol. Direct 2009, 4, 19. [Google Scholar] [CrossRef]
- Arcus, V.L.; McKenzie, J.L.; Robson, J.; Cook, G.M. The PIN-domain ribonucleases and the prokaryotic VapBC toxin-antitoxin array. Protein Eng. Des. Sel. 2011, 24, 33–40. [Google Scholar] [CrossRef]
- Fraikin, N.; Goormaghtigh, F.; Van Melderen, L. Type II Toxin-Antitoxin Systems: Evolution and Revolutions. J. Bacteriol. 2020, 202, e00763-19. [Google Scholar] [CrossRef]
- Page, R.; Peti, W. Toxin-antitoxin systems in bacterial growth arrest and persistence. Nat. Chem. Biol. 2016, 12, 208–214. [Google Scholar] [CrossRef]
- Ghinet, M.G.; Bordeleau, E.; Beaudin, J.; Brzezinski, R.; Roy, S.; Burrus, V. Uncovering the prevalence and diversity of integrating conjugative elements in actinobacteria. PLoS ONE 2011, 6, e27846. [Google Scholar] [CrossRef]
- Guedon, G.; Libante, V.; Coluzzi, C.; Payot, S.; Leblond-Bourget, N. The Obscure World of Integrative and Mobilizable Elements, Highly Widespread Elements that Pirate Bacterial Conjugative Systems. Genes 2017, 8, 337. [Google Scholar] [CrossRef] [PubMed]
- Puymege, A.; Bertin, S.; Guedon, G.; Payot, S. Analysis of Streptococcus agalactiae pan-genome for prevalence, diversity and functionality of integrative and conjugative or mobilizable elements integrated in the tRNA(Lys CTT) gene. Mol. Genet. Genom. 2015, 290, 1727–1740. [Google Scholar] [CrossRef] [PubMed]
- Willingham-Lane, J.M.; Berghaus, L.J.; Berghaus, R.D.; Hart, K.A.; Giguere, S. Effect of Macrolide and Rifampin Resistance on the Fitness of Rhodococcus equi. Appl. Environ. Microbiol. 2019, 85, e02665-18. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Name | Sequence 5′–3′ | Amplicon Size (bp) | Purpose | Source |
|---|---|---|---|---|
| IME2287_repAB_F | GGAGCACTACTACTGGACG | 1746 | IME2287 backbone marker | [43] |
| IME2287_repAB_R | GTTGACTGTGAACTCGGTGT | |||
| IME2287_sigma70_F | CTTGCGAGTAGGACATGAAG | 1752 | IME2287 backbone marker | [43] |
| IME2287_sigma70_R | GACCTTCGTCAGGGAGTAAG | |||
| IME2287_tnpR- helix_F | TCTACGTCGACAAGAAGTCC | 1745 | IME2287 backbone marker | [43] |
| IME2287_tnpR- helix_R | GTATGTGAACCGACCTTGTG | |||
| ChoE_F | AGTTGTCGATTCCCATCGTC | 672 | choE gene, chromosomal marker | [5] |
| ChoE_R | AAGCGCAACTACTTCGAGGAG | |||
| TraA-F1 | AGAGTTCATGCGTGACAACG | 959 | traA gene, pVAPA backbone marker | [5] |
| TraA-R1 | GTCCACAGGTCACCGTTCTT | |||
| erm(46)F | TATGGAGTCGATCTGCAACG | 1098 | macrolide resistance gene erm(46) | [34] |
| erm(46)R | GAGATCGGACGAGTCTGACA | |||
| pRErm46_traG_F | ACCGTCGTAGCAGTAGCC | 1533 | traG gene, pRErm46 backbone marker | [35] |
| pRErm46_traG_R | CCTCAGCGAGTGTCTTCTC | |||
| ApraF | GGCCACTTGGACTGATCGAG | 937 | apramycin cassette aac(3)IV inserted in chromosome | [40] |
| ApraR | GCATGACCGACTGGACCTTC |
| IME2287 Locus Tag | Location (nt Position) | Size (bp) | Product (BlastX) | Coverage (AA Level) | Identity (AA Level) | Species | |
|---|---|---|---|---|---|---|---|
| direct_repeat | 1 | 4 | 3 | - | - | - | - |
| inverted_repeat | 5 | 34 | 29 | - | - | - | - |
| IME2287_0010 | 46 | 303 | 257 | ParG | >90% | >80% | Conserved in Actinobacteria |
| IME2287_0020 | 306 | 1067 | 761 | ParA | >50% | >50% | Conserved in Actinobacteria |
| IME2287_0030 | 1165 | 1488 | 323 | Conserved hypothetical protein | >90% | >80% | Conserved in Rhodococcus spp. |
| IME2287_0040 | 1478 | 1765 | 287 | RepB | >70% | >80% | Conserved in Rhodococcus spp. |
| IME2287_0050 | 1765 | 2651 | 886 | RepA | >70% | >70% | Conserved in Rhodococcus spp. |
| IME2287_0060 | 2873 | 3520 | 647 | Conserved hypothetical protein | >70% | >50% | Conserved in Actinobacteria |
| IME2287_0070 | 3531 | 3854 | 323 | Conserved hypothetical protein | >50% | >50% | Conserved in Rhodococcus spp. |
| IME2287_0080 | 3814 | 4413 | 599 | Conserved hypothetical protein | >50% | >50% | Conserved in Rhodococcus spp. |
| IME2287_0090 | 4528 | 5208 | 680 | Hypothetical protein | no match | no match | No match |
| IME2287_0100 | 5167 | 5409 | 242 | Conserved hypothetical protein | >70% | >50% | Conserved in Rhodococcus spp. |
| IME2287_0110 | 5503 | 6471 | 968 | TIR domain-containing protein | >90% | >40% | Conserved bacterial protein |
| IME2287_0120 | 6543 | 6731 | 188 | Transmembrane protein | no match | no match | No match |
| IME2287_0130 | 7010 | 7903 | 893 | Sigma70 | >70% | >35% | Conserved in Actinobacteria |
| IME2287_0140 | 7900 | 8241 | 341 | TPR-like domian-containing protein | no match | no match | No match |
| IME2287_0150 | 8392 | 9024 | 632 | PIN domain-containing protein | >85% | >40% | Conserved in Actinobacteria |
| IME2287_0160 | 9073 | 9543 | 470 | MerR-like DNA binding protein | >60% | 60% | Conserved in Actinobacteria |
| IME2287_0170 | 10,014 | 11,267 | 1253 | Serine recombinase | >40% | >70% | Conserved in Rhodococcus spp. |
| IME2287_0180 | 11,264 | 12,223 | 959 | Conserved hypothetical protein | >85% | >25% | Conserved in Actinobacteria |
| IME2287_0190 | 12,470 | 12,886 | 416 | Conserved hypothetical protein | >80% | >50% | Conserved in Actinobacteria |
| IME2287_0200 | 13,038 | 13,931 | 893 | Conserved hypothetical protein | >90% | >40% | Conserved in Actinobacteria |
| inverted_repeat | 14,284 | 14,313 | 29 | - | - | - | - |
| direct_repeat | 14,314 | 14,317 | 3 | - | - | - | - |
| Source | Genetic Background | pRErm46/tnRErm46 and IME2287 | pRErm46/ tnRErm46 Only | No pRErm46/tnRErm46 or IME2287 | Total |
|---|---|---|---|---|---|
| Clinical | Clone 2287 | 40 | 0 | 0 | 40 |
| Clone G2016 | 1 | 1 | 0 | 2 | |
| Clone G2017 | 0 | 0 | 0 | 0 | |
| Singletons | 2 | 6 | 0 | 8 | |
| Susceptible | 0 | 0 | 22 | 22 | |
| Environmental | Clone 2287 | 45 | 0 | 0 | 45 |
| Clone G2016 | 0 | 1 | 0 | 1 | |
| Clone G2017 | 3 | 4 | 32 | 39 | |
| Singletons | 7 | 2 | 3 | 12 | |
| Susceptible | 0 | 0 | 38 | 38 | |
| Total | 98 | 14 | 95 | 207 |
| CFU with Macrolide-Resistant Phenotype | ||||
|---|---|---|---|---|
| DHS | Soil | |||
| Passage | RT | 37 °C | RT | 37 °C |
| 0 | 100% | 100% | 100% | 100% |
| 15 | 100% | 99% | 100% | 100% |
| 30 | 99% | 97% | 100% | 100% |
| 45 | 100% | 64% | 100% | 95% |
| Media | Temp. | Passage | Loss of pRErm46 Only | Loss of pRErm46 and IME2287 |
|---|---|---|---|---|
| Soil | 37 °C | 45 | 5 | 0 |
| DHS | 37 °C | 30 | 1 | 0 |
| RT | 15 | 1 | 0 | |
| 30 | 3 | 0 | ||
| 45 | 13 | 23 | ||
| Total | 23 | 23 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez Narvaez, S.; Sanchez, S. Exploring the Accessory Genome of Multidrug-Resistant Rhodococcus equi Clone 2287. Antibiotics 2023, 12, 1631. https://doi.org/10.3390/antibiotics12111631
Alvarez Narvaez S, Sanchez S. Exploring the Accessory Genome of Multidrug-Resistant Rhodococcus equi Clone 2287. Antibiotics. 2023; 12(11):1631. https://doi.org/10.3390/antibiotics12111631
Chicago/Turabian StyleAlvarez Narvaez, Sonsiray, and Susan Sanchez. 2023. "Exploring the Accessory Genome of Multidrug-Resistant Rhodococcus equi Clone 2287" Antibiotics 12, no. 11: 1631. https://doi.org/10.3390/antibiotics12111631